Melittin from Apis florea Venom as a Promising Therapeutic Agent for Skin Cancer Treatment

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Protein Analysis of Crude Bee Venom Extracted from A. florea
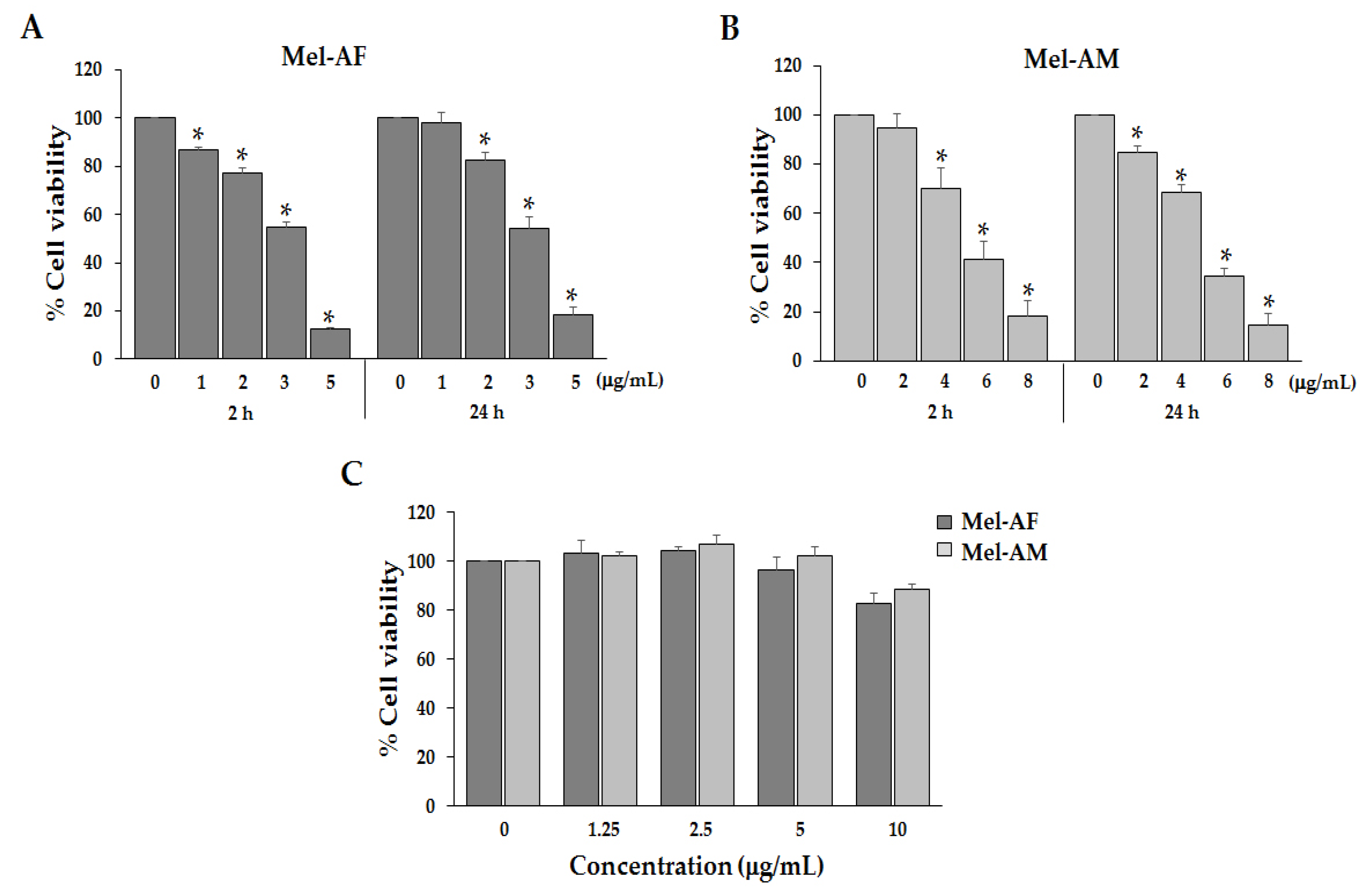
2.2. Anti-Proliferative Effect of Mel-AF on A375 Cells
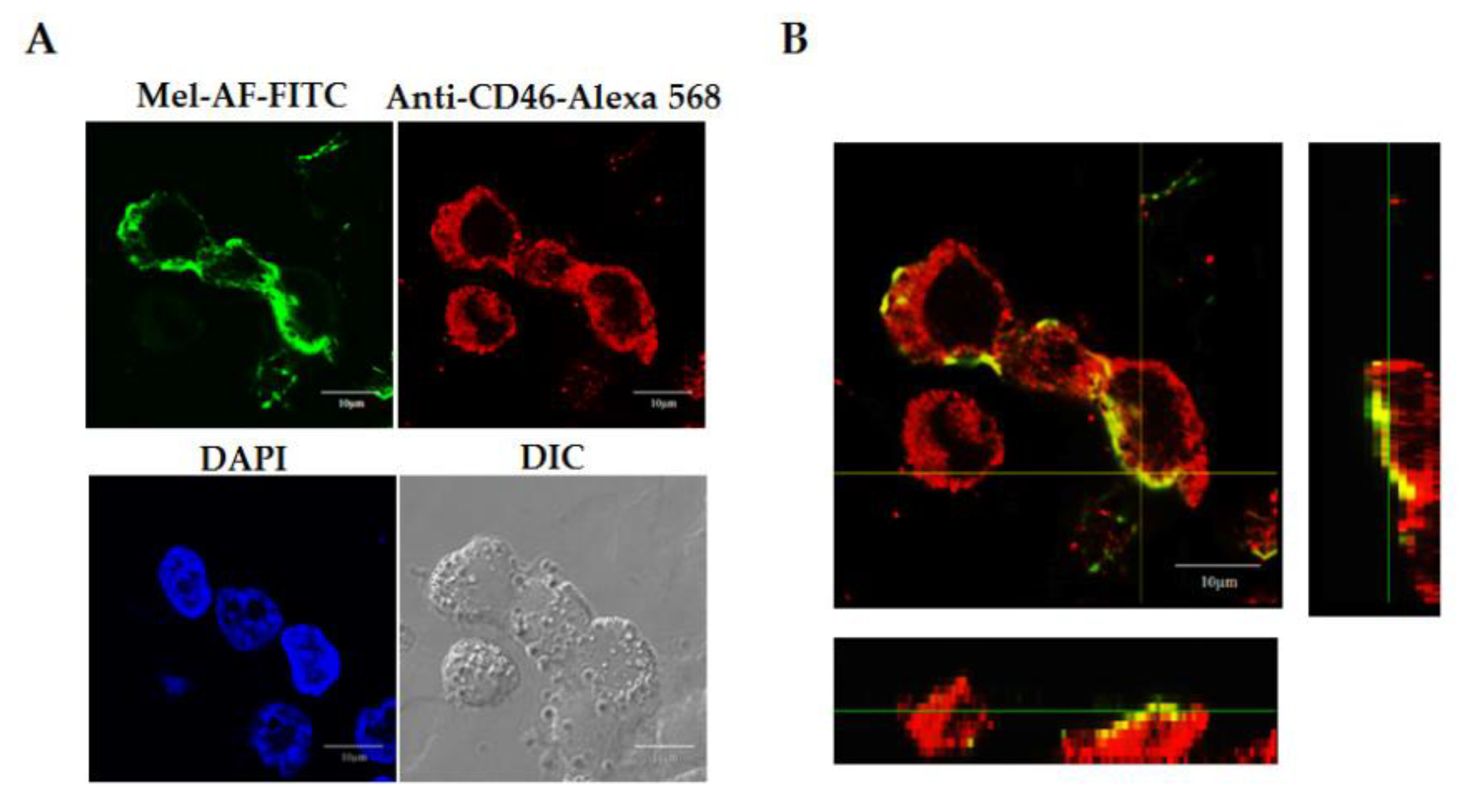
2.3. Localization of Melittin Peptide
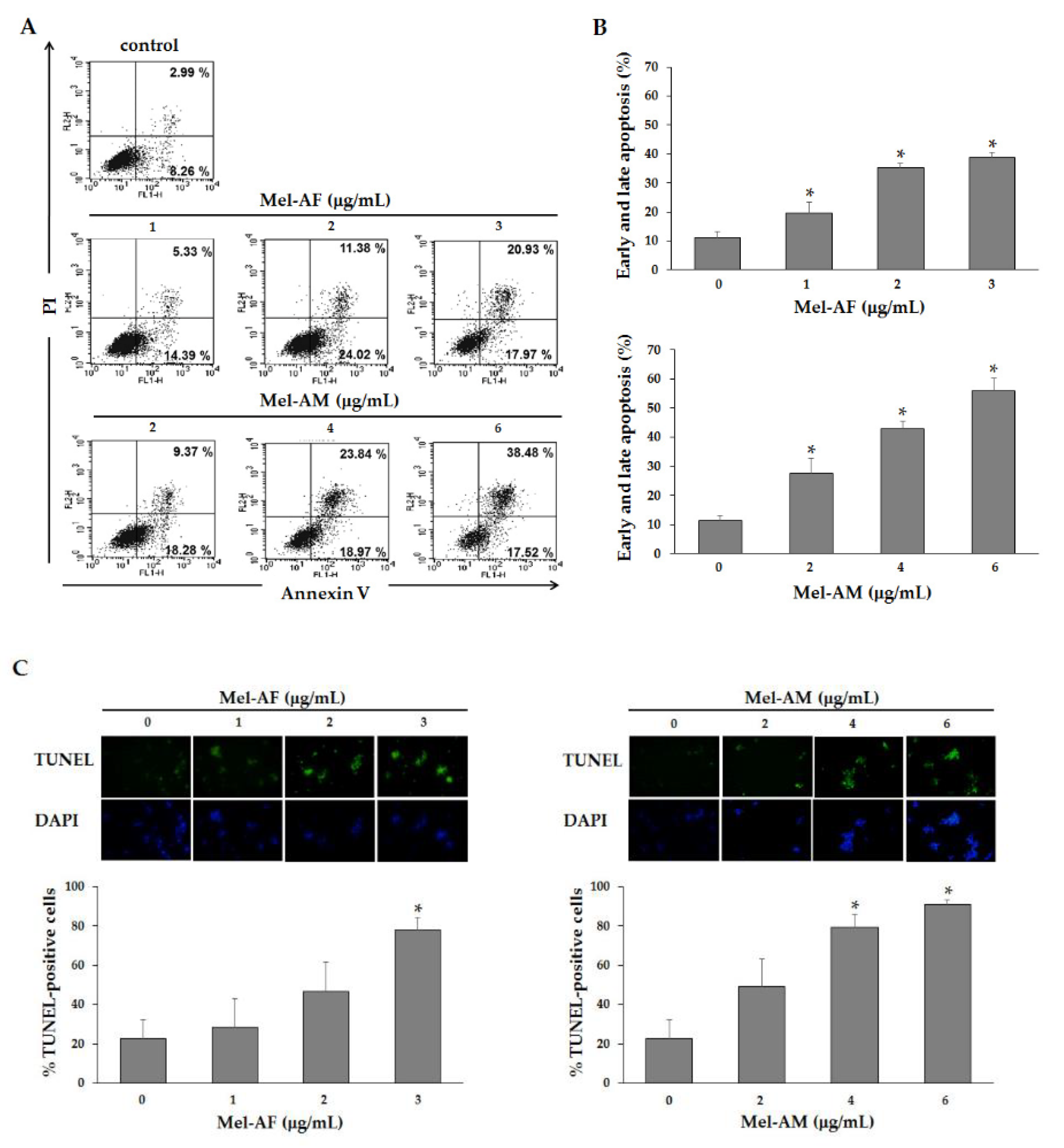
2.4. Apoptosis Induction by Mel-AF
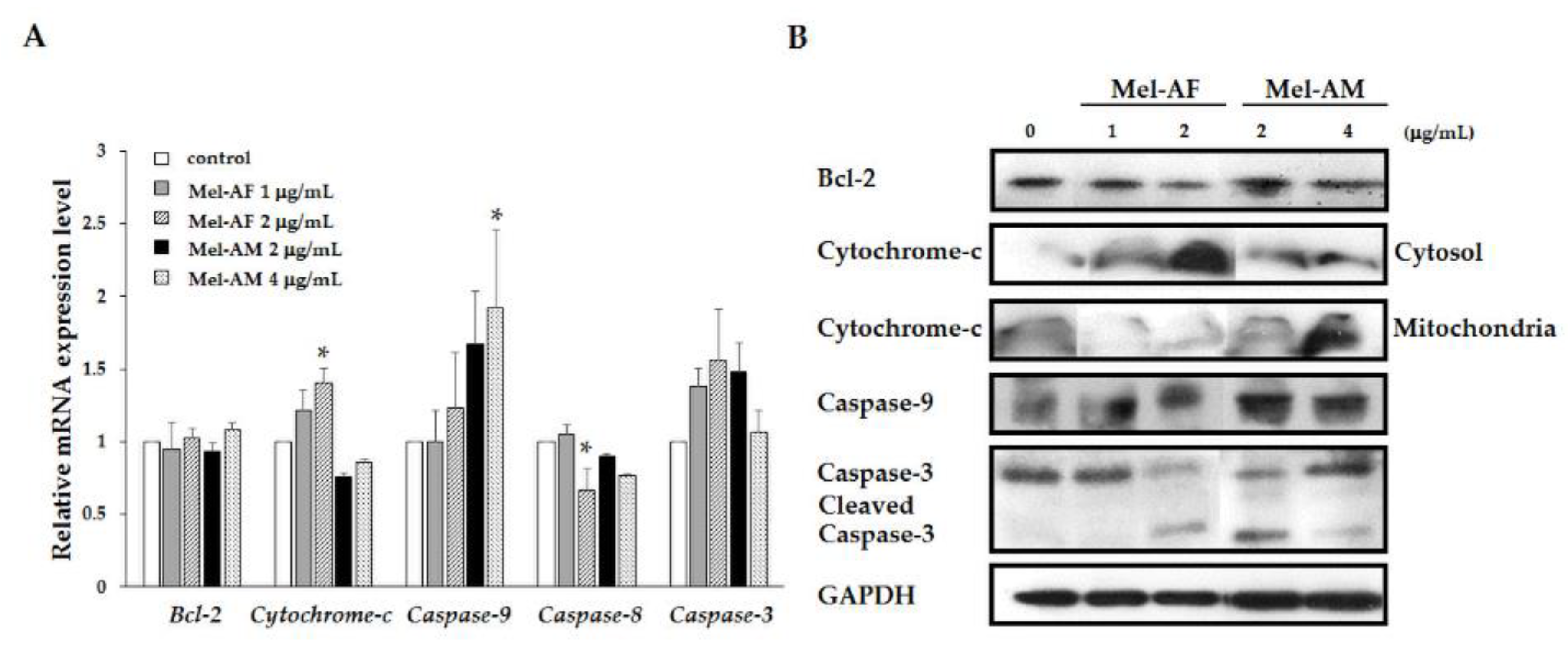
2.5. Activation of Apoptosis-Related Genes and Proteins
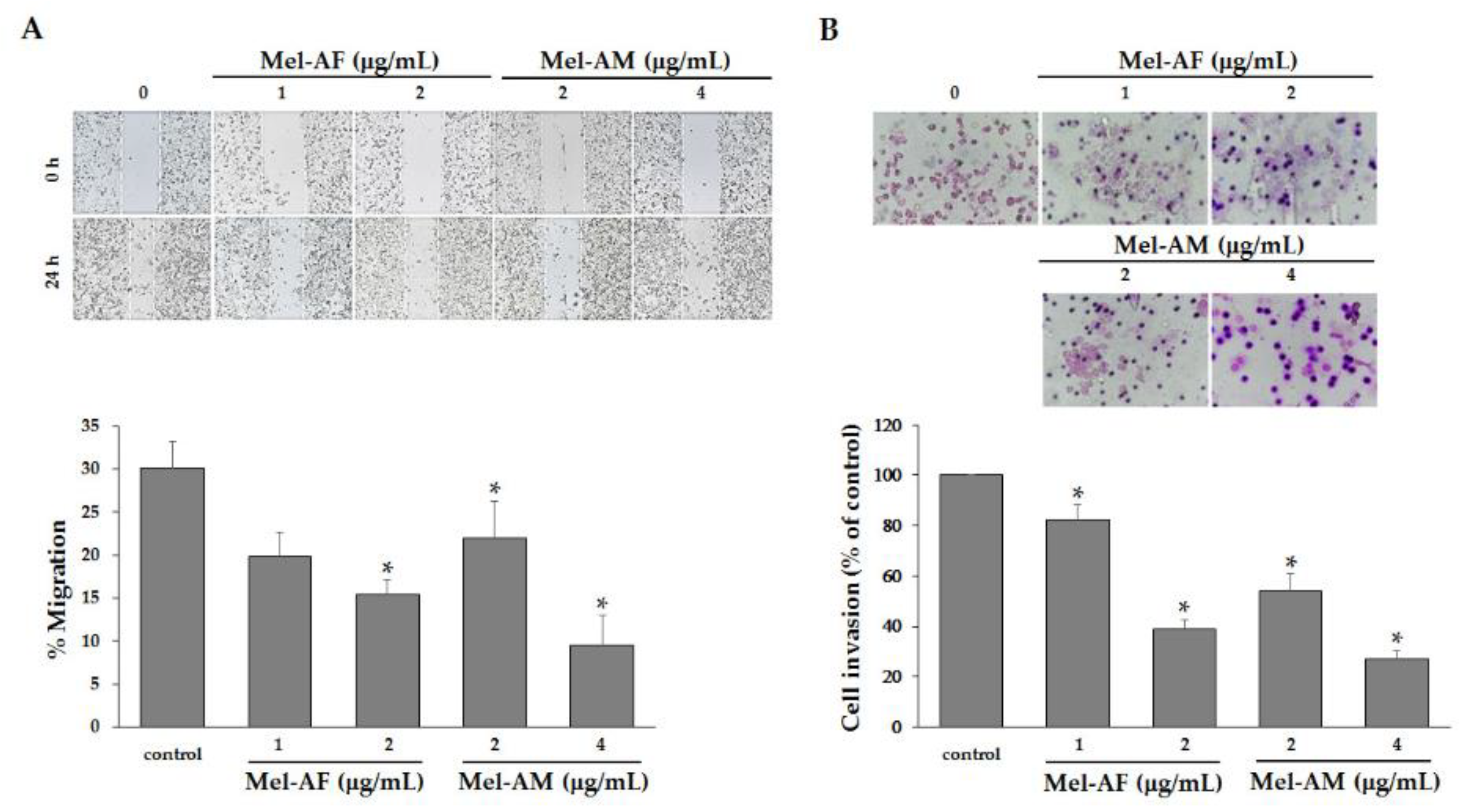
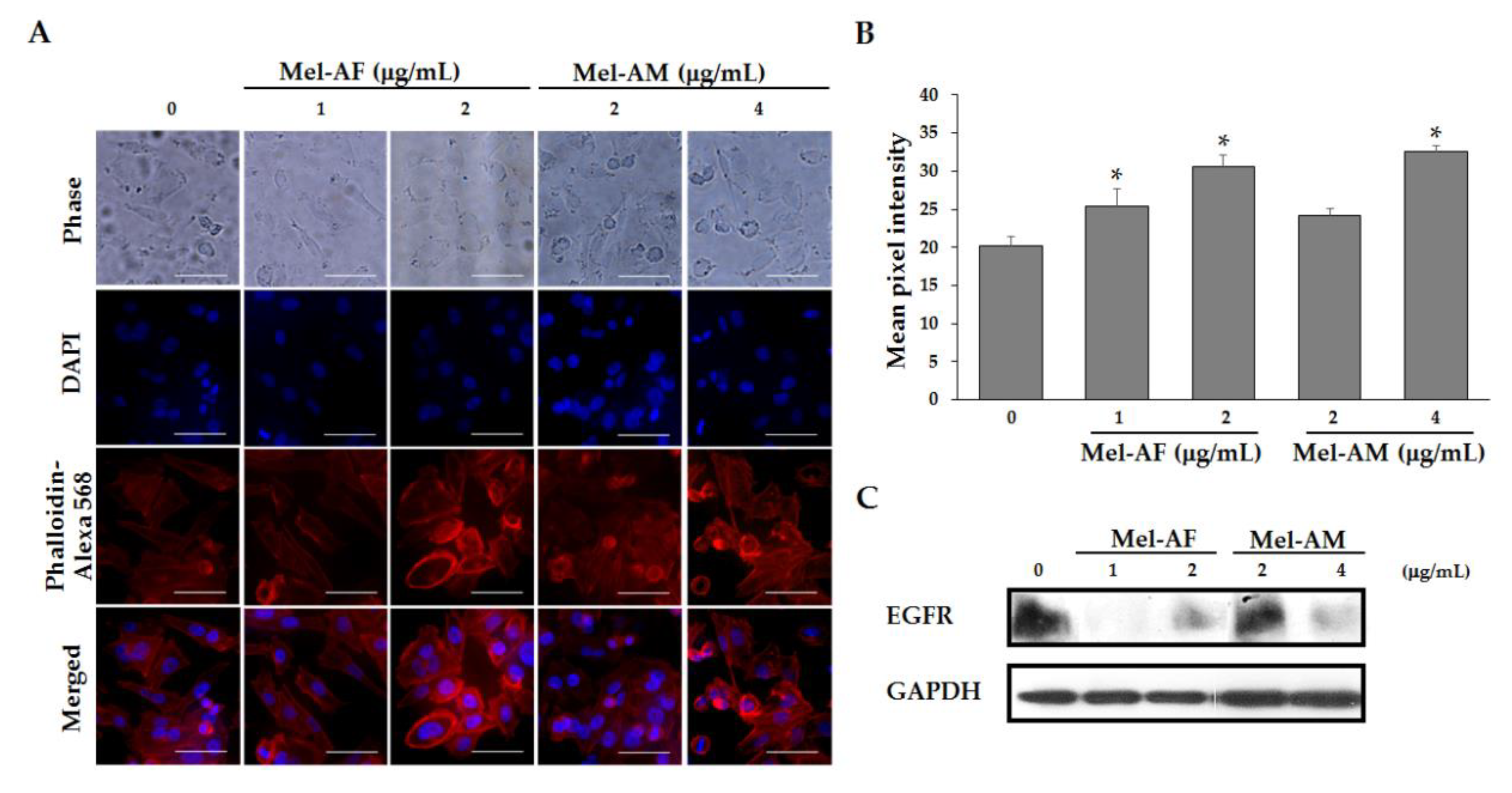
2.6. Mel-AF Suppressed Migration and Invasion Abilities of A375 Cells by Interfering with F-actin Reorganization and Diminishing Epidermal Growth Factor Receptor (EGFR) Activity
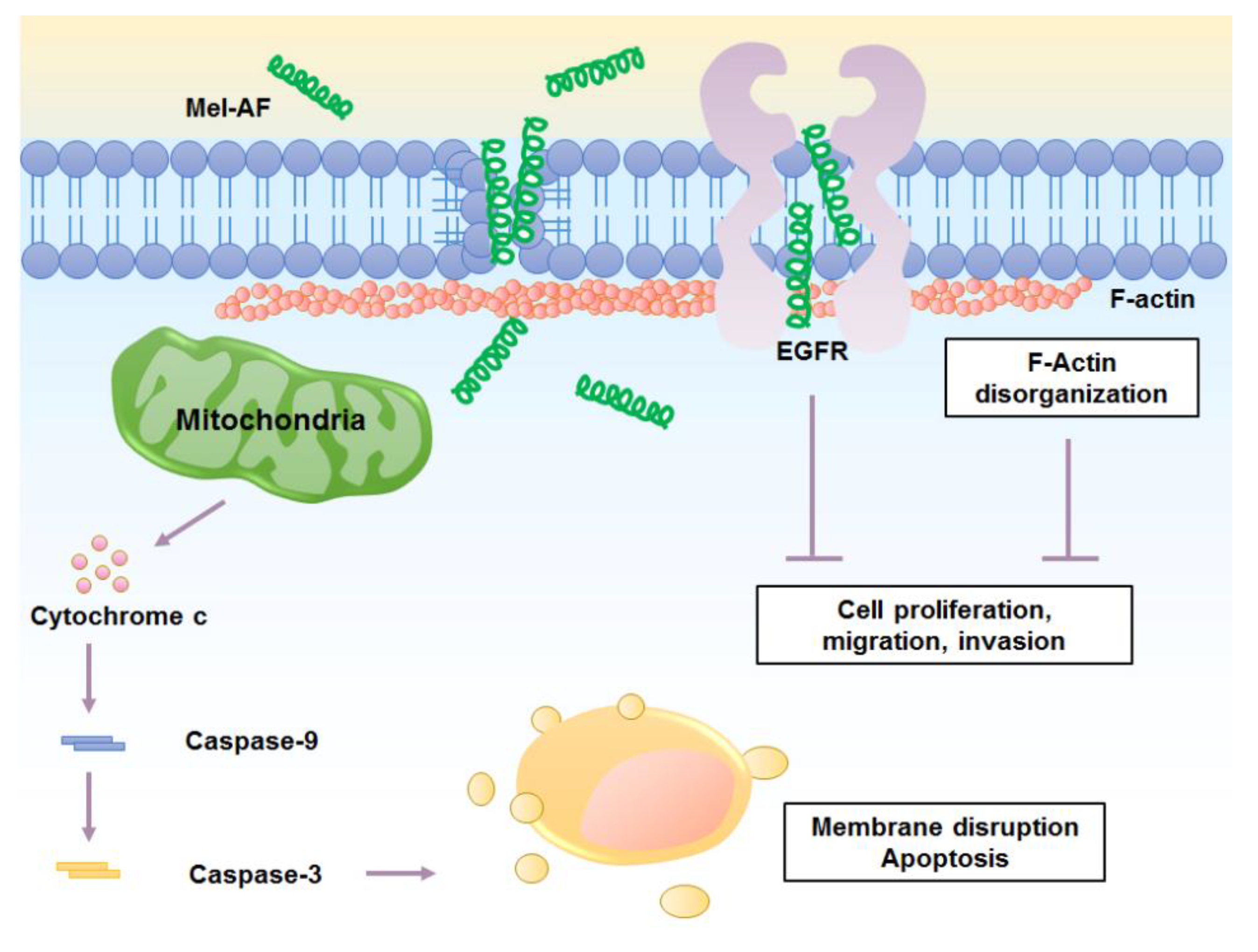
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bee Venom and Synthetic Melittin Peptide
5.2. Cell Culture
5.3. Cell Viability Assay
5.4. Peptide Localization Analysis
5.5. Phosphatidylserine Externalization Analysis
5.6. DNA Fragmentation Analysis
5.7. Apoptosis-Related Genes Analysis
5.8. Determination of Apoptosis-Related Protein Expression
5.9. Cell Migration Assay
5.10. Transwell Invasion Assay
5.11. F-actin Staining
5.12. Expression of Epidermal Growth Factor Receptor (EGFR)
5.13. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pejkova, S.; Dzokic, G.; Tudzarova-Gjorgova, S.; Panov, S. Molecular biology and genetic mechanisms in the progression of the malignant skin melanoma. Prilozi 2016, 37, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.J.; Ao, H.F.; Huang, W.T.; Chen, F.; Sun, X.C.; Wang, J.J.; Lui, Z.F.; Han, W.W.; Fry, A.N.; Wang, D.H. Impact of different surgical and postoperative adjuvant treatment modalities on survival of sinonasal malignant melanoma. BMC Cancer 2014, 14, 608. [Google Scholar] [CrossRef] [Green Version]
- Gray-Schopfer, V.; Wellbrock, C.; Marais, R. Melanoma biology and new targeted therapy. Nature 2007, 445, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.P.; Li, S.; Chen, Y.M.; Li, H.B. Natural polyphenols for prevention and treatment of cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Balunas, M.J.; Kinghorn, A.D. Drug discovery from medicinal plants. Life Sci. 2005, 78, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.; Watari, H.; AbuAlmaaty, A.; Ohba, Y.; Sakuragi, N. Apoptosis and molecular targeting therapy in cancer. Biomed Res. Int. 2014, 150845. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.; Bae, H. Anti-inflammatory applications of melittin, a major component of bee venom: Detailed mechanism of action and adverse effects. Molecules 2016, 21, 616. [Google Scholar] [CrossRef]
- Chen, J.; Guan, S.M.; Sun, W.; Fu, H. Melittin, the major pain-producing substance of bee venom. Neurosci. Bull. 2016, 32, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Aufschnaiter, A.; Kohler, V.; Khalifa, S.; El-Wahed, A.A.; Du, M.; El-Seedi, H.; Büttner, S. Apitoxin and its components against cancer, neurodegeneration and rheumatoid arthritis: Limitations and possibilities. Toxins 2020, 12, 66. [Google Scholar] [CrossRef] [Green Version]
- Son, D.J.; Lee, J.W.; Lee, Y.H.; Song, H.S.; Lee, C.K.; Hong, J.T. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol. Ther. 2007, 115, 246–270. [Google Scholar] [CrossRef] [PubMed]
- Orsolic, N. Bee venom in cancer therapy. Cancer Metastasis Rev. 2012, 31, 173–194. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, F.B.; Ng, F.S.; Nguyen, T.H.; Lim, L.H. Honeybee venom secretory phospholipase A2 induces leukotriene production but not histamine release from human basophils. Clin. Exp. Immunol. 2008, 151, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Somwongin, S.; Chantawannakul, P.; Chaiyana, W. Antioxidant activity and irritation property of venoms from Apis species. Toxicon 2018, 145, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Gajski, G.; Garaj-Vrhovac, V. Melittin: A lytic peptide with anticancer properties. Environ. Toxicol. Pharmacol. 2013, 36, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Rady, I.; Siddiqui, I.A.; Rady, M.; Mukhtar, H. Melittin, a major peptide component of bee venom, and its conjugates in cancer therapy. Cancer Lett. 2017, 402, 16–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Therrien, A.; Fournier, A.; Lafleur, M. Role of the cationic C-terminal segment of melittin on membrane fragmentation. J. Phys. Chem. B 2016, 120, 3993–4002. [Google Scholar] [CrossRef] [Green Version]
- Jo, M.; Park, M.H.; Kollipara, P.S.; An, B.J.; Song, H.S.; Han, S.B.; Kim, J.H.; Song, M.J.; Hong, J.T. Anti-cancer effect of bee venom toxin and melittin in ovarian cancer cells through induction of death receptors and inhibition of JAK2/STAT3 pathway. Toxicol. Appl. Pharmacol. 2012, 258, 72–81. [Google Scholar] [CrossRef]
- Alonezi, S.; Tusiimire, J.; Wallace, J.; Dufton, M.J.; Parkinson, J.A.; Young, L.C.; Clements, C.J.; Park, J.K.; Jeon, J.W.; Ferro, V.A.; et al. Metabolomic Profiling of the Synergistic Effects of Melittin in Combination with Cisplatin on Ovarian Cancer Cells. Metabolites 2017, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Raghuraman, H.; Chattopadhyay, A. Melittin: A membrane-active peptide with diverse functions. Biosci. Rep. 2007, 27, 189–223. [Google Scholar] [CrossRef]
- Kreil, G. Structure of melittin isolated from two species of honey bees. FEBS Lett. 1973, 33, 241–244. [Google Scholar] [CrossRef] [Green Version]
- Li, J.H.; Zhang, C.X.; Tang, Z.H. Expression of melittin gene in the venom gland of the Chinese honeybee, Apis cerana cerana. Apidologie 2005, 36, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, J.O. Toxinology of venoms from the honeybee genus Apis. Toxicon 1995, 33, 917–927. [Google Scholar] [CrossRef]
- Sobral, F.; Sampaio, A.; Falcao, S.; Queiroz, M.J.; Calhelha, R.C.; Vilas-Boas, M.; Ferreira, I.C. Chemical characterization, antioxidant, anti-inflammatory and cytotoxic properties of bee venom collected in Northeast Portugal. Food Chem. Toxicol. 2016, 94, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Tu, W.C.; Wu, C.C.; Hsieh, H.L.; Chen, C.Y.; Hsu, S.L. Honeybee venom induces calcium-dependent but caspase-independent apoptotic cell death in human melanoma A2058 cells. Toxicon 2008, 52, 318–329. [Google Scholar] [CrossRef]
- Ip, S.W.; Wei, H.C.; Lin, J.P.; Kuo, H.M.; Liu, K.C.; Hsu, S.C.; Yang, J.S.; Mei, D.; Chiu, T.H.; Han, S.M.; et al. Bee venom induced cell cycle arrest and apoptosis in human cervical epidermoid carcinoma Ca Ski cells. Anticancer Res. 2008, 28, 833–842. [Google Scholar]
- van den Bogaart, G.; Guzman, J.V.; Mika, J.T.; Poolman, B. On the mechanism of pore formation by melittin. J. Biol. Chem. 2008, 283, 33854–33857. [Google Scholar] [CrossRef] [Green Version]
- Sani, M.A.; Separovic, F. How membrane-active peptides get into lipid membranes. Acc. Chem. Res. 2016, 49, 1130–1138. [Google Scholar] [CrossRef]
- Gajski, G.; Domijan, A.M.; Zegura, B.; Stern, A.; Geric, M.; Novak Jovanovic, I.; Vrhovac, I.; Madunic, J.; Breljak, D.; Filipic, M.; et al. Melittin induced cytogenetic damage, oxidative stress and changes in gene expression in human peripheral blood lymphocytes. Toxicon 2016, 110, 56–67. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Kong, G.M.; Tao, W.H.; Diao, Y.L.; Fang, P.H.; Wang, J.J.; Bo, P.; Qian, F. Melittin induces human gastric cancer cell apoptosis via activation of mitochondrial pathway. World J. Gastroenterol. 2016, 22, 3186–3195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Yang, S.; Sun, H.; Sun, T.; Ji, F.; Wang, Y.; Xu, L.; Zhou, D. Keap1 inhibits metastatic properties of NSCLC cells by stabilizing architectures of F-actin and focal adhesions. Mol. Cancer Res. 2018, 16, 508–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, A.L.; Schrecengost, R.S.; Guerrero, M.S.; Thomas, K.S.; Bouton, A.H. Breast cancer antiestrogen resistance 3 (BCAR3) promotes cell motility by regulating actin cytoskeletal and adhesion remodeling in invasive breast cancer cells. PLoS ONE 2013, 8, e65678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrecengost, R.S.; Riggins, R.B.; Thomas, K.S.; Guerrero, M.S.; Bouton, A.H. Breast cancer antiestrogen resistance-3 expression regulates breast cancer cell migration through promotion of p130Cas membrane localization and membrane ruffling. Cancer Res. 2007, 67, 6174–6182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardama, G.A.; Comin, M.J.; Hornos, L.; Gonzalez, N.; Defelipe, L.; Turjanski, A.G.; Alonso, D.F.; Gomez, D.E.; Menna, P.L. Preclinical development of novel Rac1-GEF signaling inhibitors using a rational design approach in highly aggressive breast cancer cell lines. Anticancer Agents Med. Chem. 2014, 14, 840–851. [Google Scholar] [CrossRef] [Green Version]
- Franklin-Tong, V.E.; Gourlay, C.W. A role for actin in regulating apoptosis/programmed cell death: Evidence spanning yeast, plants and animals. Biochem. J. 2008, 413, 389–404. [Google Scholar] [CrossRef] [Green Version]
- May, M.; Wang, T.; Muller, M.; Genth, H. Difference in F-actin depolymerization induced by toxin B from the Clostridium difficile strain VPI 10463 and toxin B from the variant Clostridium difficile serotype F strain 1470. Toxins 2013, 5, 106–119. [Google Scholar] [CrossRef]
- Audoly, G.; Popoff, M.R.; Gluschankof, P. Involvement of a small GTP binding protein in HIV-1 release. Retrovirology 2005, 2, 48. [Google Scholar] [CrossRef] [Green Version]
- Kazami, S.; Usui, T.; Osada, H. Actin stress fiber retraction and aggresome formation is a common cellular response to actin toxins. Biosci. Biotechnol. Biochem. 2011, 75, 1853–1855. [Google Scholar] [CrossRef]
- Yarmola, E.G.; Somasundaram, T.; Boring, T.A.; Spector, I.; Bubb, M.R. Actin-latrunculin A structure and function. Differential modulation of actin-binding protein function by latrunculin A. J. Biol. Chem. 2000, 275, 28120–28127. [Google Scholar]
- Stehn, J.R.; Haass, N.K.; Bonello, T.; Desouza, M.; Kottyan, G.; Treutlein, H.; Zeng, J.; Nascimento, P.R.; Sequeira, V.B.; Butler, T.L.; et al. A novel class of anticancer compounds targets the actin cytoskeleton in tumor cells. Cancer Res. 2013, 73, 5169–5182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foerster, F.; Braig, S.; Moser, C.; Kubisch, R.; Busse, J.; Wagner, E.; Schmoeckel, E.; Mayr, D.; Schmitt, S.; Huettel, S.; et al. Targeting the actin cytoskeleton: Selective antitumor action via trapping PKCvarepsilon. Cell Death Dis. 2014, 5, e1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, A.; Niemetz-Rahn, A.; Nonnenmacher, A.; Tucholski, J.; Keilholz, U.; Fusi, A. Expression and activity of EGFR in human cutaneous melanoma cell lines and influence of vemurafenib on the EGFR pathway. Target. Oncol. 2015, 10, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Grahn, J.C.; Isseroff, R.R. Human melanocytes do not express EGF receptors. J. Investig. Dermatol. 2004, 123, 244–246. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Gross, D.J. Regulated EGF receptor binding to F-actin modulates receptor phosphorylation. Biochem. Biophys. Res. Commun. 2003, 312, 930–936. [Google Scholar] [CrossRef]
- Scaltriti, M.; Baselga, J. The epidermal growth factor receptor pathway: A model for targeted therapy. Clin. Cancer Res. 2006, 12, 5268–5272. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.J.; Choi, Y.; Shin, J.M.; Cho, H.J.; Kang, J.H.; Park, K.K.; Choe, J.Y.; Bae, Y.S.; Han, S.M.; Kim, C.H.; et al. Melittin suppresses EGF-induced cell motility and invasion by inhibiting PI3K/Akt/mTOR signaling pathway in breast cancer cells. Food Chem. Toxicol. 2014, 123, 218–225. [Google Scholar] [CrossRef]
- Kim, S.K.; Park, K.Y.; Yoon, W.C.; Park, S.H.; Park, K.K.; Yoo, D.H.; Choe, J.Y. Melittin enhances apoptosis through suppression of IL-6/sIL-6R complex-induced NF-kB and STAT3 activation and Bcl-2 expression for human fibroblast-like synoviocytes in rheumatoid arthritis. Jt. Bone Spine 2011, 78, 471–477. [Google Scholar] [CrossRef]
- Sun, K.W.; Ma, Y.Y.; Guan, T.P.; Xia, Y.J.; Shao, C.M.; Chen, L.G.; Ren, Y.J.; Yao, H.B.; Yang, Q.; He, X.J. Oridonin induces apoptosis in gastric cancer through Apaf-1, cytochrome c and caspase-3 signaling pathway. World J. Gastroenterol. 2012, 18, 7166–7174. [Google Scholar] [CrossRef]
- Xin Ge, X.L.; Chen, Y.; Li, M.; Jiang, S.; Wang, X. Rhein induces apoptosis of HCT-116 human colon cancer cells via activation of the intrinsic apoptotic pathway. Afr. J. Biotechnol. 2011, 10, 13244–13251. [Google Scholar]
- Janssens, N.; Janicot, M.; Perera, T.; Bakker, A. Housekeeping genes as internal standards in cancer research. Mol. Diagn. 2004, 8, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Sangboonruang, S.; Thammasit, P.; Intasai, N.; Kasinrerk, W.; Tayapiwatana, C.; Tragoolpua, K. EMMPRIN reduction via scFv-M6-1B9 intrabody affects a3b1-integrin and MCT1 functions and results in suppression of progressive phenotype in the colorectal cancer cell line Caco-2. Cancer Gene Ther. 2014, 21, 246–255. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 (Mean ± SD) μg/mL |
|---|---|
| Mel-AF | 3.38 ± 0.16 |
| Mel-AM | 4.97 ± 0.23 |
| Doxorubicin (DXR) | 4.31 ± 0.30 |
| 5-Fluorouracil (5-FU) | >100 |
| Primers | Sequences | References |
|---|---|---|
| Bcl-2 | forward 5′-GTCTGGGAATCGATCTGGAAATCC-3′ | [48] |
| reverse 5′-TTTGAAACTTCCCAATGAATCAGGAG-3′ | ||
| Cytochrome c | forward 5′-GAGCGGGAGTGTTCGTTGT-3′ | [49] |
| reverse 5′-GTCTGCCCTTTCTTCCTTCT-3′ | ||
| Caspase-9 | forward 5′-TCAGGCCCCATATGATCG-3′ | [48] |
| reverse 5′-GACTCCCTCGAGTCTCCAGAT-3′ | ||
| Caspase-8 | forward 5′-GTGGAGGAAAGCAATCTGTC-3′ | [50] |
| reverse 5′-TATTAGCCCTGCCTGGTGTCT-3′ | ||
| Caspase-3 | forward 5′-TGTTTGTGTGCTTCTGAGCC-3′ | [48] |
| reverse 5′-TCAAGCTTGTCGGCATACTG-3′ | ||
| GAPDH | forward 5′-GAAGGTGAAGGTCGGAGTC-3′ | [51] |
| reverse 5′-GAAGATGGTGATGGGATTTC-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sangboonruang, S.; Kitidee, K.; Chantawannakul, P.; Tragoolpua, K.; Tragoolpua, Y. Melittin from Apis florea Venom as a Promising Therapeutic Agent for Skin Cancer Treatment. Antibiotics 2020, 9, 517. https://doi.org/10.3390/antibiotics9080517
Sangboonruang S, Kitidee K, Chantawannakul P, Tragoolpua K, Tragoolpua Y. Melittin from Apis florea Venom as a Promising Therapeutic Agent for Skin Cancer Treatment. Antibiotics. 2020; 9(8):517. https://doi.org/10.3390/antibiotics9080517
Chicago/Turabian StyleSangboonruang, Sirikwan, Kuntida Kitidee, Panuwan Chantawannakul, Khajornsak Tragoolpua, and Yingmanee Tragoolpua. 2020. "Melittin from Apis florea Venom as a Promising Therapeutic Agent for Skin Cancer Treatment" Antibiotics 9, no. 8: 517. https://doi.org/10.3390/antibiotics9080517