
Neuroprotective Effects of Myrtenal in an Experimental Model of Dementia Induced in Rats

 ,
,  , , , and
, , , and 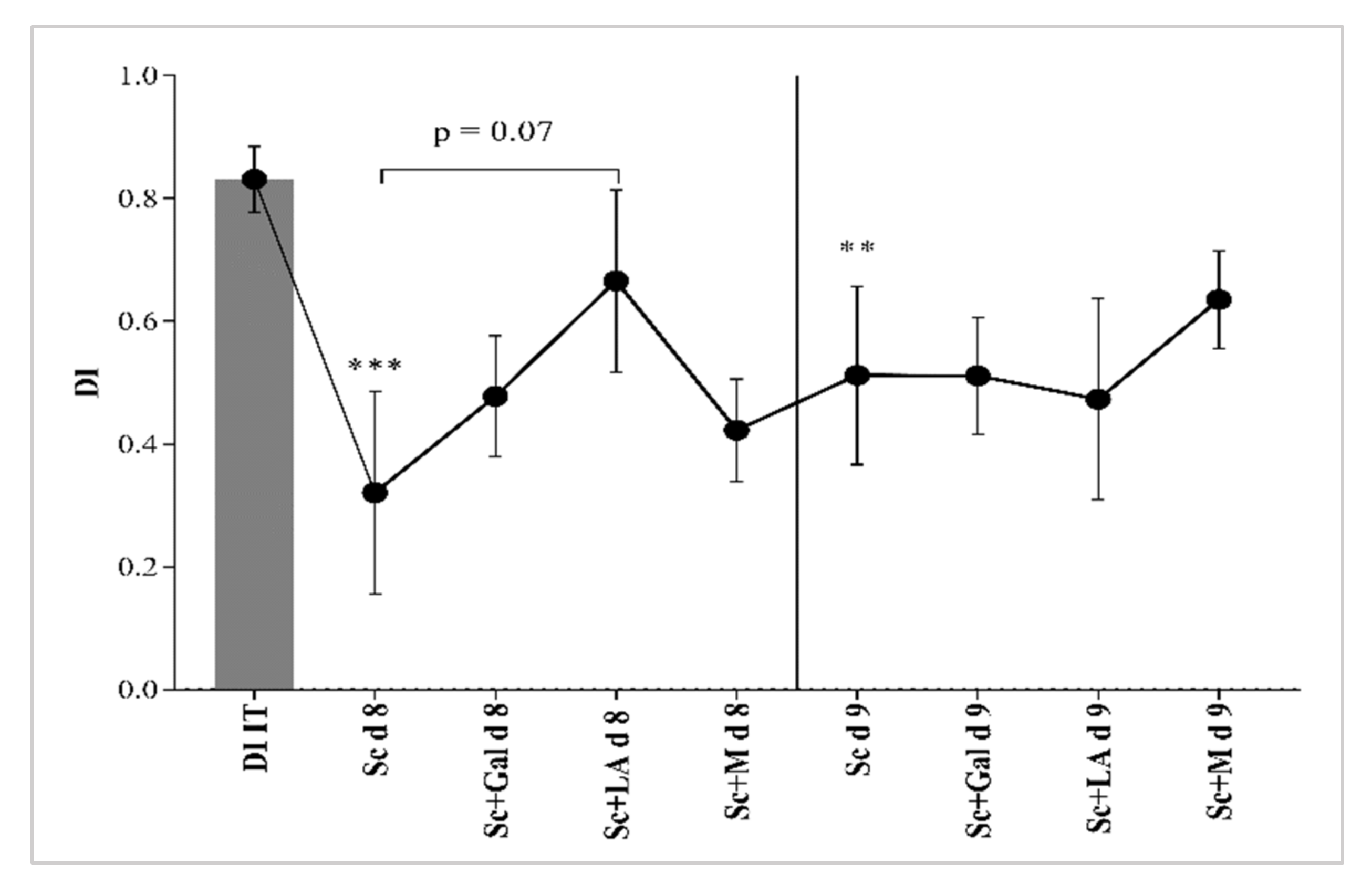{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
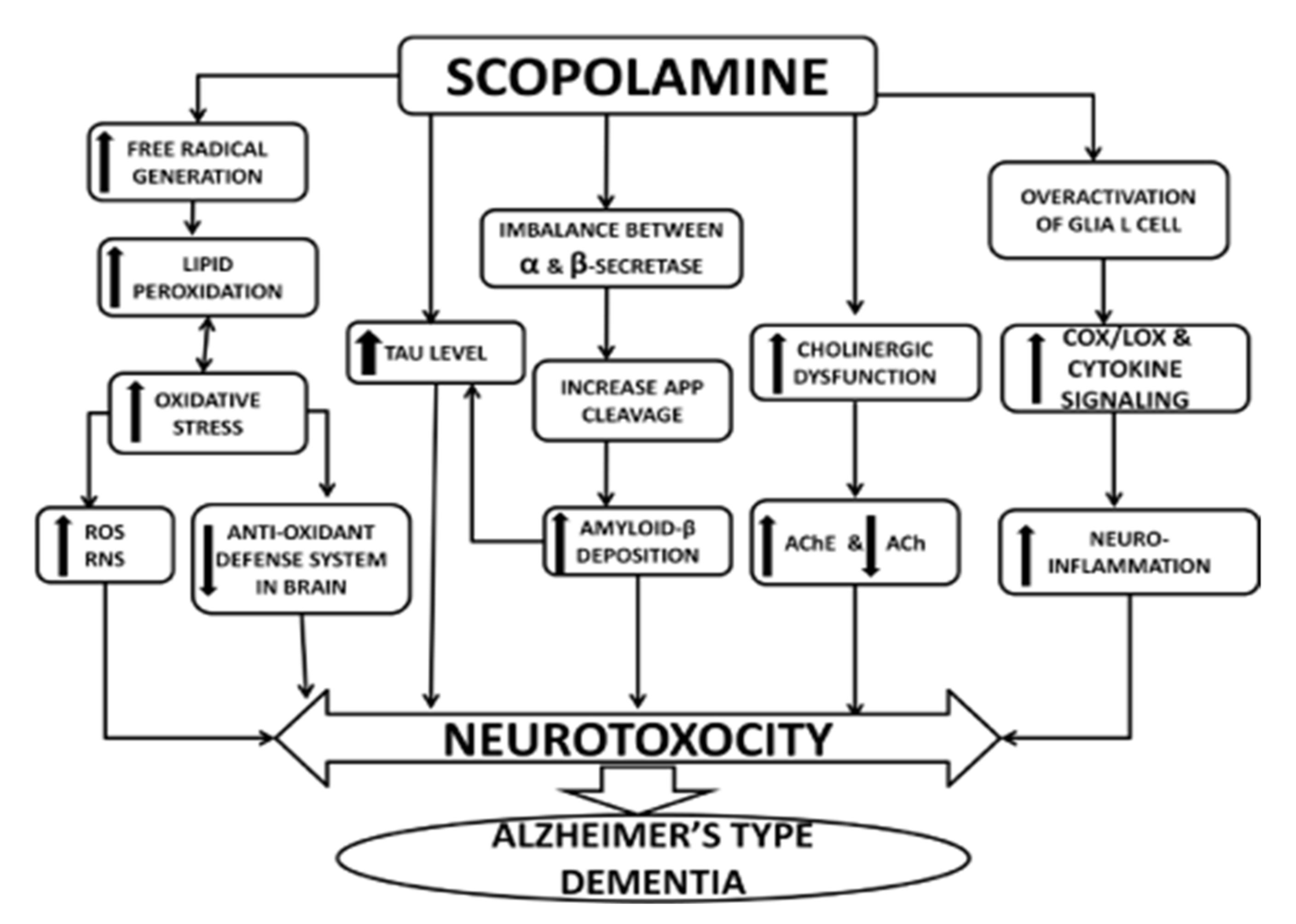
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Docking Study
2.3. Experimental Animals
2.4. Experimental Model of Dementia
2.5. Drug Treatment and Experimental Design
2.6. Behavioral Tests
2.7. Biochemical Studies
2.7.1. Evaluation of Oxidative Brain Status
2.7.2. Evaluation of Brain AChE Activity
2.7.3. Evaluation of Brain ACh Content
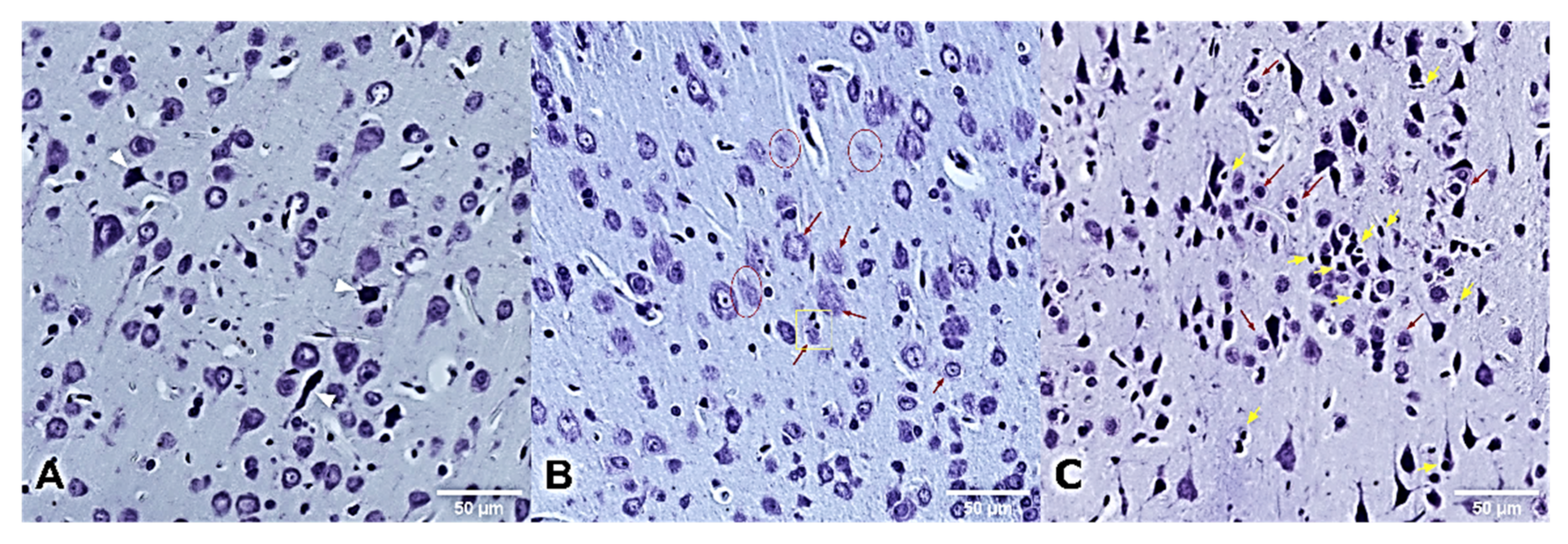
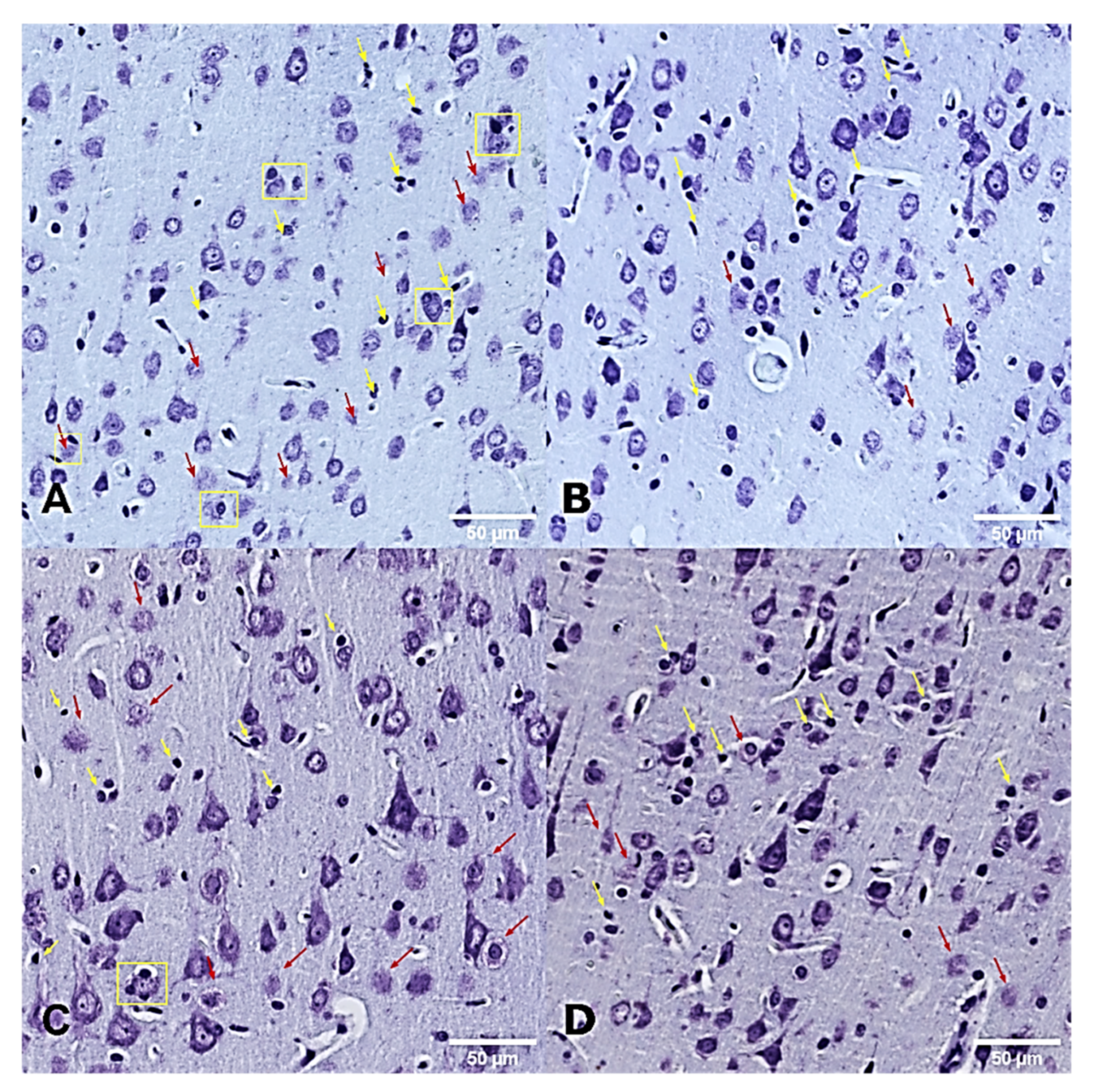
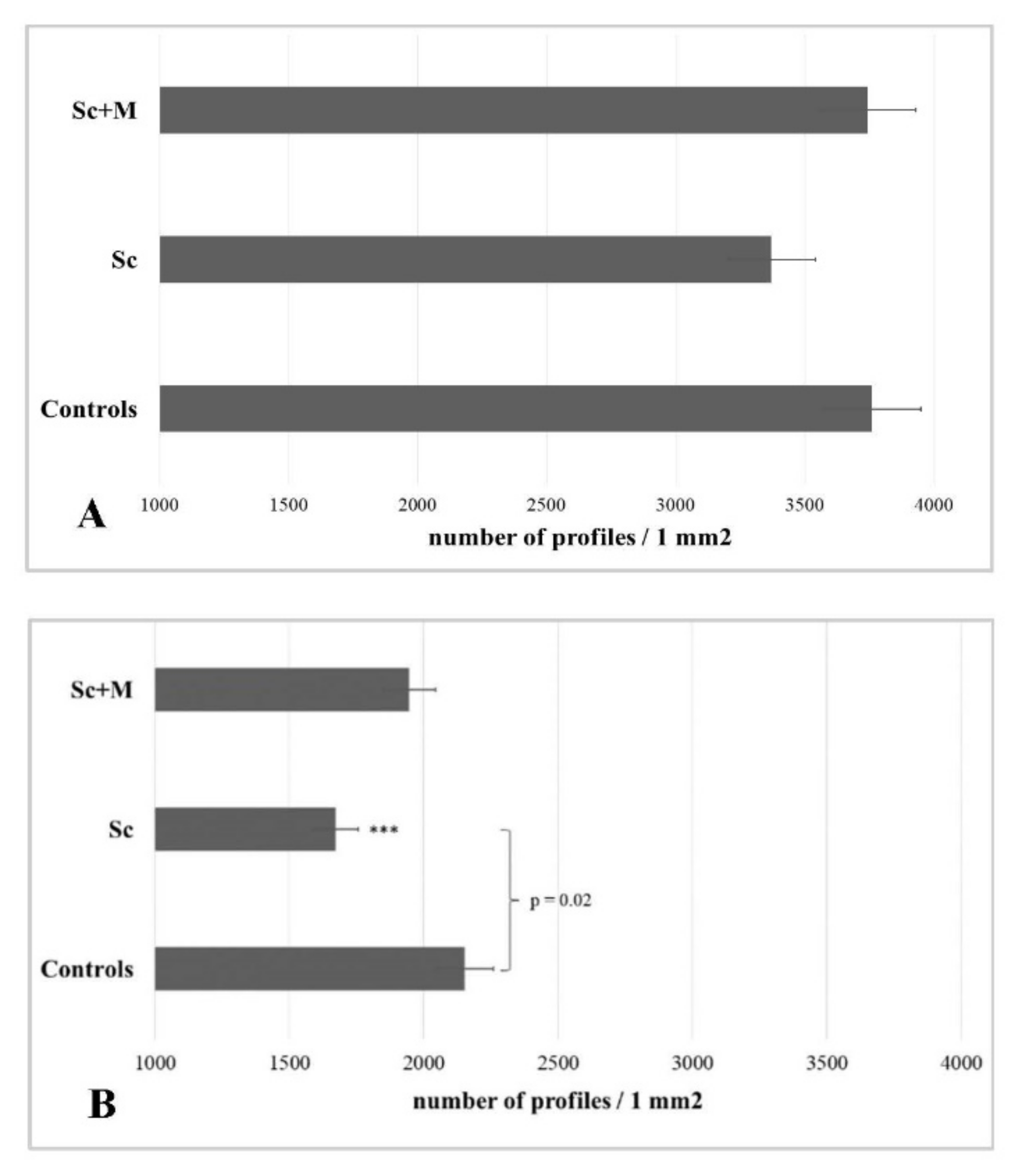
2.8. Histological Analysis
2.9. Statistical Analysis
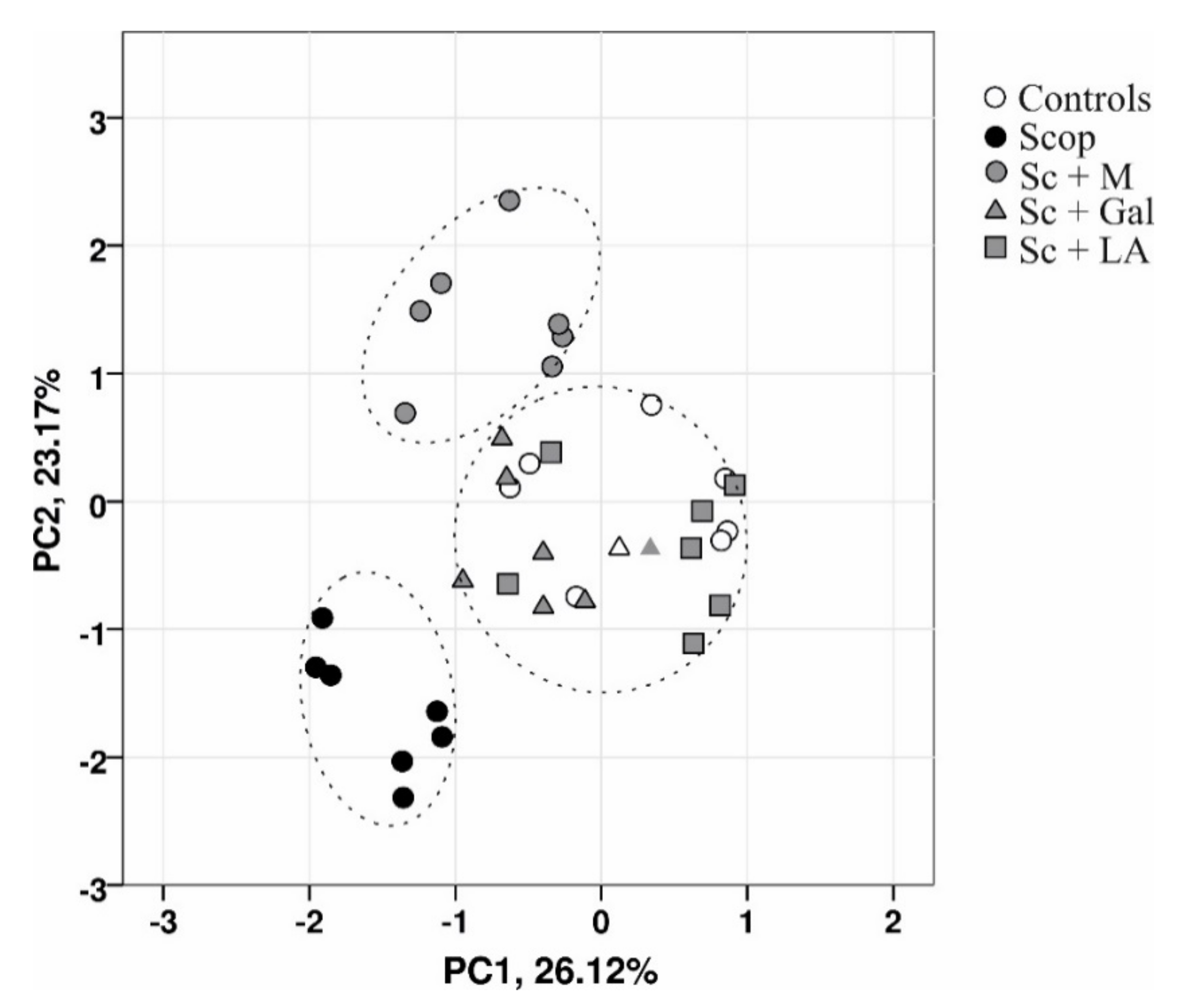
2.10. Principal Component Analysis
3. Results
3.1. Effects of Myrtenal on Demented Rats Behavior
3.1.1. Recognition Memory Status
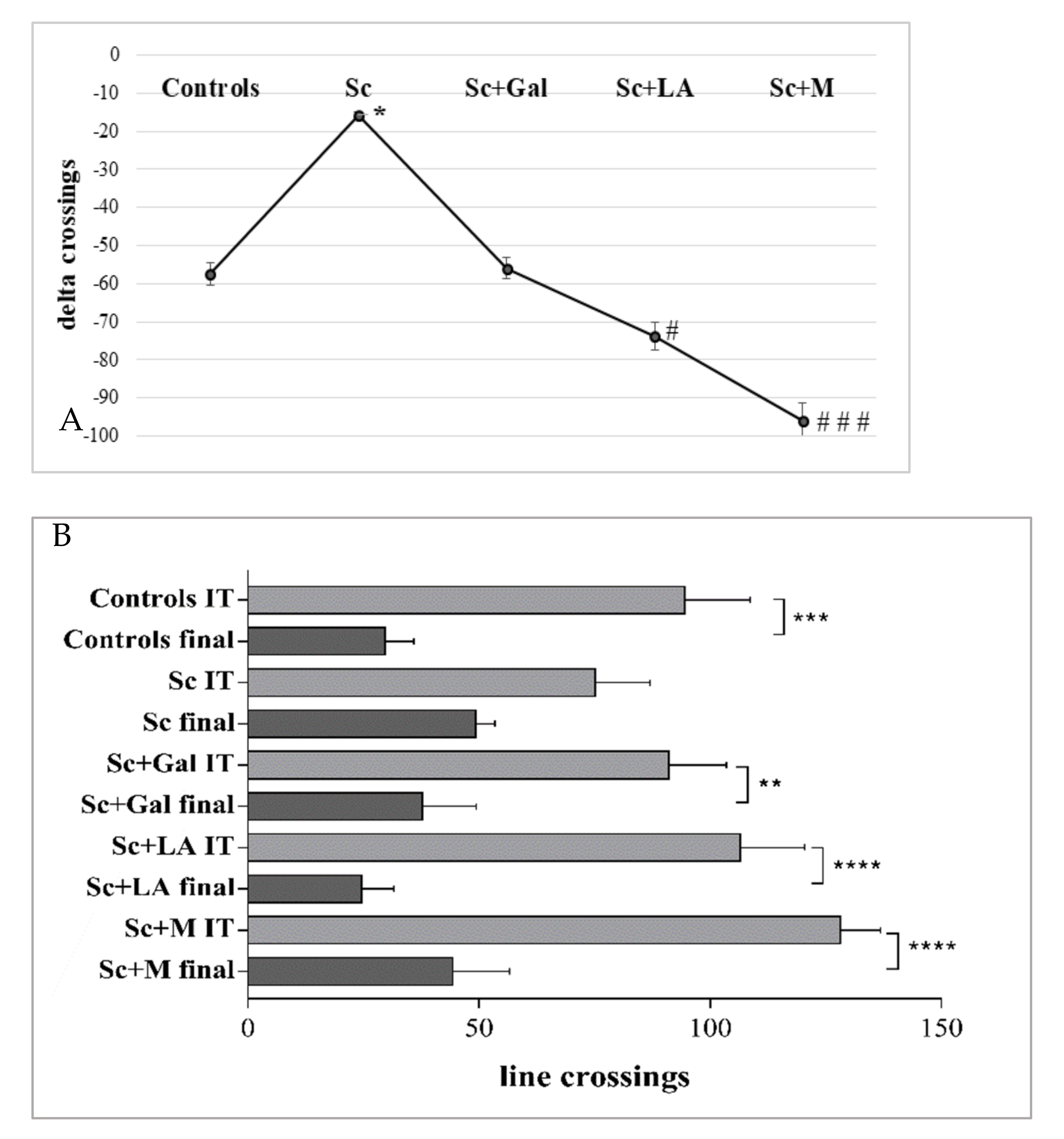
3.1.2. Effects of Myrtenal on Exploratory Activity
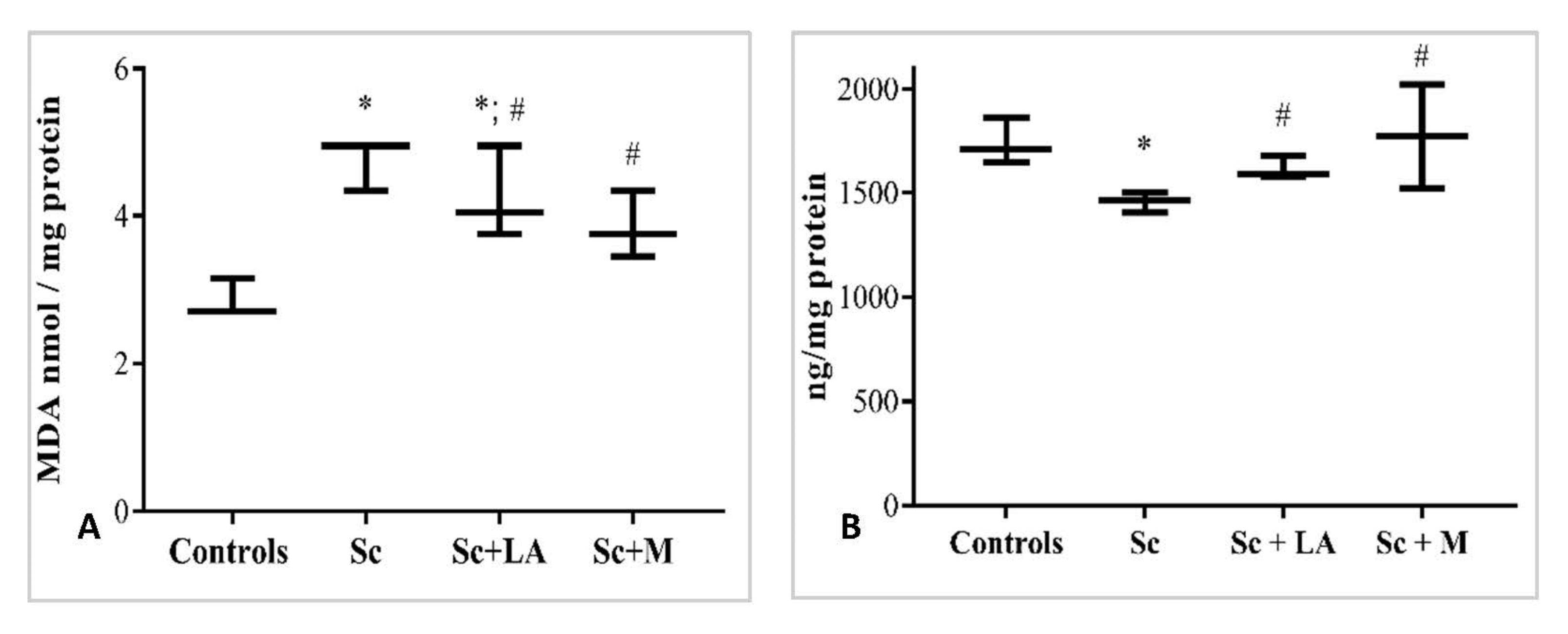
3.2. Effects of Myrtenal on Brain Oxidative Status in Dement Rodents
3.3. Effects of Myrtenal on Brain Cholinergic Transmission in Demented Rodents
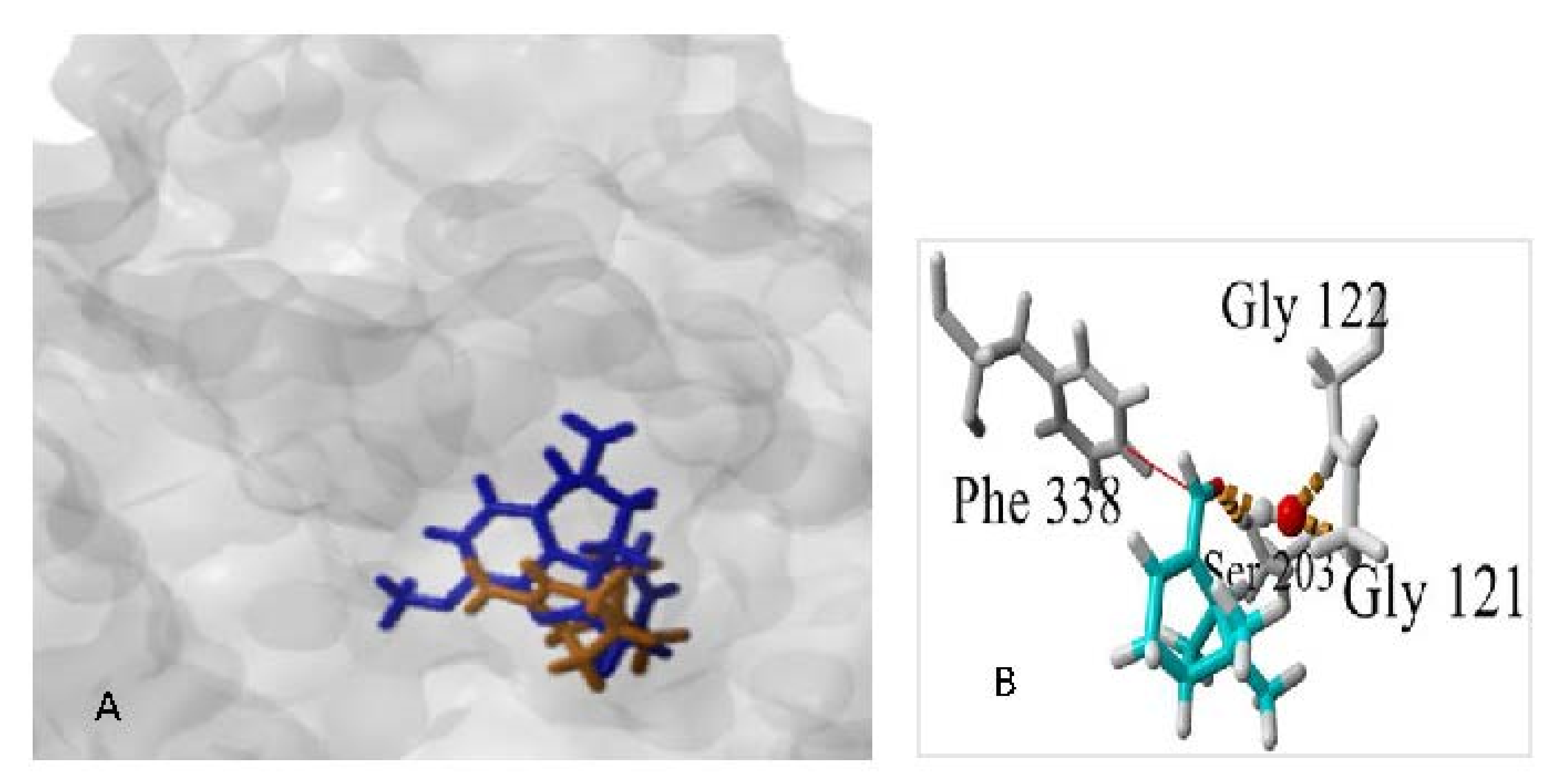
3.3.1. Docking Studies
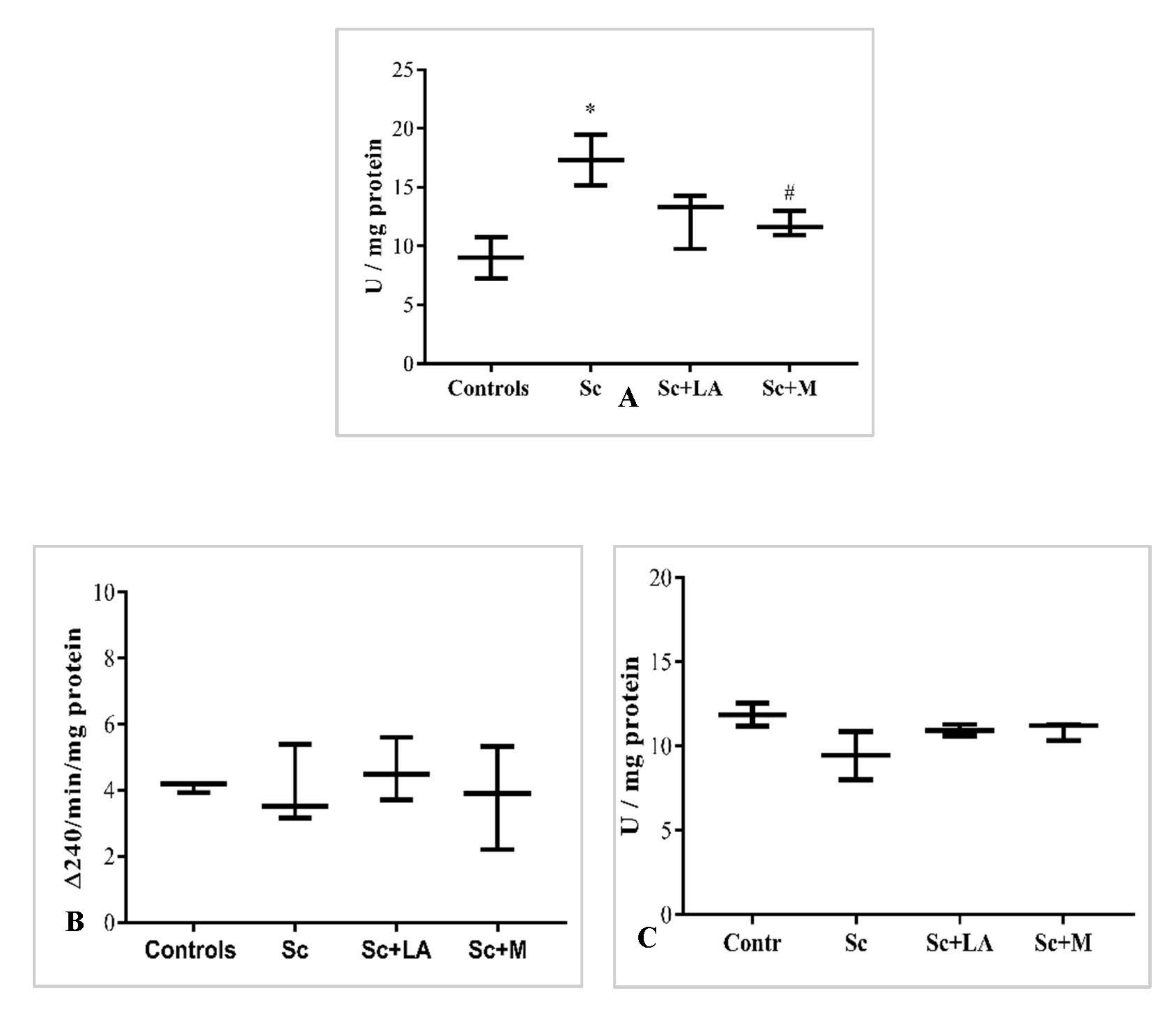
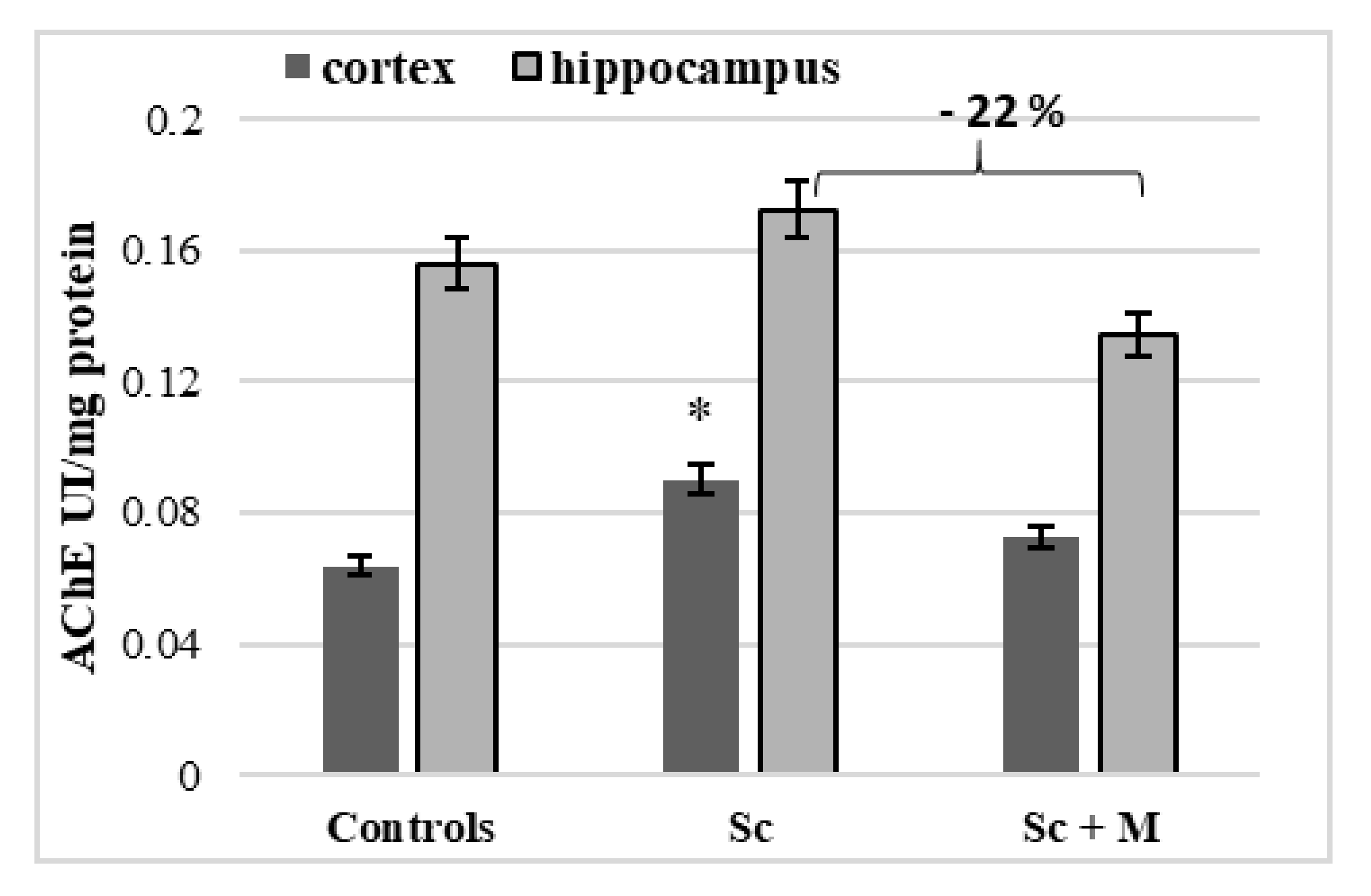
3.3.2. In Vivo Effect of Myrtenal on Brain AChE Activity
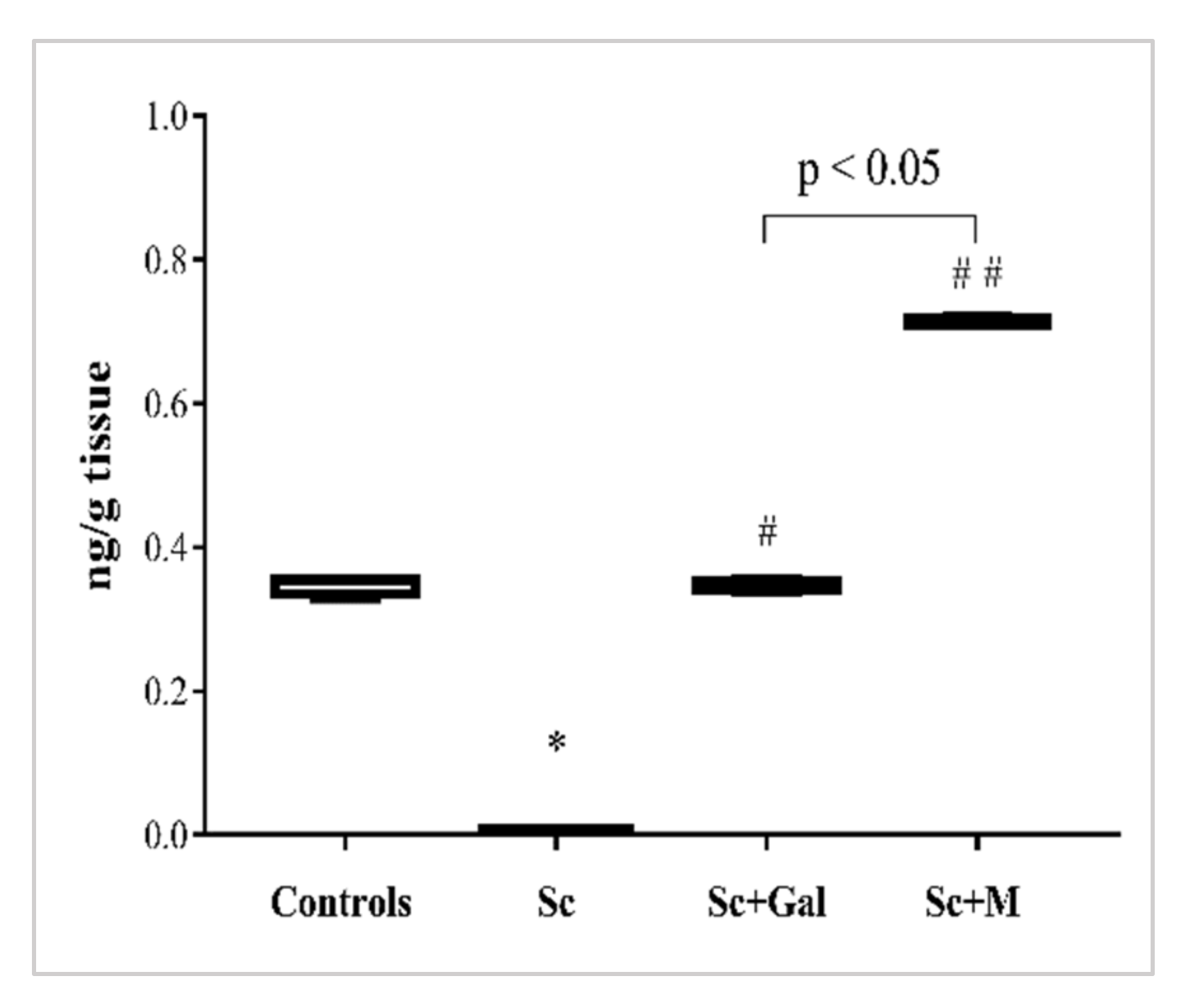
3.3.3. Effect of Myrtenal on Brain ACh Levels
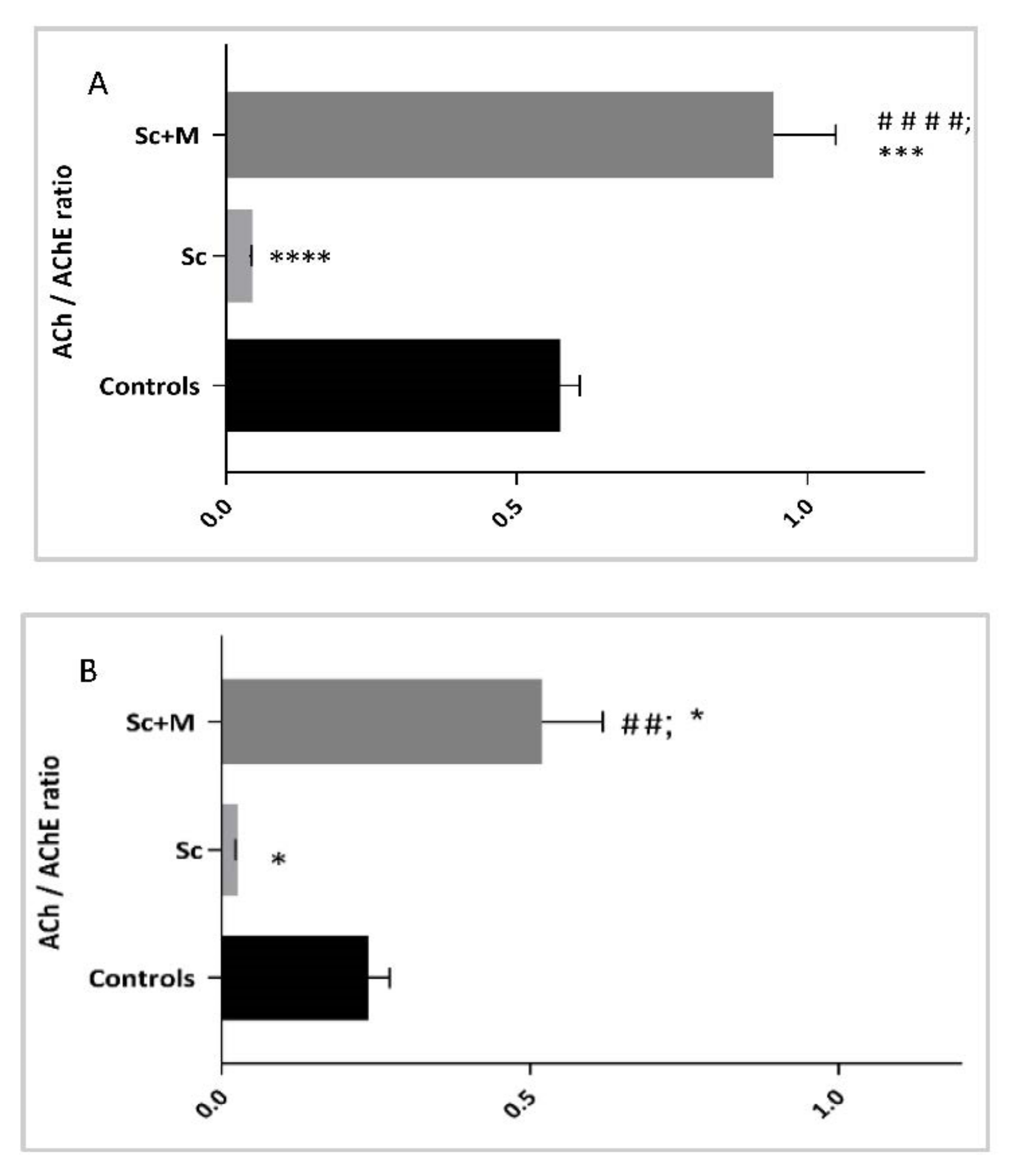
3.3.4. ACh Content/AChE Activity Ratio in the Brain
3.4. Histopathological Examinations
3.5. Summary Cluster Analysis of the Effects of Myrtenal on Scopolamine Induced Damage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rubinsztein, D.C. The roles of intracellular protein-degradation pathways in neurodegeneration. Nature 2006, 443, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Rockenstein, E.; Crews, L.; Masliah, E. Role of protein aggregation in mitochondrial dysfunction and neurodegeneration in Alzheimer’s and Parkinson’s diseases. Neuromol. Med. 2003, 4, 21–36. [Google Scholar] [CrossRef]
- Estrada Sánchez, A.M.; Mejía-Toiber, J.; Massieu, L. Excitotoxic neuronal death and the pathogenesis of Huntington’s disease. Arch. Med. Res. 2008, 39, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Pepeu, G.; Giovannini, M.G. The fate of the brain cholinergic neurons in neurodegenerative diseases. Brain Res. 2017, 167, 173–184. [Google Scholar] [CrossRef]
- Kar, S.; Slowikowski, S.P.; Westaway, D.; Mount, H.T. Interactions between Beta-Amyloid and Central Cholinergic Neurons: Implications for Alzheimer’s Disease. J. Psychiatry Neurosci. 2004, 29, 427–441. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC524960/ (accessed on 11 September 2020).
- Chen, X.; Decker, M. Multi-Target Compounds Acting in the Central Nervous System Designed from Natural Products. Curr. Med. Chem. 2013, 20, 1673–1685. [Google Scholar] [CrossRef]
- Wang, J.Y.; Wen, L.L.; Huang, Y.N.; Chen, Y.T.; Ku, M.C. Dual effects of antioxidants in neurodegeneration: Direct neuroprotection against oxidative stress and indirect protection via suppression of glia-mediated inflammation. Curr. Pharm. Des. 2006, 12, 3521–3533. [Google Scholar] [CrossRef]
- Ansari, M.A.; Scheff, S.W. Oxidative stress in the progression of Alzheimer disease in the frontal cortex. J. Neuropathol. Exp. Neurol. 2010, 69, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhao, B. Oxidative Stress and the Pathogenesis of Alzheimer’s disease. Oxid. Med. Cell. Longev. 2013, 2013, 316523. [Google Scholar] [CrossRef] [Green Version]
- Yoo, K.Y.; Park, S.-Y. Terpenoids as Potential Anti-Alzheimer’s Disease Therapeutics. Molecules 2012, 17, 3524–3538. [Google Scholar] [CrossRef]
- Hardie, J.; Isaacs, R.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Methyl salicylate and (−)-(1R, 5S)-myrtenal are plant-derived repellents for black bean aphid, Aphis fabae Scop. (Homoptera: Aphididae). J. Chem. Ecol. 1994, 20, 2847–2855. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.Y.; Lee, C.; Park, G.H.; Jang, J.H. Amelioration of Scopolamine-Induced Learning and Memory Impairment by α-Pinene in C57BL/6 Mice. Evid. Based Complement. Alternat. Med. 2017, 2017, 4926815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: https://patentimages.storage.googleapis.com/55/70/50/83e976db1028ce/US4190675.pdf (accessed on 10 July 2019).
- Kamchonwongpaisan, S.; Nilanonta, C.; Tarnchompoo, B.; Thebtaranonth, C.; Thebtaranonth, Y.; Clardy, J.; Kongsaeree, P.; Clardy, J. An antimalarial peroxide from Amomum krevanh Peirre. Tetrahedron Lett. 1995, 36, 1821–1824. [Google Scholar] [CrossRef]
- Saito, K.; Okabe, T.; Inamori, Y.; Tsujibo, H.; Miyake, Y.; Hiraoka, K.; Ishida, N. The biological properties of monoterpenes: Hypotensive effects on rats and antifungal activities on plant pathogenic fungi of monoterpenes. Mokuzai Gakkaishi 1996, 42, 677–680. [Google Scholar]
- Lingaiah, H.B.; Perumal, S.; Balasubramanian, M.P. Myrtenal attenuates diethylnitrosamine-induced hepatocellular carcinoma in rats by stabilizing intrinsic antioxidants and modulating apoptotic and anti-apoptotic cascades. Cell. Oncol. 2012, 35, 269–283. [Google Scholar]
- Dragomanova, S.T. Pharmacological, Toxicological and Neurobiological Studies of Myrtenal—A Biciclic Monoterpenoid of Natural Origin. Ph.D. Thesis, Institute of Neurobiology at Bulgarian Academy of Sciences, Sofia, Bulgaria, 2020. [Google Scholar]
- Kaufmann, D.; Dogra, A.K.; Wink, M. Myrtenal inhibits acetylcholinesterase, a known Alzheimer target. J. Pharm. Pharmacol. 2011, 63, 1368–1371. [Google Scholar] [CrossRef]
- Tancheva, L.P.; Lazarova, M.I.; Alexandrova, A.V.; Dragomanova, S.T.; Nicoletti, F.; Tzvetanova, E.R.; Hodzhev, Y.K.; Kalfin, R.E.; Miteva, S.A.; Mazzon, E.; et al. Neuroprotective Mechanisms of Three Natural Antioxidants on a Rat Model of Parkinson’s Disease: A Comparative Study. Antioxidants 2020, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Aggleton, J.P. One-trial object recognition by rats. Q. J. Exp. Psychol. 1985, 37, 279–294. [Google Scholar] [CrossRef]
- Ennaceur, A.; Delacour, J. A new one-trial test for neurobiological studies of memory in rats—1: Behavioral data. Behav. Brain Res. 1988, 31, 47–59. [Google Scholar] [CrossRef]
- Walsh, R.N.; Cummins, R.A. The open-field test: A critical review. Psychol. Bull. 1976, 83, 482–504. [Google Scholar] [CrossRef]
- Gould, T.D.; Dao, D.T.; Kovacsics, C.E. The Open Field Test. In Mood and Anxiety Related Phenotypes in Mice—Neuromethods; Gould, T., Ed.; Humana Press: Totowa, NJ, USA, 2009; Volume 42. [Google Scholar] [CrossRef]
- Tietze, F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: Applications to mammalian blood and other tissues. Anal. Biochem. 1969, 27, 502–522. [Google Scholar] [CrossRef]
- Hunter, F.E., Jr.; Gebicki, J.M.; Hoffsten, P.E.; Weinstein, J.; Scott, A. Swelling and Lysis of Rat Liver Mitochondria Induced by Ferrous Ions. J. Biol. Chem. 1963, 238, 828–835. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Günzler, W.A.; Vergin, H.; Müller, I.; Flohé, L. Glutathion-Peroxidase, VI. Die Reaktion der Glutathion-Peroxidase mit verschiedenen Hydroperoxiden. Hoppe Seyler’s Z. Physiol. Chem. 1972, 353, 1001–1004. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Elman, G.L.; Diane, C.K.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric de-termination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Fellman, J.H. A chemical method for the determination of acetylcholine: Its application in a study of presynaptic release and a choline acetyltransferase assay. J. Neurochem. 1969, 16, 135–143. [Google Scholar] [CrossRef]
- Kozlov, V.A. Localization and State of Tissue Transmitter Systems in Normal and Experimental Conditions. Available online: http://www.dslib.net/fiziologia/lokalizacija-i-sostojanie-tkanevyh-transmitternyh-sistem-v-norme-i-jeksperimente.html (accessed on 1 March 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org (accessed on 19 January 2022).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 19 January 2022).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S. In Statistics and Computing, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Halekoh, U.; Højsgaard, S. A Kenward-Roger Approximation and Parametric Bootstrap Methods for Tests in Linear Mixed Models—The R Package pbkrtest. J. Stat. Softw. 2014, 59, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Length, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. 2022. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 19 January 2022).
- Green, P.; MacLeod, C.J. SIMR: An R package for power analysis of generalized linear mixed models by simulation. Methods Ecol. Evol. 2016, 7, 493–498. [Google Scholar] [CrossRef]
- Sharma, C.; Kim, S.; Nam, Y.; Jung, U.J.; Kim, S.R. Mitochondrial Dysfunction as a Driver of Cognitive Impairment in Alzheimer’s disease. Int. J. Mol. Sci. 2021, 22, 4850. [Google Scholar] [CrossRef]
- Gonzalez-Burgos, E.; Gomez-Serranillos, M.P. Terpene Compounds in Nature: A Re-view of Their Potential Antioxidant Activity. Curr. Med. Chem. 2012, 19, 5319–5341. [Google Scholar] [CrossRef]
- Baccouri, B.; Rajhi, I. Potential Antioxidant Activity of Terpenes. In Terpenes and Terpenoids—Recent Advances; Perveen, S., Al-Taweel, A.M., Eds.; Intechopen: London, UK, 2021. [Google Scholar]
- Mata, A.T.; Proença, C.; Ferreira, A.R.; Serralheiro, M.L.M.; Nogueira, J.M.F.; Araújo, M.E.M. Antioxidant and anti-acetylcholinesterase activities of five plants used as Portuguese food spices. Food Chem. 2007, 103, 778–786. [Google Scholar] [CrossRef]
- Aazza, S.; Lyoussi, B.; Miguel, M.G. Antioxidant and Anti-acetylcholinesterase Activities of Some Commercial Essential Oils and Their Major Compounds. Molecules 2011, 16, 7672–7690. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.N.; Yeong, K.Y. Scopolamine, a Toxin-Induced Experimental Model, Used for Research in Alzheimer’s Disease. CNS Neurol. Disord. Drug Targets 2020, 19, 85–93. [Google Scholar] [CrossRef]
- Kaur, R.; Mehan, S.; Khanna, D.; Kalra, S. Ameliorative Treatment with Ellagic Acid in Scopolamine Induced Alzheimer’s Type Memory and Cognitive Dysfunctions in Rats. Austin J. Clin. Neurol. 2015, 2, 1053. [Google Scholar]
- Tzvetanova, E.R.; Georgieva, A.P.; Alexandrova, A.V.; Tancheva, L.P.; Lazarova, M.I.; Dragomanova, S.T.; Alova, L.G.; Stefanova, M.O.; Kalfin, R.E. Antioxidant mechanisms in neuroprotective action of lipoic acid on learning and memory of rats with experimental dementia, Bulg. Chem. Commun. 2018, 50, 52–57. [Google Scholar]
- Tancheva, L.P.; Popatanasov, A.B.; Dragomanova, S.T.; Tzvetanova, E.R.; Aleksandrova, S.M.; Alova, L.G.; Stefanova, M.O.; Kalfin, R.E. New mechanisms in preventive effect of el-lagic acid on cognition in mice with Alzheimer’s disease type dementia. Bulg. Chem. Commun. 2018, 50, 20–24. [Google Scholar]
- Lazarova, M.I.; Tsekova, D.S.; Tancheva, L.P.; Kirilov, K.T.; Uzunova, D.N.; Vezenkov, L.T.; Tsvetanova, E.R.; Alexandrova, A.V.; Georgieva, A.P.; Gavrilova, P.T.; et al. New Galantamine Derivatives with Inhibitory Effect on Acetylcholinesterase Activity. J. Alzheimer’s Dis. 2021, 83, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Lazarova, M.; Tancheva, L.; Alexandrova, A.; Tsvetanova, E.; Georgieva, A.; Stefanova, M.; Tsekova, D.; Vezenkov, L.; Kalfin, R.; Uzunova, D.; et al. Effects of New Galantamine Derivatives in a Scopolamine Model of Dementia in Mice. J. Alzheimer’s Dis. 2021, 84, 671–690. [Google Scholar] [CrossRef] [PubMed]
- Platel, A.; Porsolt, R. Habituation of exploratory activity in mice: A screening test for memory enhancing drugs. Psychopharmacology 1982, 78, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Platel, A.; Jalfre, M.; Pawelec, C.; Roux, S.; Porsolt, R. Habituation of exploratory activity in mice: Effects of combinations of piracetam and choline on memory processes. Pharm. Biochem. Behav. 1984, 21, 209–212. [Google Scholar] [CrossRef]
- Jomova, K.; Vondrakova, D.; Lawson, M.; Valko, M. Metals, oxidative stress and neurodegenerative disorders. Mol. Cell. Biochem. 2010, 345, 91–104. [Google Scholar] [CrossRef]
- Pocernich, C.B.; Lange, M.L.B.; Sultana, R.; Butterfield, D.A. Nutritional approaches to modulate oxidative stress in Alzheimer’s disease. Curr. Alzheimer′s Res. 2011, 8, 452–469. [Google Scholar] [CrossRef]
- El-Sherbiny, D.A.; Khalifa, A.E.; Attia, A.S.; Eldenshary, E.E.-D.S. Hypericum perforatum extract demonstrates antioxidant properties against elevated rat brain oxidative status induced by amnestic dose of scopolamine. Pharmacol. Biochem. Behav. 2003, 76, 525–533. [Google Scholar] [CrossRef]
- Fan, Y.; Hu, J.; Li, J.; Yang, Z.; Xin, X.; Wang, J.; Ding, J.; Geng, M. Effect of acidic oligosaccharide sugar chain on scopolamine-induced memory impairment in rats and its related mechanisms. Neurosci. Lett. 2005, 374, 222–226. [Google Scholar] [CrossRef]
- Jeong, E.J.; Lee, K.Y.; Kim, S.H.; Sung, S.H.; Kim, Y.C. Cognitive-enhancing and antioxidant activities of iridoid glycosides from Scrophularia buergeriana in scopolamine-treated mice. Eur. J. Pharmacol. 2008, 588, 78–84. [Google Scholar] [CrossRef]
- Goverdhan, P.; Sravanthi, A.; Mamatha, T. Neuroprotective Effects of Meloxicam and Selegiline in Scopolamine-Induced Cognitive Impairment and Oxidative Stress. Int. J. Alzheimer’s Dis. 2012, 2012, 974013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budzynska, B.; Boguszewska-Czubara, A.; Kruk-Slomka, M.; Skalicka-Woźniak, K.; Michalak, A.; Musik, I.; Biala, G. Effects of imperatorin on scopolamine-induced cognitive impairment and oxidative stress in mice. Psychopharmacology 2015, 232, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, C.; Qi, Y.; Fang, L.; Luo, J.; Bi, K.; Jia, Y. Timosaponin B-II ameliorates scopolamine-induced cognition deficits by attenuating acetylcholinesterase activity and brain oxidative damage in mice. Metab. Brain Dis. 2016, 31, 1455–1461. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Kim, B.-W.; Song, S.-Y.; Kim, J.-S.; Kim, I.-S.; Kwon, Y.-S.; Koppula, S.; Choi, D.-K. Cognitive enhancing effects of alpha asarone in amnesic mice by influencing cholinergic and antioxidant defense mechanisms. Biosci. Biotechnol. Biochem. 2012, 76, 1518–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Kim, J.; Seo, S.G.; Choi, B.R.; Han, J.S.; Lee, K.W.; Kim, J. Sulforaphane alleviates scopolamine-induced memory impairment in mice. Pharmacol. Res. 2014, 85, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Gutierres, J.M.; Carvalho, F.B.; Schetinger, M.R.; Agostinho, P.; Marisco, P.C.; Vieira, J.M.; Rosa, M.M.; Bohnert, C.; Rubin, M.; Morsch, V.M.; et al. Neuroprotective effect of anthocyanins on acetylcholinesterase activity and attenuation of scopolamine-induced amnesia in rats. Int. J. Dev. Neurosci. 2014, 33, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Heo, H.J.; Suh, Y.-M.; Kim, M.-J.; Choi, S.-J.; Mun, N.S.; Kim, H.-K.; Kim, E.; Kim, C.-J.; Cho, H.-Y.; Kim, Y.J.; et al. Daidzein Activates Choline Acetyltransferase from MC-IXCCells and Improves Drug-Induced Amnesia. Biosci. Biotechnol. Biochem. 2006, 70, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Karami, A.; Eriksdotter, M.; Kadir, A.; Almkvist, O.; Nordberg, A.; Darreh-Shori, T. csf cholinergic index, a new biomeasure of treatment effect in patients with Alzheimer’s disease. Front. Mol. Neurosci. 2019, 12, 239. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dragomanova, S.; Pavlov, S.; Marinova, D.; Hodzev, Y.; Petralia, M.C.; Fagone, P.; Nicoletti, F.; Lazarova, M.; Tzvetanova, E.; Alexandrova, A.; et al. Neuroprotective Effects of Myrtenal in an Experimental Model of Dementia Induced in Rats. Antioxidants 2022, 11, 374. https://doi.org/10.3390/antiox11020374
Dragomanova S, Pavlov S, Marinova D, Hodzev Y, Petralia MC, Fagone P, Nicoletti F, Lazarova M, Tzvetanova E, Alexandrova A, et al. Neuroprotective Effects of Myrtenal in an Experimental Model of Dementia Induced in Rats. Antioxidants. 2022; 11(2):374. https://doi.org/10.3390/antiox11020374
Chicago/Turabian StyleDragomanova, Stela, Stoyan Pavlov, Desislava Marinova, Yordan Hodzev, Maria Cristina Petralia, Paolo Fagone, Ferdinando Nicoletti, Maria Lazarova, Elina Tzvetanova, Albena Alexandrova, and et al. 2022. "Neuroprotective Effects of Myrtenal in an Experimental Model of Dementia Induced in Rats" Antioxidants 11, no. 2: 374. https://doi.org/10.3390/antiox11020374