
Protective Effect of Red Algae (Rhodophyta) Extracts on Essential Dietary Components of Heat-Treated Salmon

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
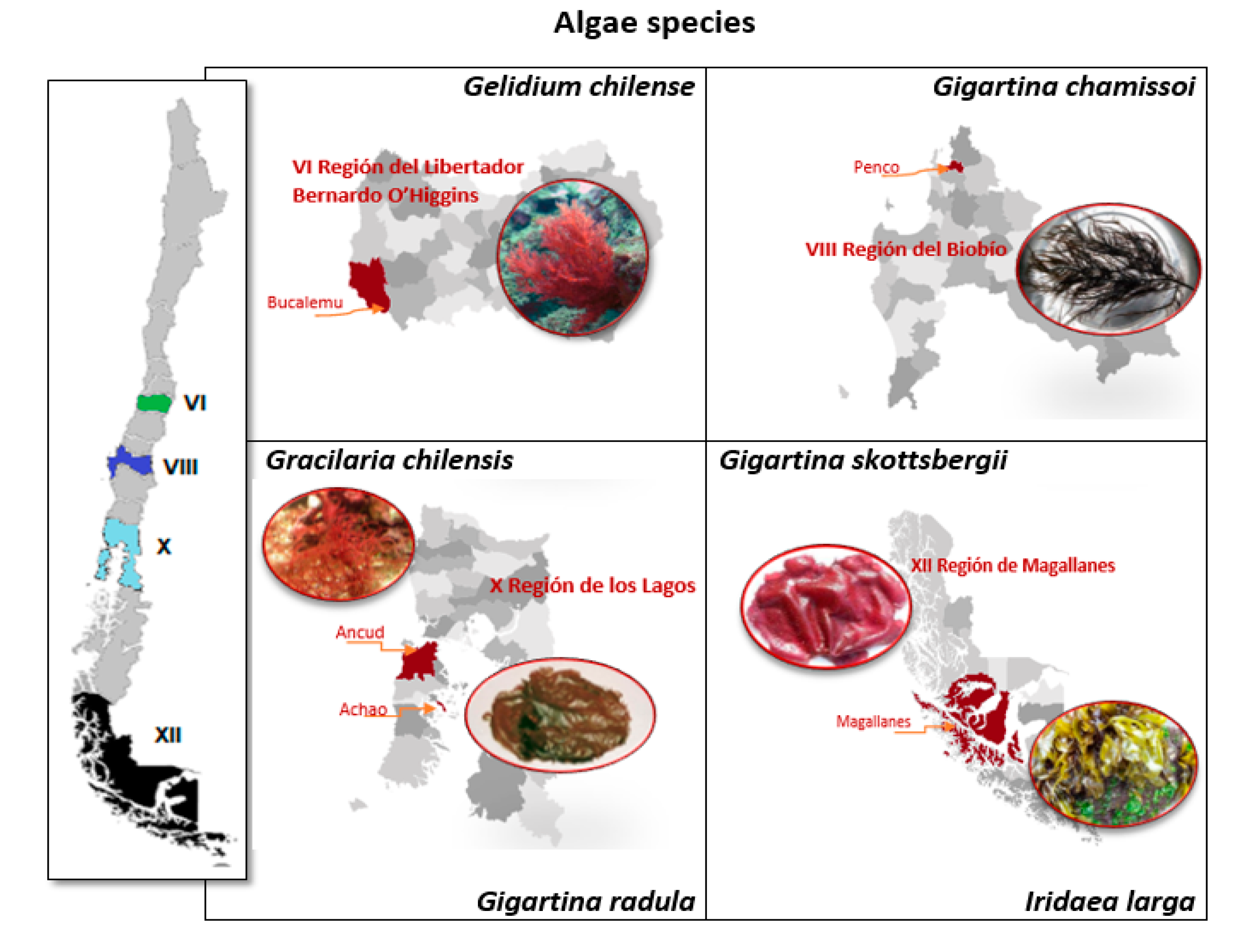
2.1. Red Algae Samples
2.2. Nutritional Content
2.3. Polyphenol Content
2.4. Determination of Flavonoids
2.5. Antiradical Activity
2.6. Antioxidant Capacity by FRAP Method
2.7. Antibacterial Activity
2.8. Antioxidant Effect of Extracts in Cooked Salmon Paste
2.9. Analysis of Polyunsaturated Fatty Acids ω-3
2.10. Tocopherols
2.11. Lipid Oxidation
2.12. Statistical Analysis
3. Results and Discussion
3.1. Nutritional Composition
3.2. Antioxidant Activity of Phenolic Extracts
3.3. Antibacterial Assays
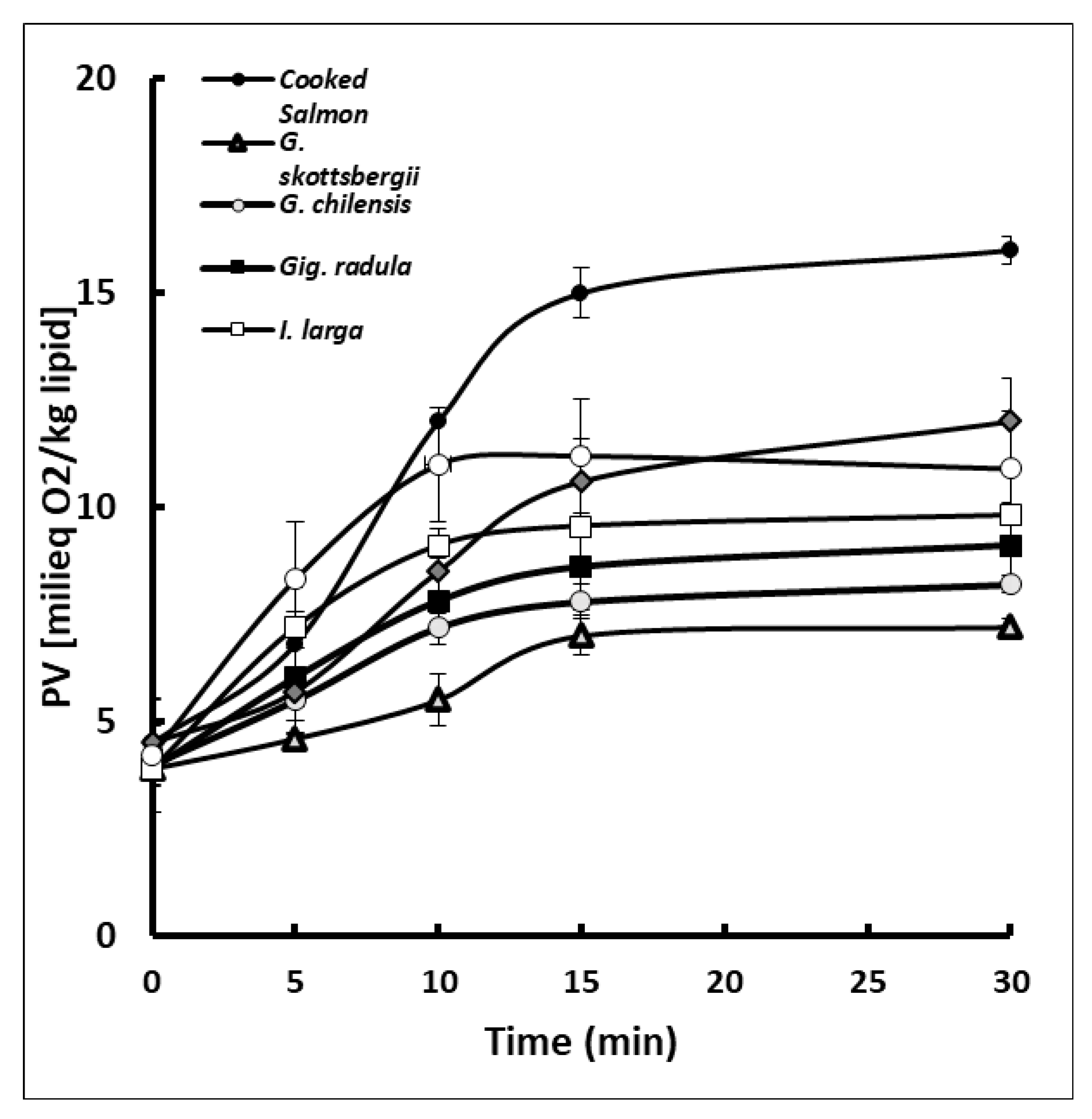
3.4. Protective Effect of Lipid Oxidation in Cooked Salmon Paste
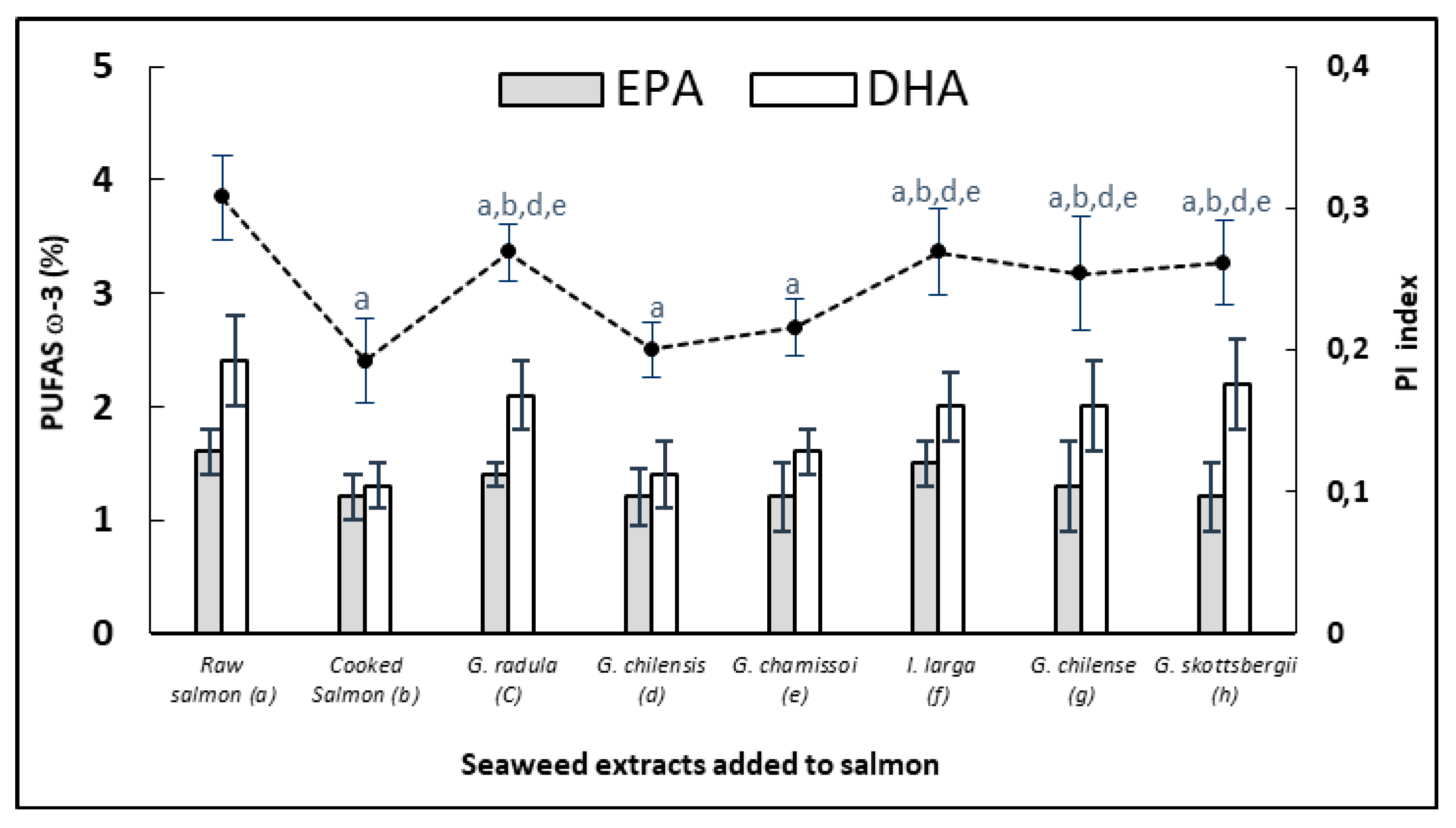
3.5. Protective Effect of EPA and DHA (PUFAS) in Cooked Salmon Paste
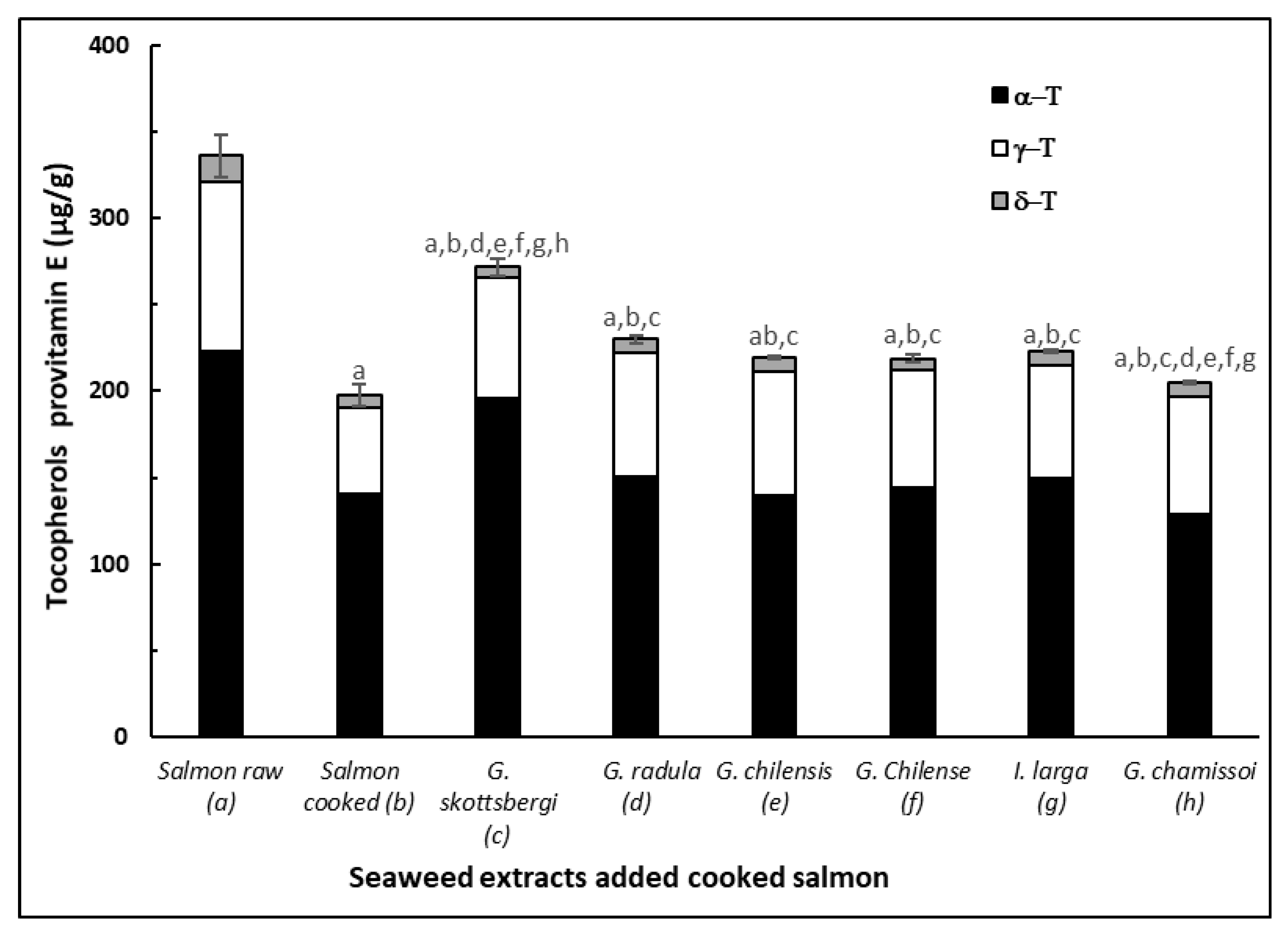
3.6. Protective Effect of Pro Vitamin E in Cooked Salmon Paste
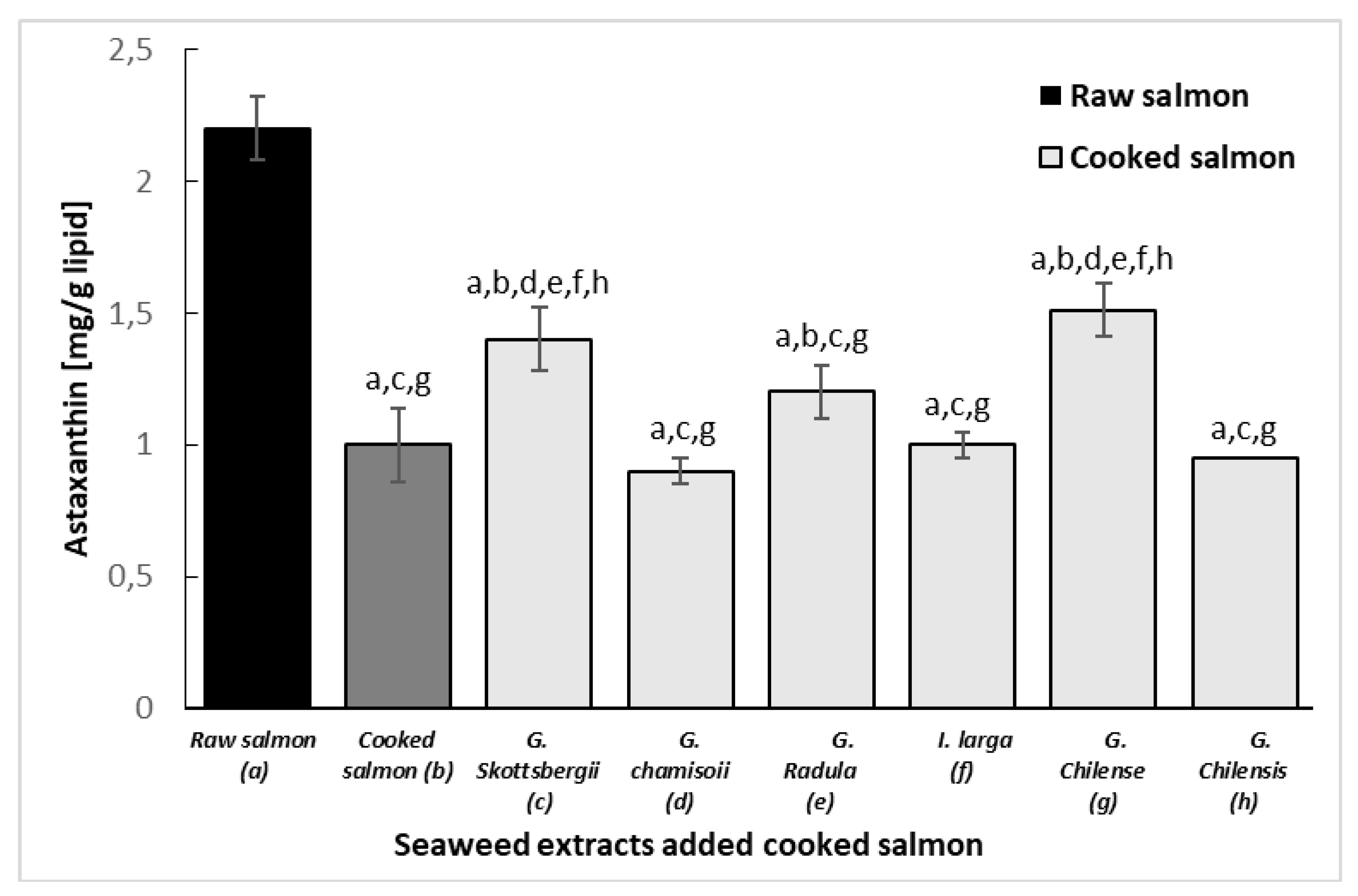
3.7. Protective Effect of Astaxanthin in Cooked Salmon Paste
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Figueroa, V.; Farfán, M.; Aguilera, J.M. Seaweeds as Novel Foods and Source of Culinary Flavors. Food Rev. Int. 2021. [Google Scholar] [CrossRef]
- Ortiz, J.; Uquiche, E.; Robert, P.; Romero, N.; Quitral, V.; Llanten, C. Functional and nutritional value of the Chilean seaweeds Codium fragile, Gracilaria chilensis and Macrocystis pyrifera. Eur. J. Lipid Sci. Technol. 2009, 111, 320–327. [Google Scholar] [CrossRef] [Green Version]
- MacArtain, P.; Gill, C.I.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Rebours, C.; Marinho-Soriano, E.; Zertuche-González, J.A.; Hayashi, L.; Vásquez, J.; Kradolfer, P.; Soriano, G.; Ugarte, R.; Abreu, M.H.; Bay-Larsen, I.; et al. Seaweeds: An opportunity for wealth and sustainable livelihood for coastal communities. J Appl. Phycol. 2014, 26, 1939–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, K.W.; Hauser, J.; Nakamura, Y.; Kanaya, S. Dietary seaweeds and obesity. Food Sci. Hum. Wellness 2015, 4, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Bobadilla, F.; Rodríguez-Tirado, C.; Imarai, M.; Galotto, M.G.; Andersson, R. Soluble -1,3/1,6-glucan in seaweed from the southern hemisphere and it immunomodulatory effect. Carbohyd. Polym. 2013, 92, 241–248. [Google Scholar] [CrossRef]
- Dovale-Rosabal, G.; Rodríguez, A.; Contreras, E.; Ortiz-Viedma, J.; Muñoz, M.; Trigo, M.; Aubourg, S.P.; Espinosa, A. Concentration of EPA and DHA from Refined Salmon Oil by Optimizing the Urea–Fatty Acid Adduction Reaction Conditions Using Response Surface Methodology. Molecules 2019, 24, 1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pando, M.E.; Rodríguez, A.; Galdames, A.; Berríos, M.; Rivera, M.; Romero, N.; Valenzuela, M.A.; Ortiz, J.; Aubourg, S. Maximization of the docosahexaenoic and eicosapentaenoic acids content in concentrates obtained from a by-product of rainbow trout (Oncorhynchus mykiss) processing. Eur. Food Res. Technol. 2020, 144, 536–543. [Google Scholar] [CrossRef] [Green Version]
- Mateluna, C.; Figueroa, V.; Ortiz, J.; Aguilera, J.M. Effect of processing on the texture and microstructure of the algae Durvillaea antarctica. J. Appl. Phycology 2020, 32, 4211–4219. [Google Scholar] [CrossRef]
- Alcicek, Z. The effects of thyme (Thymus vulgaris L.) oil concentration on liquid-smoked vacuum-packed rainbow trout (Oncorhynchus mykiss Walbaum) fillets during chilled storage. Food Chem. 2011, 128, 683–688. [Google Scholar] [CrossRef]
- Trigo, M.; López, M.; Dovale, G.; Ortiz, J.; Rodríguez, A.; Aubourg, S.P. Enhancement of sensory acceptance of frozen mackerel by alga-extract glazing. Bulg. Chem. Commun. 2019, 51, 216–223. [Google Scholar]
- Miranda, J.M.; Trigo, M.; Barros-Velazquez, J.; Aubourg, S.P. Effect of icing medium containing the alga focus spiralis on the microbiological activity and lipid oxidation in chilled megrim (Lepidorhombus whiffiagonis). Food Cont. 2016, 59, 290–297. [Google Scholar] [CrossRef] [Green Version]
- El Shafay, S.M.; Ali, S.S.; El-Sheekh, M.M. Antimicrobial activity of some seaweeds species from Red sea, against multidrug resistant bacteria. Egypt. J. Aquat. Res. 2016, 42, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, J.; Vivanco, J.P.; Aubourg, S.P. Lipid and Sensory quality of canned Atlantic salmon (Salmo salar): Effect of the use of different seaweed extracts as covering liquids. European J. Lipid Sci. and Technol. 2014, 116, 596–605. [Google Scholar] [CrossRef]
- Arulkumar, A.; Satheeshkumar, K.; Paramasivam, S.; Rameshthangam, P.; Miranda, J.M. Chemical Biopreservative Effects of Red Seaweed on the Shelf Life of Black Tiger Shrimp (Penaeus monodon). Foods 2020, 9, 634. [Google Scholar] [CrossRef]
- Ortiz, J.; Lemus-Mondaca, R.; Vega-Gálvez, A.; Ah-Hen, K.; Puente-Diaz, L.; Zura-Bravo, L.; Aubourg, S.P. Influence of air-drying temperature on drying kinetics, colour, firmness and biochemical characteristics of Atlantic salmon (Salmo salar L.) fillets. Food Chem. 2013, 4, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.; Vivanco, J.P.; Quitral, V.; Larraín, M.A.; Concha, G.; Aubourg, S.P. Effect of the antioxidant profile in the diet of farmed coho salmon (Oncorhynchus kisutch) on the nutritional value retention during frozen storage. Grasas Aceites 2013, 64, 3. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis of Association of Official Analytical Chemists International, 16th ed.; AOAC: Gaithersburg, MD, USA, 1995; Volume I, II, p. 870. [Google Scholar]
- AOCS. Official Methods and Recommended Practices of the American Oil Chemists Society, 4th ed.; AOCS Press: Champaign, IL, USA, 1993; pp. 54–56. [Google Scholar]
- AOAC. Official Methods of Analysis of Association of Official Analytical Chemists International; Official Method 995.16; AOAC Press: Champaign, IL, USA, 2005. [Google Scholar]
- Yildiz, G.; Vatan, O.; Çelikler, S.; Dere, Ş. Determination of the Phenolic Compounds and Antioxidative Capacity in Red Algae Gracilaria bursa-pastoris. Int. J. Food Prop. 2011, 14, 496–502. [Google Scholar] [CrossRef] [Green Version]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.; y Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of Antioxidant Power China. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- González del Val, A.; Platas, G.; Basilio, A.; Cabello, A.; Gorrochategui, J.; Suay, I.; Vicente, F.; Portillo, E.; Jiménez del Río, M.; García Reina, G.; et al. Screening of antimicrobial activities in red, green and Brown macroalga from Gran Canaria (Canary Islands, Spain). Int. J. Microbiol. 2001, 4, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method of extraction and purification of total lipids. Can. J. Biochem Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, J.; Romero, N.; Robert, P.; Araya, J.; Lopez-Hernández, J.; Bozzo, C.; Navarrete, E.; Osorio, A.; Rios, A. Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea Antarctica. Food Chem. 2006, 99, 98–104. [Google Scholar] [CrossRef]
- Sanchez-Machado, D.; Lopez Hernandez, J.; Paseiro Lozada, P. High performance liquid chromatographic determination of alfa-tocopherol in macroalgae. J. Chromatogr. A 2002, 976, 277–284. [Google Scholar] [CrossRef]
- Jiménez-Escrig, A.; Goñi, I. Nutritional evaluation and physiological effects of edible marine macroalgae. Arch. Latinoam Nutr. 1999, 49, 114–120. [Google Scholar]
- Schmidt-Hebbel, H.; Pennacchiotti, I.; Masson, L.; Mella, M.A. Tabla de Composición Química de los Alimentos, 8th ed.; Universidad de Chile: Santiago, Chile, 1992. [Google Scholar]
- Chandini, S.K.; Ganesan, P.; Suresh, P.V.; Bhaskar, N. Seaweeds as a source of nutritionally beneficial compounds—A review. J Food Sci Tech. 2008, 45, 1–13. [Google Scholar]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [Green Version]
- Rohani-Ghadikolaei, K.; Abdulalian, E.; Ng, W. Evaluation of the proximate, fatty acid and mineral composition of representative green, brown and red seaweeds from the Persian Gulf of Iran as potential food and feed resources. J Food Sci Tech. 2012, 49, 774–780. [Google Scholar] [CrossRef] [Green Version]
- Dhargalkar, V.K.; Neelam, P. Seaweed: Promising plant of the millennium. Sci. Cult. 2005, 71, 60–66. [Google Scholar]
- Lechat, H.; Amat, M.; Mazoyer, J.; Buleón, A.; Lahaye, M. Structure and distribution of glucomannan and sulfated glucan in the cell walls of the red alga Kappaphycus alvarezii (Gigartinales, Rhodophyta). J. Phycol. 2001, 36, 891–902. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, X.; Li, J.; Jiang, H.; Shan, X.; Wang, Y.; Ma, W.; Hao, J.; Yu, G. A β-glucan from Durvillaea Antarctica has immunomodulatory effects on RAW264.7 macrophages via Toll-like receptor 4. Carbohyd. Polym. 2018, 191, 255–265. [Google Scholar] [CrossRef]
- Sathya, R.; Kanaga, N.; Sankar, P.; Jeeva, S. Antioxidant properties of phlorotannins from brown seaweed Cystoseira trinodis (Forsskål) C. Agardh. Arab. J. Chem. 2017, 10, S2608–S2614. [Google Scholar] [CrossRef] [Green Version]
- Devi, G.K.; Manivannan, K.; Thirumaran, G.; Rajathi, G.; Anantharaman, P. In vitro antioxidant activities of selected seaweeds from Southeast coast of India. Asian Pac. J. Trop. Med. 2011, 4, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Kajal, C.; Deepu, J.; Nammunayathuputhenkotta, K.P. Antioxidant activities and phenolic contents of three red seaweeds (Division: Rhodophyta) harvested from the Gulf of Mannar of Peninsular India. J Food Sci Tech. 2015, 52, 1924–1935. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, K.; Krishnankartha, N.; Praveen, K.; KizekadathVijayan, K.; Syda Rao, G. Evaluation of phenolic contents and antioxidant activities of brown seaweeds belonging to Turbinaria spp. (Phaeophyta, Sargassaceae) collected from Gulf of Mannar. Asian Pac. J. Trop. Biomed. 2013, 3, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Jónsdóttir, R.; Olafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Yoshie-Stark, Y.; Hsieh, Y.P.; Suzuki, T. Distribution of flavonoids and related compounds from seaweeds in Japan. J. Tokyo Univ. Fish. 2003, 89, 1–6. [Google Scholar]
- Moubayed, N.M.S.; Al Houri, H.J.M.; Khulaifi, A.; Al Farraj, D.A. Antimicrobial, antioxidant properties and chemical composition of seaweeds collected from Saudi Arabia (Red Sea and Arabian Gulf). Saudi J. Biol. Sci. 2017, 24, 162–169. [Google Scholar] [CrossRef] [Green Version]
- De Alencar, D.B.; De Carvalho, F.C.T.; Rebouças, R.H.; Dos Santos, D.R.; Dos Santos Pires-Cavalcante, K.M.; De Lima, R.L.; Baracho, B.M.; Bezerra, R.M.; Viana, F.A.; Dos Fernandes Vieira, R.H.S.; et al. Bioactive extracts of red seaweeds Pterocladiella capillacea and Osmundaria obtusiloba (Floridophyceae: Rhodophyta) with antioxidant and bacterial agglutination potential. Asian Pac. J. Trop. Med. 2016, 9, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Oucif, H.; Miranda, J.M.; Trigo, M.; Iglesias, R.; Toro, J.; Al-Mehidi, S.; Barros-Velázquez, J.; Aubourg, S.P. Antimicrobial and antioxidant effects of alga Cystoseira compressa extract during the chilled storage of horse mackerel (Trachurus trachurus). In Proceedings of the 47th West European Fish Technologists Association Conference, Dublin, Ireland, 9–12 October 2017. [Google Scholar]
- Miranda, J.M.; Ortiz, J.; Barros-Velázquez, J.; Aubourg, S.P. Quality Enhancement of Chilled Fish by Including Alga Bifurcaria bifurcata Extract in the Icing Medium. Food Bioprocess Technol. 2016, 9, 387–395. [Google Scholar] [CrossRef] [Green Version]
- Medina, I.; Sacchi, R.; Aubourg, S.P. A 13C-NMR study of lipid alterations during fish canning: Effect of filling medium. J. Sci. Food Agric. 1995, 69, 445–450. [Google Scholar] [CrossRef]
- Rodríguez, A.; Cruz, J.M.; Paseiro-Losada, P.; Aubourg, S.P. Effect of a polyphenol-vacuum packaging on lipid deterioration during an 18-month frozen storage of coho salmon (Oncorhynchus kisutch). Food Bioprocess Technol. 2012, 5, 2602–2611. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Viedma, J.; Romero, N.; Puente, L.; Burgos, K.; Toro, M.; Ramirez, L.; Rodriguez, A.; J Barros-Velazquez, J.; Aubourg, S.P. Antioxidant and antimicrobial effects of stevia (Stevia rebaudiana Bert.) extracts during preservation of refrigerated salmon paste. Eur. J. Lipid Sci. Technol. 2017, 119, 1600467. [Google Scholar] [CrossRef]
- Iglesias, J.; Pazos, M.; Andersen, M.L.; Skibsted, L.H.; Medina, I. Caffeic Acid as Antioxidant in Fish Muscle: Mechanism of Synergism with Endogenous Ascorbic Acid and α-Tocopherol. Food Chem. 2006, 99, 98–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iglesias-Neira, J.; Pazos, M.; Maestre, R.; Torres, J.L.; Medina, I. Galloylated Polyphenols as Inhibitors of Hemoglobin-Catalyzed Lipid Oxidation in Fish Muscle. J. Agric. Food Chem. 2011, 59, 5684–5691. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algae | Ash | Lipids | Proteins | Carbohydrates | Fiber | β-Glucans |
|---|---|---|---|---|---|---|
| I. larga (a) | 21.29 ± 1.69c | 0.73 ± 0.19b | 1.23 ± 1.45b | 66.74 ± 4.49b | 59.76 ± 2.09c | 7.0 ± 0.4 b,d,f |
| G. chilensis (b) | 12.28 ± 2.45b | 1.36 ± 0.25c | 19.94 ± 1.13d | 66.42 ± 0.16b | 59.99 ± 5.24c | 4.9 ± 0.1 a,c,d,f |
| G. chilense (c) | 8.77 ± 1.44a | 1.40 ± 0.26c | 20.26 ± 1.48d | 69.57 ± 4.30a | 55.45 ± 2.10b | 6.0 ± 0.6 b,d,f |
| G. chamissoi(d) | 13.66 ± 1.21d | 3.73 ± 0.05d | 14.08 ± 1.50c | 68.53 ± 3.45a | 55.16 ± 1.32b | 3.2 ± 0.8 a,b,ce,f |
| G. rádula (e) | 19.58 ± 1.52c | 0.95 ± 0.14b | 11.18 ± 1.18b | 68.29 ± 2.20a | 48.91 ± 1.71a | 6.4 ± 0.2 b,d,f |
| G.skottsbergii (f) | 25.72 ± 1.24b | 0.20 ± 0.03a | 7.57 ± 0.91a | 66.51 ± 5.32d | 59.18 ± 5.11c | 5.6 ± 0.3 a,b,d,e |
| Algae | TP | TF | DPPH | FRAP |
|---|---|---|---|---|
| mgGAE/g dw | mgCE/g dw | (%) | g Fe2+/100 g | |
| I. larga (a) | 6.9 ± 1.2 b,c,d,f | 5.8 ± 1.2 b,c,d,f | 26.6 ± 3.5 b,c,e,f | 0.36 ± 0.01 b,c,d |
| G. chilensis (b) | 2.6 ± 0.6 a,c,e,f | 2.2 ± 1.2 a,c,e,f | 34.2 ± 6.0 a,c,e,d,e | 0.57 ± 0.11 a,c,d,e,f |
| G. chilense (c) | 9.9 ± 1.3 a,b,c,d,e | 8.4 ± 1.2 a,b,d,e,f | 51.2 ± 9.1 a,b,c,d,e,d | 0.47 ± 0.04 a,b,d,e,f |
| G. chamissoi (d) | 3.4 ± 0.4 a,b,d,e,f | 3.1 ± 1.2 a,b,c,d,e | 21.6 ± 4.1 b,c,d,e | 0.62 ± 0.08 a,c,d,f |
| G. rádula (e) | 6.1 ± 0.9 a,b,c,d | 5.1 ± 1.2 b,c,d,e | 72.2 ± 10.8 a,b,c,d | 0.31 ± 0.01 b,c,d |
| G. skottsbergii (f) | 11.3 ± 2.1 a,b,c,d,e | 9.6 ± 1.2 a,b,c,d,e | 66.9 ± 16.4 a,b,c,d | 0.34 ± 0.01 b,c,d |
| Bacterial Strain | ||||||
|---|---|---|---|---|---|---|
| Algae Extract | S. enteritidis | E. coli | B. cereus | P. aeruginosa | St. aureus | P. mirabilis |
| Iridaea larga | + | + | + | − | − | − |
| Gracilaria chilensis | + | + | + | + | + | + |
| Gelidium chilense | + | + | + | + | + | + |
| Gigartina chamissoi | + | + | + | + | + | + |
| Gigartina radula | + | + | + | + | + | + |
| Gigartina skottsbergii | + | + | + | + | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortiz-Viedma, J.; Aguilera, J.M.; Flores, M.; Lemus-Mondaca, R.; Larrazabal, M.J.; Miranda, J.M.; Aubourg, S.P. Protective Effect of Red Algae (Rhodophyta) Extracts on Essential Dietary Components of Heat-Treated Salmon. Antioxidants 2021, 10, 1108. https://doi.org/10.3390/antiox10071108
Ortiz-Viedma J, Aguilera JM, Flores M, Lemus-Mondaca R, Larrazabal MJ, Miranda JM, Aubourg SP. Protective Effect of Red Algae (Rhodophyta) Extracts on Essential Dietary Components of Heat-Treated Salmon. Antioxidants. 2021; 10(7):1108. https://doi.org/10.3390/antiox10071108
Chicago/Turabian StyleOrtiz-Viedma, Jaime, José M. Aguilera, Marcos Flores, Roberto Lemus-Mondaca, María José Larrazabal, José M. Miranda, and Santiago P. Aubourg. 2021. "Protective Effect of Red Algae (Rhodophyta) Extracts on Essential Dietary Components of Heat-Treated Salmon" Antioxidants 10, no. 7: 1108. https://doi.org/10.3390/antiox10071108