
Apoptotic p53 Gene Expression in the Regulation of Persistent Organic Pollutant (POP)-Induced Oxidative Stress in the Intertidal Crab Macrophthalmusjaponicus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
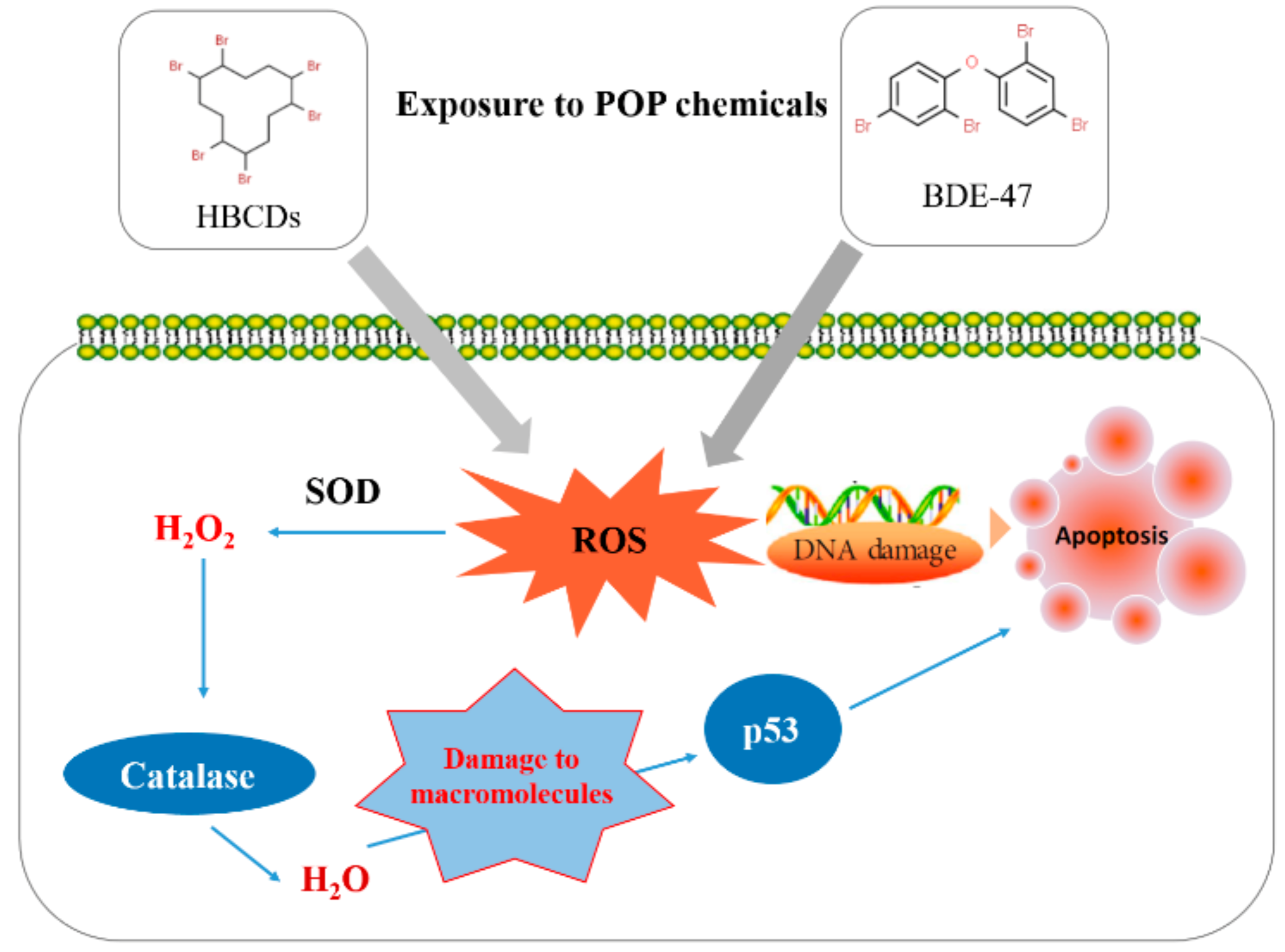
:1. Introduction
2. Materials and Methods
2.1. Organisms and Exposure Experiments
2.2. Macrophthalmus Japonicus p53 (Mjp53)
2.3. Basal Levels of Mjp53 by Tissue and Expression Analysis of Mjp53 Gene
2.4. Data Analysis
3. Results
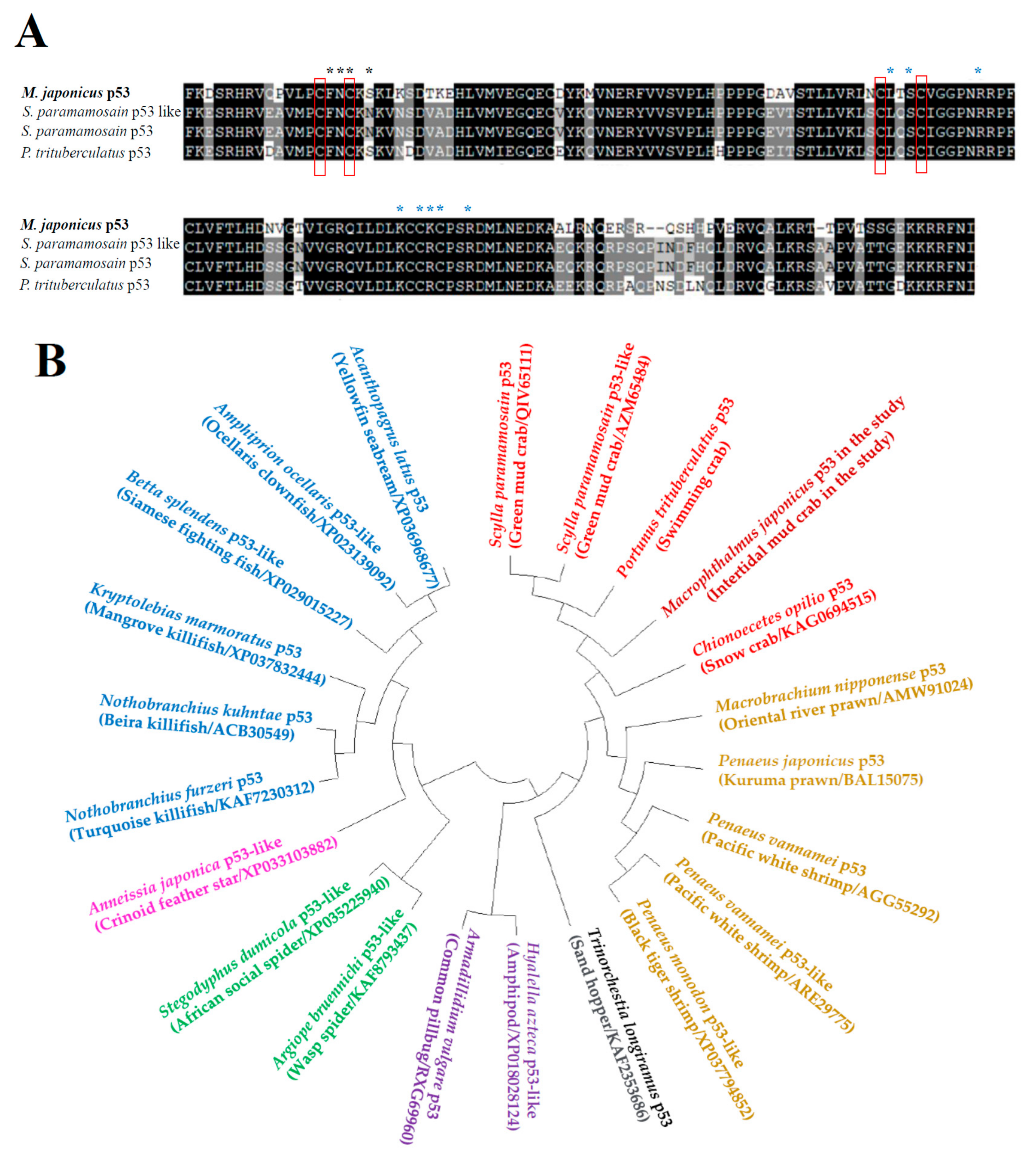
3.1. Identification and Phylogenetic Analysis of Mjp53 Gene
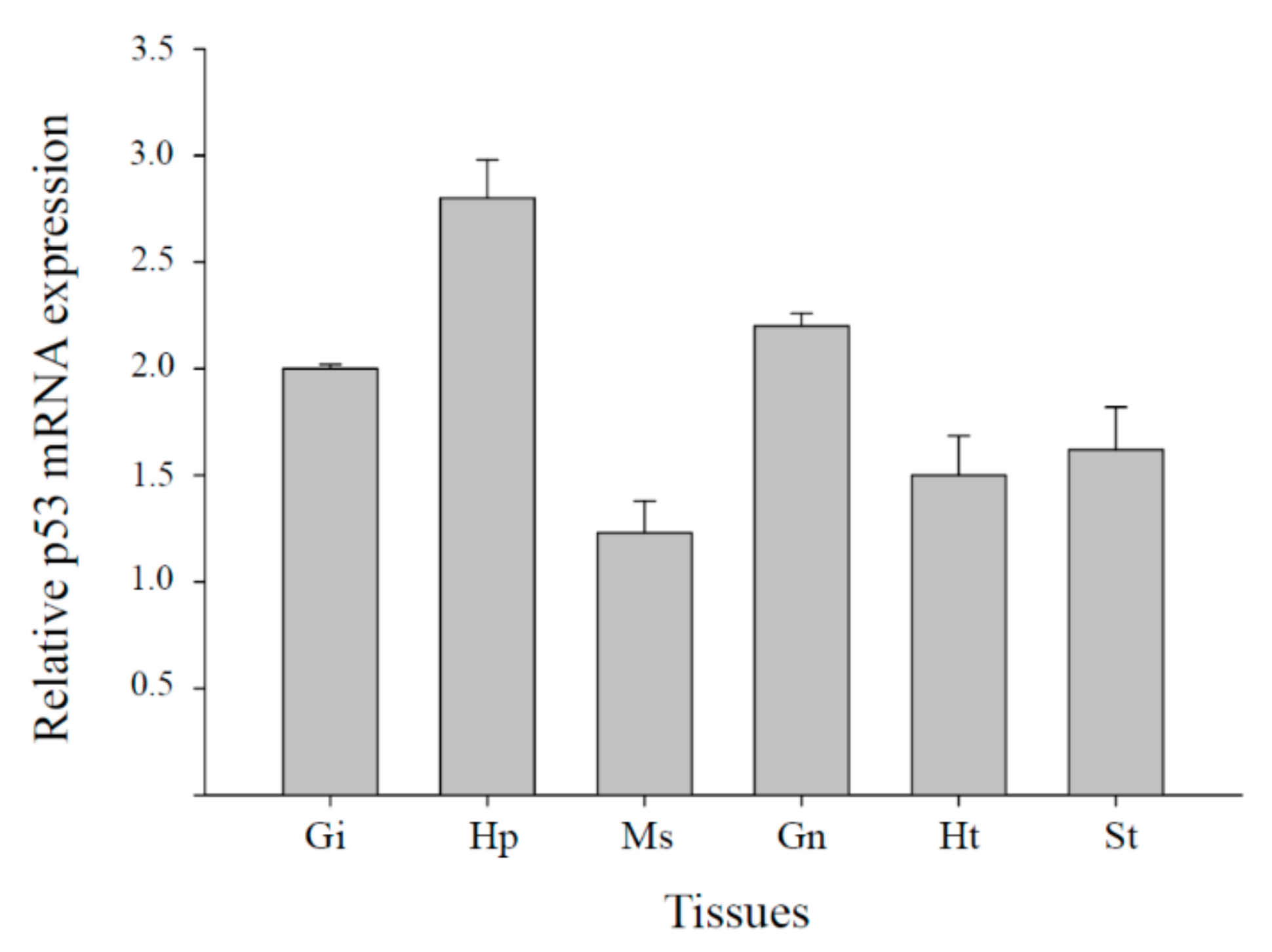
3.2. Tissue Distribution of Mjp53 Expression
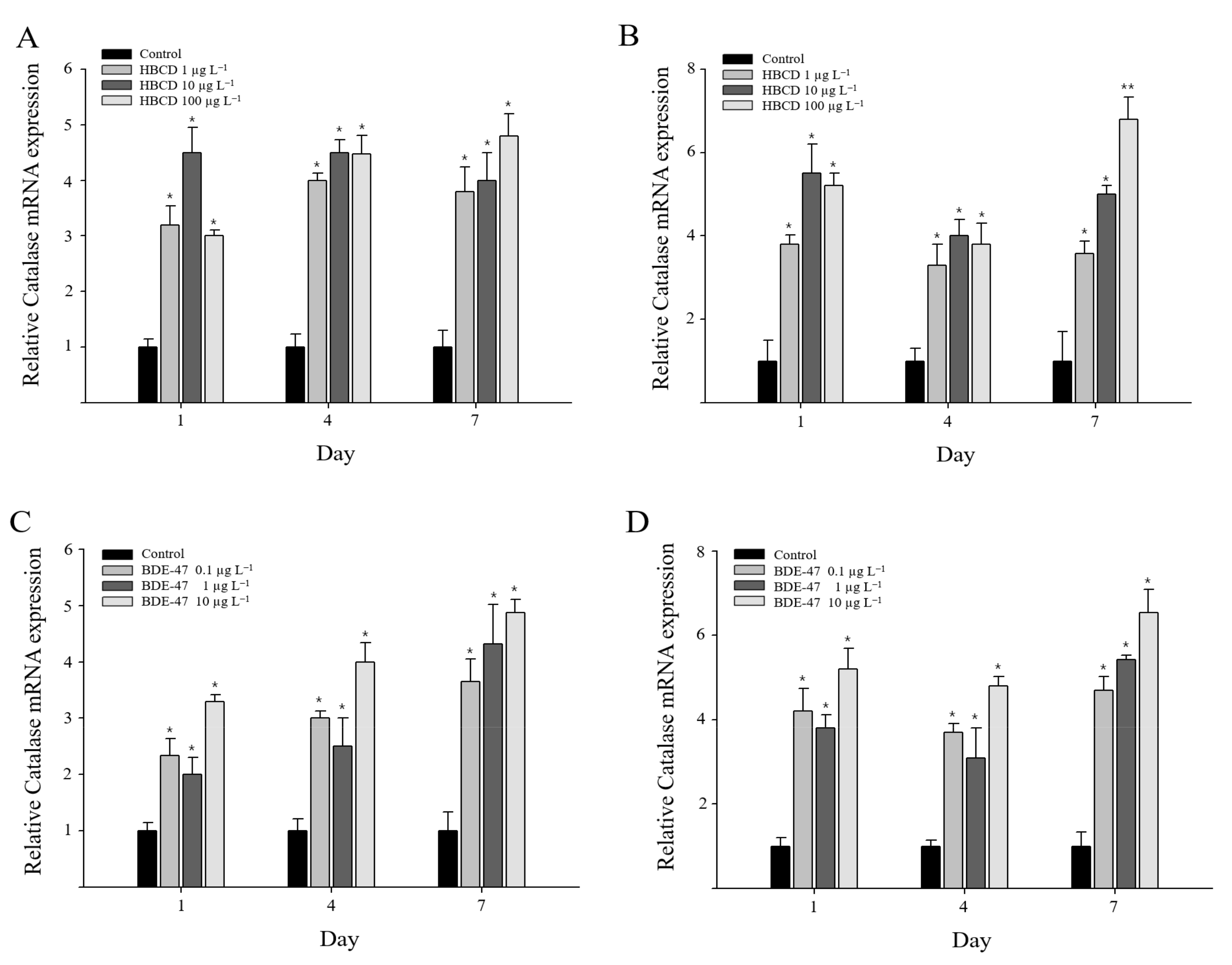
3.3. Catalase Gene Expression in Oxidative Stress Responses to Exposure of HBCD or BDE-47
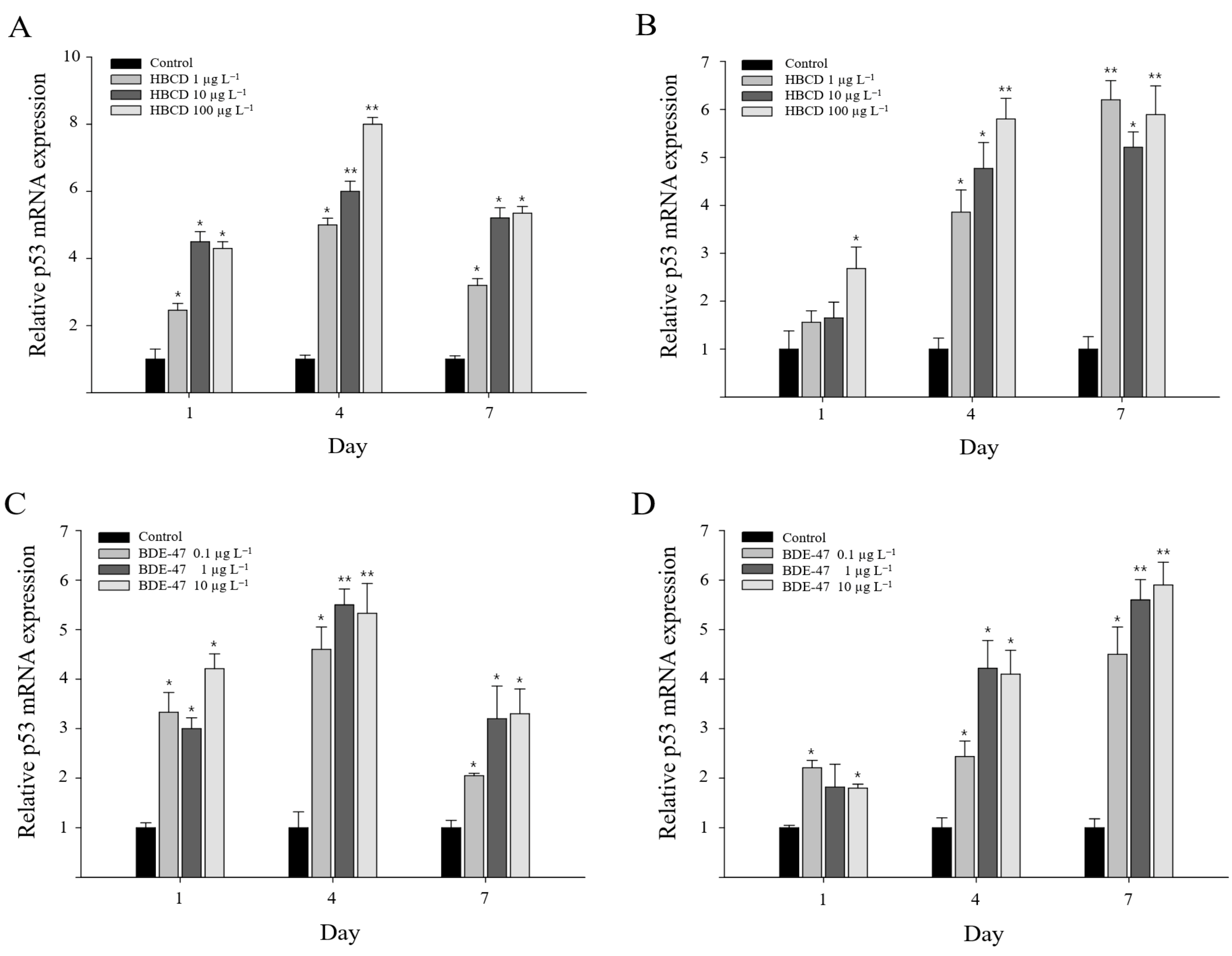
3.4. Mjp53 Gene Expression Responses to HBCD or BDE-47 Exposure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, X.; Sun, R.; Chen, Y.; Zhang, X.; Cui, Z. Temporal-spatial distribution and diastereoisomer pattern of hexabromocyclododecane in the vicinity of a chemical plant. J. Environ. Sci. 2019, 82, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Feng, Y.; Li, J.; Zhou, D.; Guo, R.; Ji, R.; Chen, J. The bioaccumulation, elimination, and trophic transfer of BDE-47 in the aquatic food chain of Chlorella pyrenoidosa-Daphnia magna. Environ. Pollut. 2020, 258, 113720. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Won, E.J.; Lee, M.C.; Seo, J.S.; Lee, S.J.; Lee, J.S. Developmental retardation, reduced fecundity, and modulated expression of the defensome in the intertidal copepod Tigriopus japonicus exposed to BDE-47 and PFOS. Aquat. Toxicol. 2015, 165, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Saquib, Q.; Siddiqui, M.A.; Ahmed, J.; Al-Salim, A.; Ansari, S.M.; Faisal, M.; Al-Khedhairy, A.A.; Musarrat, J.; AlWathnani, H.A.; Alatar, A.A.; et al. Hazards of low dose flame-retardants (BDE-47 and BDE-32): Influence on transcriptome regulation and cell death in human liver cells. J. Hazard. Mater. 2016, 308, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, N.; Gewurtz, S.B.; Drouillard, K.G.; Kolic, T.; Macpherson, K.; Reiner, E.J.; Bhavsar, S.P. Polybrominated diphenyl ethers (PBDEs) in Great Lakes fish: Levels, patterns, trends and implications for human exposure. Sci. Total Environ. 2017, 576, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Pan, Z.J.; Li, M.; Hong, F.S.; Zhu, C.K.; Wu, N.; Chang, G.; Wang, H.; Zhao, X.X. BDE-47 induced apoptosis in zebrafish embryos through mitochondrial ROS-mediated JNK signaling. Chemosphere 2020, 258, 127385. [Google Scholar] [CrossRef]
- Chang, C.J.; Terrell, M.L.; Marcus, M.; Marder, M.E.; Panuwet, P.; Ryan, P.B.; Pearson, M.; Barton, H.; Barr, D.B. Serum concentrations of polybrominated biphenyls (PBBs), polychlorinated biphenyls (PCBs) and polybrominated diphenyl ethers (PBDEs) in the Michigan PBB Registry 40 years after the PBB contamination incident. Environ. Int. 2020, 137, 105526. [Google Scholar] [CrossRef]
- Barghi, M.; Shin, E.S.; Son, M.H.; Choi, S.D.; Pyo, H.; Chang, Y.S. Hexabromocyclododecane (HBCD) in the Korean food basket and estimation of dietary exposure. Environ. Pollut. 2016, 213, 268–277. [Google Scholar] [CrossRef]
- Dong, H.; Lu, G.; Yan, Z.; Liu, J.; Yang, H.; Nkoom, M. Bioconcentration and effects of hexabromocyclododecane exposure in crucian carp (Carassius auratus). Ecotoxicology 2018, 27, 313–324. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, F.; Zhang, J.; Shi, J.; Tao, H.; Ge, H.; Guo, W.; Liu, D.; Cai, Z. Toxicity and accumulation of 6-OH-BDE-47 and newly synthesized 6,6′-diOH-BDE-47 in early life-stages of Zebrafish (Danio rerio). Sci. Total Environ. 2021, 763, 143036. [Google Scholar] [CrossRef]
- Zhang, S.; Kuang, G.; Zhao, G.; Wu, X.; Zhang, C.; Lei, R.; Xia, T.; Chen, J.; Wang, Z.; Ma, R.; et al. Involvement of the mitochondrial p53 pathway in PBDE-47-induced SH-SY5Y cells apoptosis and its underlying activation mechanism. Food Chem. Toxicol. 2013, 62, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.C.; Yang, W.X. Characterization and expression pattern of p53 during spermatogenesis in the Chinese mitten crab Eriocheir sinensis. Mol. Biol. Rep. 2013, 40, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Qiu, N.; Wang, R.; Sun, Y.; Wang, X.; Jiang, D.; Meng, Y.; Zhou, F. Toxic effects and mechanism of 2,2′,4,4′-tetrabromodiphenyl ether (BDE-47) on Lemna minor. Chemosphere 2018, 193, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Yu, L.; Liu, C.; Yu, K.; Shi, X.; Yeung, L.W.Y.; Lam, P.K.S.; Wu, R.S.S.; Zhou, B. Hexabromocyclododecane-induced developmental toxicity and apoptosis in zebrafish embryos. Aquat. Toxicol. 2009, 93, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Li, D.; Shen, R.; Wang, X.; Shi, D. Mechanisms of hexabromocyclododecanes induced developmental toxicity in marine medaka (Oryzias melastigma) embryos. Aquat. Toxicol. 2014, 152, 173–185. [Google Scholar] [CrossRef]
- Chon, H.S.; Ohandja, D.G.; Voulvoulis, N. The role of sediments as a source of metals in river catchments. Chemosphere 2012, 88, 1250–1256. [Google Scholar] [CrossRef]
- Minor, M.L.; Mullarney, J.C.; Pilditch, C.A.; Huhn, K. Crab burrow aspect ratio influences particle capture rates on intertidal sandflats. Geo-Maine Lett. 2020, 40, 197–216. [Google Scholar] [CrossRef]
- Park, K.; Kim, W.S.; Choi, B.; Kwak, I.S. Expression levels of the immune-related p38 mitogen-activated protein kinase transcript in response to environmental pollutants on Macrophthalmus japonicus crab. Genes 2020, 11, 958. [Google Scholar] [CrossRef]
- Hwang, D.S.; Lee, J.S.; Rhee, J.S.; Han, J.; Lee, Y.M.; Kim, I.C.; Park, G.S.; Lee, J.; Lee, J.S. Modulation of p53 gene expression in the intertidal copepod Tigriopus japonicus exposed to alkylphenols. Mar. Environ. Res. 2010, 69, S77–S80. [Google Scholar] [CrossRef]
- Shi, D.; Lv, D.; Liu, W.; Shen, R.; Li, D.; Hong, H. Accumulation and developmental toxicity of hexabromocyclododecanes (HBCDs) on the marine copepod Tigriopus japonicus. Chemosphere 2017, 167, 155–162. [Google Scholar] [CrossRef]
- Park, K.; Kim, W.S.; Kwak, I.S. Endocrine-disrupting chemicals impair the innate immune prophenoloxidase system in the intertidal mud crab, Macrophthalmus japonicus. Fish Shellfish Immunol. 2019, 87, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Nikapitiya, C.; Kwak, I.S. Identification and expression of proteolysis response genes for Macrophthalmus japonicus exposure to irgarol toxicity. Ann. Limnol. Int. J. Limnol. 2016, 52, 65–74. [Google Scholar] [CrossRef]
- Chai, M.; Li, R.; Cong, S. Contamination of polybrominated diphenyl ethers (PBDEs) in urban mangroves of Southern China. Sci. Total Environ. 2019, 646, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Windsor, F.M.; Pereira, M.G.; Tyler, C.R.; Ormerod, S.J. River organisms as indicators of the distribution and sources of persistent organic pollutants in contrasting catchments. Environ. Pollut. 2019, 255, 113144. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guo, R.; Tang, S.; Zhu, F.; Zhang, S.; Yan, Z.; Chen, J. Single and mixture toxicities of BDE-47, 6-OH-BDE-47 and 6-MeO-BDE-47 on the feeding activity of Daphnia magna: From behavior assessment to neurotoxicity. Chemosphere 2018, 195, 542. [Google Scholar] [CrossRef]
- Tanaka, Y.; Fujiwara, M.; Shindo, A.; Yin, G.; Teraoka, H. Aroclor 1254 and BDE-47 inhibit dopaminergic function manifesting as changes in locomotion behaviors in zebrafish embryos. Chemosphere 2018, 193, 1207–1215. [Google Scholar] [CrossRef]
- Darnerud, P.O. Toxic effects of brominated flame retardants in man and in wildlife. Environ. Int. 2003, 29, 841–853. [Google Scholar] [CrossRef]
- Miguel, N.C.O.; Wajsenzon, I.J.R.; Takiya, C.M.; Andrade, L.R.; Tortelote, G.G.; Einicker-Lamas, M.; Allodi, S. Catalase, Bax and p53 expression in the visual system of the crab Ucides cordatus following exposure to ultraviolet radiation. Cell Tissue Res. 2007, 329, 159–168. [Google Scholar] [CrossRef]
- Hollmann, G.; Linden, R.; Giangrande, A.; Allodi, S. Increased p53 and decreased p21 accompany apoptosis induced by ultraviolet radiation in the nervous system of a crustacean. Aquat. Toxicol. 2016, 173, 1–8. [Google Scholar] [CrossRef]
- Cheng, C.H.; Su, Y.L.; Ma, H.L.; Deng, Y.Q.; Feng, J.; Chen, X.L.; Jie, Y.K.; Guo, Z.X. Effect of nitrite exposure on oxidative stress, DNA damage and apoptosis in mud crab (Scylla paramamosain). Chemosphere 2020, 239, 124668. [Google Scholar] [CrossRef]
- Cheng, C.H.; Ma, H.L.; Deng, Y.Q.; Feng, J.; Jie, Y.K.; Guo, Z.X. Oxidative stress, cell cycle arrest, DNA damage and apoptosis in the mud crab (Scylla paramamosain) induced by cadmium exposure. Chemosphere 2021, 263, 128277. [Google Scholar] [CrossRef] [PubMed]
- Vogt, G. Functional cytology of the hepatopancreas of decapod crustaceans. J. Morphol. 2019, 280, 1405–1444. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Sun, H.; Qian, Y.; Yang, J. Impairments of cadmium on vitellogenin accumulation in the hepatopancreas of freshwater crab Sinopotamon henanense. Environ. Sci. Pollut. Res. Int. 2017, 24, 18160–18167. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, K.; Kwak, I.-S. Apoptotic p53 Gene Expression in the Regulation of Persistent Organic Pollutant (POP)-Induced Oxidative Stress in the Intertidal Crab Macrophthalmusjaponicus. Antioxidants 2022, 11, 771. https://doi.org/10.3390/antiox11040771
Park K, Kwak I-S. Apoptotic p53 Gene Expression in the Regulation of Persistent Organic Pollutant (POP)-Induced Oxidative Stress in the Intertidal Crab Macrophthalmusjaponicus. Antioxidants. 2022; 11(4):771. https://doi.org/10.3390/antiox11040771
Chicago/Turabian StylePark, Kiyun, and Ihn-Sil Kwak. 2022. "Apoptotic p53 Gene Expression in the Regulation of Persistent Organic Pollutant (POP)-Induced Oxidative Stress in the Intertidal Crab Macrophthalmusjaponicus" Antioxidants 11, no. 4: 771. https://doi.org/10.3390/antiox11040771