
A Review of Resistance to Polymyxins and Evolving Mobile Colistin Resistance Gene (mcr) among Pathogens of Clinical Significance



Abstract
:1. Introduction
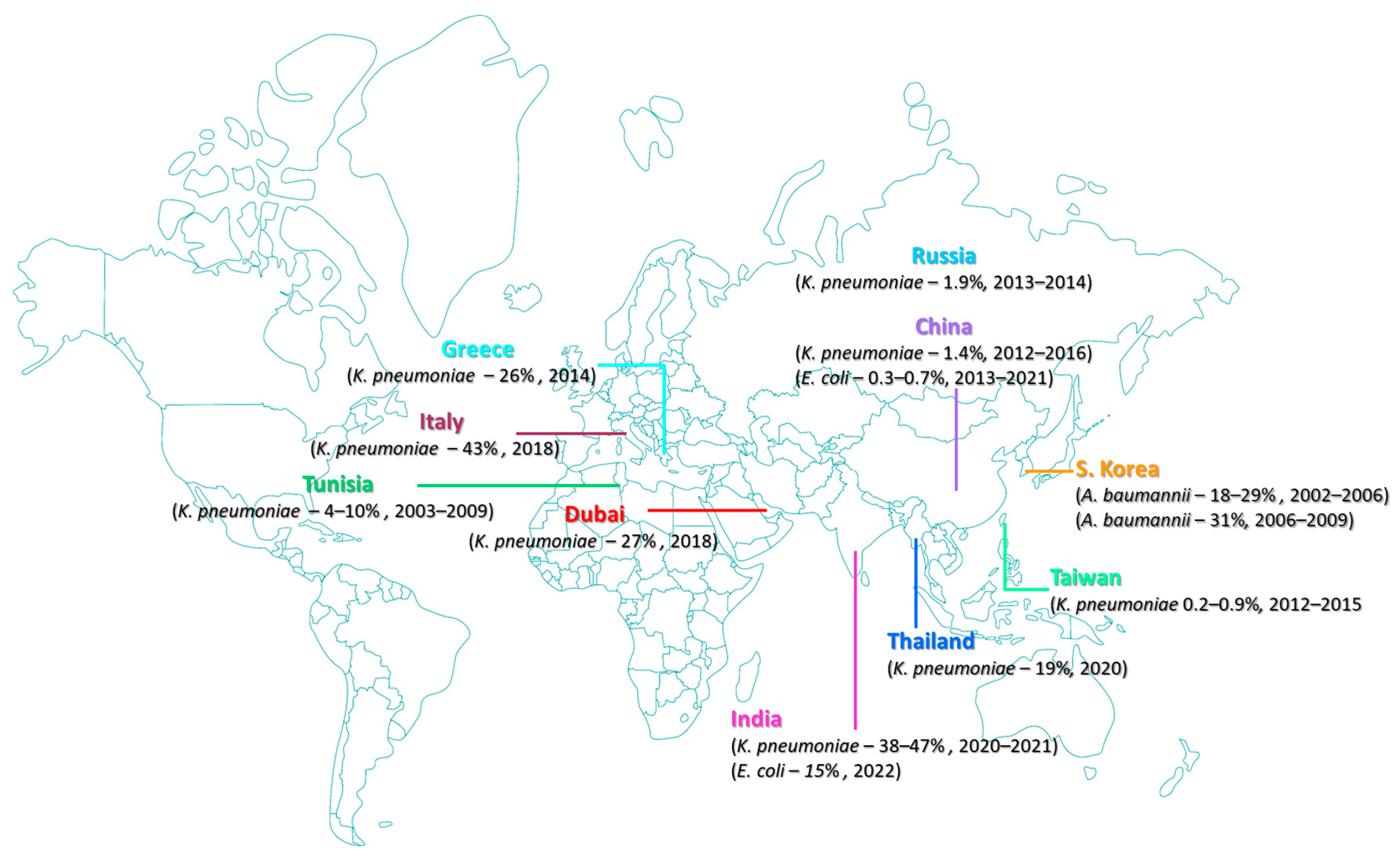
2. Rate of Resistance to Polymyxins among Pathogens of Clinical Significance
3. Mechanisms of Resistance to Polymyxins in Different Bacteria
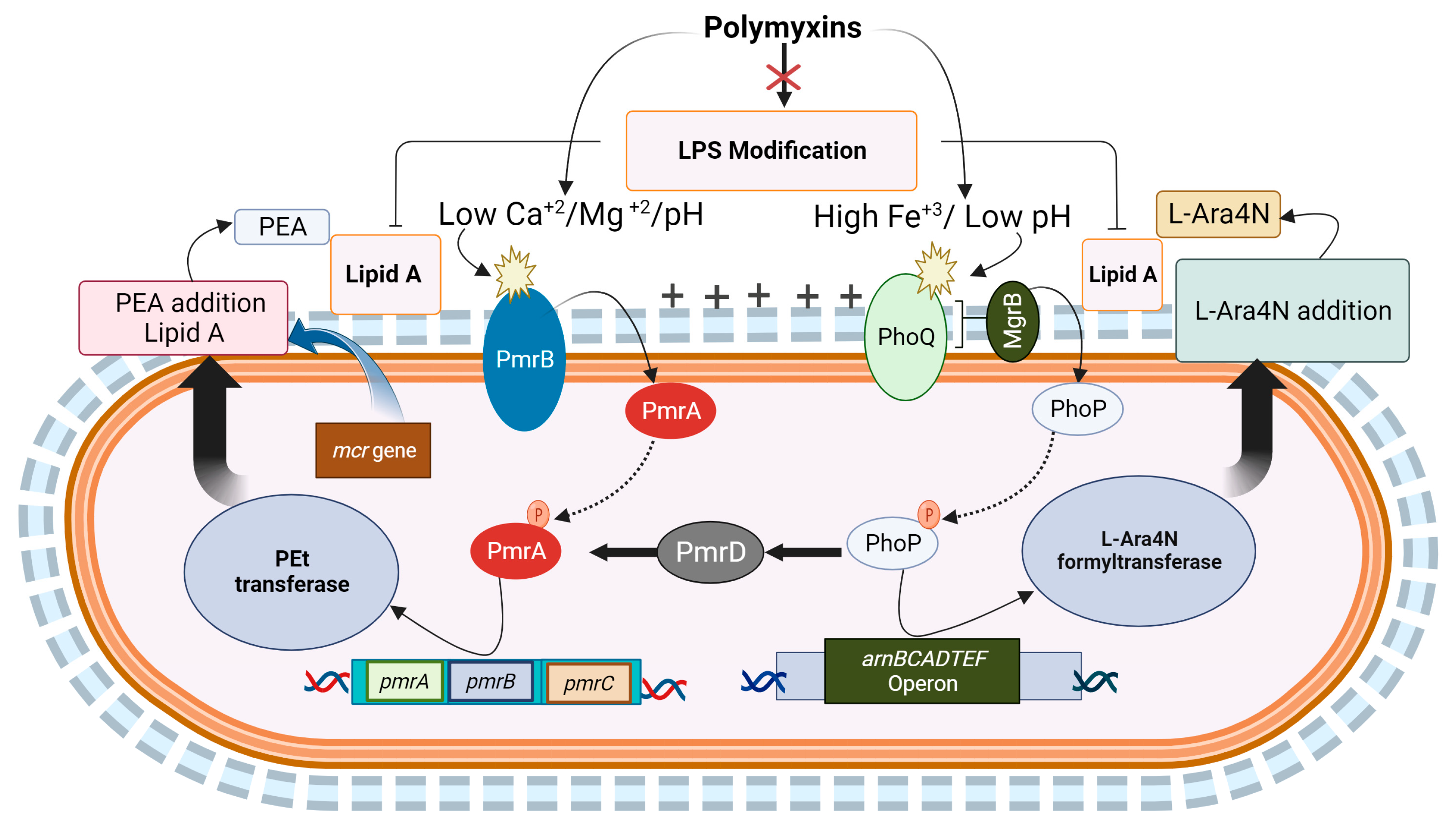
Evolving Intrinsic Mechanisms of Resistance to Polymyxins
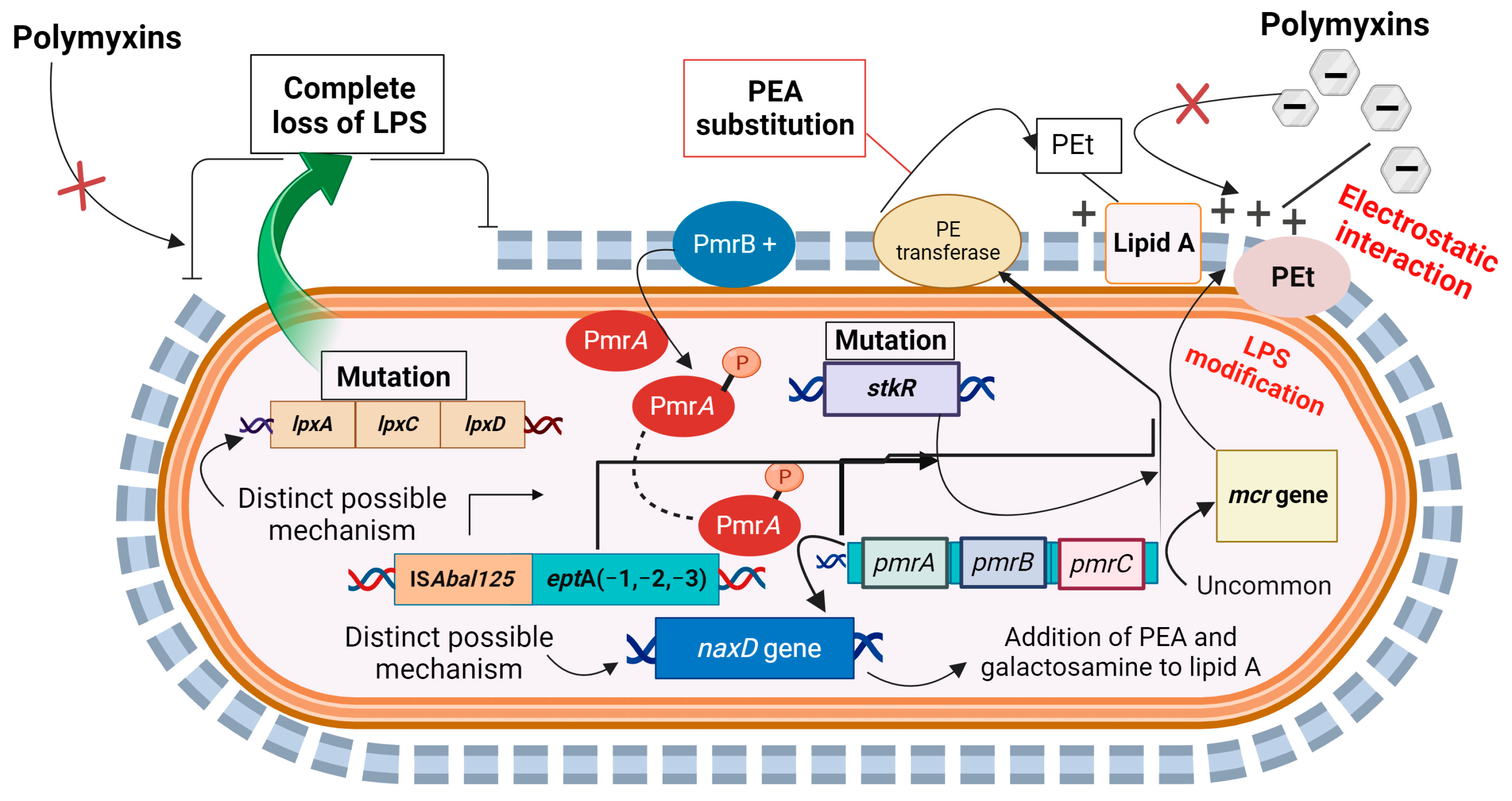
Distinct Mechanisms of Intrinsic Polymyxin Resistance in A. baumannii
4. Plasmid-Mediated mcr Gene-Based Polymyxins
4.1. Global Dissemination of mcr among Different Bacteria in Different Environments
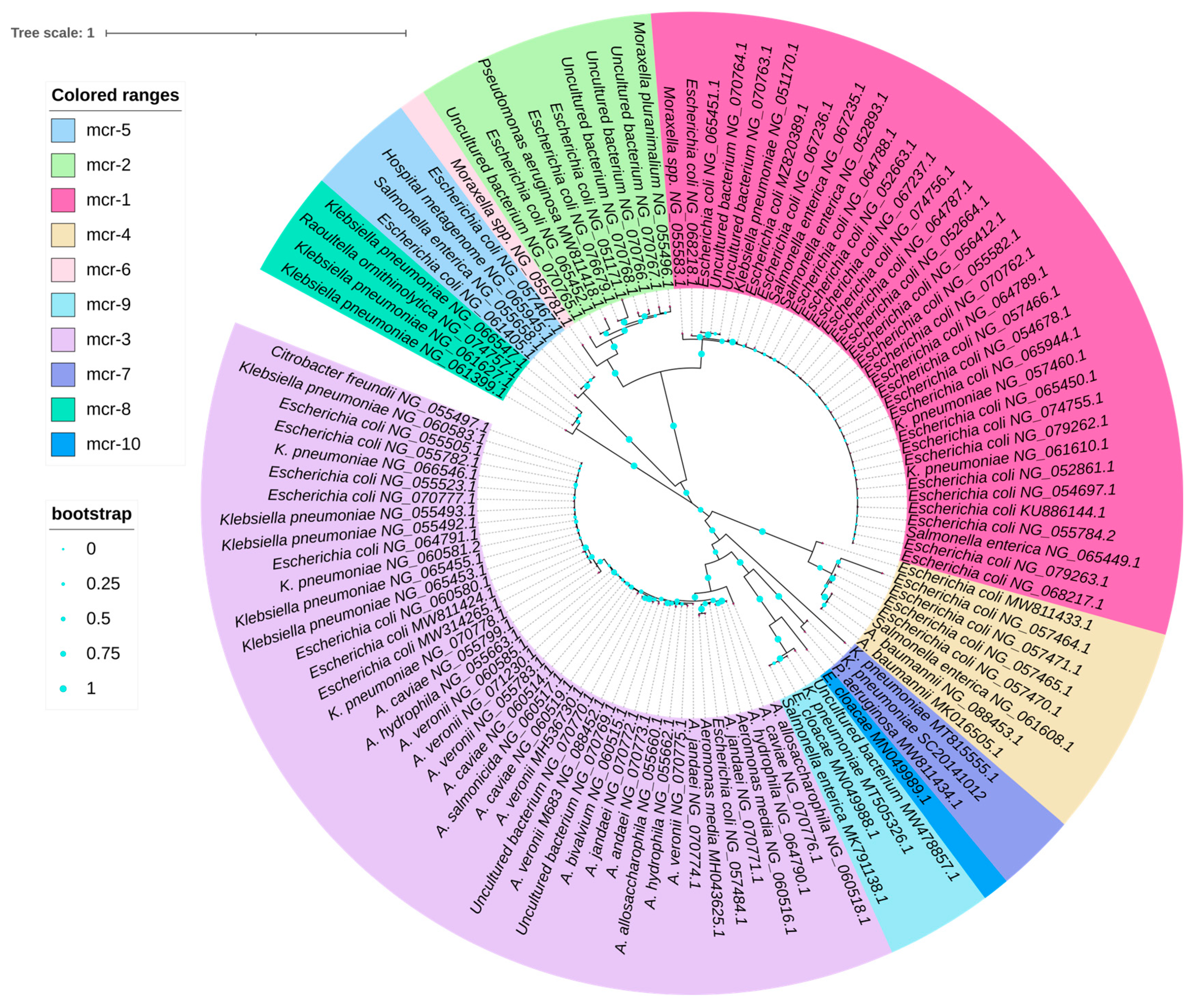
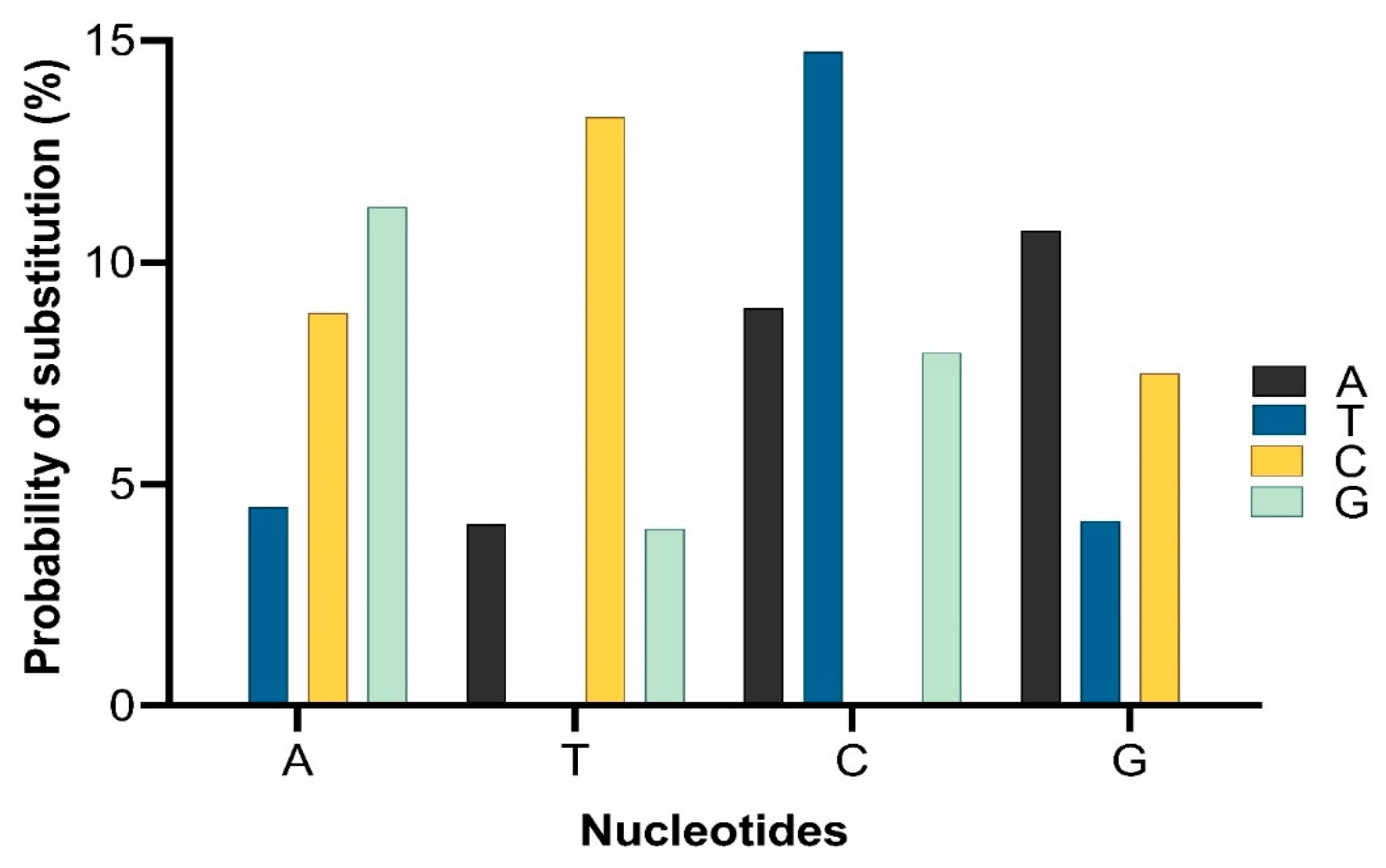
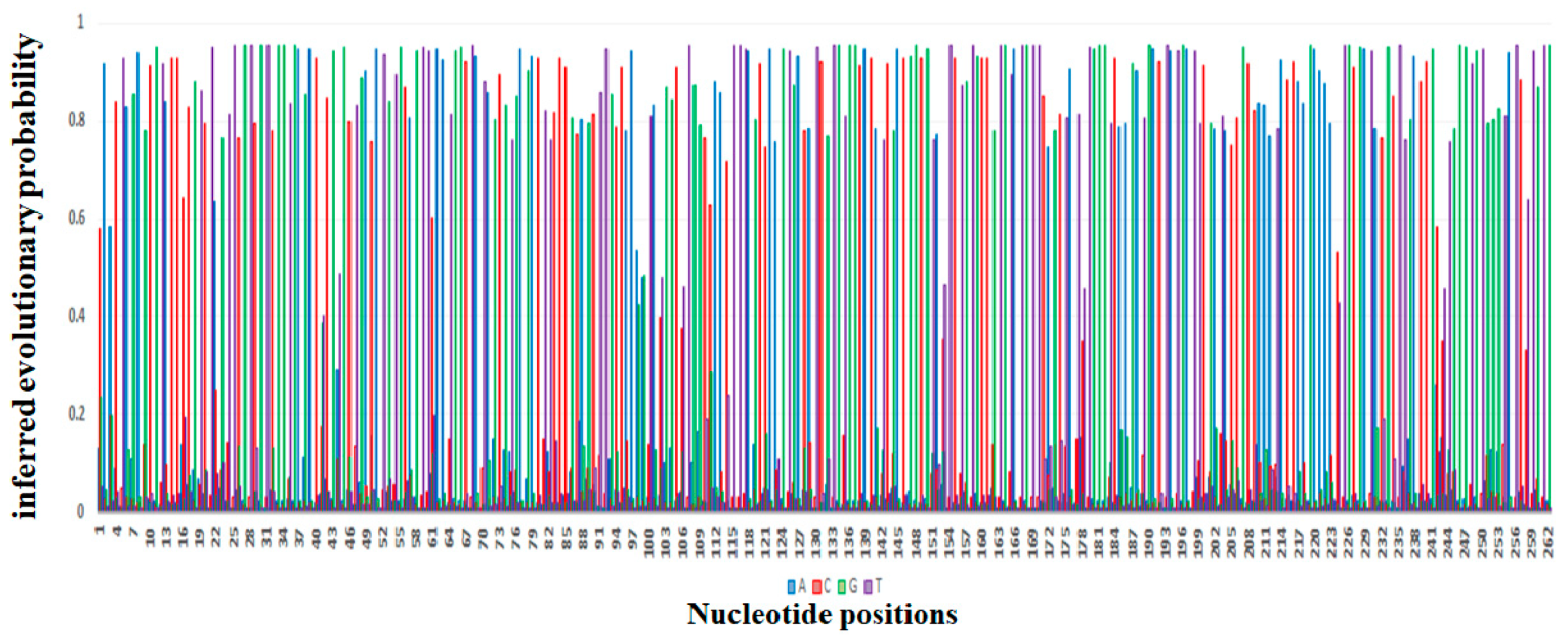
4.2. Evolution of mcr Gene Variants from mcr-1 to mcr-10
The Processes and Molecular Vehicles Responsible for the Transmission of mcr Variants

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mcr Variants | Insertion Sequences Structure | Transposon | Plasmids | Organism | Host (Isolated from) | Year of Discovery | References |
|---|---|---|---|---|---|---|---|
| mcr-1 | (ISApl1-mcr-1-pap2-ISApl1 and Tn7511) | Novel transposon Tn7511 | IncI1 plasmid, pMCR-E2899 | E. coli DH5α | Turkey meat | 2022 | [160] |
| mcr-1 | Combination of ISApl1 and IS91 (ISApl1-mcr-1-IS91) | Chromosomal Tn6330 transposon | IncI2 plasmid | E. coli | Community and hospital settings | 2022 | [74] |
| mcr-1 | IS26-mcr-1-PAP2, and ISAPl1-mcr-1-PAP2 and ISEcp1-blaCTX₋M₋₅₅-mcr-1-PAP2 | --- | IncI2, IncX4, and IncHI2 plasmids | E. coli and Salmonella spp. | Food products, food supply chain, and clinical samples | 2021 | [161,162] |
| mcr-1.1 | IS26-parA-mcr-1.1-pap2 | --- | IncX4-type plasmid | E. coli | Dog feces | 2020 | [150] |
| mcr-1 | I ISApl1-mcr-1-orf ISApl1 | ISApl1 transposon | IncHI2 and IncX4 plasmids | Enterobacteriaceae | Livestock | 2018 | [163] |
| mcr-1 | ISApl1-mcr-1-pap2-ISApl1 | Tn6330 | IncI2 and IncX4 plasmids | Novel Moraxella spp. | Pig | 2018 | [140] |
| mcr-1 | mcr-1-orf, ISApl1-mcr-1-orf and Tn6330 | Novel transposon Tn6330 | IncX4 and IncI2 plasmids | E. coli | Pig farms in China | 2017 | [162] |
| mcr-2 | (ISEc69-mcr-2-ORF-ISEc69 | Tn7052 | IncX4 conjugative plasmid | Moraxella osloensis | --- | 2021 | [164] |
| mcr-2 | ISEc69-mcr-2-ISEc69 | --- | IncX4 plasmid | M. bovoculi | Pigs, pork and chicken meat, and humans | 2017 | [165] |
| mcr- 3.1 | TnAs2-mcr-3.1-dgkA-ISKpn40 | Novel transposon Tn6330 | pCP61-IncFIB plasmid | E. coli | Pigs | 2021 | [166] |
| mcr-3.5 | IS4321R-TnAs2-mcr-3.5-dgkA-IS15 | Novel transposon Tn6330 | IncFIItype plasmid pCP55-IncFII | E. coli | Pigs | 2021 | [166] |
| mcr-3.7 | TnAs2-mcr-3.7-dgkA-IS26 | --- | IncP1 plasmid | E. coli | Dogs | 2020 | [150] |
| mcr-8 | IS903B-ampC-hp-hphp-Giy-T-dgkA-baeS-copR-IS3-mcr-8-Gly-T-IS5 | _ Δ IS66 transposases | IncFIA plasmid | K. pneumoniae | Patients from intensive care | 2022 | [167] |
| mcr-8 | IS903B-ymoA-inhA-mcr-8-copR-baeS-dgkA-ampC | Composite transposon | pABC264-OXA-181 plasmid | K. pneumoniae | Patient with bacteremia | 2022 | [168] |
| mcr-8.2 | ISEcl1-mcr-8.2-orf-ISKpn26 | --- | IncFII/FIA | K. pneumoniae | Patient’s Intestinal sample | 2022 | [169] |
| mcr-9.1 | IS903B-mcr-9.1-wbuC-IS26 | Tn6360 | IncHI2/2A plasmid | E. cloacae complex | Clinical isolates | 2022 | [170] |
| mcr-10 | ISKpn26 is present at upstream of xerC-mcr-10 and an IS26 | Transposon Tn1722 | IncFIA plasmid | Enterobacter roggenkampii | Clinical isolate | 2020 | [101] |
| mcr-10.1 | hsdSMR-ISEc36-mcr-10.1-xerC | --- | IncFIIK plasmids | K. pneumoniae | Clinical isolates | 2022 | [170] |
4.3. Methods for Detecting Polymyxin Resistance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Sayed Ahmed, M.A.E.; Zhong, L.L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.B. Colistin and its role in the Era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef] [PubMed]
- Trebosc, V.; Gartenmann, S.; Tötzl, M.; Lucchini, V.; Schellhorn, B.; Pieren, M.; Lociuro, S.; Gitzinger, M.; Tigges, M.; Bumann, D.; et al. Dissecting colistin resistance mechanisms in extensively drug-resistant Acinetobacter baumannii clinical isolates. mBio 2019, 10, e01083-19. [Google Scholar] [CrossRef] [PubMed]
- Stansly, P.G.; Shepherd, R.G.; White, H.J. Polymyxin: A new chemotherapeutic agent. Bull. Johns Hopkins Hosp. 1947, 81, 43–54. [Google Scholar] [PubMed]
- Vaara, M. Polymyxins and their potential next generation as therapeutic antibiotics. Front. Microbiol. 2019, 10, 1689. [Google Scholar] [CrossRef]
- Koyama, Y. A new antibiotic ‘colistin’ produced by spore-forming soil bacteria. J. Antibiot. 1950, 3, 457–458. [Google Scholar]
- Kwa, A.; Kasiakou, S.K.; Tam, V.H.; Falagas, M.E. Polymyxin B: Similarities to and differences from colistin (polymyxin E). Expert Rev. Anti. Infect. Ther. 2007, 5, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Katz, E.; Demain, A.L. The peptide antibiotics of Bacillus: Chemistry, biogenesis, and possible functions. Bacteriol. Rev. 1977, 41, 449–474. [Google Scholar] [CrossRef]
- Orwa, J.A.; Govaerts, C.; Busson, R.; Roets, E.; Van Schepdael, A.; Hoogmartens, J. Isolation and structural characterization of colistin components. J. Antibiot. 2001, 54, 595–599. [Google Scholar] [CrossRef]
- He, J.; Ledesma, K.R.; Lam, W.Y.; Figueroa, D.A.; Lim, T.P.; Chow, D.S.; Tam, V.H. Variability of polymyxin B major components in commercial formulations. Int. J. Antimicrob. Agents 2010, 35, 308–310. [Google Scholar] [CrossRef]
- Kassamali, Z.; Wang, W.; Jones, R.; Rotschafer, J.; Danziger, L.; Prince, R. Is what you see what you get? Product variation in clinical and reference formula-tions of polymyxin B. In Proceedings of the 23rd European Congress of Clinical Microbiology and Infectious Diseases (ECCMID), Berlin, Germany, 27–30 April 2013. [Google Scholar]
- Paterson, D.L.; Harris, P.N. Colistin resistance: A major breach in our last line of defence. Lancet Infect. Dis. 2016, 16, 132–133. [Google Scholar] [CrossRef]
- WHO. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Shahzad, S.; Ashraf, M.A.; Sajid, M.; Shahzad, A.; Rafique, A.; Mahmood, M.S. Evaluation of synergistic antimicrobial effect of vitamins (A, B1, B2, B6, B12, C, D, E and K) with antibiotics against resistant bacterial strains. J. Glob. Antimicrob. Resist. 2018, 13, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Trimble, M.J.; Mlynárčik, P.; Kolář, M.; Hancock, R.E. Polymyxin: Alternative mechanisms of action and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025288. [Google Scholar] [CrossRef] [PubMed]
- Gales, A.C.; Jones, R.N.; Sader, H.S. Contemporary activity of colistin and polymyxin B against a worldwide collection of Gram-negative pathogens: Results from the SENTRY Antimicrobial Surveillance Program (2006–2009). J. Antimicrob. Chemother. 2011, 66, 2070–2074. [Google Scholar] [CrossRef] [PubMed]
- Kontopidou, F.; Plachouras, D.; Papadomichelakis, E.; Koukos, G.; Galani, I.; Poulakou, G.; Dimopoulos, G.; Antoniadou, A.; Armaganidis, A.; Giamarellou, H. Colonization and infection by colistin-resistant Gram-negative bacteria in a cohort of critically ill patients. Clin. Microbiol. Infect. 2011, 17, E9–E11. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Wang, Y.; Shen, Y.; Shen, J.; Wu, C. Early emergence of mcr-1 in Escherichia coli from food-producing animals. Lancet Infect. Dis. 2016, 16, 293. [Google Scholar] [CrossRef] [PubMed]
- Skov, R.L.; Monnet, D.L. Plasmid-mediated colistin resistance (mcr-1 gene): Three months later, the story unfolds. Eurosurveillance 2016, 21, 30155. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.W.; Zhang, T.; Ma, J.H.; Fang, Y.; Wang, H.Y.; Huang, Z.X.; Wang, Y.; Wu, C.; Chen, G.X. Occurrence of plasmid- and chromosome-carried mcr-1 in waterborne Enterobacteriaceae in China. Antimicrob. Agents Chemother. 2017, 61, 00017-17. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Sun, J.; Liu, Y.; Feng, Y. The MCR-1 colistin resistance: A new challenge to global public health. Chin. Sci. Bull. 2017, 62, 1018–1029. [Google Scholar]
- Sun, J.; Li, X.P.; Fang, L.X.; Sun, R.Y.; He, Y.Z.; Lin, J.; Liao, X.P.; Feng, Y.; Liu, Y.H. Co-occurrence of mcr-1 in the chromosome and on an IncHI2 plasmid: Persistence of colistin resistance in Escherichia coli. Int. J. Antimicrob. Agents 2018, 51, 842–847. [Google Scholar] [CrossRef]
- Battikh, H.; Harchay, C.; Dekhili, A.; Khazar, K.; Kechrid, F.; Zribi, M.; Masmoudi, A.; Fendri, C. Clonal spread of colistin-resistant Klebsiella pneumoniae coproducing KPC and VIM carbapenemases in neonates at a Tunisian university hospital. Microb. Drug Resist. 2017, 23, 468–472. [Google Scholar] [CrossRef]
- Mezghani Maalej, S.; Rekik Meziou, M.; Mahjoubi, F.; Hammami, A. Epidemiological study of Enterobacteriaceae resistance to colistin in Sfax (Tunisia). Med. Mal. Infect. 2012, 42, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, X.; Wang, J.; Ouyang, P.; Jin, C.; Wang, R.; Zhang, Y.; Jin, L.; Chen, H.; Wang, Z.; et al. Phenotypic and genotypic characterization of carbapenem-resistant Enterobacteriaceae: Data from a longitudinal large-scale CRE study in China (2012–2016). Clin. Infect. Dis. 2018, 67, S196–S205. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Liu, Y.; Zhang, Q.; Jin, L.; Wang, Q.; Zhang, Y.; Wang, X.; Hu, M.; Li, L.; Qi, J.; et al. The prevalence of colistin resistance in Escherichia coli and Klebsiella pneumoniae isolated from food animals in China: Coexistence of mcr-1 and bla(NDM) with low fitness cost. Int. J. Antimicrob. Agents 2018, 51, 739–744. [Google Scholar] [CrossRef]
- Hejnar, P.; Kolár, M.; Hájek, V. Characteristics of Acinetobacter strains (phenotype classification, antibiotic susceptibility and production of beta-lactamases) isolated from haemocultures from patients at the Teaching Hospital in Olomouc. Acta Univ. Palacki. Olomuc. Fac. Med. 1999, 142, 73–77. [Google Scholar] [PubMed]
- Mahamat, A.; Bertrand, X.; Moreau, B.; Hommel, D.; Couppie, P.; Simonnet, C.; Kallel, H.; Demar, M.; Djossou, F.; Nacher, M. Clinical epidemiology and resistance mechanisms of carbapenem-resistant Acinetobacter baumannii, French Guiana, 2008–2014. Int. J. Antimicrob. Agents 2016, 48, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Sheck, E.A.; Edelstein, M.V.; Sukhorukova, M.V.; Ivanchik, N.V.; Skleenova, E.Y.; Dekhnich, A.V.; Azizov, I.S.; Kozlov, R.S. Epidemiology and genetic diversity of colistin nonsusceptible nosocomial Acinetobacter baumannii strains from Russia for 2013–2014. Can. J. Infect. Dis. Med. Microbiol. 2017, 2017, 1839190. [Google Scholar] [CrossRef] [PubMed]
- Giamarellou, H. Epidemiology of infections caused by polymyxin-resistant pathogens. Int. J. Antimicrob. Agents 2016, 48, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.S.; Suh, J.Y.; Kwon, K.T.; Jung, S.I.; Park, K.H.; Kang, C.I.; Chung, D.R.; Peck, K.R.; Song, J.H. High rates of resistance to colistin and polymyxin B in subgroups of Acinetobacter baumannii isolates from Korea. J. Antimicrob. Chemother. 2007, 60, 1163–1167. [Google Scholar] [CrossRef]
- Genteluci, G.L.; Gomes, D.B.; Souza, M.J.d.; Carvalho, K.R.; Villas-Bôas, M.H.S. Emergence of polymyxin B-resistant Acinetobacter baumannii in hospitals in Rio de Janeiro. J. Bras. Patol. Med. Lab. 2016, 52, 91–95. [Google Scholar] [CrossRef]
- Carrasco, L.D.M.; Dabul, A.N.G.; Boralli, C.; Righetto, G.M.; Carvalho, I.S.E.; Dornelas, J.V.; Martins da Mata, C.P.S.; de Araújo, C.A.; Leite, E.M.M.; Lincopan, N.; et al. Polymyxin resistance among XDR ST1 carbapenem-resistant Acinetobacter baumannii clone expanding in a teaching hospital. Front. Microbiol. 2021, 12, 622704. [Google Scholar] [CrossRef]
- European Centre for Disease Prevetion and Control (ECDC). Surveillance of Antimicrobial Resistance in Europe—Annual Report of the European Antimicrobial Resistance Surveillance Network (EARS-Net) 2017; ECDC: Solna, Sweden, 2017. [Google Scholar]
- Liu, L.; Liu, B.; Li, Y.; Zhang, W. Successful control of resistance in Pseudomonas aeruginosa using antibiotic stewardship and infection control programs at a Chinese university hospital: A 6-year prospective study. Infect. Drug Resist. 2018, 11, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Zhu, Y.; Yang, Z.; Shi, D.; Ni, Y.; Hua, L.; Li, J. Prevalence and molecular characteristics of polymyxin-resistant Pseudomonas aeruginosa in a chinese tertiary teaching hospital. Antibiotics 2022, 11, 799. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Stapleton, F.; Summers, S.; Rice, S.A.; Willcox, M.D.P. Antibiotic resistance characteristics of Pseudomonas aeruginosa isolated from keratitis in Australia and India. Antibiotics 2020, 9, 600. [Google Scholar] [CrossRef] [PubMed]
- Antoniadou, A.; Kontopidou, F.; Poulakou, G.; Koratzanis, E.; Galani, I.; Papadomichelakis, E.; Kopterides, P.; Souli, M.; Armaganidis, A.; Giamarellou, H. Colistin-resistant isolates of Klebsiella pneumoniae emerging in intensive care unit patients: First report of a multiclonal cluster. J. Antimicrob. Chemother. 2007, 59, 786–790. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevetion and Control (ECDC). Surveillance of Antimicrobial Resistance in Europe-2014; ECDC: Solna, Sweden, 2014. [Google Scholar]
- Moubareck, C.A.; Mouftah, S.F.; Pál, T.; Ghazawi, A.; Halat, D.H.; Nabi, A.; AlSharhan, M.A.; AlDeesi, Z.O.; Peters, C.C.; Celiloglu, H.; et al. Clonal emergence of Klebsiella pneumoniae ST14 co-producing OXA-48-type and NDM carbapenemases with high rate of colistin resistance in Dubai, United Arab Emirates. Int. J. Antimicrob. Agents 2018, 52, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Uzairue, L.I.; Rabaan, A.A.; Adewumi, F.A.; Okolie, O.J.; Folorunso, J.B.; Bakhrebah, M.A.; Garout, M.; Alfouzan, W.A.; Halwani, M.A.; Alamri, A.A.; et al. Global Prevalence of colistin resistance in Klebsiella pneumoniae from bloodstream infection: A systematic review and meta-analysis. Pathogens 2022, 11, 1092. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, K.; Mittal, V.; Arya, M.; Kumar, M.; Phillips, A.; Kajla, B. Pattern of colistin resistance in Klebsiella isolates in an intensive care unit of a tertiary care hospital in India. J. Infect. Public Health 2020, 13, 1018–1021. [Google Scholar] [CrossRef]
- Singh, A.; Keshri, A.K.; Rawat, S.S.; Swami, D.; Uday, K.V.; Prasad, A. Identification and characterization of colistin-resistant E. coli and K. pneumoniae isolated from lower Himalayan region of India. SN Appl. Sci. 2021, 3, 615. [Google Scholar] [CrossRef]
- Kuo, S.C.; Huang, W.C.; Wang, H.Y.; Shiau, Y.R.; Cheng, M.F.; Lauderdale, T.L. Colistin resistance gene mcr-1 in Escherichia coli isolates from humans and retail meats, Taiwan. J. Antimicrob. Chemother. 2016, 71, 2327–2329. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Ke, B.; Zhao, X.; Guo, Y.; Wang, W.; Wang, X.; Zhu, H. Antimicrobial resistance profile of mcr-1 positive clinical isolates of Escherichia coli in China from 2013 to 2016. Front. Microbiol. 2018, 9, 2514. [Google Scholar] [CrossRef]
- Yan, W.; Zhang, Q.; Zhu, Y.; Jing, N.; Yuan, Y.; Zhang, Y.; Ren, S.; Hu, D.; Zhao, W.; Zhang, X.; et al. Molecular mechanism of polymyxin resistance in multidrug-resistant Klebsiella pneumoniae and Escherichia coli isolates from Henan Province, China: A multicenter study. Infect. Drug Resist. 2021, 14, 2657–2666. [Google Scholar] [CrossRef] [PubMed]
- Bir, R.; Gautam, H.; Arif, N.; Chakravarti, P.; Verma, J.; Banerjee, S.; Tyagi, S.; Mohapatra, S.; Sood, S.; Dhawan, B.; et al. Analysis of colistin resistance in carbapenem-resistant Enterobacterales and XDR Klebsiella pneumoniae. Ther. Adv. Infect. Dis. 2022, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- López-Rojas, R.; Domínguez-Herrera, J.; McConnell, M.J.; Docobo-Peréz, F.; Smani, Y.; Fernández-Reyes, M.; Rivas, L.; Pachón, J. Impaired virulence and In Vivo fitness of colistin-resistant Acinetobacter baumannii. J. Infect. Dis. 2011, 203, 545–548. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Beaudry, F.; Thériault, W.; Letellier, A. Colistin in pig production: Chemistry, mechanism of antibacterial action, microbial resistance emergence, and one health perspectives. Front. Microbiol. 2016, 7, 1789. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S. The Salmonella PmrAB regulon: Lipopolysaccharide modifications, antimicrobial peptide resistance and more. Trends Microbiol. 2008, 16, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Cannatelli, A.; D’Andrea, M.M.; Giani, T.; Di Pilato, V.; Arena, F.; Ambretti, S.; Gaibani, P.; Rossolini, G.M. In Vivo emergence of colistin resistance in Klebsiella pneumoniae producing KPC-type carbapenemases mediated by insertional inactivation of the PhoQ/PhoP mgrB regulator. Antimicrob. Agents Chemother. 2013, 57, 5521–5526. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Guan, Z.; Raetz, C.R. An undecaprenyl phosphate-aminoarabinose flippase required for polymyxin resistance in Escherichia coli. J. Biol. Chem. 2007, 282, 36077–36089. [Google Scholar] [CrossRef]
- Wright, M.S.; Suzuki, Y.; Jones, M.B.; Marshall, S.H.; Rudin, S.D.; van Duin, D.; Kaye, K.; Jacobs, M.R.; Bonomo, R.A.; Adams, M.D. Genomic and transcriptomic analyses of colistin-resistant clinical isolates of Klebsiella pneumoniae reveal multiple pathways of resistance. Antimicrob. Agents Chemother. 2015, 59, 536–543. [Google Scholar] [CrossRef]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef]
- Baron, S.; Leulmi, Z.; Villard, C.; Olaitan, A.O.; Telke, A.A.; Rolain, J.M. Inactivation of the arn operon and loss of aminoarabinose on lipopolysaccharide as the cause of susceptibility to colistin in an atypical clinical isolate of Proteus vulgaris. Int. J. Antimicrob. Agents 2018, 51, 450–457. [Google Scholar] [CrossRef]
- Phan, M.D.; Nhu, N.T.K.; Achard, M.E.S.; Forde, B.M.; Hong, K.W.; Chong, T.M.; Yin, W.F.; Chan, K.G.; West, N.P.; Walker, M.J.; et al. Modifications in the pmrB gene are the primary mechanism for the development of chromosomally encoded resistance to polymyxins in uropathogenic Escherichia coli. J. Antimicrob. Chemother. 2017, 72, 2729–2736. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, M.R.; Casella, L.G.; Jones, J.W.; Adams, M.D.; Zurawski, D.V.; Hazlett, K.R.; Doi, Y.; Ernst, R.K. Unique structural modifications are present in the lipopolysaccharide from colistin-resistant strains of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2013, 57, 4831–4840. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wei, W.; Lei, S.; Lin, J.; Srinivas, S.; Feng, Y. An evolutionarily conserved mechanism for intrinsic and transferable polymyxin resistance. mBio 2018, 9, e02317-17. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Lin, T.L.; Lin, Y.T.; Wang, J.T. A putative RND-type efflux pump, H239_3064, contributes to colistin resistance through CrrB in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2018, 73, 1509–1516. [Google Scholar] [CrossRef]
- Herrera, C.M.; Crofts, A.A.; Henderson, J.C.; Pingali, S.C.; Davies, B.W.; Trent, M.S. Correction for Herrera et al., The Vibrio cholerae VprA-VprB two-component system controls virulence through endotoxin modification. mBio 2015, 6, e00155. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; Jenssen, H.; Bains, M.; Wiegand, I.; Gooderham, W.J.; Hancock, R.E. The two-component system CprRS senses cationic peptides and triggers adaptive resistance in Pseudomonas aeruginosa independently of ParRS. Antimicrob. Agents Chemother. 2012, 56, 6212–6222. [Google Scholar] [CrossRef]
- Jones, C.L.; Singh, S.S.; Alamneh, Y.; Casella, L.G.; Ernst, R.K.; Lesho, E.P.; Waterman, P.E.; Zurawski, D.V. In Vivo fitness adaptations of colistin-resistant Acinetobacter baumannii isolates to oxidative stress. Antimicrob. Agents Chemother. 2017, 61, e00598-16. [Google Scholar] [CrossRef] [PubMed]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter baumannii antibiotic resistance mechanisms. Pathogens 2021, 10, 373. [Google Scholar] [CrossRef]
- Da Silva, G.J.; Domingues, S. Interplay between colistin resistance, virulence and fitness in Acinetobacter baumannii. Antibiotics 2017, 6, 28. [Google Scholar] [CrossRef]
- Jaidane, N.; Naas, T.; Mansour, W.; Radhia, B.B.; Jerbi, S.; Boujaafar, N.; Bouallegue, O.; Bonnin, R.A. Genomic analysis of In Vivo acquired resistance to colistin and rifampicin in Acinetobacter baumannii. Int. J. Antimicrob. Agents 2018, 51, 266–269. [Google Scholar] [CrossRef]
- Boinett, C.J.; Cain, A.K.; Hawkey, J.; Do Hoang, N.T.; Khanh, N.N.T.; Thanh, D.P.; Dordel, J.; Campbell, J.I.; Lan, N.P.H.; Mayho, M.; et al. Clinical and laboratory-induced colistin-resistance mechanisms in Acinetobacter baumannii. Microb. Genom. 2019, 5, e000246. [Google Scholar] [CrossRef] [PubMed]
- Boll, J.M.; Crofts, A.A.; Peters, K.; Cattoir, V.; Vollmer, W.; Davies, B.W.; Trent, M.S. A penicillin-binding protein inhibits selection of colistin-resistant, lipooligosaccharide-deficient Acinetobacter baumannii. Proc. Natl. Acad. Sci. USA 2016, 113, E6228–E6237. [Google Scholar] [CrossRef] [PubMed]
- Sabnis, A.; Hagart, K.L.; Klöckner, A.; Becce, M.; Evans, L.E.; Furniss, R.C.D.; Mavridou, D.A.; Murphy, R.; Stevens, M.M.; Davies, J.C.; et al. Colistin kills bacteria by targeting lipopolysaccharide in the cytoplasmic membrane. Elife 2021, 10, e65836. [Google Scholar] [CrossRef] [PubMed]
- Bakthavatchalam, Y.D.; Pragasam, A.K.; Biswas, I.; Veeraraghavan, B. Polymyxin susceptibility testing, interpretative breakpoints and resistance mechanisms: An update. J. Glob. Antimicrob. Resist. 2018, 12, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Beceiro, A.; Llobet, E.; Aranda, J.; Bengoechea, J.A.; Doumith, M.; Hornsey, M.; Dhanji, H.; Chart, H.; Bou, G.; Livermore, D.M.; et al. Phosphoethanolamine modification of lipid A in colistin-resistant variants of Acinetobacter baumannii mediated by the pmrAB two-component regulatory system. Antimicrob. Agents Chemother. 2011, 55, 3370–3379. [Google Scholar] [CrossRef] [PubMed]
- Girardello, R.; Visconde, M.; Cayô, R.; Figueiredo, R.C.; Mori, M.A.; Lincopan, N.; Gales, A.C. Diversity of polymyxin resistance mechanisms among Acinetobacter baumannii clinical isolates. Diagn. Microbiol. Infect. Dis. 2017, 87, 37–44. [Google Scholar] [CrossRef]
- Deveson Lucas, D.; Crane, B.; Wright, A.; Han, M.L.; Moffatt, J.; Bulach, D.; Gladman, S.L.; Powell, D.; Aranda, J.; Seemann, T.; et al. Emergence of high-level colistin resistance in an Acinetobacter baumannii clinical isolate mediated by inactivation of the global regulator H-NS. Antimicrob. Agents Chemother. 2018, 62, e02442-17. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Yong, D.; Jeong, S.H.; Chong, Y. Multidrug-resistant Acinetobacter spp.: Increasingly problematic nosocomial pathogens. Yonsei Med. J. 2011, 52, 879–891. [Google Scholar] [CrossRef]
- Giles, S.K.; Stroeher, U.H.; Papudeshi, B.; Edwards, R.A.; Carlson-Jones, J.A.; Roach, M.; Brown, M.H. The StkSR two-component system influences colistin resistance in Acinetobacter baumannii. Microorganisms 2022, 10, 985. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Gao, R.; Hu, Y.; Li, Z.; Sun, J.; Wang, Q.; Lin, J.; Ye, H.; Liu, F.; Srinivas, S.; Li, D.; et al. Dissemination and mechanism for the MCR-1 colistin resistance. PLoS Pathog. 2016, 12, e1005957. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Yang, Y.; Li, W.; Ma, X.; Zhang, C.; Zhang, J.; Sun, B.; Ding, T.; Tian, G.B. Comparative genomic analyses of polymyxin-resistant Enterobacteriaceae strains from China. BMC Genom. 2022, 23, 88. [Google Scholar] [CrossRef] [PubMed]
- Anandan, A.; Evans, G.L.; Condic-Jurkic, K.; O’Mara, M.L.; John, C.M.; Phillips, N.J.; Jarvis, G.A.; Wills, S.S.; Stubbs, K.A.; Moraes, I.; et al. Structure of a lipid A phosphoethanolamine transferase suggests how conformational changes govern substrate binding. Proc. Natl. Acad. Sci. USA 2017, 114, 2218–2223. [Google Scholar] [CrossRef] [PubMed]
- Hanson, S.R.; Best, M.D.; Wong, C.H. Sulfatases: Structure, mechanism, biological activity, inhibition, and synthetic utility. Angew. Chem. Int. Ed. 2004, 43, 5736–5763. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, M.; Abo, H.; Ozawa, M.; Uchiyama, M.; Shirakawa, T.; Suzuki, S.; Shima, A.; Yamashita, A.; Sekizuka, T.; Kato, K.; et al. Prevalence of colistin resistance gene mcr-1 and absence of mcr-2 in Escherichia coli isolated from healthy food-producing animals in Japan. Antimicrob. Agents Chemother. 2017, 61, e02057-16. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, L.; Wang, J.; Yassin, A.K.; Butaye, P.; Kelly, P.; Gong, J.; Guo, W.; Li, J.; Li, M.; et al. Molecular detection of colistin resistance genes (mcr-1, mcr-2 and mcr-3) in nasal/oropharyngeal and anal/cloacal swabs from pigs and poultry. Sci. Rep. 2018, 8, 3705. [Google Scholar] [CrossRef] [PubMed]
- Belaynehe, K.M.; Shin, S.W.; Park, K.Y.; Jang, J.Y.; Won, H.G.; Yoon, I.J.; Yoo, H.S. Emergence of mcr-1 and mcr-3 variants coding for plasmid-mediated colistin resistance in Escherichia coli isolates from food-producing animals in South Korea. Int. J. Infect. Dis. 2018, 72, 22–24. [Google Scholar] [CrossRef]
- Xu, Y.; Zhong, L.L.; Srinivas, S.; Sun, J.; Huang, M.; Paterson, D.L.; Lei, S.; Lin, J.; Li, X.; Tang, Z.; et al. Spread of MCR-3 colistin resistance in China: An epidemiological, genomic and mechanistic study. EBioMedicine 2018, 34, 139–157. [Google Scholar] [CrossRef]
- Ling, Z.; Yin, W.; Li, H.; Zhang, Q.; Wang, X.; Wang, Z.; Ke, Y.; Wang, Y.; Shen, J. Chromosome-mediated mcr-3 Variants in Aeromonas veronii from chicken meat. Antimicrob. Agents Chemother. 2017, 61, e01272-17. [Google Scholar] [CrossRef]
- Xiang, R.; Liu, B.H.; Zhang, A.Y.; Lei, C.W.; Ye, X.L.; Yang, Y.X.; Chen, Y.P.; Wang, H.N. Colocation of the polymyxin resistance gene mcr-1 and a variant of mcr-3 on a plasmid in an Escherichia coli isolate from a chicken farm. Antimicrob. Agents Chemother. 2018, 62, e00501-18. [Google Scholar] [CrossRef]
- Liu, L.; Feng, Y.; Zhang, X.; McNally, A.; Zong, Z. New Variant of mcr-3 in an extensively drug-resistant Escherichia coli clinical isolate carrying mcr-1 and bla(NDM-5). Antimicrob. Agents Chemother. 2017, 61, e01757-17. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, J.; Wang, J.; Butaye, P.; Kelly, P.; Li, M.; Yang, F.; Gong, J.; Yassin, A.K.; Guo, W.; et al. Newly identified colistin resistance genes, mcr-4 and mcr-5, from upper and lower alimentary tract of pigs and poultry in China. PLoS ONE 2018, 13, e0193957. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Villa, L.; Feudi, C.; Curcio, L.; Orsini, S.; Luppi, A.; Pezzotti, G.; Magistrali, C.F. Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. EuroSurveillance 2017, 22, 30589. [Google Scholar] [CrossRef]
- Zhang, H.; Hou, M.; Xu, Y.; Srinivas, S.; Huang, M.; Liu, L.; Feng, Y. Action and mechanism of the colistin resistance enzyme MCR-4. Commun. Biol. 2019, 2, 36. [Google Scholar] [CrossRef] [PubMed]
- Carretto, E.; Brovarone, F.; Nardini, P.; Russello, G.; Barbarini, D.; Pongolini, S.; Gagliotti, C.; Carattoli, A.; Sarti, M. Detection of mcr-4 positive Salmonella enterica serovar Typhimurium in clinical isolates of human origin, Italy, October to November 2016. EuroSurveillance 2018, 23, 17-00821. [Google Scholar] [CrossRef]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. EuroSurveillance 2018, 23, 17-00672. [Google Scholar] [CrossRef] [PubMed]
- Hammerl, J.A.; Borowiak, M.; Schmoger, S.; Shamoun, D.; Grobbel, M.; Malorny, B.; Tenhagen, B.A.; Käsbohrer, A. mcr-5 and a novel mcr-5.2 variant in Escherichia coli isolates from food and food-producing animals, Germany, 2010 to 2017. J. Antimicrob. Chemother. 2018, 73, 1433–1435. [Google Scholar] [CrossRef]
- Partridge, S.R.; Di Pilato, V.; Doi, Y.; Feldgarden, M.; Haft, D.H.; Klimke, W.; Kumar-Singh, S.; Liu, J.H.; Malhotra-Kumar, S.; Prasad, A.; et al. Proposal for assignment of allele numbers for mobile colistin resistance (mcr) genes. J. Antimicrob. Chemother. 2018, 73, 2625–2630. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Cerdeira, L.; Silva, M.M.; Sellera, F.P.; Muñoz, M.; Junior, F.G.; Azevedo, S.S.; Power, P.; Gutkind, G.; Lincopan, N. Novel mcr-5.3 variant in a CTX-M-8-producing Escherichia coli ST711 isolated from an infected horse. J. Antimicrob. Chemother. 2018, 73, 3520–3522. [Google Scholar] [CrossRef]
- Fleres, G.; Couto, N.; Schuele, L.; Chlebowicz, M.A.; Mendes, C.I.; van der Sluis, L.W.M.; Rossen, J.W.A.; Friedrich, A.W.; García-Cobos, S. Detection of a novel mcr-5.4 gene variant in hospital tap water by shotgun metagenomic sequencing. J. Antimicrob. Chemother. 2019, 74, 3626–3628. [Google Scholar] [CrossRef]
- AbuOun, M.; Stubberfield, E.J.; Duggett, N.A.; Kirchner, M.; Dormer, L.; Nunez-Garcia, J.; Randall, L.P.; Lemma, F.; Crook, D.W.; Teale, C.; et al. mcr-1 and mcr-2 variant genes identified in Moraxella species isolated from pigs in Great Britain from 2014 to 2015. J. Antimicrob. Chemother. 2017, 72, 2745–2749. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Q.; Li, Y.X.; Lei, C.W.; Zhang, A.Y.; Wang, H.N. Novel plasmid-mediated colistin resistance gene mcr-7.1 in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2018, 73, 1791–1795. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Zhou, Y.; Li, J.; Yin, W.; Wang, S.; Zhang, S.; Shen, J.; Shen, Z.; Wang, Y. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg. Microbes Infect. 2018, 7, 122. [Google Scholar] [CrossRef]
- Qin, S.; Zhang, C.; Schwarz, S.; Li, L.; Dong, H.; Yao, H.; Du, X.D. Identification of a novel conjugative mcr-8.2-bearing plasmid in an almost pan-resistant hypermucoviscous Klebsiella pneumoniae ST11 isolate with enhanced virulence. J. Antimicrob. Chemother. 2020, 75, 2696–2699. [Google Scholar] [CrossRef] [PubMed]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of novel mobilized colistin resistance gene mcr-9 in a multidrug-resistant, colistin-susceptible Salmonella enterica serotype Typhimurium isolate. mBio 2019, 10, e00853-19. [Google Scholar] [CrossRef] [PubMed]
- Tyson, G.H.; Li, C.; Hsu, C.H.; Ayers, S.; Borenstein, S.; Mukherjee, S.; Tran, T.T.; McDermott, P.F.; Zhao, S. The mcr-9 gene of Salmonella and Escherichia coli is not associated with colistin resistance in the United States. Antimicrob. Agents Chemother. 2020, 64, e00573-20. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Feng, Y.; Liu, L.; Wei, L.; Kang, M.; Zong, Z. Identification of novel mobile colistin resistance gene mcr-10. Emerg. Microbes Infect. 2020, 9, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Tamura, K.; Sanderford, M.; Gray, V.E.; Kumar, S. A molecular evolutionary reference for the human variome. Mol. Biol. Evol. 2016, 33, 245–254. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Buess, S.; Nüesch-Inderbinen, M.; Stephan, R.; Zurfluh, K. Assessment of animals as a reservoir for colistin resistance: No MCR-1/MCR-2-producing Enterobacteriaceae detected in Swiss livestock. J. Glob. Antimicrob. Resist. 2017, 8, 33–34. [Google Scholar] [CrossRef]
- Girardello, R.; Piroupo, C.M.; Martins, J., Jr.; Maffucci, M.H.; Cury, A.P.; Franco, M.R.G.; Malta, F.M.; Rocha, N.C.; Pinho, J.R.R.; Rossi, F.; et al. Genomic characterization of mcr-1.1-producing Escherichia coli recovered from human infections in São Paulo, Brazil. Front. Microbiol. 2021, 12, 663414. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, R.; Card, R.M.; Nunez, J.; Pomba, C.; Mendonça, N.; Anjum, M.F.; Da Silva, G.J. Detection of an mcr-1-encoding plasmid mediating colistin resistance in Salmonella enterica from retail meat in Portugal. J. Antimicrob. Chemother. 2016, 71, 2338–2340. [Google Scholar] [CrossRef] [PubMed]
- Gogry, F.A.; Siddiqui, M.T.; Haq, Q.M.R. Emergence of mcr-1 conferred colistin resistance among bacterial isolates from urban sewage water in India. Environ. Sci. Pollut. Res. Int. 2019, 26, 33715–33717. [Google Scholar] [CrossRef] [PubMed]
- Bilal, H.; Rehman, T.U.; Khan, M.A.; Hameed, F.; Jian, Z.G.; Han, J.; Yang, X. Molecular epidemiology of mcr-1, bla (KPC-2,) and bla (NDM-1) harboring clinically isolated Escherichia coli from Pakistan. Infect. Drug Resist. 2021, 14, 1467–1479. [Google Scholar] [CrossRef] [PubMed]
- Vu Thi Ngoc, B.; Le Viet, T.; Nguyen Thi Tuyet, M.; Nguyen Thi Hong, T.; Nguyen Thi Ngoc, D.; Le Van, D.; Chu Thi, L.; Tran Huy, H.; Penders, J.; Wertheim, H.; et al. Characterization of genetic elements carrying mcr-1 gene in Escherichia coli from the community and hospital settings in Vietnam. Microbiol. Spectr. 2022, 10, e0135621. [Google Scholar] [CrossRef] [PubMed]
- Hadjadj, L.; Baron, S.A.; Olaitan, A.O.; Morand, S.; Rolain, J.M. Co-occurrence of variants of mcr-3 and mcr-8 Genes in a Klebsiella pneumoniae isolate from Laos. Front. Microbiol. 2019, 10, 2720. [Google Scholar] [CrossRef] [PubMed]
- McGann, P.; Snesrud, E.; Maybank, R.; Corey, B.; Ong, A.C.; Clifford, R.; Hinkle, M.; Whitman, T.; Lesho, E.; Schaecher, K.E. Escherichia coli harboring mcr-1 and blaCTX-M on a novel IncF plasmid: First report of mcr-1 in the United States. Antimicrob. Agents Chemother. 2016, 60, 4420–4421. [Google Scholar] [CrossRef] [PubMed]
- Cannatelli, A.; Giani, T.; Antonelli, A.; Principe, L.; Luzzaro, F.; Rossolini, G.M. First detection of the mcr-1 colistin resistance gene in Escherichia coli in Italy. Antimicrob. Agents Chemother. 2016, 60, 3257–3258. [Google Scholar] [CrossRef]
- Bhat, A.H. Bacterial zoonoses transmitted by household pets and as reservoirs of antimicrobial resistant bacteria. Microb. Pathog. 2021, 155, 104891. [Google Scholar] [CrossRef]
- Skarżyńska, M.; Zaja, C.M.; Bomba, A.; Bocian, Ł.; Kozdruń, W.; Polak, M.; Wia Cek, J.; Wasyl, D. Antimicrobial resistance glides in the Sky-Free-Living Birds as a reservoir of resistant Escherichia coli with zoonotic potential. Front. Microbiol. 2021, 12, 656223. [Google Scholar] [CrossRef]
- Zurfluh, K.; Nüesch-Inderbinen, M.; Klumpp, J.; Poirel, L.; Nordmann, P.; Stephan, R. Key features of mcr-1-bearing plasmids from Escherichia coli isolated from humans and food. Antimicrob. Resist. Infect. Control 2017, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.R.; Sellera, F.P.; Esposito, F.; Sabino, C.P.; Cerdeira, L.; Lincopan, N. Colistin-resistant mcr-1-positive Escherichia coli on public beaches, an infectious threat emerging in recreational waters. Antimicrob. Agents Chemother. 2017, 61, e00234-17. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Feng, Y.; Lü, X.; McNally, A.; Zong, Z. IncP plasmid carrying colistin resistance gene mcr-1 in Klebsiella pneumoniae from hospital sewage. Antimicrob. Agents Chemother. 2017, 61, e02229-16. [Google Scholar] [CrossRef]
- Hembach, N.; Schmid, F.; Alexander, J.; Hiller, C.; Rogall, E.T.; Schwartz, T. Occurrence of the mcr-1 colistin resistance gene and other clinically relevant antibiotic resistance genes in microbial populations at different municipal wastewater treatment plants in Germany. Front. Microbiol. 2017, 8, 1282. [Google Scholar] [CrossRef]
- Sun, P.; Bi, Z.; Nilsson, M.; Zheng, B.; Berglund, B.; Stålsby Lundborg, C.; Börjesson, S.; Li, X.; Chen, B.; Yin, H.; et al. Occurrence of bla(KPC-2), bla(CTX-M), and mcr-1 in Enterobacteriaceae from Well Water in Rural China. Antimicrob. Agents Chemother. 2017, 61, e02569-16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, J.; Chen, L.; Yassin, A.K.; Kelly, P.; Butaye, P.; Li, J.; Gong, J.; Cattley, R.; Qi, K.; et al. Housefly (Musca domestica) and blow fly (Protophormia terraenovae) as vectors of bacteria carrying colistin resistance genes. Appl. Environ. Microbiol. 2018, 84, e01736-17. [Google Scholar] [CrossRef] [PubMed]
- Bean, D.C.; Wigmore, S.M.; Abdul Momin, M.H.F.; Wareham, D.W. Polymyxin resistant bacteria in Australian poultry. Front. Sustain. Food Syst. 2020, 4, 550318. [Google Scholar] [CrossRef]
- Yoon, E.J.; Hong, J.S.; Yang, J.W.; Lee, K.J.; Lee, H.; Jeong, S.H. Detection of mcr-1 plasmids in Enterobacteriaceae isolates from human specimens: Comparison with those in Escherichia coli isolates from livestock in Korea. Ann. Lab. Med. 2018, 38, 555–562. [Google Scholar] [CrossRef]
- Zeng, K.J.; Doi, Y.; Patil, S.; Huang, X.; Tian, G.B. Emergence of the plasmid-mediated mcr-1 gene in colistin-resistant Enterobacter aerogenes and Enterobacter cloacae. Antimicrob. Agents Chemother. 2016, 60, 3862–3863. [Google Scholar] [CrossRef]
- Liu, B.T.; Song, F.J.; Zou, M.; Hao, Z.H.; Shan, H. Emergence of colistin resistance gene mcr-1 in Cronobacter sakazakii producing NDM-9 and in Escherichia coli from the same animal. Antimicrob. Agents Chemother. 2017, 61, 01444-16. [Google Scholar] [CrossRef]
- Li, X.P.; Fang, L.X.; Jiang, P.; Pan, D.; Xia, J.; Liao, X.P.; Liu, Y.H.; Sun, J. Emergence of the colistin resistance gene mcr-1 in Citrobacter freundii. Int. J. Antimicrob. Agents 2017, 49, 786–787. [Google Scholar] [CrossRef] [PubMed]
- Mendes, A.C.; Novais, Â.; Campos, J.; Rodrigues, C.; Santos, C.; Antunes, P.; Ramos, H.; Peixe, L. mcr-1 in carbapenemase-producing Klebsiella pneumoniae with hospitalized patients, Portugal, 2016–2017. Emerg. Infect. Dis. 2018, 24, 762–766. [Google Scholar] [CrossRef]
- Yi, L.; Wang, J.; Gao, Y.; Liu, Y.; Doi, Y.; Wu, R.; Zeng, Z.; Liang, Z.; Liu, J.H. mcr-1-harboring Salmonella enterica serovar Typhimurium sequence type 34 in pigs, China. Emerg. Infect. Dis. 2017, 23, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Huang, Y.; Wang, J.; Xu, X.; Hawkey, J.; Yang, C.; Liang, B.; Hu, X.; Wu, F.; Yang, X.; et al. Multidrug-resistant Shigella sonnei carrying the plasmid-mediated mcr-1 gene in China. Int. J. Antimicrob. Agents 2018, 52, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Yao, X.; Lv, L.; Doi, Y.; Huang, X.; Huang, S.; Liu, J.H. Emergence of mcr-1 in Raoultella ornithinolytica and Escherichia coli isolates from retail vegetables in China. Antimicrob. Agents Chemother. 2017, 61, e01139-17. [Google Scholar] [CrossRef] [PubMed]
- Ellem, J.A.; Ginn, A.N.; Chen, S.C.; Ferguson, J.; Partridge, S.R.; Iredell, J.R. Locally acquired mcr-1 in Escherichia coli, Australia, 2011 and 2013. Emerg. Infect. Dis. 2017, 23, 1160–1163. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.M.; Lubian, A.F.; Partridge, S.R.; Gottlieb, T.; Iredell, J.; Daley, D.A.; Coombs, G.W. Australian Group on Antimicrobial Resistance (AGAR) Australian Gram-negative Sepsis Outcome Programme (GnSOP) Annual Report 2020. Commun. Dis. Intell. 2022, 46, 1–12. [Google Scholar] [CrossRef]
- Arnott, A.; Wang, Q.; Bachmann, N.; Sadsad, R.; Biswas, C.; Sotomayor, C.; Howard, P.; Rockett, R.; Wiklendt, A.; Iredell, J.R.; et al. Multidrug-resistant Salmonella enterica 4,[5],12:i:- Sequence Type 34, New South Wales, Australia, 2016–2017. Emerg. Infect. Dis. 2018, 24, 751. [Google Scholar] [CrossRef]
- Ingle, D.J.; Ambrose, R.L.; Baines, S.L.; Duchene, S.; Gonçalves da Silva, A.; Lee, D.Y.J.; Jones, M.; Valcanis, M.; Taiaroa, G.; Ballard, S.A.; et al. Evolutionary dynamics of multidrug resistant Salmonella enterica serovar 4,[5],12:i:- in Australia. Nat. Commun. 2021, 12, 4786. [Google Scholar] [CrossRef]
- Sun, J.; Fang, L.X.; Wu, Z.; Deng, H.; Yang, R.S.; Li, X.P.; Li, S.M.; Liao, X.P.; Feng, Y.; Liu, Y.H. Genetic analysis of the IncX4 plasmids: Implications for a unique pattern in the mcr-1 acquisition. Sci. Rep. 2017, 7, 424. [Google Scholar] [CrossRef]
- Martins-Sorenson, N.; Snesrud, E.; Xavier, D.E.; Cacci, L.C.; Iavarone, A.T.; McGann, P.; Riley, L.W.; Moreira, B.M. A novel plasmid-encoded mcr-4.3 gene in a colistin-resistant Acinetobacter baumannii clinical strain. J. Antimicrob. Chemother. 2020, 75, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Kumar, S. On estimating evolutionary probabilities of population variants. BMC Evol. Biol. 2019, 19, 133. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Tamura, K.; Tao, Q.; Kumar, S. Theoretical Foundation of the RelTime method for estimating divergence times from variable evolutionary rates. Mol. Biol. Evol. 2018, 35, 1770–1782. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, S.; Fillol-Salom, A.; Quiles-Puchalt, N.; Ibarra-Chávez, R.; Haag, A.F.; Chen, J.; Penadés, J.R. Bacterial chromosomal mobility via lateral transduction exceeds that of classical mobile genetic elements. Nat. Commun. 2021, 12, 6509. [Google Scholar] [CrossRef] [PubMed]
- Strepis, N.; Voor In ‘t Holt, A.F.; Vos, M.C.; Zandijk, W.H.A.; Heikema, A.P.; Hays, J.P.; Severin, J.A.; Klaassen, C.H.W. Genetic analysis of mcr-1-carrying plasmids from Gram-negative bacteria in a Dutch tertiary care hospital: Evidence for intrapatient and interspecies transmission events. Front. Microbiol. 2021, 12, 727435. [Google Scholar] [CrossRef]
- Goodman, R.N.; Tansirichaiya, S.; Brouwer, M.S.M.; Roberts, A.P. Intracellular transposition of mobile genetic elements associated with the colistin resistance gene mcr-1. Microbiol. Spectr. 2023, 11, e0327822. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, J.; Li, J.; Ding, Y.; Li, X.P.; Lin, J.; Hassan, B.; Feng, Y. Expanding landscapes of the diversified mcr-1-bearing plasmid reservoirs. Microbiome 2017, 5, 70. [Google Scholar] [CrossRef]
- Sellera, F.P.; Fernandes, M.R.; Sartori, L.; Carvalho, M.P.; Esposito, F.; Nascimento, C.L.; Dutra, G.H.; Mamizuka, E.M.; Pérez-Chaparro, P.J.; McCulloch, J.A.; et al. Escherichia coli carrying IncX4 plasmid-mediated mcr-1 and blaCTX-M genes in infected migratory Magellanic penguins (Spheniscus magellanicus). J. Antimicrob. Chemother. 2017, 72, 1255–1256. [Google Scholar] [CrossRef]
- Maluta, R.P.; Logue, C.M.; Casas, M.R.; Meng, T.; Guastalli, E.A.; Rojas, T.C.; Montelli, A.C.; Sadatsune, T.; de Carvalho Ramos, M.; Nolan, L.K.; et al. Overlapped sequence types (STs) and serogroups of avian pathogenic (APEC) and human extra-intestinal pathogenic (ExPEC) Escherichia coli isolated in Brazil. PLoS ONE 2014, 9, e105016. [Google Scholar] [CrossRef]
- Mshana, S.E.; Imirzalioglu, C.; Hain, T.; Domann, E.; Lyamuya, E.F.; Chakraborty, T. Multiple ST clonal complexes, with a predominance of ST131, of Escherichia coli harbouring blaCTX-M-15 in a tertiary hospital in Tanzania. Clin. Microbiol. Infect. 2011, 17, 1279–1282. [Google Scholar] [CrossRef]
- Guenther, S.; Falgenhauer, L.; Semmler, T.; Imirzalioglu, C.; Chakraborty, T.; Roesler, U.; Roschanski, N. Environmental emission of multiresistant Escherichia coli carrying the colistin resistance gene mcr-1 from German swine farms. J. Antimicrob. Chemother. 2017, 72, 1289–1292. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, R.; Li, J.; Wu, Z.; Yin, W.; Schwarz, S.; Tyrrell, J.M.; Zheng, Y.; Wang, S.; Shen, Z.; et al. Comprehensive resistome analysis reveals the prevalence of NDM and MCR-1 in Chinese poultry production. Nat. Microbiol. 2017, 2, 16260. [Google Scholar] [CrossRef] [PubMed]
- El Garch, F.; Sauget, M.; Hocquet, D.; LeChaudee, D.; Woehrle, F.; Bertrand, X. mcr-1 is borne by highly diverse Escherichia coli isolates since 2004 in food-producing animals in Europe. Clin. Microbiol. Infect. 2017, 23, 51.e51–51.e54. [Google Scholar] [CrossRef] [PubMed]
- Boueroy, P.; Wongsurawat, T.; Jenjaroenpun, P.; Chopjitt, P.; Hatrongjit, R.; Jittapalapong, S.; Kerdsin, A. Plasmidome in mcr-1 harboring carbapenem-resistant Enterobacterales isolates from human in Thailand. Sci. Rep. 2022, 12, 19051. [Google Scholar] [CrossRef]
- Wang, R.; van Dorp, L.; Shaw, L.P.; Bradley, P.; Wang, Q.; Wang, X.; Jin, L.; Zhang, Q.; Liu, Y.; Rieux, A.; et al. The global distribution and spread of the mobilized colistin resistance gene mcr-1. Nat. Commun. 2018, 9, 1179. [Google Scholar] [CrossRef] [PubMed]
- Matamoros, S.; van Hattem, J.M.; Arcilla, M.S.; Willemse, N.; Melles, D.C.; Penders, J.; Vinh, T.N.; Thi Hoa, N.; Bootsma, M.C.J.; van Genderen, P.J.; et al. Global phylogenetic analysis of Escherichia coli and plasmids carrying the mcr-1 gene indicates bacterial diversity but plasmid restriction. Sci. Rep. 2017, 7, 15364. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, J.; Ding, Y.; Li, X.P.; Liu, Y.H.; Feng, Y. Genomic insights into mcr-1-positive plasmids carried by colistin-resistant Escherichia coli isolates from inpatients. Antimicrob. Agents Chemother. 2017, 61, e00361-17. [Google Scholar] [CrossRef]
- Tegetmeyer, H.E.; Jones, S.C.; Langford, P.R.; Baltes, N. ISApl1, a novel insertion element of Actinobacillus pleuropneumoniae, prevents ApxIV-based serological detection of serotype 7 strain AP76. Vet. Microbiol. 2008, 128, 342–353. [Google Scholar] [CrossRef]
- Geurts, A.M.; Hackett, C.S.; Bell, J.B.; Bergemann, T.L.; Collier, L.S.; Carlson, C.M.; Largaespada, D.A.; Hackett, P.B. Structure-based prediction of insertion-site preferences of transposons into chromosomes. Nucleic Acids Res. 2006, 34, 2803–2811. [Google Scholar] [CrossRef]
- Sun, J.; Xu, Y.; Gao, R.; Lin, J.; Wei, W.; Srinivas, S.; Li, D.; Yang, R.S.; Li, X.P.; Liao, X.P.; et al. Deciphering MCR-2 colistin resistance. mBio 2017, 8, e00625-17. [Google Scholar] [CrossRef]
- Xavier, B.B.; Lammens, C.; Ruhal, R.; Kumar-Singh, S.; Butaye, P.; Goossens, H.; Malhotra-Kumar, S. Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. EuroSurveill 2016, 21, 30280. [Google Scholar] [CrossRef] [PubMed]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed]
- Cain, A.K.; Liu, X.; Djordjevic, S.P.; Hall, R.M. Transposons related to Tn1696 in IncHI2 plasmids in multiply antibiotic resistant Salmonella enterica serovar Typhimurium from Australian animals. Microb. Drug Resist. 2010, 16, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Snesrud, E.; McGann, P.; Chandler, M. The birth and demise of the ISApl1-mcr-1-ISApl1 composite transposon: The vehicle for transferable colistin resistance. mBio 2018, 9, e02381-17. [Google Scholar] [CrossRef]
- Li, W.; Yan, Y.; Chen, J.; Sun, R.; Wang, Y.; Wang, T.; Feng, Z.; Peng, K.; Wang, J.; Chen, S.J. Genomic characterization of conjugative plasmids carrying the mcr-1 gene in foodborne and clinical strains of Salmonella and Escherichia coli. Food Control. 2021, 125, 108032. [Google Scholar] [CrossRef]
- Du, C.; Feng, Y.; Wang, G.; Zhang, Z.; Hu, H.; Yu, Y.; Liu, J.; Qiu, L.; Liu, H.; Guo, Z.; et al. Co-occurrence of the mcr-1.1 and mcr-3.7 genes in a multidrug-resistant Escherichia coli isolate from China. Infect. Drug Resist. 2020, 13, 3649–3655. [Google Scholar] [CrossRef] [PubMed]
- He, Y.Z.; Long, T.F.; He, B.; Li, X.P.; Li, G.; Chen, L.; Liao, X.P.; Liu, Y.H.; Sun, J. ISEc69-mediated mobilization of the colistin resistance gene mcr-2 in Escherichia coli. Front. Microbiol. 2020, 11, 564973. [Google Scholar] [CrossRef]
- Li, R.; Xie, M.; Zhang, J.; Yang, Z.; Liu, L.; Liu, X.; Zheng, Z.; Chan, E.W.; Chen, S. Genetic characterization of mcr-1-bearing plasmids to depict molecular mechanisms underlying dissemination of the colistin resistance determinant. J. Antimicrob. Chemother. 2017, 72, 393–401. [Google Scholar] [CrossRef]
- Partridge, S.R. mcr-2 in the IncX4 plasmid pKP37-BE is flanked by directly oriented copies of ISEc69. J. Antimicrob. Chemother. 2017, 72, 1533–1535. [Google Scholar] [CrossRef]
- Li, R.; Du, P.; Zhang, P.; Li, Y.; Yang, X.; Wang, Z.; Wang, J.; Bai, L. Comprehensive genomic investigation of coevolution of mcr genes in Escherichia coli strains via nanopore sequencing. Glob. Chall. 2021, 5, 2000014. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.C.; Li, R.B.; Yang, Y.; Liao, X.P. Sporadic dissemination of mcr-8-ST11 Klebsiella pneumoniae isolates in China. Enferm. Infecc. Microbiol. Clin. 2022, 40, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Qiao, J.; Xu, H.; Liu, R.; Chen, R.; Li, C.; Hu, X.; Zhou, J.; Guo, X.; Zheng, B. First report of Klebsiella pneumoniae co-producing OXA-181, CTX-M-55, and MCR-8 isolated from the patient with bacteremia. Front. Microbiol. 2022, 13, 1020500. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wu, Y.; Fang, Y.; Sang, Z.; Huang, L.; Dong, N.; Zeng, Y.; Lu, J.; Zhang, R.; Chen, G. Emergence of an ST1326 (CG258) multi-drug resistant Klebsiella pneumoniae co-harboring mcr-8.2, ESBL genes, and the resistance-nodulation-division efflux pump gene cluster tmexCD1-toprJ1 in China. Front. Microbiol. 2022, 13, 800993. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Wang, X.; Yu, H.; Zhang, J.; Wang, J.; Li, J.; Li, X.; Hu, K.; Gong, X.; Gou, X.; et al. Molecular antibiotic resistance mechanisms and co-transmission of the mcr-9 and metallo-β-lactamase genes in carbapenem-resistant Enterobacter cloacae complex. Front. Microbiol. 2022, 13, 1032833. [Google Scholar] [CrossRef]
- Liu, M.C.; Jian, Z.; Liu, W.; Li, J.; Pei, N. One healthaAnalysis of mcr-carrying plasmids and emergence of mcr-10.1 in three species of Klebsiella recovered from humans in China. Microbiol. Spectr. 2022, 10, e0230622. [Google Scholar] [CrossRef] [PubMed]
- Abdul Momin, M.H.F.; Bean, D.C.; Hendriksen, R.S.; Haenni, M.; Phee, L.M.; Wareham, D.W. CHROMagar COL-APSE: A selective bacterial culture medium for the isolation and differentiation of colistin-resistant Gram-negative pathogens. J. Med. Microbiol. 2017, 66, 1554–1561. [Google Scholar] [CrossRef] [PubMed]
- Przybysz, S.M.; Correa-Martinez, C.; Köck, R.; Becker, K.; Schaumburg, F. SuperPolymyxin™ medium for the screening of colistin-resistant gram-negative bacteria in stool samples. Front. Microbiol. 2018, 9, 2809. [Google Scholar] [CrossRef] [PubMed]
- Bardet, L.; Le Page, S.; Leangapichart, T.; Rolain, J.M. LBJMR medium: A new polyvalent culture medium for isolating and selecting vancomycin and colistin-resistant bacteria. BMC Microbiol. 2017, 17, 220. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, Y.; Liu, C.; Kudinha, T.; Liu, X.; Luo, Y.; Yang, Q.; Sun, H.; Hu, J.; Xu, Y.C. Comparison of five commonly used automated susceptibility testing methods for accuracy in the China Antimicrobial Resistance Surveillance System (CARSS) hospitals. Infect. Drug Resist. 2018, 11, 1347–1358. [Google Scholar] [CrossRef]
- Cordovana, M.; Ambretti, S. Antibiotic susceptibility testing of anaerobic bacteria by broth microdilution method using the MICRONAUT-S Anaerobes MIC plates. Anaerobe 2020, 63, 102217. [Google Scholar] [CrossRef]
- Carretto, E.; Brovarone, F.; Russello, G.; Nardini, P.; El-Bouseary, M.M.; Aboklaish, A.F.; Walsh, T.R.; Tyrrell, J.M. Clinical validation of SensiTest colistin, a broth microdilution-based nethod to evaluate colistin MICs. J. Clin. Microbiol. 2018, 56, e01523-17. [Google Scholar] [CrossRef]
- Poirel, L.; Larpin, Y.; Dobias, J.; Stephan, R.; Decousser, J.W.; Madec, J.Y.; Nordmann, P. Rapid Polymyxin NP test for the detection of polymyxin resistance mediated by the mcr-1/mcr-2 genes. Diagn. Microbiol. Infect. Dis. 2018, 90, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Jouy, E.; Haenni, M.; Le Devendec, L.; Le Roux, A.; Châtre, P.; Madec, J.Y.; Kempf, I. Improvement in routine detection of colistin resistance in E. coli isolated in veterinary diagnostic laboratories. J. Microbiol. Methods 2017, 132, 125–127. [Google Scholar] [CrossRef] [PubMed]
- Coppi, M.; Cannatelli, A.; Antonelli, A.; Baccani, I.; Di Pilato, V.; Sennati, S.; Giani, T.; Rossolini, G.M. A simple phenotypic method for screening of MCR-1-mediated colistin resistance. Clin. Microbiol. Infect. 2018, 24, 201.e201–201.e203. [Google Scholar] [CrossRef] [PubMed]
- Kon, H.; Dalak, M.A.B.; Schwartz, D.; Carmeli, Y.; Lellouche, J. Evaluation of the MICRONAUT MIC-strip colistin assay for colistin susceptibility testing of carbapenem-resistant Acinetobacter baumannii and Enterobacterales. Diagn. Microbiol. Infect. Dis. 2021, 100, 115391. [Google Scholar] [CrossRef]
- Bardet, L.; Okdah, L.; Le Page, S.; Baron, S.A.; Rolain, J.M. Comparative evaluation of the UMIC Colistine kit to assess MIC of colistin of gram-negative rods. BMC Microbiol. 2019, 19, 60. [Google Scholar] [CrossRef] [PubMed]
- Sękowska, A.; Bogiel, T. The Evaluation of Eazyplex® SuperBug CRE assay usefulness for the detection of ESBLs and carbapenemases genes directly from urine samples and positive blood cultures. Antibiotics 2022, 11, 138. [Google Scholar] [CrossRef]
- Chabou, S.; Leangapichart, T.; Okdah, L.; Le Page, S.; Hadjadj, L.; Rolain, J.M. Real-time quantitative PCR assay with Taqman® probe for rapid detection of MCR-1 plasmid-mediated colistin resistance. New Microbes New Infect. 2016, 13, 71–74. [Google Scholar] [CrossRef]
- Zhong, L.L.; Zhou, Q.; Tan, C.Y.; Roberts, A.P.; El-Sayed Ahmed, M.A.E.; Chen, G.; Dai, M.; Yang, F.; Xia, Y.; Liao, K.; et al. Multiplex loop-mediated isothermal amplification (multi-LAMP) assay for rapid detection of mcr-1 to mcr-5 in colistin-resistant bacteria. Infect. Drug Resist. 2019, 12, 1877–1887. [Google Scholar] [CrossRef]
- Borowiak, M.; Baumann, B.; Fischer, J.; Thomas, K.; Deneke, C.; Hammerl, J.A.; Szabo, I.; Malorny, B. Development of a novel mcr-6 to mcr-9 multiplex PCR and assessment of mcr-1 to mcr-9 occurrence in colistin-resistant Salmonella enterica isolates from environment, feed, animals and food (2011–2018) in Germany. Front. Microbiol. 2020, 11, 80. [Google Scholar] [CrossRef]
- Li, J.; Shi, X.; Yin, W.; Wang, Y.; Shen, Z.; Ding, S.; Wang, S. A multiplex SYBR green real-time PCR assay for the detection of three colistin resistance genes from cultured bacteria, feces, and environment samples. Front. Microbiol. 2017, 8, 2078. [Google Scholar] [CrossRef]
- Neumann, B.; Rackwitz, W.; Hunfeld, K.P.; Fuchs, S.; Werner, G.; Pfeifer, Y. Genome sequences of two clinical Escherichia coli isolates harboring the novel colistin-resistance gene variants mcr-1.26 and mcr-1.27. Gut Pathog. 2020, 12, 40. [Google Scholar] [CrossRef]
- Nicolas, I.; Bordeau, V.; Bondon, A.; Baudy-Floc’h, M.; Felden, B. Novel antibiotics effective against gram-positive and -negative multi-resistant bacteria with limited resistance. PLoS Biol. 2019, 17, e3000337. [Google Scholar] [CrossRef]
- Flament-Simon, S.C.; de Toro, M.; Mora, A.; García, V.; García-Meniño, I.; Díaz-Jiménez, D.; Herrera, A.; Blanco, J. Whole genome sequencing and characteristics of mcr-1-harboring plasmids of porcine Escherichia coli isolates belonging to the high-risk clone O25b:H4-ST131 clade B. Front. Microbiol. 2020, 11, 387. [Google Scholar] [CrossRef]






| mcr Gene Number | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| mcr gene number and source | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| mcr-1 Escherichia coli KU886144.1 | 0.18 | 0.67 | 0.57 | 0.54 | 0.22 | 0.47 | 0.68 | 0.71 | 0.71 | |
| mcr-2 Pseudomonas aeruginosa MW811418.1 | 0.18 | 0.68 | 0.58 | 0.56 | 0.12 | 0.49 | 0.69 | 0.7 | 0.72 | |
| mcr-3 Escherichia coli MW811424.1 | 0.67 | 0.68 | 0.62 | 0.75 | 0.68 | 0.7 | 0.76 | 0.38 | 0.38 | |
| mcr-4 Escherichia coli MW811433.1 | 0.57 | 0.58 | 0.62 | 0.56 | 0.58 | 0.49 | 0.65 | 0.65 | 0.61 | |
| mcr-5.1 Salmonella enterica NG055658.1 | 0.54 | 0.56 | 0.75 | 0.56 | 0.55 | 0.43 | 0.64 | 0.72 | 0.73 | |
| mcr-6.1 Moraxella sp. NG055781.1 | 0.22 | 0.12 | 0.68 | 0.58 | 0.55 | 0.51 | 0.72 | 0.72 | 0.74 | |
| mcr-7 Pseudomonas aeruginosa MW811434.1 | 0.47 | 0.49 | 0.7 | 0.49 | 0.43 | 0.51 | 0.65 | 0.71 | 0.68 | |
| mcr-8 Klebsiella pneumoniae MT815555.1 | 0.68 | 0.69 | 0.76 | 0.65 | 0.64 | 0.72 | 0.65 | 0.69 | 0.72 | |
| mcr-9 Uncultured bacterium MW478857.1 | 0.71 | 0.7 | 0.38 | 0.65 | 0.72 | 0.72 | 0.71 | 0.69 | 0.22 | |
| mcr-10.1 Enterobacter cloacae MN044989.1 | 0.71 | 0.72 | 0.38 | 0.61 | 0.73 | 0.74 | 0.68 | 0.72 | 0.22 | |
| Average evolutionary divergence | 0.53 | 0.52 | 0.62 | 0.59 | 0.61 | 0.54 | 0.57 | 0.69 | 0.61 | 0.61 |
| Standard Deviation | 0.20 | 0.23 | 0.14 | 0.05 | 0.11 | 0.23 | 0.11 | 0.04 | 0.18 | 0.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahzad, S.; Willcox, M.D.P.; Rayamajhee, B. A Review of Resistance to Polymyxins and Evolving Mobile Colistin Resistance Gene (mcr) among Pathogens of Clinical Significance. Antibiotics 2023, 12, 1597. https://doi.org/10.3390/antibiotics12111597
Shahzad S, Willcox MDP, Rayamajhee B. A Review of Resistance to Polymyxins and Evolving Mobile Colistin Resistance Gene (mcr) among Pathogens of Clinical Significance. Antibiotics. 2023; 12(11):1597. https://doi.org/10.3390/antibiotics12111597
Chicago/Turabian StyleShahzad, Shakeel, Mark D. P. Willcox, and Binod Rayamajhee. 2023. "A Review of Resistance to Polymyxins and Evolving Mobile Colistin Resistance Gene (mcr) among Pathogens of Clinical Significance" Antibiotics 12, no. 11: 1597. https://doi.org/10.3390/antibiotics12111597