1. Introduction
In the past two decades, nosocomial infections have become a significant public health problem worldwide [
1], representing a healthcare failure that leads to adverse effects during in-hospital stays and an increased risk of mortality among patients [
2]. These infections are associated with various medical procedures such as invasive interventions, extensive surgeries and the use of indwelling medical devices, giving rise to central-line-associated bloodstream infections (CLABSIs), catheter-associated urinary-tract infections (CAUTIs), surgical-site infections (SSIs) and ventilator-associated pneumonia (VAP) [
3].
Factors such as prolonged hospital stays also contribute to the acquisition of bloodstream infections not associated with central catheter use, gastrointestinal infections (GIs), urinary-tract infections (UTIs) not associated with catheter use and non-ventilator-associated hospital-acquired pneumonia (NV-HAP). Various body sites are affected by healthcare-associated infections (HAIs), including the ear, eye, nose, throat, lower respiratory tract, skin, soft tissue, cardiovascular system, bone, joints, central nervous system and reproductive tract. HAIs have diverse causes, including bacterial, viral or fungal agents. Among the main bacteria responsible for these infections are Gram-positive microorganisms like coagulase-negative Staphylococci,
Staphylococcus aureus, Streptococcus species, Enterococcus faecalis, Enterococcus faecium and
Clostridium difficile [
4], as well as Gram-negative microorganisms such as some members of the Enterobacteriaceae family (e.g.,
Escherichia coli, Klebsiella pneumoniae, Klebsiella oxytoca, Proteus mirabilis), Enterobacter species (
Pseudomonas aeruginosa, Acinetobacter baumanii and
Burkholderia cepacian). These pathogens are associated with high morbidity and mortality rates in hospitalized patients due to their inherent multi-drug-resistant properties [
5].
The extensive use of antibiotics in hospitals exerts significant selection pressure on nosocomial bacterial populations, especially on bacteria like
Escherichia coli, Acinetobacter baumanii, Pseudomonas aeruginosa, Enterococcus faecium, Proteus mirabilis and
Stenotrophomonas maltophilia, leading to the emergence of resistant, multi-resistant and pan-resistant nosocomial infections [
6].
Escherichia coli, a pathogenic microorganism, can be found in various environmental sources and the intestines of humans and animals. While most strains are harmless, certain pathogenic strains have been associated with gastrointestinal diseases and extraintestinal tissue infections. In hospitals,
Escherichia coli has been identified as the causative agent of various healthcare-associated infections like BSI, CLABSI, UTI, CAUTI, SSI, VAP, NV-HAP and GI [
7].
Due to excessive and sometimes indiscriminate antibiotic use,
Escherichia coli strains in hospitals have become highly multi-drug-resistant (MDR). This resistance is particularly common in beta-lactam antibiotics, severely limiting effective treatment options [
8]. The resistance to beta-lactam antibiotics is encoded by different classes of beta-lactamases, including
blaCTX,
blaVIM,
blaNMC,
blaOXA,
blaIMP,
blaLAP,
blaROB,
blaDHA,
blaVEB,
blaCMY,
blaLAP,
blaSHV,
blaTEM,
blaSME,
blaSFC,
blaKPC,
blaAmpC,
blaGES,
blaSME,
blaPER and
blaNDM [
9,
10,
11,
12,
13,
14,
15,
16,
17,
18], which confer a wide spectrum of resistance to beta-lactam antibiotics [
19].
In this study, we focus on characterizing the beta-lactam resistome expressed by Escherichia coli-causing nosocomial infections in the “October first” Regional Hospital of ISSSTE in Mexico.
2. Materials and Methods
2.1. Primer Design
The bioinformatic analysis and primer design process were previously described and reported by our research group [
20]. In summary, we retrieved beta-lactamase integron and DNA sequences from the GenBank of NCBI. All available sequences of beta-lactamases were gathered for the purpose of PCR primer design. The DNA sequences were aligned using ClustalW v.2 software [
21]. Subsequently, we used FigTree V1.4.0 software [
22] to generate phylogenetic trees based on the alignments. From the alignment results, conserved sequences were identified and used as a basis for designing a set of specific and degenerate primers. The PerPrimer v1.1.21 Software [
23] was utilized, employing stringent criteria, including primer length (18–25 bp), a melting temperature (Tm) of 60–62
C, a GC content of 40–60%, with a
T
of 1
C in the annealing temperature and an amplicon size ranging from 83 to 230 bp [
20] (
Table 1).
2.2. Biological Samples
A total of five-hundred-and-eighty-five clinical samples were collected from various secretions, including urethral, vaginal, nasal, diabetic foot ulcers, bronchial, surgical and pharyngeal wound secretions. Additionally, body fluids such as blood, dialysis fluid, cerebrospinal fluid, peritoneal, pleural and synovial fluid, urine, feces, sputum and semen, as well as catheter tips from patients diagnosed with nosocomial infections, were also included in the study. Among these samples, 218 Escherichia coli strains showing multi-drug resistance were cultured and selected for further analysis. The biochemical identification of these strains was performed by the Microbiology section of the Central Clinical Laboratory at the “October first” Regional Hospital (ISSSTE).
2.3. Escherichia coli Isolation Culture
This study was conducted at the “October first” Regional Hospital (ISSSTE) in Mexico City. The collected samples were cultured on various agar media, including Blood Agar (COS), MacConkey Agar, Chocolate Mueller Hilton Agar (PVX), Chromogenic Candida Agar, Sabouraud Agar, Triptone Soya Broth, Chromogenic Agar for Staphylococcus aureus identification in human specimens (SAID), Salted Mannitol Agar, Chromogenic Agar for the detection of Salmonella, SS Agar for Shigella and Salmonella and CromlDTM CPS Agar.
The cultured plates were incubated at 35 C for 18 to 24 h. All bacterial growth on the MacConkey agar plates was subjected to biochemical identification tests. However, only the cultures that were identified as Escherichia coli were further selected for antimicrobial susceptibility testing and genotyping.
2.4. Biochemical Identification
The biochemical identification of Escherichia coli strains was carried out using the VITEK 2XL system, an automated method provided by bioMérieux Inc. (Durham, NC, USA) Specifically, the VITEK 2XL GN 21341 cards were used, containing 21 biochemical tests. These tests included -galactosidase (OPNG), arginine dehydrolase (ADH), lysine decarboxylase (LDC), ornithine decarboxylase (ODC), citrate utilization (CIT), hydrogen sulfide (H2S) production, urease (URE) activity, tryptophan deaminase (TDA) production, indole production (IND), Voges–Proskauer acetoin production (VP), gelatinase (GEL) activity and fermentation/oxidation tests for various sugars such as glucose (GLU), mannitol, inositol, sorbitol, ramnose, sucrose, melibiose, amygdalin (vitamin B17), arabinose and cytochrome oxidase (OX) activity.
To confirm the Escherichia coli identification, the strains must test positive for -galactosidase (OPNG), lysine decarboxylase (LDL), ornithine decarboxylase (ODC), indole production (IND), glucose fermentation (GLU), mannitol (MAN), sorbitol (SOR), ramnose (RHA), sucrose (SAC), melibiose (MEL) and arabinose (ARA) tests among the set of biochemical tests conducted using the VITEK 2XL GN 21341 cards.
2.5. Antimicrobial Susceptibility Testing
Antibiotic susceptibility testing was performed using the VITEK 2XL automated system with ASTGN70 cards (REF 413 and 401) specifically designed for Gram-negative bacteria by BioMérieux Inc.(Durham, NC, USA) The following antibiotics were tested with varying concentrations (in g/mL): amikacin (8, 16, 64 g/mL), ampicillin (4, 8, 32 g/mL), ampicillin/sulbactam (4/2, 16/8, 32/16 g/mL), aztreonam (2, 8, 32 g/mL), cefazolin (4, 16, 64 g/mL), cefepime (2, 8, 16, 32 g/mL), cefotetan (2, 8, 32 g/mL), ceftazidime (1, 2, 8, 32 g/mL), ceftriaxone (1, 2, 8, 32 g/mL), cefuroxime (2, 8, 32 g/mL), ciprofloxacin (0.5, 2, 4 g/mL), gentamicin (4, 16, 32 g/mL), imipenem (2, 4, 16g/mL), meropenem (0.5, 4, 16 g/mL), piperacillin/tazobactam (4/4, 16/4, 128/4 g/mL), ticarcillin/clavulanic acid (8/2, 32/2, 64/2 g/mL), tobramycin (8, 16, 64 g/mL), trimethoprim/sulfamethoxazole (0.5/9.5, 2/38, 16/304 g/mL), nitrofurantoin (16, 32, 64 g/mL) and gatifloxacin (1, 4, 8 g/mL).
The resistance or susceptibility profile of each strain was determined by measuring the minimum inhibitory concentration (MIC) of the antibiotics, which was assessed by measuring the optical density of bacterial cultures after incubation.
2.6. Beta-Lactam Selection
For genomic and plasmid DNA sequencing, it was necessary to obtain generic cultures of isolated pure Escherichia coli colonies. The growth of each colony was achieved by placing it in 10 mL of 25% lysogeny broth (LB) medium, which was then sterilized at 121 C for 15 min. Subsequently, each Escherichia coli strain was grown in a set of beta-lactam drugs, including ampicillin (10 g), ampicillin/sulbactam (10/10 g), carbenicillin (100 g), cefaclor (30 g), cefazolin (30 g), cefepime (30 g), cefoperazone (75 g), cefotetan (30 g), mezlocillin (75 g) and piperacillin/tazobactam (100/10 g). All of these beta-lactam drugs were obtained from Becton Dickinson (Franklin Lakes, NJ, USA). Each medium was incubated at 37 C for 24 h under aerobic conditions. The inoculum was adjusted using a Mac Farland 0.5 reading, which corresponds to an expected colony-forming units per milliliter (CFU/mL) of approximately 1.5 × 108. The cultures were then placed in a Shaker Series Innova at 37 C and 240 rpm for 20 h.
2.7. DNA Extraction
2.7.1. Crude Extract
Total DNA was extracted using a modified version of the heat-shock method described by Maugeri et al. One milliliter of Luria medium containing a well-identified, multi-resistant Escherichia coli strain grown for 20 h at 35 C was used. The selected medium was lysed by heating it in a Thermoblock for 1 min at 100 C, followed by immediate placement in an ice bath for 5 min, inducing thermal shock and facilitating the release of genetic material. Two milliliters of the cell lysates containing DNA were purified using chelex (glass beads). The purified DNA served as a template for both real-time PCR assays and NGS (next-generation sequencing). The extracted DNA was stored at −80 C for further analysis and experimentation.
2.7.2. Plasmid DNA Extraction
Plasmid DNA extraction was conducted using a previously reported method [
20]. The PureLink HiPure Plasmid DNA Purification Kit (Invitrogen, Carlsbad, CA, USA) was employed, following the manufacturer’s instructions. The extracted plasmid DNA solution was quantified using a Qubit Flex Fluorometer (Thermo Fisher Scientific, Wilmington, DE, USA) based on fluorometry. To verify the integrity of the plasmid DNA, electrophoresis was performed in 2% agarose gel. Subsequently, the samples were stored at −80
C until further use.
2.8. Real-Time PCR
The beta-lactamases SYBR Green-based qPCR assays were performed on the Real-Time HT FAST 7900 system (Applied Biosystems, Waltham, MA, USA). Each reaction was conducted in a total volume of 10 L, comprising 5 L of 21.5 × SYBR Green PCR Master Mix (Applied Biosystems, Waltham, MA, USA), 10 mM of specific or degenerate primers and 3 L of the DNA template. The cycling conditions for both assays consisted of an initial denaturation step at 95 C for 5 min, followed by 45 cycles of denaturation at 95 C for 15 s, annealing at 50 C for 40 s and extension at 60 C for 40 s. To confirm the specificity of the qPCR products, the amplicons were analyzed using melting curves. Additionally, the products were visualized on a 2% agarose gel pre-stained with ethidium bromide and the resulting image was digitized using a GelLogistic 3000 photodocumenter.
2.9. Whole DNA Sequencing
For DNA sequencing, a previously reported method by the research group [
20] was followed. Both genomic and plasmid DNA were mixed in a 1:1 proportion and used to prepare indexed libraries using the Illumina Nextera XT DNA Sample Preparation Kit (FC-131-1096) designed for small genomes. The libraries were then sequenced on the MiSeq platform (Illumina; San Diego, CA, USA). Adapters and barcodes were trimmed using the default settings in the Illumina experiment manager, resulting in 300 bp paired-end reads. The quality of the unprocessed reads was assessed using FastQC High-Throughput Sequence QC Report v:0.11.5 from Babraham Bioinformatics, Babraham Institute (Cambridge, UK) [
24]. A minimum quality score (Q score) of more than 30 was achieved for at least 85% of all reads. For read mapping, the BWA-MEM aligner version 0.7.7-r441 from the Wellcome Trust Sanger Institute (Hinxton, UK) was used. Genome assembly was performed using the SPAdes Genome Assembler software version 3.11 from CAB, St. Petersburg State University (St. Petersburg, Russia). The reported ASM74325v1 genome of Escherichia coli strain 25922 ATCC with a size of 5.2 Mbp was used as the assembly reference [
25]. The DNA metagenomic analysis for the taxonomic classification of the Escherichia coli bacteria was conducted using the Kraken taxonomic sequence classification system, version 0.10.5-beta, from CCB, Johns Hopkins University (Baltimore, MD, USA) [
26]. For identifying beta-lactamase genes, a comparative analysis was performed using the Basic Local Alignment Search Tool (BLAST) from NCBI-NIH (Bethesda, MD, USA) [
27].
2.10. Study Outcomes
The main objective of this study was to characterize the beta-lactam resistome expressed by Escherichia coli strains causing nosocomial infections. The focus was on understanding the patterns of antibiotic resistance induced by the expression of beta-lactamases, which often complicate the treatment of nosocomial infections. By examining the resistome and identifying the specific beta-lactamase genes responsible for the multi-drug resistance, this study aimed to provide valuable insights for the better management and treatment of nosocomial infections caused by these bacteria.
4. Discussion
Escherichia coli is a Gram-negative bacteria that has coexisted with humans since ancient times. It is one of the best-adapted microorganisms to the human environment and has become a member of the intestinal microbiota. It has a worldwide distribution and is known for its high genetic plasticity, which enables it to easily adapt to different environments. In our research, we found that 22.48% of all multi-resistant
Escherichia coli were cultured from urine, 21.10% from vaginal discharge, 6.42% from catheter tips, 5.96% from blood cultures, 5.50% from feces cultures, 5.05% from sputum, 3.67% from biopsies and 0.92% from cerebrospinal fluid. Additionally, 28.90% were isolated from diverse discharge. These findings are consistent with previous reports where
Escherichia coli was isolated from patients with nosocomial infections, urine infections, gut infections, neonatal meningitis, pneumonia, cholecystitis, peritonitis, osteomyelitis, infectious arthritis and even otitis. Moreover,
Escherichia coli is the most frequent cause of bacteremia [
28,
29,
30,
31,
32,
33,
34,
35,
36,
37].
4.1. Escherichia coli Nosocomial Characterization
Escherichia coli has been identified as a member of the nosocomial bacteriome in hospitals worldwide [
38]. In our study, we determined the nosocomial origin of
Escherichia coli by quantifying the multiple-antibiotic-resistance index (MAR index) using dilution antibiogram data (
Table 3). Our analysis revealed a wide range of antibiotic resistance in the
Escherichia coli strains obtained from in-hospital patients, with 99.08% having an MAR index greater than 0.25 and a mean of 0.58 ± 0.21 for the antibiotics conforming to the general dilution antibiogram (
Table 2). For the beta-lactam selection antibiogram, the MAR index was 0.82 ± 0.09 and for both sets of antibiotics, the MAR index was larger than 4, indicating that all
Escherichia coli strains were isolated from an environment of high antibiotic use, specifically nosocomial sources.
Our findings are consistent with other reports where
Escherichia coli isolated from tertiary hospitals in South-West Nigeria had an MAR index higher than 0.2 [
39]. Similar results were found in
Escherichia coli-causing urine infections in patients from a referral hospital in Eastern Nepal [
40]. The high MAR index is a major health problem caused by uncontrolled antibiotic use worldwide, especially in the nosocomial environment. It has been identified as a risk factor for neonatal mortality and prolonged hospital stays [
41]. Similar results have been published in adult in-hospital patients infected with Gram-negative bacilli that have a high MAR index, with these patients showing a significant increase in mortality, morbidity, length of hospitalization and healthcare costs [
42].
4.2. Escherichia coli Characterization and Genome Size
The metagenomic DNA analysis of sequencing data confirmed that the
Escherichia coli strains obtained from the microbiology section of the clinic laboratory of the “October first” Regional Hospital had a 100% identity (
Table 5). All strains showed at least 60% of all sequencing reads that identified the species, with a mean of 85% of sequencing reads. These findings are consistent with previous reports by our research group [
20].
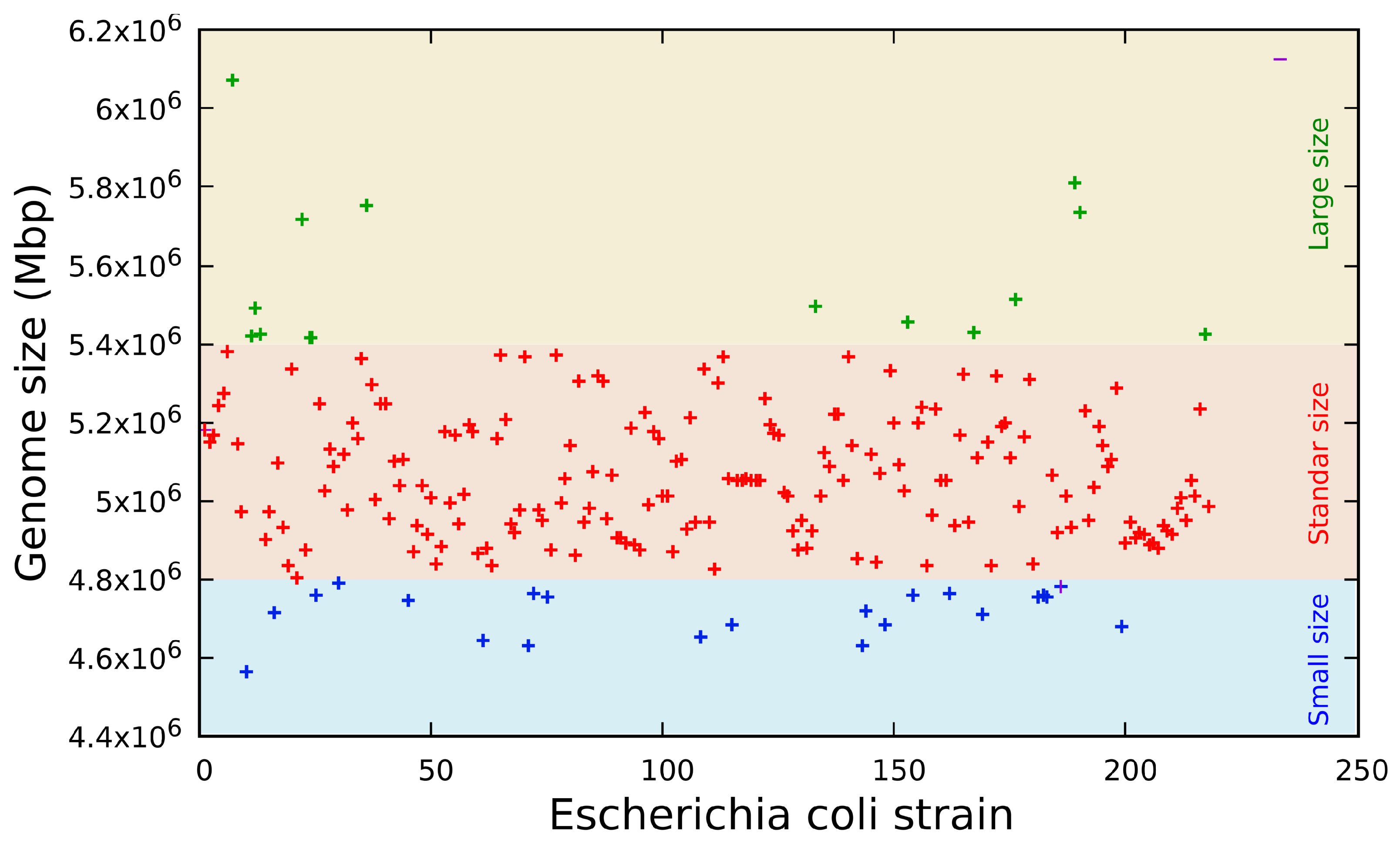
The genome sequence revealed three distinct groups (
Figure 1) classified by size in base pairs: a short genome size composed of sequences from 4.56 to 4.83 Mbp (
n = 10, 4.59%), a standard genome size composed of sequences from 4.84 to 5.27 Mbp (
n = 176, 80.73%) and a long genome size composed of sequences from 5.28 to 6.07 Mbp (
n = 32, 14.68%). These genome sizes are consistent with previous data published on the 61 genomes of
Escherichia coli [
43] and the data bank published by the SRA of NCBI [
44].
4.3. Beta-Lactam Gene Families
The isolated
Escherichia coli strains were cultured and selected in the presence of beta-lactam drugs in liquid culture media (
Table 4). All strains were expected to be carriers of at least one member of the different beta-lactamase gene families. Real-time PCR was used to identify the gene families transcribed and the phenotype responsible for beta-lactam resistance. The results showed that 95.87% of the isolated strains were positive for
blaTEM, 71.10% were positive for
blaCTX, 33.94% were positive for
blaSHV, 29.82% were positive for
blaBIL, 19.27% were positive for
blaDHA, 18.81% were positive for
blaCMY, 11.01% were positive for
blaIMP, 6.88% were positive for
blaLAP, 14.59% were positive for
blaP, 4.13% were positive for
blaVIM, 1.82% were positive for
blaCTX and 0.92% were positive for both
blaKPC and
blaCTX (
Table 8). No
Escherichia coli strains positive for members of the
blaOXA and
blaROB gene families were found in all obtained strains from patients with nosocomial infections. In this study, the
blaTEM gene family was detected in 95.87% of the studied
Escherichia coli bacteria from patients diagnosed with nosocomial infection in the“October first” Regional Hospital. This result is consistent with previous reports that found the
blaTEM gene family in 74.5% of all multi-drug-resistant ESBL-producing
Escherichia coli isolates from hospitals in Malaysia [
45]. The seven most frequent gene families that code for beta-lactamases expressed by
Escherichia coli that cause nosocomial infections are
blaTEM,
blaCTX,
blaSHV,
blaBIL,
blaDHA,
blaCMY and
blaIMP, representing 93.85% of the beta-lactamase problem. However, the
blaTEM,
blaCTX and
blaSHV families together represent the greatest problem with regard to resistance to beta-lactam drugs, as they are expressed in 67.38% of all multi-resistant
Escherichia coli strains that cause nosocomial infections in the in-patients of the “October first” Regional Hospital of the ISSSTE. These results are consistent with previous works that found that
Escherichia coli isolated from in-hospital patients expressed different members of the
blaCTX,
blaSHV and
blaTEM gene families [
46,
47,
48,
49,
50,
51]. On the other hand, the gene families
blaBIL,
blaDHA,
blaCMY,
blaIMP,
blaLAP,
blaP,
blaVIM,
blaCTX and
blaKPC represent 32.62% of the beta-lactam antibiotic resistance problem. These results are consistent with previous reports where the expression of the following gene families in
Escherichia coli isolated from in-hospital patients was found:
blaBIL [
52],
blaDHA [
53],
blaCMY [
54],
blaIMP,
blaLAP, which has been associated with quinolone resistance gene expression [
55] and
blaP [
56].
4.4. Beta-Lactamase Genome Family Resistome
Out of the 218
Escherichia coli strains resistant to beta-lactam drugs, isolated from 351 in-hospital patients with nosocomial infections, a total of 52 different combinations of gene families coding for beta-lactamases were identified as contributing to the beta-lactam resistome. Interestingly, 13 of the most frequently occurring combinations accounted for a substantial 72.9% of all beta-lactam resistomes (
Table 6). On the other hand, the remaining 40 less common combinations were responsible just for 27.1% of all multi-resistance to beta-lactam drugs (
Table 7). The common combinations consisted of one to six distinct gene families that encode beta-lactamases (
Table 6), whereas the less common combinations were composed of two to nine different gene families coding for beta-lactamases (
Table 7).
These observations lead us to postulate that the rare combinations represent natural trials in which beta-lactam resistomes are formed by integrating a large number of beta-lactamase gene families. These combinations are likely to be subjected to selection pressure and might potentially evolve into highly efficient resistomes in the future. However, currently, the most successful resistomes are the common ones composed of six or fewer gene families, as they are prevalent in the majority of nosocomial infections caused by multi-resistant Escherichia coli strains.
4.5. Genomic Beta-Lactamase Intrafamilial Variability
4.5.1. blaTEM Gene Family
The sequencing data revealed that 95.87% of all resistant
Escherichia coli strains carried a member of the
blaTEM gene family, with 21.56% of them exhibiting a unique beta-lactam resistance mechanism. In contrast, 74.31% of the strains (
Table 6) showed multiple beta-lactam resistance mechanisms. These findings align with previous reports where the
blaTEM gene family (
n = 61 strains, 100%) was identified as the primary beta-lactam drug resistance mechanism in
Escherichia coli isolated from the river Yamuna [
57]. Characterization of the blaTEM members identified six different alleles of this gene family, including blaTEM-1 expressed in 55.5% of all isolated
Escherichia coli strains, followed by
blaTEM-12 in 13.88%,
blaTEM-2 in 10.05%,
blaTEM-52 in 9.09%,
blaTEM-24 in 7.18% and
blaTEM-10 in 4.31% (
Table 6). These findings are consistent with another study where 24% (
n = 48) of strains obtained from five hospitals in Tehran were found to carry the
blaTEM-1 gene [
58]. The blaTEM-1 enzyme is particularly noteworthy as it hydrolyzes ampicillin at a higher rate compared to carbenicillin, oxacillin and cephalothin but does not hydrolyze extended-spectrum cephalosporins. Furthermore,
blaTEM-1 is considered an ancestral gene for
blaTEM-10 and
blaTEM-12, which are among the most commonly encountered
blaTEM alleles in the United States of America [
59].
Additionally, the
blaTEM-2 allele exhibits similar hydrolytic activity to
blaTEM-1 but possesses greater transcriptional activity [
60]. Another less common beta-lactamase identified in this study includes
blaTEM-24 and
blaTEM-52, both of which have been described in bacteria-causing nosocomial infections in a Tunisian hospital [
61]. These findings are consistent with similar results published by a Korean group from Clinical Isolates of
Escherichia coli expressing the
blaTEM-1,
blaTEM-19,
blaTEM-20 and
blaTEM-52 genes [
62], conferring resistance characteristics to penicillins, aztreonam and cephalosporins. The expression of all these genes has been previously cloned, sequenced and reported in
Escherichia coli strains [
63].
4.5.2. blaCTX Gene Family
The second most frequent gene family identified in our study was
blaCTX (54.72%), which was expressed in six isoforms, distributed across three clusters. Within the
blaCTX-M-1 cluster, we found two members:
blaCTX-M-1, present in 18.06% and
blaCTX-M-15, found in 68.39% of the samples. In the
blaCTX-9 cluster, only the
blaCTX-M-27 was detected in 4.52% of the strains. The
blaCTX-M-25 cluster had the
blaCTX-M-100 isoform as its sole member, accounting for 9.03% of the isolates. Notably, the
blaCTX-M-15 isoform emerged as the most dominant among the clusters. These findings are consistent with results obtained from clinically significant
Escherichia coli isolates in Kuwait Hospitals, where the
blaCTX-M-15 isoform was present in 84.1% of all isolates (
Table 6) [
64]. Additionally, another study conducted in the UK using
Escherichia coli-producing CTX-M extended-spectrum beta-lactamases identified the
blaCTX-M-15 isoform in 95.9% of all studied strains [
65]. In contrast, our study found the blaCTX-M-1 isoform in 18.06% of the samples. Interestingly, previous works published by three university hospitals in Tehran reported that 61.8% of 144 CTX-resistant
Escherichia coli isolates carried the
blaCTX-M-1 gene [
66], conferring resistance characteristics to penicillin, aztreonam and cephalosporin. The expression of all these genes has been previously cloned, sequenced and reported in
Escherichia coli strains [
67].
4.5.3. blaSHV Gene Family
The third most frequently expressed beta-lactamase by
Escherichia coli strains causing nosocomial infections was
blaSHV, accounting for 33.94% of all analyzed multi-resistant strains. Within this gene family, five isoforms were identified, with
blaSHV-12 being the most dominant (78.38%), followed by
blaSHV-1 (9.46%),
blaSHV-100 (5.41%),
blaSHV-121 (4.05%) and
blaSHV-11 (2.70%) (
Table 6). Although all isoforms were expressed at low frequencies,
blaSHV-12 stood out as the most prevalent member, isolated from diverse sources in a recent study conducted in Catalonia, where it was found at a frequency of 23% [
68]. These isoforms confer characteristic resistance to penicillin, aztreonam and cephalosporin. The expression of all these gene isoforms has been previously cloned, sequenced and reported in
Escherichia coli strains [
69].
4.5.4. blaIMP Gene Family
The blaIMP gene family was detected in 11.01% of the strains and it comprises six different members, namely
blaIMP-1 (8.33%;
n = 2),
blaIMP-2 (4.17%;
n = 1),
blaIMP-4 (4.17%;
n = 1),
blaIMP-6 (4.17%,
n = 1),
blaIMP-11 (8.33%;
n = 2) and the dominant gene
blaIMP-14 (8.33%;
n = 17). These genes confer characteristic resistance to imipenem and carbapenem [
55]. The expression of all these genes has been previously cloned and sequenced from
Escherichia coli strains [
70].
4.5.5. blaCTX-M, blaVIM and blaP Gene Family
All of these beta-lactamase gene families were expressed in only one isoform each. Specifically, the blaCTX-M-15 isoform was found in 1.82% (n = 4) of the strains, blaVIM-1 in 4.13% (n = 9) and blaP in 6.88% (n = 15) of the 218 Escherichia coli strains studied, isolated from patients attending the “October first” Regional Hospital in Mexico City. These specific isoforms of blaCTX-M-15, blaVIM-1 and blaP beta-lactamases were detected in the given samples, which might explain the observed differences from their more commonly reported counterparts worldwide, like blaCTX-M-1 and blaCTX-M-15, which are frequently identified extended-spectrum beta-lactamases (ESBLs) globally. The prevalence of specific beta-lactamase isoforms could be influenced by the sample type and origin.
It is worth noting that both
blaCTX-M-1,
blaVIM-1 and
blaP beta-lactamases have been previously cloned and sequenced in
Escherichia coli strains [
71,
72,
73].
4.5.6. blaLAP and blaKPC Gene Families
The blaLAP gene was expressed in 6.88% of the isolated
Escherichia coli strains, with only two detected isoforms:
blaLAP-1 was the dominant gene expressed, accounting for 73.33% of all positive
blaLAP strains, followed by the
blaLAP-2 gene isoform, representing 26.67%. This gene, in combination with other beta-lactamases, contributes to the multi-drug resistance characteristics [
74].
Similarly, the
blaKPC gene family was expressed in two isoforms, namely
blaKPC-2 and
blaKPC-3, both of which have been previously reported as members of the
Escherichia coli genotype [
75,
76]. These isoforms provide carbapenem resistance characteristics [
77].
4.5.7. blaBIL, blaDHA and blaCMY Gene Families
The
blaBIL gene family was the fourth most frequent, but only the
blaBIL-1 isoform was identified. Previous reports have confirmed the expression of
blaBIL-1 in
Escherichia coli, conferring resistance characteristics to penicillin, carbenicillin, cefaclor and cephalosporin [
52].
Additionally, the
blaDHA (19.27%) and
blaCMY (18.81%) gene families were expressed at similar frequencies. Within the
blaDHA gene family, three isoforms were identified:
blaDHA-1 (80.95%),
blaDHA-2 (14.29%) and
blaDHA-7 (4.76%). The
blaCMY gene family was expressed in four isoforms:
blaCMY-2 (90.24%),
blaCMY-38 (4.88%),
blaCMY-9 and
blaCMY-12 (both expressed in 2.44%). The most dominant isoforms carried by
Escherichia coli-causing nosocomial infections were
blaDHA-1 and
blaCMY-2. These genes were previously described as co-expressed with the
blaCTX-M-15 isoform in a multi-resistant
Escherichia coli strain causing the death of a puppy in Italy, indicating ESBL characteristics [
78].
All of the blaBIL, blaDHA and blaCMY genes found in this study have been previously cloned and sequenced in
Escherichia coli [
52,
79,
80].
4.6. Beta-Lactam Resistome
The beta-lactam antibiotics, particularly penicillin, ampicillin and third-generation cephalosporin, play a crucial role in the treatment of nosocomial infections caused by
Escherichia coli [
81]. However, their extensive use in hospitals exerts significant selection pressure on the entire bacteriome, leading to the emergence of new beta-lactamases. This has become the most significant mechanism of resistance to beta-lactam antibiotics [
82]. These resistance mechanisms are encoded in the bacterial chromosome, plasmids or transposons, facilitating the rapid spread of beta-lactamase genes among bacterial populations. As a result, the development of novel strategies to combat antibiotic resistance becomes essential to tackle the growing challenges posed by nosocomial infections.
In this study, our focus was on the identification of 14 of the most clinically important beta-lactamase families. These included the
blaBIL,
blaCFX,
blaCMY,
blaCTX,
blaDHA,
blaIMP,
blaKPC,
blaLAP,
blaOXA,
blaP,
blaROB,
blaSHV,
blaTEM and
blaVIM gene families, collectively covering 99% of the beta-lactam resistance reported in
Escherichia coli strains causing various intestinal, extraintestinal and systemic pathologies [
83,
84]. By focusing on these specific gene families, we aimed to gain valuable insights into the prevalence and diversity of beta-lactam-resistance mechanisms in clinically relevant
Escherichia coli strains, which is crucial for understanding and combating antibiotic resistance in medical settings.
Among the Escherichia coli strains analyzed, we observed that they express from one to nine members of beta-lactamase gene families. It is worth noting that the accumulation of four or more members of the beta-lactamase gene families is a rare occurrence, as they are often unique strains. Nonetheless, these strains remain of utmost importance as they represent potential sources for the expansion of pan-resistant Escherichia coli strains with resistance to all beta-lactam drugs. The presence and diversity of these beta-lactamase genes emphasize the urgent need for effective measures to control and combat antibiotic resistance in nosocomial infections
In our study, the dominant beta-lactamase genotypes in the analyzed Escherichia coli strains were formed by blaTEM (21.6%) and its combinations with blaCTX (19.3%), blaCTX and blaSHV (6%) and blaCTX and blaBIL (5.5%). Together, these four genotypes constituted 52.4% of all identified genotypes. Based on these findings, the analyzed Escherichia coli strains are classified as extended-spectrum beta-lactamase-producing (ESBL) strains. The remaining combinations of beta-lactamase gene families represented from 0.5% to 4.1% each, but they still represent pan-resistant Escherichia coli strains against beta-lactam drugs. Our method of evaluation using real-time PCR and corroborated by next-generation sequencing provides a comprehensive panorama of the beta-lactam drug resistome in the analyzed Escherichia coli strains.
These results are in line with previous reports indicating that
Escherichia coli is the bacterial species that most frequently produces ESBLs [
85]. Similarly, in
Klebsiella pneumoniae, it has been reported that multiple beta-lactamase gene carriers express from two to four different members of the beta-lactamase gene families [
86]. However, it is essential to note that our method provides a broader and more generalized perspective of the beta-lactam drug resistome in the studied
Escherichia coli strains, allowing for a comprehensive understanding of the resistance mechanisms.
The molecular methods used for the identification and characterization of the
Escherichia coli beta-lactam resistome are of critical importance in drug therapy. In nosocomial infections caused by ESBL-producing
Escherichia coli, the available drug therapy options are significantly reduced due to the pan-resistant phenotype. This can lead to an increase in mortality rates, prolonged hospitalization time and higher healthcare costs [
87,
88]. Patients who have acquired nosocomial infections caused by ESBL-producing
Escherichia coli have been reported to face challenges in treatment [
89]. For instance, the identification of
Escherichia coli strains producing ESBLs of the CTX-M type has been linked to treatment failure in urinary-tract infections (UTIs) when treated with commonly used antibiotics such as penicillin, ampicillin and first- to fourth-generation cephalosporins. Even next-generation antibiotics like carbapenem and imipenem may prove ineffective against these strains, as some of them have also developed resistance to beta-lactamase inhibitors [
90,
91].
Our study’s results are consistent with those published in a multicenter study conducted in eight hospitals in Mexico, where the dominant isolated microorganisms were ESBL-producing
Escherichia coli strains (83.9%) and many carried genes encoding beta-lactamases of
blaSHV and/or
blaCTX-M types [
92]. These findings emphasize the urgent need for robust molecular methods to identify and characterize beta-lactam resistance in
Escherichia coli, as they play a crucial role in guiding appropriate and effective drug therapy decisions for nosocomial infections.

 ,
, 



{kind=link}
{kind=link}