
Comparative Physiological and Transcriptomic Analyses Reveal Mechanisms of Exogenous Spermidine-Induced Tolerance to Low-Iron Stress in Solanum lycopersicum L

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions and Experimental Treatments
2.2. RNA-Seq Analysis and Quantitative Real-Time PCR Analysis
2.3. Determination of Biomass and Root Morphology, Root Vigor and Root Fe3+ Reductase Activity
2.4. Determination of Photosynthetic Pigment Content and Photosynthetic Index
2.5. Determination of Antioxidant Properties and Osmoregulatory Substances
2.6. Determination of Sucrose Content and Metabolism-Related Enzyme Activities
2.7. Determination of Polyamine Content
2.8. Statistical Analysis
3. Results
3.1. Overview of Sequencing Data-Quality Control
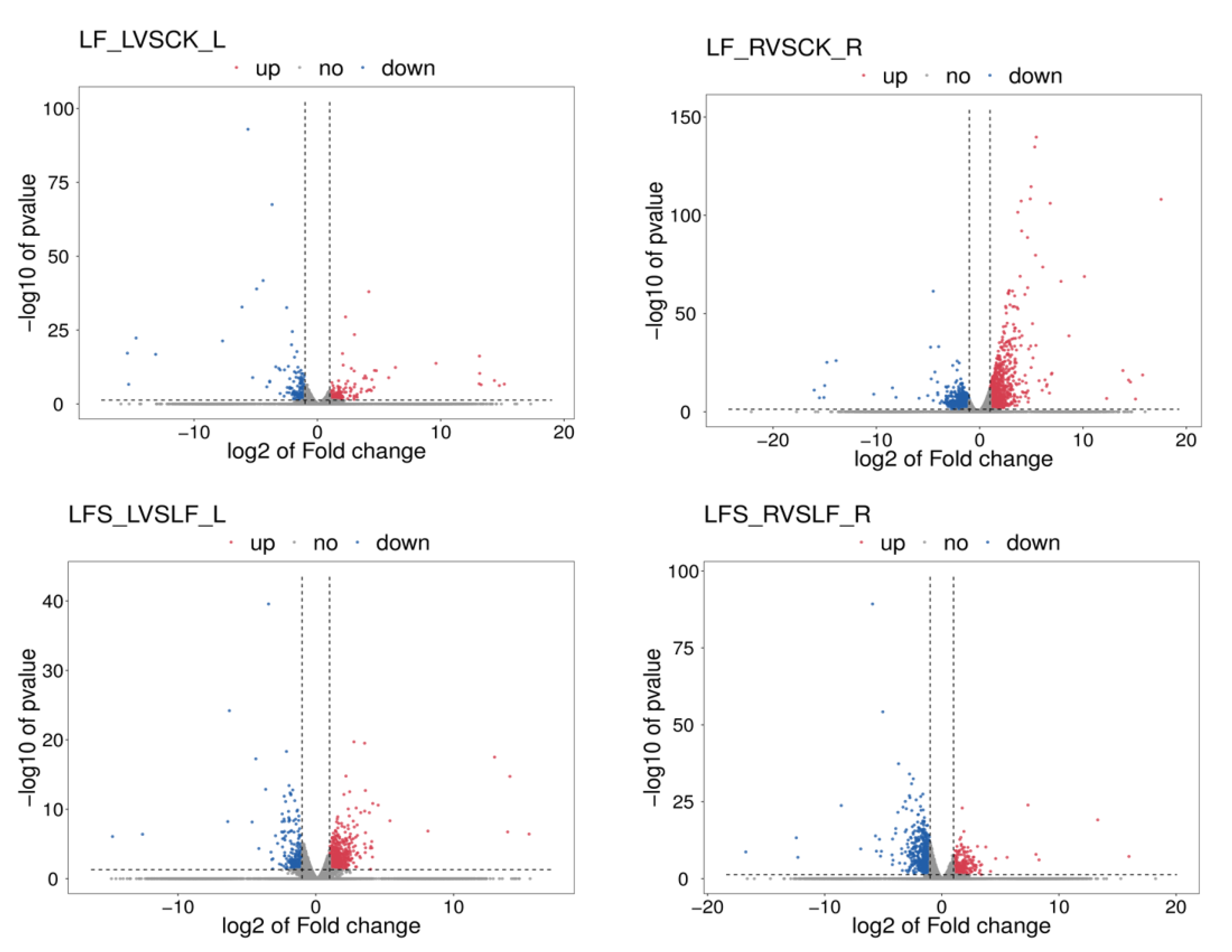
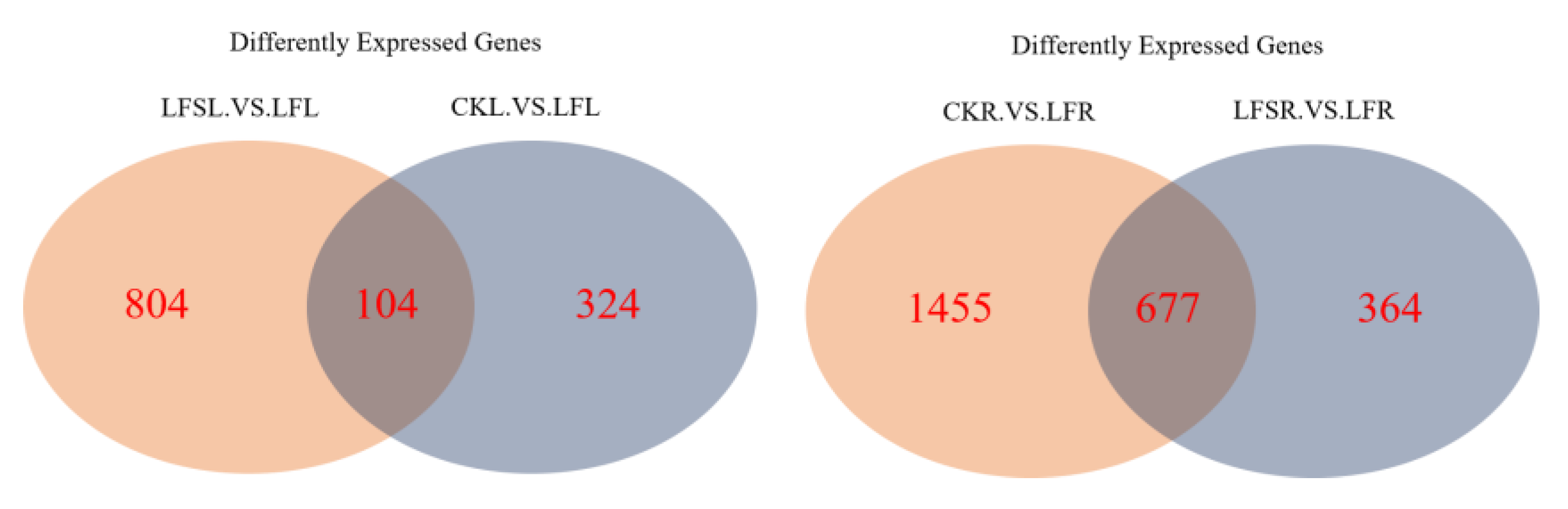
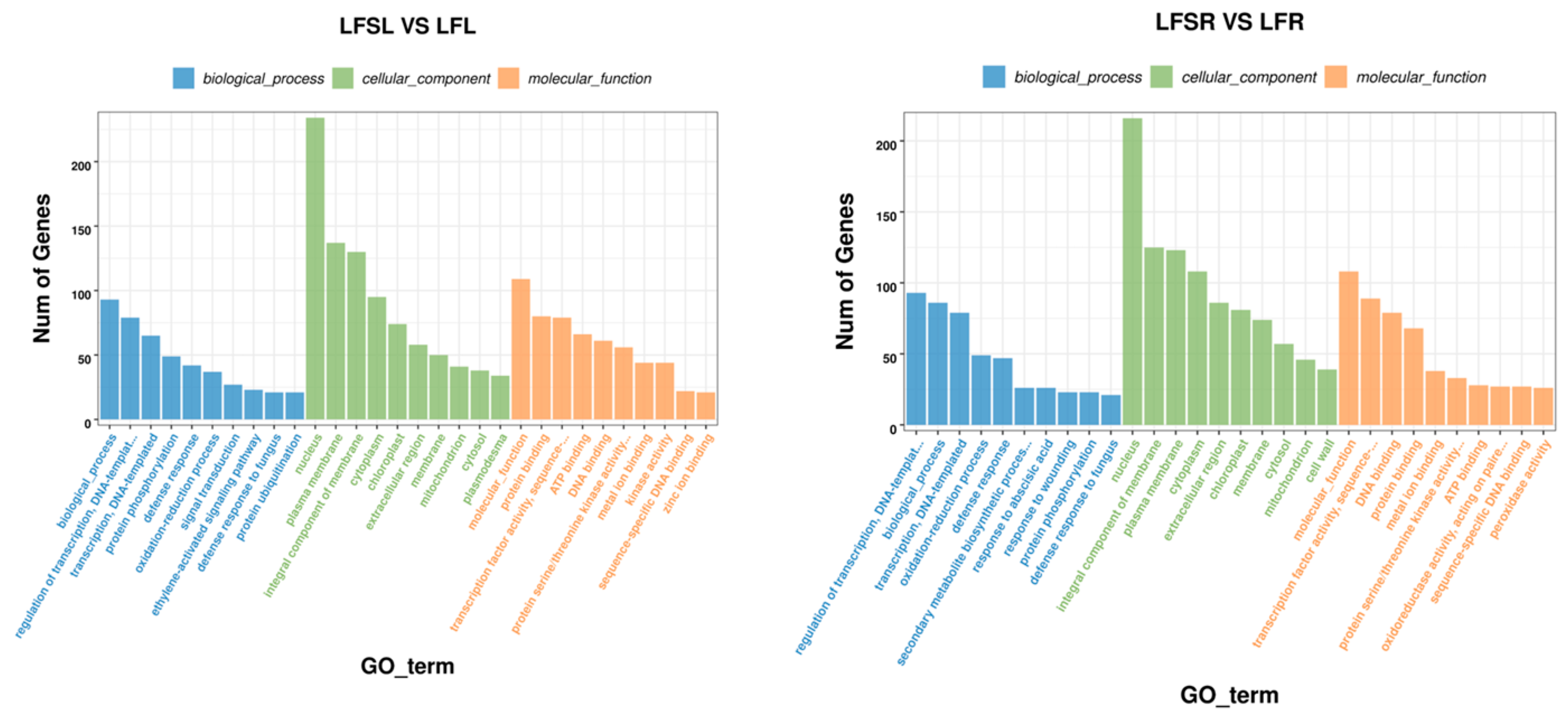
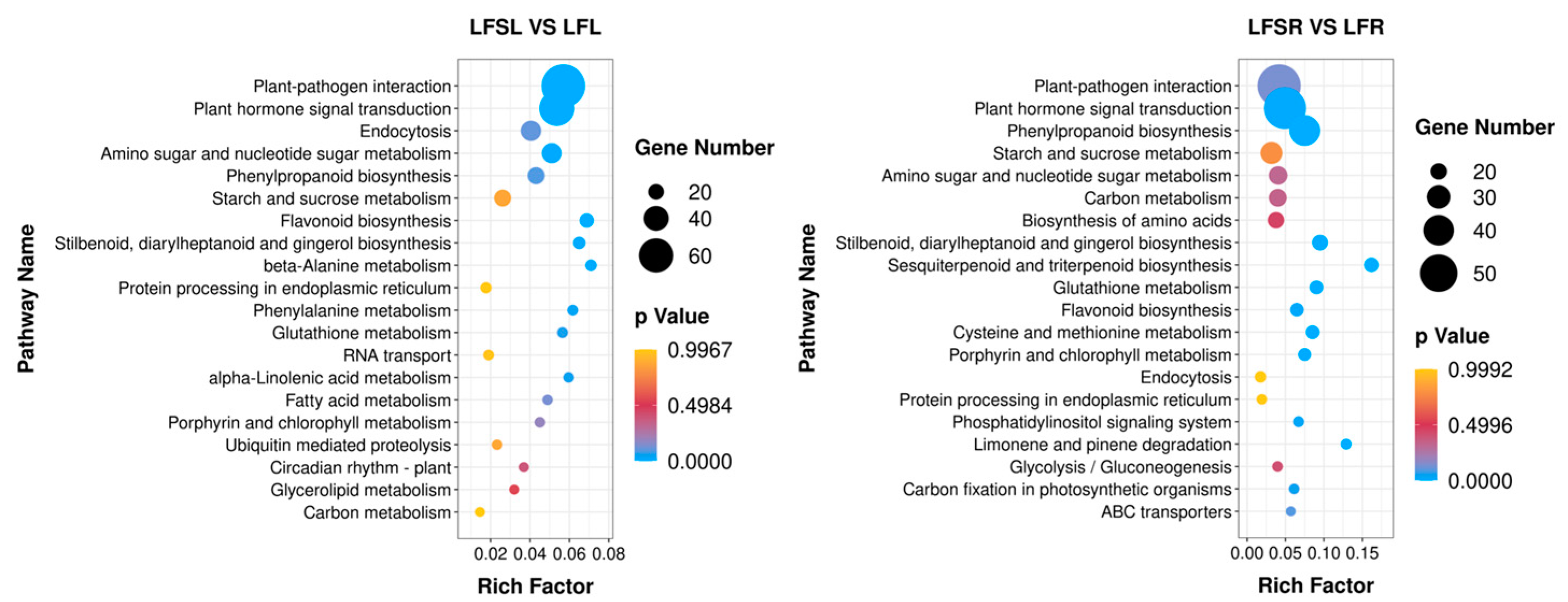
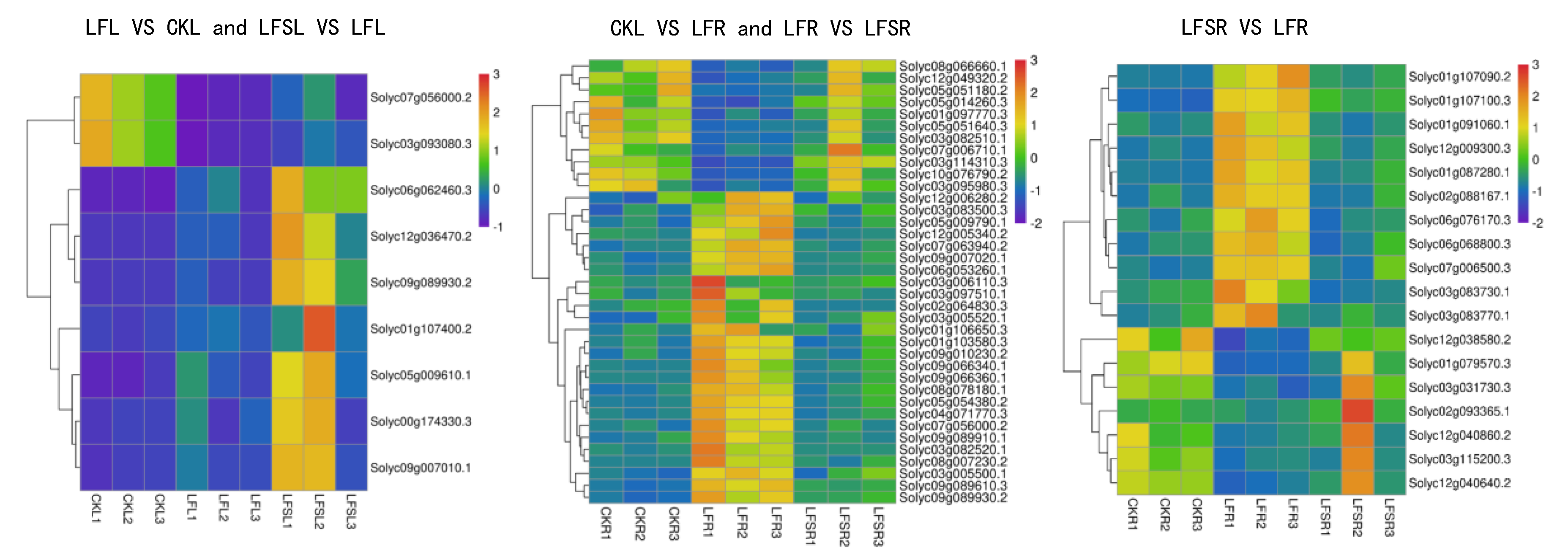
3.2. Analysis of Differentially Expressed Genes
3.3. Exogenous Spd Improved the Growth and Photosynthetic Efficiency of Tomato Plants under Low-Iron Stress
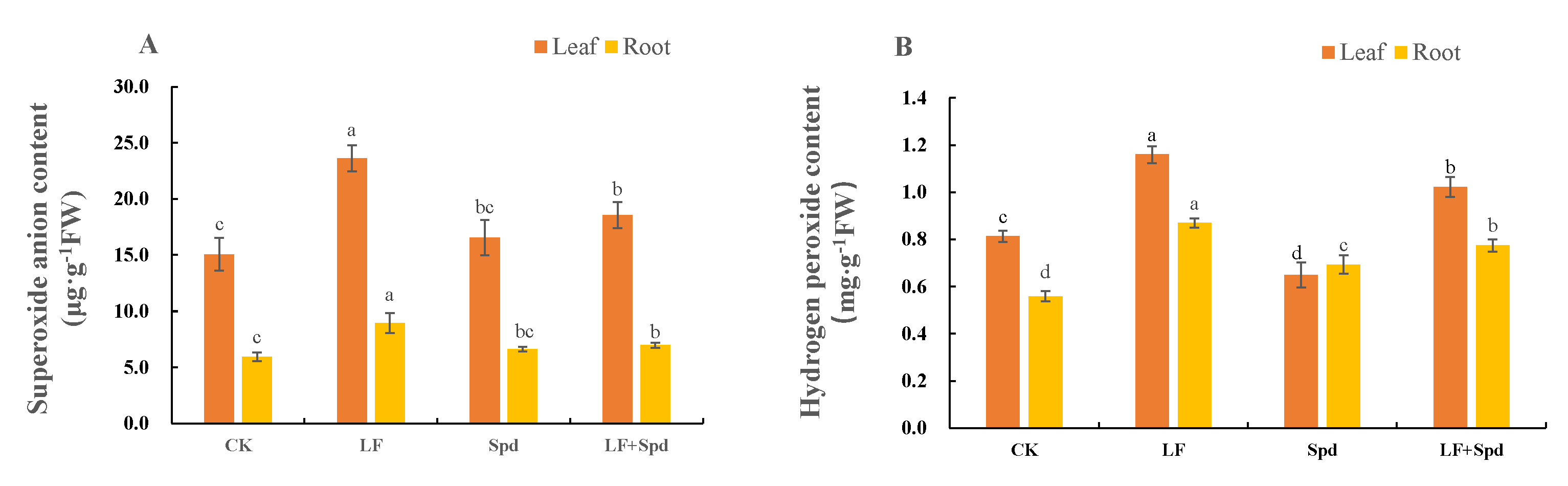
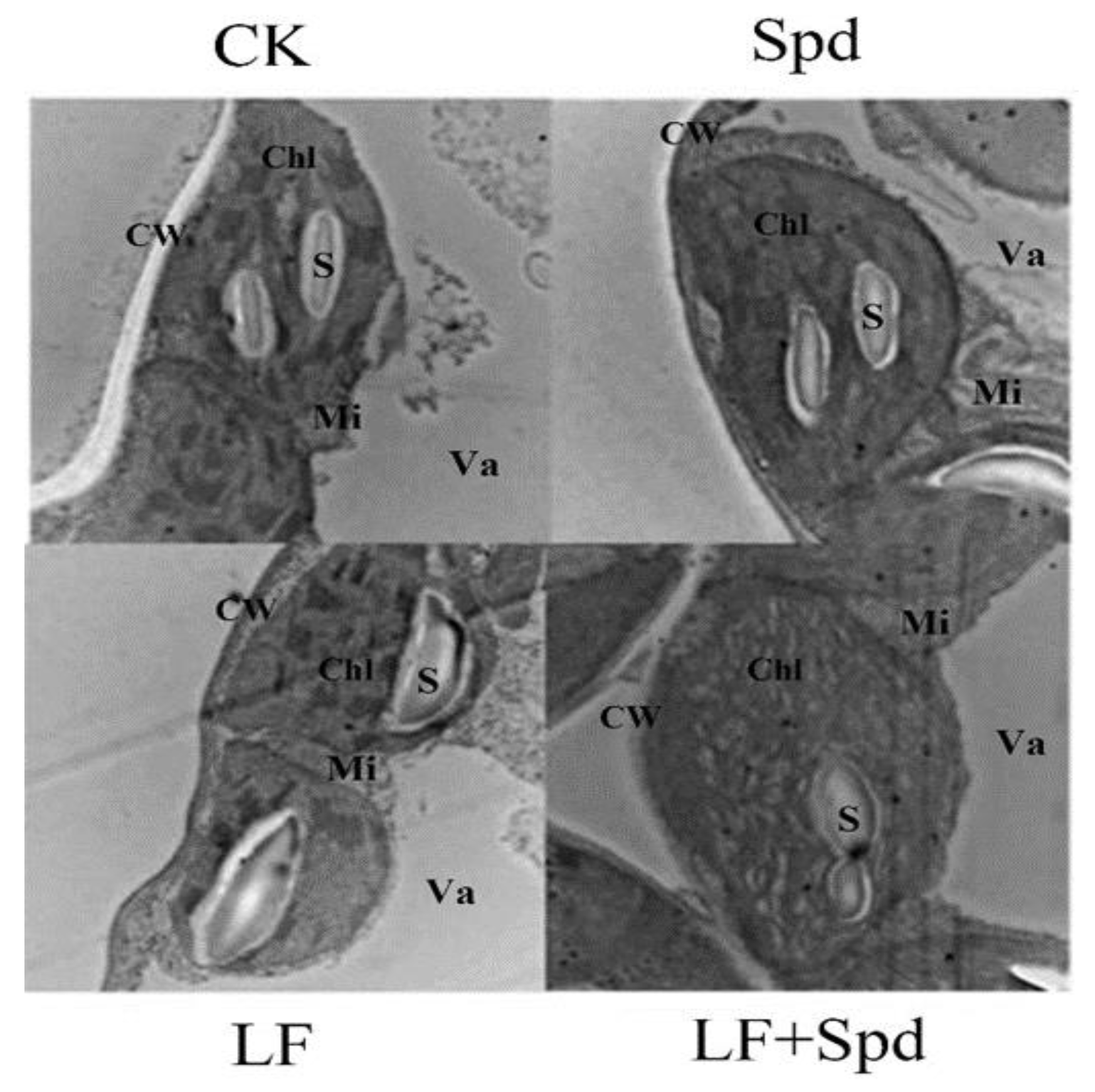
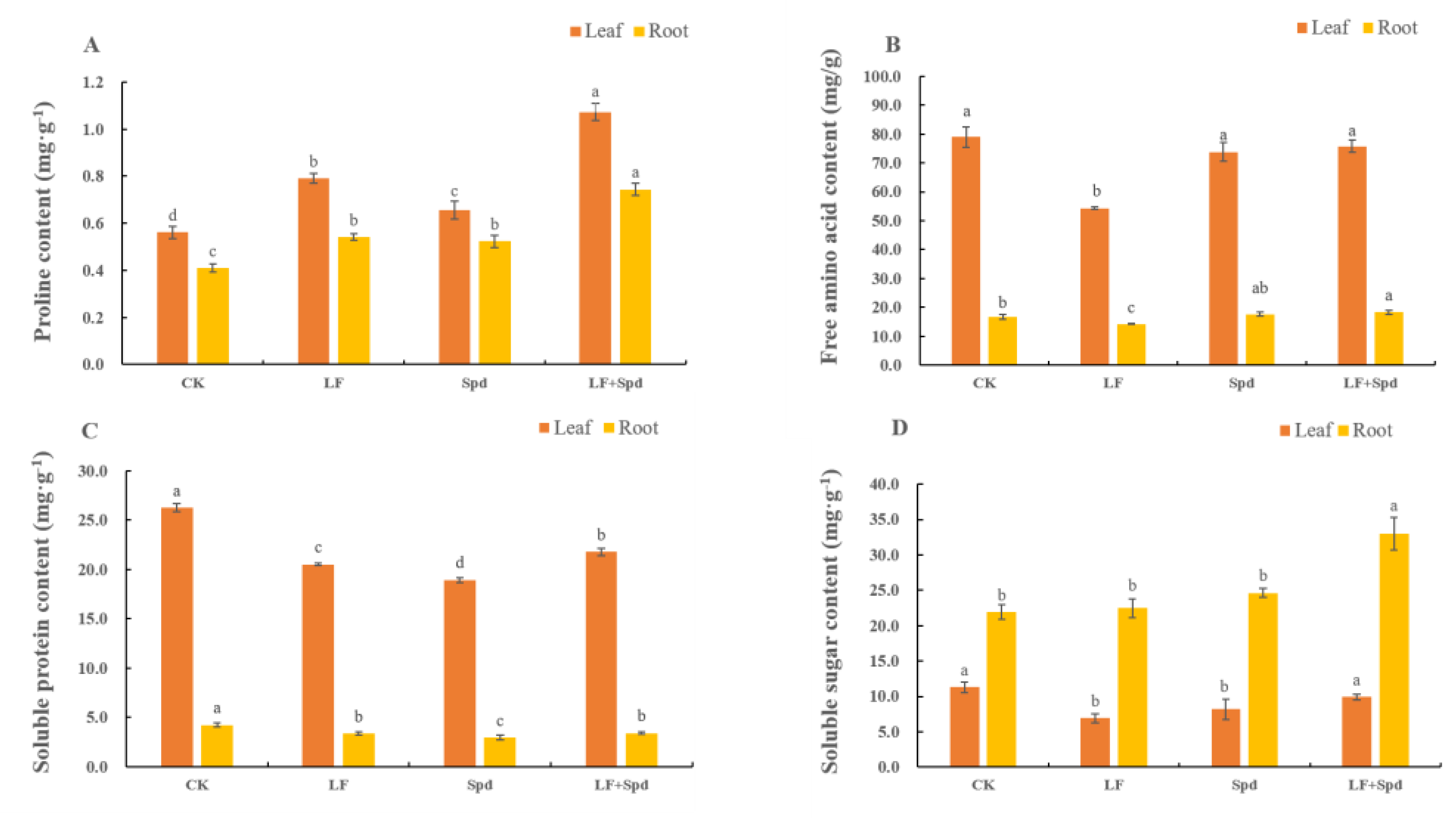
3.4. Effect of Exogenous Spd on ROS Accumulation, Antioxidant System and Osmoregulatory Substances in Tomatoes under Low-Iron Stress
3.5. Effect of Exogenous Spd on the Organic Acid Content in Roots and the Polyamine Content in Leaves under Low-Iron Stress in Tomato Plants
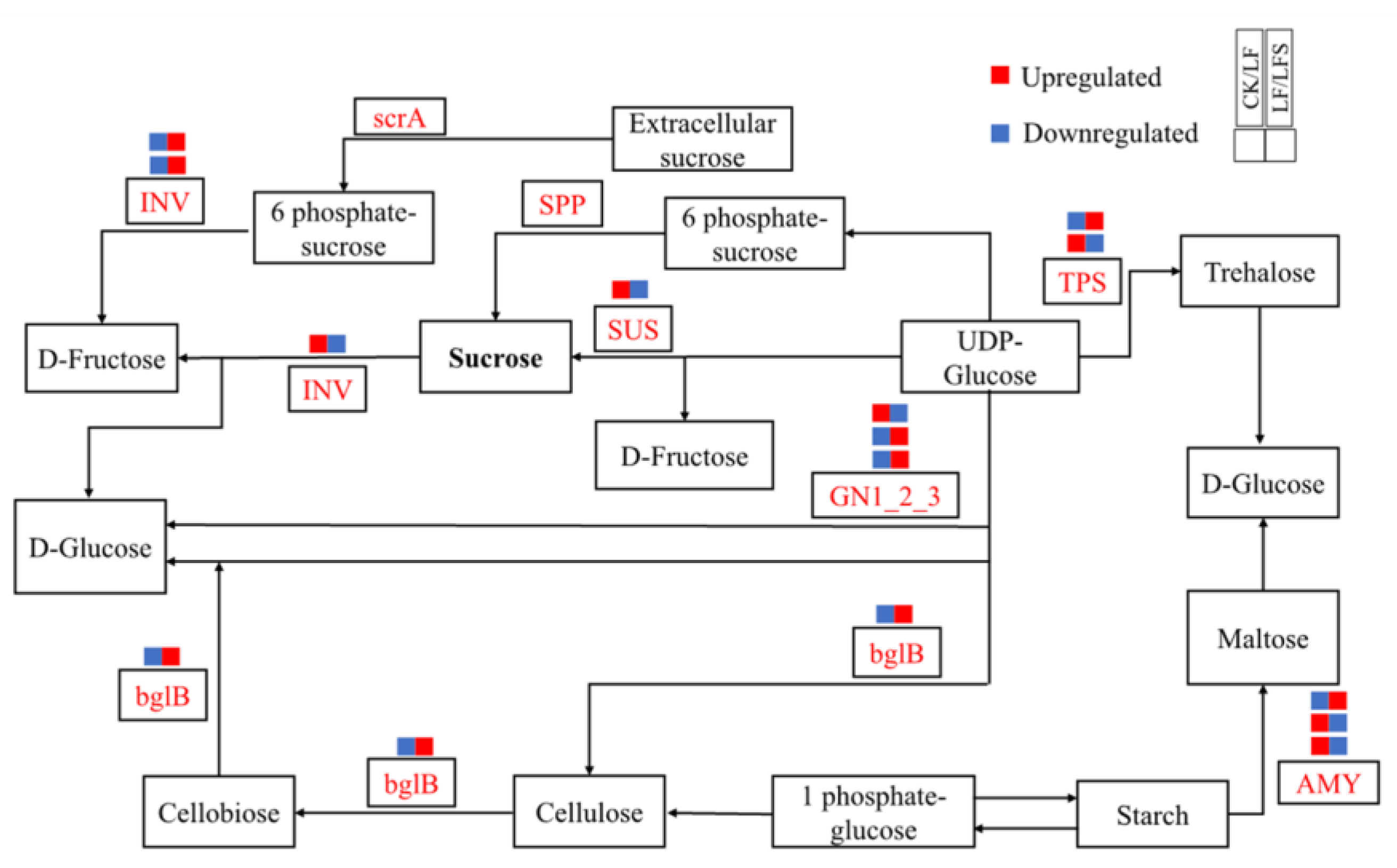
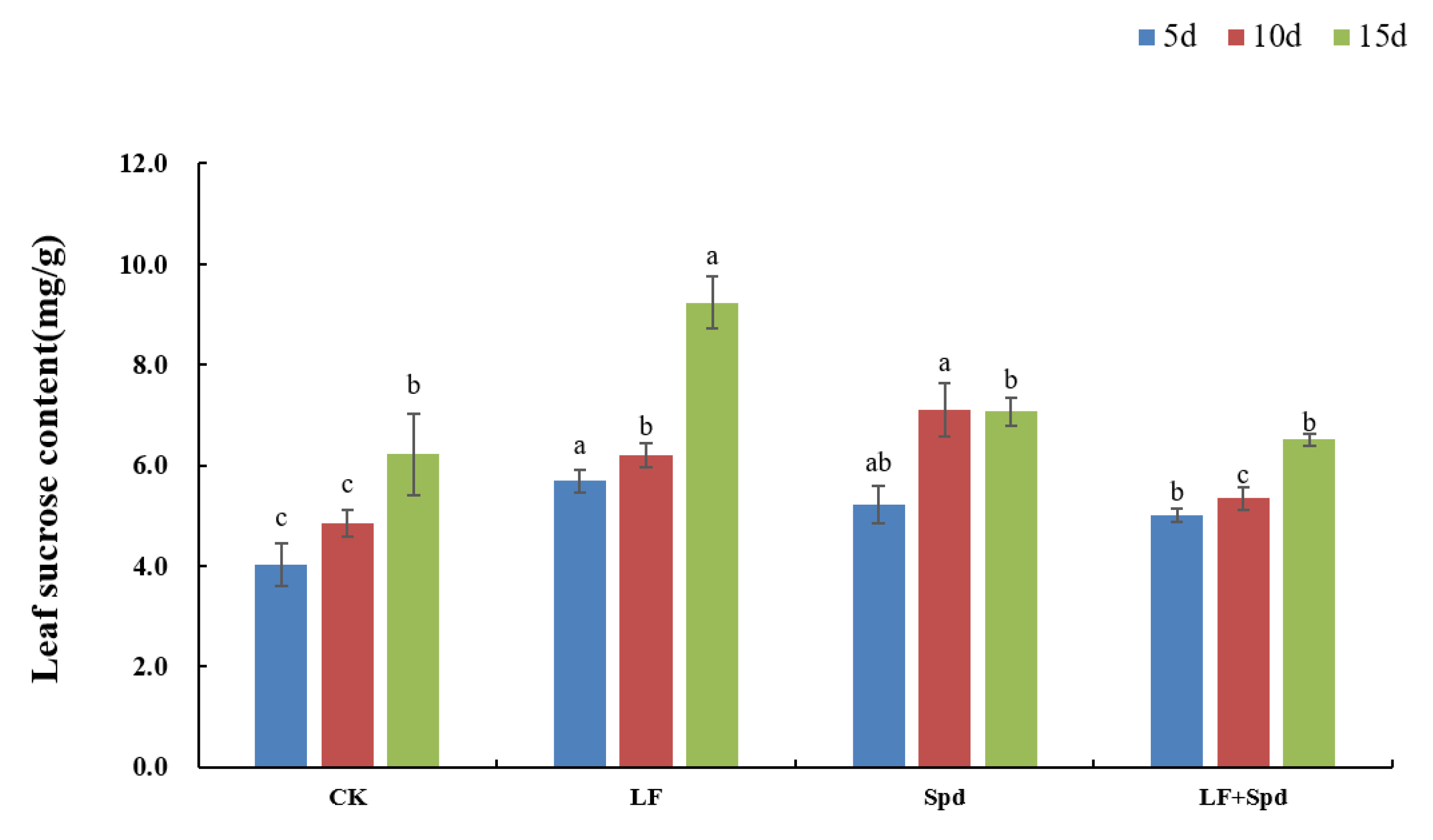
3.6. Effect of Exogenous Spd on Sugar Metabolism in Tomato Leaves under Low-Iron Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaya, C.; Akram, N.; Ashraf, M. Influence of exogenously applied nitric oxide on strawberry (Fragaria × ananassa) plants grown under iron deficiency and/or saline stress. Physiol. Plant. 2018, 165, 247–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Shukla, V.; Meena, V.; Kumar, A.; Singh, J.; Kandoth, K.; Mantri, S.; Raouched, H.; Pandey, A.K. Underpinning wheat physiological and molecular responses to co-occurring iron and phosphate deficiency stress. HAL 2020, hal-02749803. [Google Scholar]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef]
- Molassiotis, A.N.; Diamantidis, G.C.; Therios, I.N.; Tsirakoglou, V.; Dimassi, K.N. Oxidative stress, antioxidant activity and Fe (III)-chelate reductase activity of five Prunus rootstocks explants in response to Fe deficiency. Plant Growth Regul. 2005, 46, 69–78. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of Nutrients: Micronutrients-ScienceDirect. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 191–248. [Google Scholar]
- Kumar, S.; Li, G.; Yang, J.; Huang, X.F.; Ji, Q.; Zhou, K.; Khan, S.; Ke, W.; Houet, H.W. Investigation of an Antioxidative System for Salinity Tolerance in Oenanthe javanica. Antioxidants 2020, 9, 940. [Google Scholar] [CrossRef] [PubMed]
- Reimer, J.; Shaaban, B.; Drummen, N.; Sanjeev Ambady, S.; Genzel, F.; Poschet, G.; Wiese-Klinkenberg, A.; Usadel, B.; Wormit, A. Capsicum Leaves under Stress: Using Multi-Omics Analysis to Detect Abiotic Stress Network of Secondary Metabolism in Two Species. Antioxidants 2022, 11, 671. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Dong, Y.; Tian, X.; Wang, W.; He, Z. Mechanisms of exogenous nitric oxide and 24-Epibrassinolide alleviating chlorosis of peanut plants under iron deficiency. Pedosphere 2018, 28, 926–942. [Google Scholar] [CrossRef]
- Chen, W.W.; Yang, L.J.; Cheng, J.; Mo, J.; Ye, T.; Zheng, S. Nitric oxide acts downstream of auxin to trigger root ferric-chelate reductase activity in response to iron deficiency in arabidopsis. Plant Physiol. 2010, 154, 810–819. [Google Scholar] [CrossRef] [Green Version]
- Römheld, V.; Marschner, H. Iron defificiency stress induced morphological and physiological changes in root tips of sunflflower. Physiol. Plant. 1981, 53, 354–360. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef] [Green Version]
- Vert, G.; Grotz, N.; Dédaldéchamp, F.; Gaymard, F.; Guerinot, M.L.; Briat, J.F.; Curie, C. Correction: IRT1, an arabidopsis transporter essential for iron uptake from the soil and for plant growth. Plant Cell 2021, 33, 439–440. [Google Scholar] [CrossRef]
- Lingam, S.; Mohrbacher, J.; Brumbarova, T.; Potuschak, T.; Fink-Straube, C.; Blondet, D.; Genschik, P.; Bauer, P. Interaction between the bHLH transcription factor fit and ethylene insensitive3/ethylene insensitive3-like1 reveals molecular linkage between the regulation of iron acquisition and ethylene signaling in arabidopsis. Plant Cell 2011, 23, 1815–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktar, F.; Islam, M.S.; Milon, M.A.-A.; Islam, N.; Islam, M.A. Polyamines: An essentially regulatory modulator of plants to abiotic stress tolerance: A review. Asian J. Appl. Sci. 2021, 9, 195–204. [Google Scholar] [CrossRef]
- Duan, J.; Li, J.; Guo, S.; Kang, Y. Exogenous spermidine affects polyamine metabolism in salinity-stressed Cucumis sativus roots and enhances short-term salinity tolerance. Plant Physiol. 2008, 165, 1620–1635. [Google Scholar] [CrossRef] [PubMed]
- ElSayed, A.I.; Rafudeen, M.S.; El-hamahmy, M.A.M.; Odero, D.C.; Hossain, M.S. Enhancing antioxidant systems by exogenous spermine and spermidine in wheat (Triticum aestivum) seedlings exposed to salt stress. Funct. Plant Biol. 2018, 45, 745–759. [Google Scholar] [CrossRef]
- Amri, E.; Shahsavar, A.R. Response of lime seedlings (Citrus aurantifolia L.) to exogenous spermidine treatments under drought stress. Aust. J. Basic Appl. Sci. 2010, 4, 4483–4489. [Google Scholar]
- Sun, X.; Xie, L.; Han, L. Effects of exogenous spermidine and spermine on antioxidant metabolism associated with cold-induced leaf senescence in Zoysiagrass (Zoysia japonica Steud.). Hortic. Environ. Biotechnol. 2019, 60, 295–302. [Google Scholar] [CrossRef]
- Xu, X.; Shi, G.; Ding, C.; Xu, Y.; Zhao, J.; Yang, H.; Pan, Q. Regulation of exogenous spermidine on the reactive oxygen species level and polyamine metabolism in Alternanthera philoxeroides (Mart.) Griseb under copper stress. Plant Growth Regul. 2011, 63, 251–258. [Google Scholar] [CrossRef]
- Sang, Q.; Shan, X.; An, Y.; Shu, S.; Sun, J.; Guo, S. Proteomic analysis reveals the positive effect of exogenous spermidine in tomato seedlings’ response to high-temperature stress. Front. Plant Sci. 2017, 8, 120. [Google Scholar] [CrossRef] [Green Version]
- Qian, R.; Ma, X.; Zhang, X.; Hu, Q.; Liu, H.; Zheng, J. Effect of exogenous spermidine on osmotic adjustment, antioxidant enzymes activity, and gene expression of gladiolus gandavensis seedlings under salt stress. Plant Growth Regul. 2020, 40, 1353–1367. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, Y.; Zhao, X.; Jin, X.; Hou, L.; Shi, Y. Silicon compensates phosphorus deficit-Induced growth inhibition by improving photosynthetic capacity, antioxidant potential, and nutrient homeostasis in tomato. Agronomy 2019, 9, 733. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Qian, Z.; Luo, M.; Fan, S.; Zhang, X.; Zhang, L. Identification of salt stress responding genes using transcriptome analysis in green alga Chlamydomonas reinhardtii. Int. J. Mol. Sci. 2018, 19, 3359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenneth, J.L.; Thomas, D.S. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-∆∆Ct Method. Methods 2001, 25, 402–408. [Google Scholar]
- Zhang, Y.; Li, S.; Liang, Y.; Zhao, H.; Hou, L.; Shi, Y.; Ahammed, G.J. Effects of exogenous spermidine and elevated CO2 on physiological and biochemical changes in tomato plants under iso-osmotic salt stress. Plant Growth Regul. 2018, 37, 1222–1234. [Google Scholar] [CrossRef] [Green Version]
- Mellor, R.S.; Pietro, A.S. Experimental plant physiology. Bioence 1974, 24, 418. [Google Scholar]
- Ekmekcioglu, C.; Strauss-Blasche, G.; Marktl, W. The plasma membrane Fe3+-reductase activity of CaCO2 cells is modulated during differentiation. Biochem. Mol. Biol. Int. 1998, 46, 951–961. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leafextracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, U.; Armond, P.A. Heat-induced changes of chlorophyll fluorescence in isolated chloroplasts and related heat-damage at the pigment level. Biochim. Biophys. Acta 1978, 502, 138–151. [Google Scholar] [CrossRef]
- Cai, S.; Zhang, Y.; Xu, Y.; Qi, Z.; Li, M.; Ahammed, G.; Xia, X.; Shi, K.; Zhou, Y.; Retier, R.; et al. HsfA1a upregulates melatonin biosynthesis to confer cadmium tolerance in tomato plants. Pineal. Res. 2017, 62, e12387. [Google Scholar] [CrossRef]
- Reuveni, R.; Shimoni, M.; Karchi, Z.; Kuc, J. Peroxidase Activity as a Biochemical Marker for Resistance of Muskmelon (Cucumis melo) to Pseudoperonospora cubensis. Phytopathology 1992, 82, 749–753. [Google Scholar] [CrossRef]
- Garcia, C.; Fedrigo, K.; Gabriel, A.; Botelho, R.V.; Rodrigues, J.D.; Ono, E.O. Control of mildew in vines with cinnamon extract and catalase activity in organic production. Res. Soc. Dev. 2021, 10, e214101018885. [Google Scholar] [CrossRef]
- Ukeda, H.; Kawana, D.; Maeda, S.; Sawamura, M. Spectrophotometric assay for superoxide dismutase based on the reduction of highly water-soluble tetrazolium salts by xanthine-xanthine oxidase. Biosci. Biotechnol. Biochem. 1999, 63, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Sheteiwy, M.; Shen, H.; Xu, J.; Guan, Y.; Song, W.; Hu, J. Seed polyamines metabolism induced by seed priming with spermidine and 5-aminolevulinic acid for chilling tolerance improvement in rice (Oryza sativa L.) seedlings. Environ. Exp. Bot. 2017, 137, 58–72. [Google Scholar] [CrossRef]
- Li, S.; Jiang, H.; Wang, J.; Wang, Y.; Pan, S.; Tian, H.; Duan, M.; Wang, S.; Tang, X.; Mo, Z. Responses of plant growth, physiological, gas exchange parameters of super and non-super rice to rhizosphere temperature at the tillering stage. Sci. Rep.-UK 2019, 9, 10618. [Google Scholar] [CrossRef]
- Shou, H.; Bordallo, P.; Fan, J.; Yeakley, J.M.; Bibikova, M.; Sheen, J.; Wang, K. Expression of an active tobacco mitogen-activated protein kinase kinase kinase enhances freezing tolerance in transgenic maize. Proc. Natl. Acad. Sci. USA 2004, 101, 3298–3303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Montagu, M.V.; Inzé, D.; Camp, W.V. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Gong, M. Improvement of measurement method for superoxide anion radical in plant. Acta Bot. Yunnanica 2005, 27, 211–216. [Google Scholar]
- Bates, L.; Waldren, R.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, X.; Shi, Y.; Zou, Z.; Yan, F.; Zhao, Y.; Zhang, H.; Zhao, J. Beneficial role of exogenous spermidine on nitrogen metabolism in tomato seedlings exposed to saline-alkaline stress. Am. Soc. Hortic. Sci. 2013, 138, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Güler, A.; Candemir, A. Determination of physicochemical characteristics, organic acid and sugar profiles of Turkish grape juices. Int. J. Agric. Environ. Food Sci. 2020, 4, 149–156. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Y.; Chen, S.; Liu, J.; Fan, K.; Li, Z.; Liu, Z.; Lin, W. Gibberellins play dual roles in response to phosphate starvation of tomato seedlings, negatively in shoots but positively in roots. Plant Physiol. 2019, 234–235, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.R.; Tyms, S. Rapid analysis of polyamines in cell culture by high performance liquid chromatography. Med. Lab. Sci. 1986, 43, 258–261. [Google Scholar] [PubMed]
- Guo, L.; Born, M.L.; Niu, W.; Liu, F. Biochar amendment improves shoot biomass of tomato seedlings and sustains water relations and leaf gas exchange rates under different irrigation and nitrogen regimes. Agric. Water Manag. 2021, 245, 106580. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.; Naz, S.; Altaf, M.M.; Khan, L.; Tiwari, R.K.; Lal, M.K.; Shahid, M.A.; Kumar, R.; et al. Melatonin Improves Drought Stress Tolerance of Tomato by Modulating Plant Growth, Root Architecture, Photosynthesis, and Antioxidant Defense System. Antioxidants 2022, 11, 309. [Google Scholar] [CrossRef]
- Carrasco-Gil, S.; Allende-Montalbán, R.L.; HernándezApaolaza, L.; Lucena, J.J. Application of seaweed organic components increases tolerance to Fe deficiency in tomato plants. Agronomy 2021, 11, 507. [Google Scholar] [CrossRef]
- Lu, Y.; Ye, H.; Geng, S.; Huang, Z.; Liu, Z.; Long, X.; Liu, Z. Effects of NaCl stress on growth, leaf photosynthetic parameters and ion distribution of Helianthus tuberosus seedling. Plant Resour. Environ. 2010, 19, 86–91. [Google Scholar]
- Roosta, H.; Estaji, A.; Niknam, F. Effect of iron, zinc and manganese shortage-induced change on photosynthetic pigments, some osmoregulators and chlorophyll fluorescence parameters in lettuce. Photosynthetica 2018, 56, 606–615. [Google Scholar] [CrossRef]
- Yao, Y.; Jiang, C. Alteration of mineral elements and FTIR characterization of Hovenia orange live seedlings under iron deficiency stress. China South Fruit Tree 2016, 45, 10–14. (In Chinese) [Google Scholar]
- Hussain, A.; Nazir, F.; Fariduddin, Q. Polyamines (spermidine and putrescine) mitigate the adverse effects of manganese induced toxicity through improved antioxidant system and photosynthetic attributes in Brassica juncea. Chemosphere 2019, 236, 124830. [Google Scholar] [CrossRef]
- Kong, J.; Dong, Y.; Xu, L.; Liu, S.; Bai, X. Effects of foliar application of salicylic acid and nitric oxide in alleviating iron deficiency induced chlorosis of Arachis hypogaea L. Bot. Stud. 2014, 55, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, S.; Wang, G.; Xu, H.; Liu, J.; Luo, J.; Shen, Y. Exogenous Spermidine Improves Chilling Tolerance in Sweet Corn Seedlings by Regulation on Abscisic Acid, ROS and Ca2+ Pathways. Plant Biol. 2021, 64, 487–499. [Google Scholar] [CrossRef]
- Li, L.; Gu, W.; Li, C.; Li, W.; Li, C.; Li, J.; Wei, S. Exogenous spermidine improves drought tolerance in maize by enhancing the antioxidant defence system and regulating endogenous polyamine metabolism. Crop Pasture Sci. 2018, 69, 1076–1091. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Y.; Ma, X.J.; Ouyang, Z.; Deng, L.; Shen, S.; Dong, X.; Du, N.; Dong, H.; Guo, Z.; et al. Melatonin Alleviates Copper Toxicity via Improving ROS Metabolism and Antioxidant Defense Response in Tomato Seedlings. Antioxidants 2022, 11, 758. [Google Scholar] [CrossRef]
- Du, J.; Shu, S.; An, Y.; Zhou, H.; Guo, S.; Sun, J. Influence of exogenous spermidine on carbon–nitrogen metabolism under Ca (NO3)2 stress in cucumber root. Plant Growth Regul. 2017, 81, 103–115. [Google Scholar] [CrossRef]
- Fatima, A.; Adnan, A.S.; Arif, S.A.; Abdullah, Q.A.A. The effects of free amino acids profiles on seeds germination/dormancy and seedlings development of two genetically different cultivars of Yemeni Pomegranates. Stress Physiol. Biochem. 2012, 8, 114–137. [Google Scholar]
- Long, W.; Li, Q.; Wan, N.; Feng, D.; Kong, F.; Zhou, Y.; Yuan, J. Root morphological and physiological characteristics in maize seedlings adapted to low iron stress. PLoS ONE 2020, 15, e0239075. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Qin, C.; Qi, M.; Dong, X.; Parvaiz, P.; FathiAbd_Allah, E.; Zhang, L. Spermine application alleviates salinity induced growth and photosynthetic inhibition in Solanum lycopersicum by modulating osmolyte and secondary metabolite accumulation and differentially regulating antioxidant metabolism. Plant Physiol. Biochem. 2019, 144, 1–13. [Google Scholar] [CrossRef]
- Kabir, A.H.; Paltridge, N.G.; Able, A.J.; Paull, J.G.; Stangoulis, J. Natural variation for Fe-efficiency is associated with upregulation of Strategy I mechanisms and enhanced citrate and ethylene synthesis in Pisum sativum L. Planta 2012, 235, 1409–1419. [Google Scholar] [CrossRef]
- Boquete, M.T.; Lang, I.; Weidinger, M.; Richards, C.L.; Alonso, C. Patterns and mechanisms of heavy metal accumulation and tolerance in two terrestrial moss species with contrasting habitat specialization. Environ. Exp. Bot. 2021, 182, 104336. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, P.; Gu, Z.; Tao, Y.; Shen, C.; Zhou, Y.; Han, Y.; Yang, R. Ca2+ involved in GABA signal transduction for phenolics accumulation in germinated hulless barley under NaCl stress. Food Chem. X 2019, 2, 100023. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Zhou, H.; Sang, T.; Shu, S.; Sun, J.; Guo, S. Effects of Exogenous Spermidine on Carbon and Nitrogen Metabolism in Tomato Seedlings under High Temperature. Am. Soc. Hortic. Sci. 2016, 141, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Lamhamdi, M.; Bakrim, A.; Aarab, A.; Lafont, R.; Sayah, F. A comparison of lead toxicity using physiological and enzymatic parameters on spinach (Spinacia oleracea L.) and wheat (Triticum aestivum L.) growth. Moroc. J. Biol. 2010, 12, 6–7. [Google Scholar]
- Srivastava, R.K.; Pandey, P.; Rajpoot, R.; Rani, A.; Dubey, R.S. Cadmium and lead interactive effects on oxidative stress and antioxidative responses in rice seedlings. Protoplasma 2014, 251, 1047–1065. [Google Scholar] [CrossRef]
- Wang, Y.Y. Studies on the Role of bZIP44 Gene in Response to Iron Deficiency in Plants; Hefei University of Technology: Hefei, China, 2019. (In Chinese) [Google Scholar]
- Xie, C.; Jin, X.; Yan, L.; Shi, X.; Liu, H.; Xu, Y.; Dong, C. Effects of iron deficiency stress on physiology and gene expression of iron absorption and transpotation in Pyrus betulaefolia. Nanjing Agric. Univ. 2019, 42, 465–473. (In Chinese) [Google Scholar]
- Giehl, R.H.; Lima, J.; von Wirén, N. Localized Iron Supply Triggers Lateral Root Elongation in Arabidopsis by Altering the AUX1-Mediated Auxin Distribution. Plant Cell 2012, 24, 33–49. [Google Scholar] [CrossRef] [Green Version]
- Liu, W. Functional Study of Ethylene Response Factor ERF4/ERF72 in Response to Iron Deficiency in Apple Rootstock; China Agricultural University: Beijing, China, 2017. (In Chinese) [Google Scholar]
- Bai, Z.Q.; Zu, H.H.; Wang, R.; Gao, X.X.; Zou, T.; Chen, G.L.; Wu, J.W. Molecular role of ethylene in fruit ripening of Ziziphus jujube Mill. Plant Signal Behav. 2020, 15, 1834749. [Google Scholar] [CrossRef]
- Chen, P.F.; Chen, L.; Jiang, Z.R.; Wang, G.P.; Wang, S.H.; Ding, Y.F. Sucrose is involved in the regulation of iron deficiency responses in rice (Oryza sativa L.). Plant Cell Rep. 2018, 37, 789–798. [Google Scholar] [CrossRef]
- Bashir, K.; Hanada, K.; Shimizu, M.; Seki, M.; Nakanishi, H.; Nishizawa, N.K. Transcriptomic analysis of rice in response to iron deficiency and excess. Rice 2014, 7, 18. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
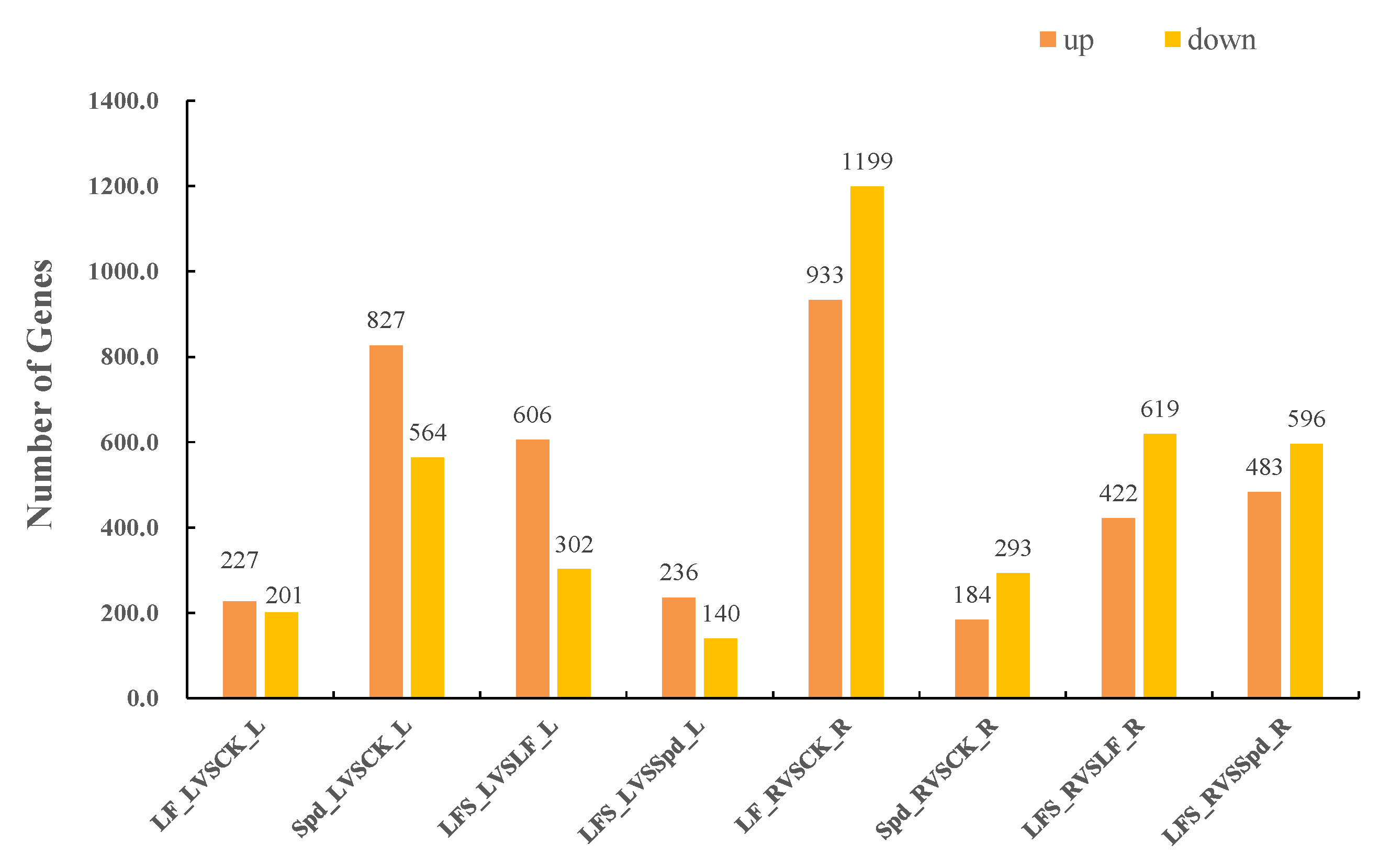{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
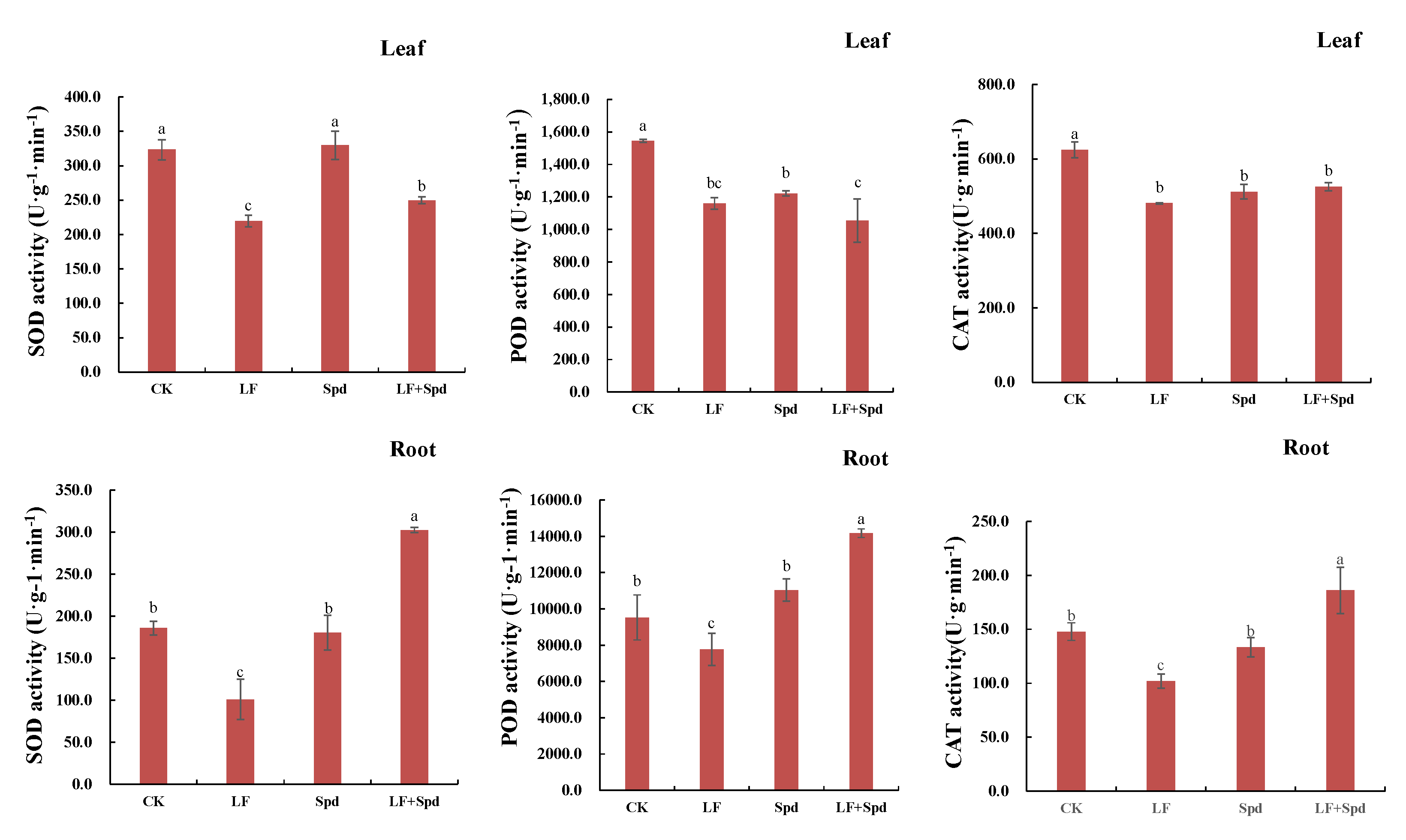{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Read | Valid Read | Valid Ratio (Reads) | Mapped Reads | Unique Mapped Reads | Q20% | Q30% |
|---|---|---|---|---|---|---|---|
| CK_L1 | 51,425,238 | 47,618,720 | 92.60 | 45,445,100 (95.44%) | 38,907,284 (81.71%) | 99.99 | 97.68 |
| CK_L2 | 41,647,086 | 39,867,334 | 95.73 | 38,103,955 (95.58%) | 32,333,283 (81.10%) | 99.99 | 97.78 |
| CK_L3 | 36,237,924 | 34,971,030 | 96.50 | 33,505,416 (95.81%) | 28,442,929 (81.33%) | 99.99 | 97.59 |
| CK_R1 | 41,884,204 | 40,904,806 | 97.66 | 35,303,817 (86.31%) | 30,123,337 (73.64%) | 99.99 | 97.38 |
| CK_R2 | 47,810,190 | 46,772,676 | 97.83 | 42,676,467 (91.24%) | 36,960,152 (79.02%) | 99.98 | 98.37 |
| CK_R3 | 51,562,904 | 50,495,438 | 97.93 | 44,954,365 (89.03%) | 38,867,119 (76.97%) | 99.98 | 98.30 |
| LF_L1 | 45,353,480 | 42,425,970 | 93.55 | 40,385,907 (95.19%) | 34,478,301 (81.27%) | 99.99 | 97.79 |
| LF_L2 | 45,112,774 | 43,318,042 | 96.02 | 41,436,986 (95.66%) | 35,189,847 (81.24%) | 99.99 | 97.62 |
| LF_L3 | 47,262,530 | 45,751,138 | 96.80 | 43,773,386 (95.68%) | 37,222,647 (81.36%) | 99.99 | 97.73 |
| LF_R1 | 43,262,284 | 42,427,286 | 98.07 | 39,214,339 (92.43%) | 33,844,982 (79.77%) | 99.99 | 98.39 |
| LF_R2 | 52,854,702 | 51,760,038 | 97.93 | 47,780,225 (92.31%) | 41,266,840 (79.73%) | 99.99 | 98.51 |
| LF_R3 | 52,525,326 | 51,551,094 | 98.15 | 45,715,483 (88.68%) | 39,276,860 (76.19%) | 99.99 | 98.45 |
| LFS_L1 | 46,825,170 | 44,333,134 | 94.68 | 42,409,857 (95.66%) | 36,246,131 (81.76%) | 99.99 | 97.50 |
| LFS_L2 | 35,688,744 | 34,009,554 | 95.29 | 32,356,247 (95.14%) | 27,660,644 (81.33%) | 99.99 | 97.25 |
| LFS_L3 | 41,306,858 | 39,800,370 | 96.35 | 38,018,606 (95.52%) | 32,451,594 (81.54%) | 99.99 | 97.60 |
| LFS_R1 | 53,734,354 | 52,663,752 | 98.01 | 47,503,814 (90.20%) | 40,935,810 (77.73%) | 99.99 | 98.41 |
| LFS_R2 | 52,372,096 | 51,298,494 | 97.95 | 45,352,741 (88.41%) | 39,262,642 (76.54%) | 99.98 | 98.37 |
| LFS_R3 | 54,358,180 | 53,210,976 | 97.89 | 48,848,829 (91.80%) | 42,040,996 (79.01%) | 99.99 | 98.43 |
| Spd_L1 | 50,060,896 | 45,020,762 | 89.93 | 43,045,480 (95.61%) | 36,727,980 (81.58%) | 99.99 | 97.52 |
| Spd_L2 | 35,827,860 | 34,695,530 | 96.84 | 33,367,320 (96.17%) | 28,459,478 (82.03%) | 99.99 | 97.74 |
| Spd_L3 | 50,682,352 | 48,290,700 | 95.28 | 46,229,329 (95.73%) | 39,485,340 (81.77%) | 99.99 | 97.53 |
| Spd_R1 | 52,910,480 | 51,911,130 | 98.11 | 46,563,164 (89.70%) | 40,136,000 (77.32%) | 99.98 | 98.42 |
| Spd_R2 | 51,487,988 | 50,453,420 | 97.99 | 45,578,298 (90.34%) | 39,515,003 (78.32%) | 99.98 | 98.44 |
| Spd_R3 | 53,683,760 | 52,593,000 | 97.97 | 46,823,515 (89.03%) | 40,562,480 (77.13%) | 99.98 | 98.48 |
| Treatments | Shoot Fresh Weight/g | Root Fresh Weight/g | Shoot Dry Weight/g | Root Dry Weight/g | Total Fresh Weight/g | Total Dry Weight/g |
|---|---|---|---|---|---|---|
| CK | 5.43 ± 0.44 ab | 1.45 ± 0.12 b | 0.40 ± 0.01 b | 0.09 ± 0.01 a | 6.88 ± 0.43 b | 0.49 ± 0.02 b |
| LF | 3.80 ± 0.61 c | 1.16 ± 0.13 c | 0.29 ± 0.05 c | 0.06 ± 0.01 b | 4.96 ± 0.73 c | 0.35 ± 0.03 c |
| Spd | 6.19 ± 0.45 a | 1.81 ± 0.12 a | 0.48 ± 0.03 a | 0.10 ± 0.01 a | 7.99 ± 0.43 a | 0.58 ± 0.03 a |
| LF + Spd | 5.07 ± 0.24 b | 1.42 ± 0.19 bc | 0.36 ± 0.02 b | 0.08 ± 0.01 a | 6.49 ± 0.35 b | 0.45 ± 0.01 b |
| Treatments | Chl a mg·g−1 FW | Chl b mg·g−1 FW | Carotenoid mg·g−1 FW | Chl a + b mg·g−1 FW |
|---|---|---|---|---|
| CK | 1.47 ± 0.06 b | 0.66 ± 0.02 b | 0.24 ± 0.02 a | 3.20 ± 0.08 b |
| LF | 1.06 ± 0.03 d | 0.56 ± 0.04 c | 0.15 ± 0.01 b | 2.41 ± 0.08 d |
| Spd | 1.64 ± 0.02 a | 0.70 ± 0.01 a | 0.25 ± 0.02 a | 3.52 ± 0.07 a |
| LF + Spd | 1.31 ± 0.10 c | 0.63 ± 0.02 b | 0.18 ± 0.02 b | 2.93 ± 0.16 c |
| Treatments | Pn/ (μmol·m−1·s−1) | Gs/ (mmol·m−1·s−1) | Ci/ (mmol·mol−1) | Tr/ (mmol·m−1·s−1) |
|---|---|---|---|---|
| CK | 7.56 ± 0.10 b | 180.55 ± 1.68 a | 130.91 ± 1.58 c | 3.55 ± 0.00 a |
| LF | 3.85 ± 0.08 d | 92.73 ± 1.55 c | 136.26 ± 2.55 b | 2.04 ± 0.02 c |
| Spd | 8.92 ± 0.03 a | 178.94 ± 1.28 a | 122.71 ± 0.80 d | 3.54 ± 0.02 a |
| LF + Spd | 4.73 ± 0.12 c | 139.87 ± 0.43 b | 153.42 ± 1.47 a | 2.90 ± 0.05 b |
| Treatments | Fv/Fm | ETR | ΦPSII | qP | NPQ |
|---|---|---|---|---|---|
| CK | 0.763 ± 0.007 b | 120.562 ± 0.95 b | 0.276 ± 0.002 b | 0.468 ± 0.026 b | 1.380 ± 0.038 c |
| LF | 0.706 ± 0.009 d | 75.706 ± 3.14 d | 0.173 ± 0.007 d | 0.302 ± 0.012 d | 2.566 ± 0.056 a |
| Spd | 0.776 ± 0.008 a | 146.958 ± 0.77 a | 0.337 ± 0.002 a | 0.518 ± 0.004 a | 1.314 ± 0.012 c |
| LF + Spd | 0.741 ± 0.002 c | 106.407 ± 1.21 c | 0.243 ± 0.003 c | 0.416 ± 0.006 c | 1.851 ± 0.022 b |
| Treatments | Oxalic Acid/(mg·g−1) | Malic Acid/(µg·g−1) | Citric Acid/(µg·g−1) | Acetic Acid/(µg·g−1) |
|---|---|---|---|---|
| CK | 2.68 ± 0.13 a | 357.77 ± 28.45 c | 90.95 ± 4.85 c | 114.52 ± 15.87 c |
| LF | 2.73 ± 0.13 a | 606.70 ± 97.39 bc | 162.00 ± 23.73 b | 57.54 ± 12.40 d |
| Spd | 2.24 ± 0.09 b | 684.47 ± 167.26 b | 133.55 ± 9.60 bc | 210.01 ± 11.88 b |
| LF + Spd | 2.75 ± 0.35 a | 1654.85 ± 218.26 a | 241.63 ± 40.10 a | 236.42 ± 15.10 a |
| Treatments | Free Polyamine (nmol·g−1) | Soluble Conjugated Polyamine (nmol·g−1) | Bound Polyamine (nmol·g−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Put | Spd | Spm | Put | Spd | Spm | Put | Spd | Spm | |
| CK | 662.64 ± 16.76 b | 429.20 ± 57.33 b | 142.69 ± 1.68 b | 111.21 ± 39.69 c | 30.44 ± 3.91 d | 60.37 ± 2.54 c | 828.56 ± 91.03 c | 238.62 ± 27.29 c | 451.48 ± 36.43 c |
| LF | 545.76 ± 43.10 c | 568.52 ± 43.73 b | 160.10 ± 3.17 ab | 229.51 ± 46.45 b | 85.60 ± 4.36 b | 96.29 ± 2.73 b | 6909.74 ± 433.32 a | 2064.22 ± 130.05 a | 794.57 ± 43.54 b |
| Spd | 848.99 ± 64.31 a | 574.61 ± 30.55 b | 160.37 ± 13.66 ab | 129.79 ± 18.81 c | 55.02 ± 4.26 c | 63.07 ± 2.60 c | 1060.74 ± 47.86 c | 308.22 ± 14.31 c | 496.38 ± 18.08 c |
| LF + Spd | 702.82 ± 55.56 b | 878.03 ± 140.96 a | 170.73 ± 12.87 a | 408.15 ± 78.32 a | 103.22 ± 4.38 a | 119.36 ± 9.19 a | 4963.23 ± 340.45 b | 1479.83 ± 102.24 b | 1103.26 ± 55.84 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Zhao, Y.; Yao, Q.; Liu, F.; Li, X.; Jin, X.; Zhang, Y.; Ahammed, G.J. Comparative Physiological and Transcriptomic Analyses Reveal Mechanisms of Exogenous Spermidine-Induced Tolerance to Low-Iron Stress in Solanum lycopersicum L. Antioxidants 2022, 11, 1260. https://doi.org/10.3390/antiox11071260
Shi Y, Zhao Y, Yao Q, Liu F, Li X, Jin X, Zhang Y, Ahammed GJ. Comparative Physiological and Transcriptomic Analyses Reveal Mechanisms of Exogenous Spermidine-Induced Tolerance to Low-Iron Stress in Solanum lycopersicum L. Antioxidants. 2022; 11(7):1260. https://doi.org/10.3390/antiox11071260
Chicago/Turabian StyleShi, Yu, Yihong Zhao, Qi Yao, Feng Liu, Xiumin Li, Xiu Jin, Yi Zhang, and Golam Jalal Ahammed. 2022. "Comparative Physiological and Transcriptomic Analyses Reveal Mechanisms of Exogenous Spermidine-Induced Tolerance to Low-Iron Stress in Solanum lycopersicum L" Antioxidants 11, no. 7: 1260. https://doi.org/10.3390/antiox11071260