
Genetic Characterization of Carbapenem-Resistant Klebsiella pneumoniae Clinical Isolates in a Tertiary Hospital in Greece, 2018–2022
 , , and
, , and
Abstract
:1. Introduction
2. Results
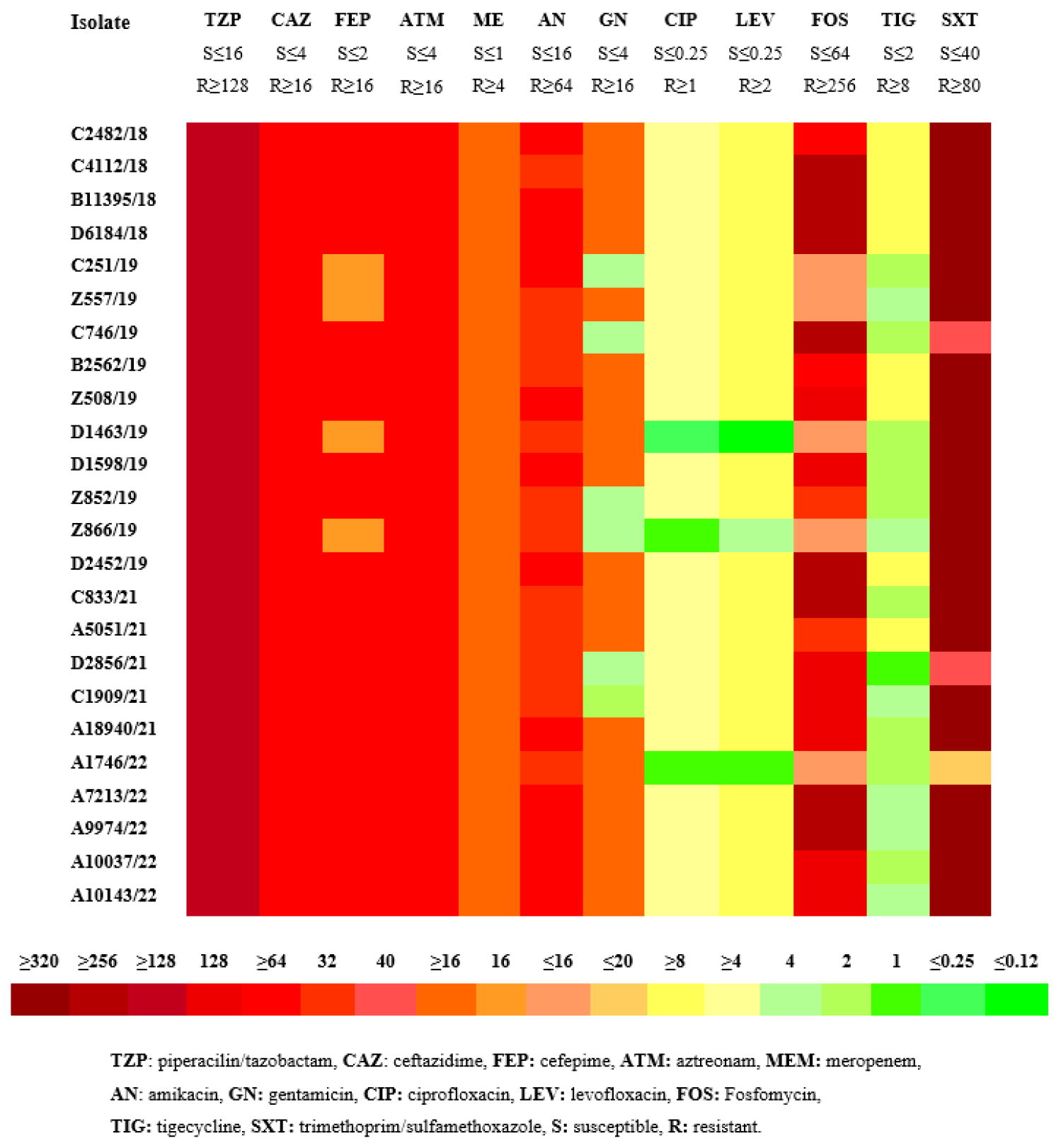
2.1. Antimicrobial Susceptibility Testing
2.2. MLST, Antimicrobial Resistance Genes, Plasmids
2.3. Virulence Factors and Efflux and Regulator Systems
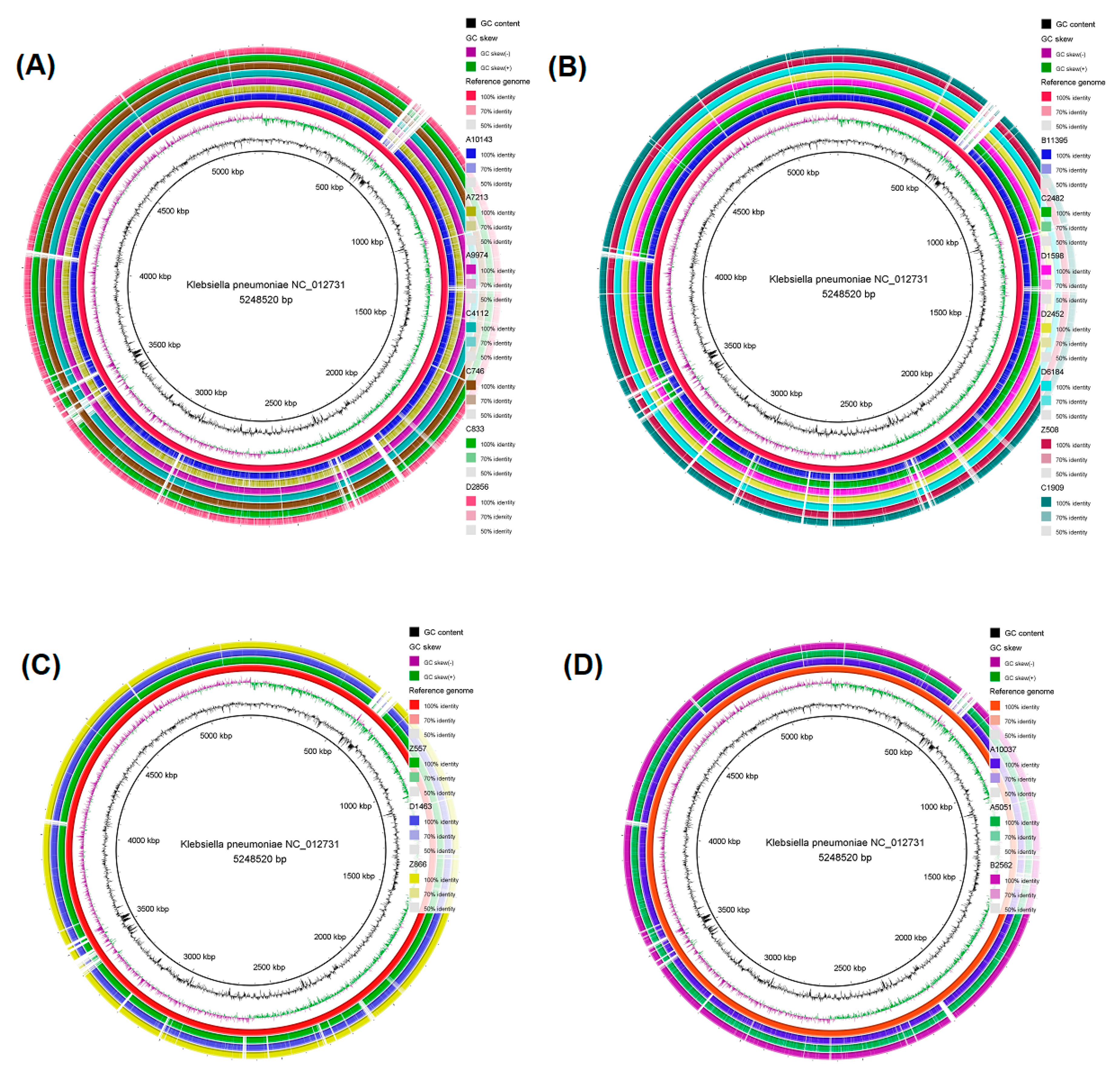
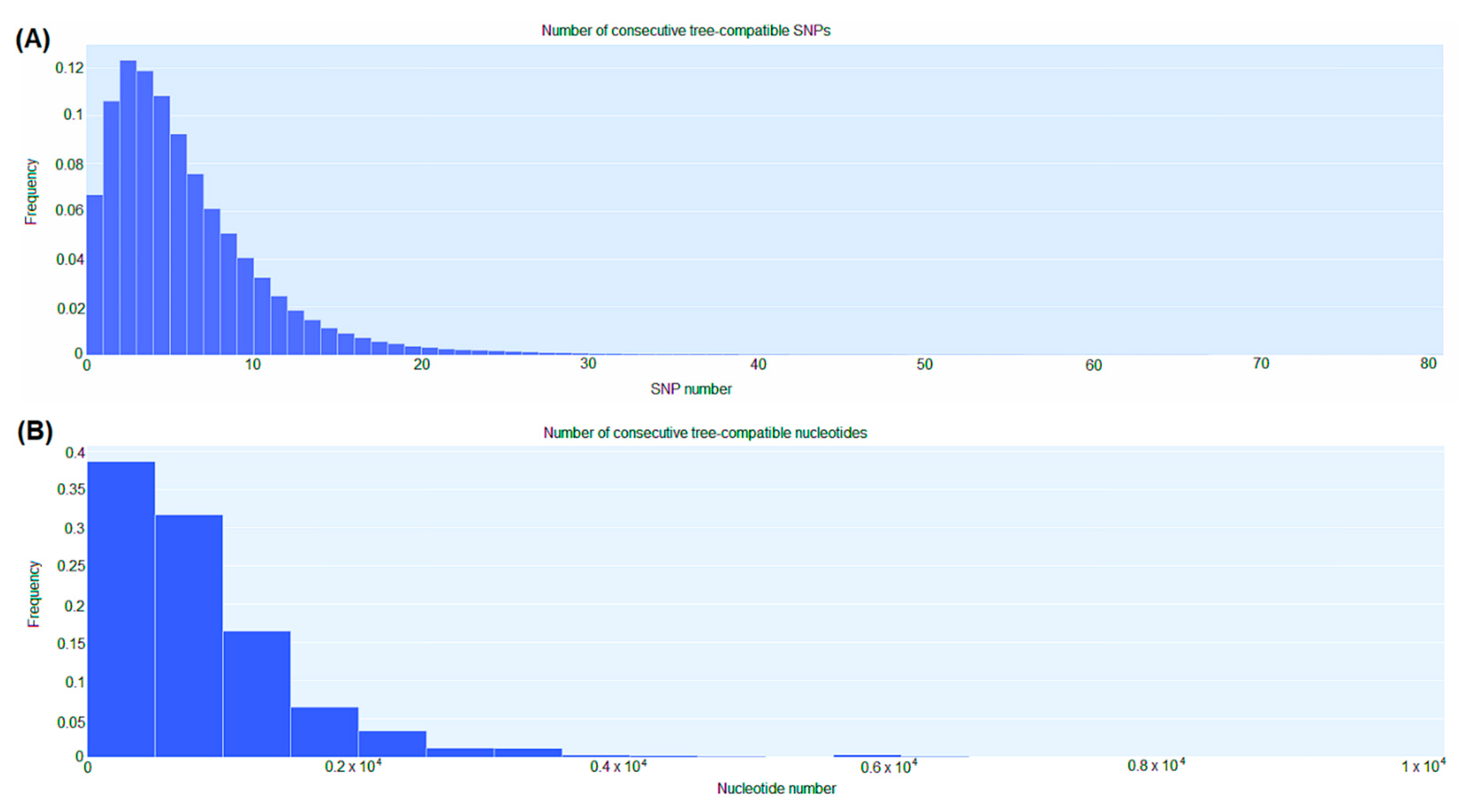
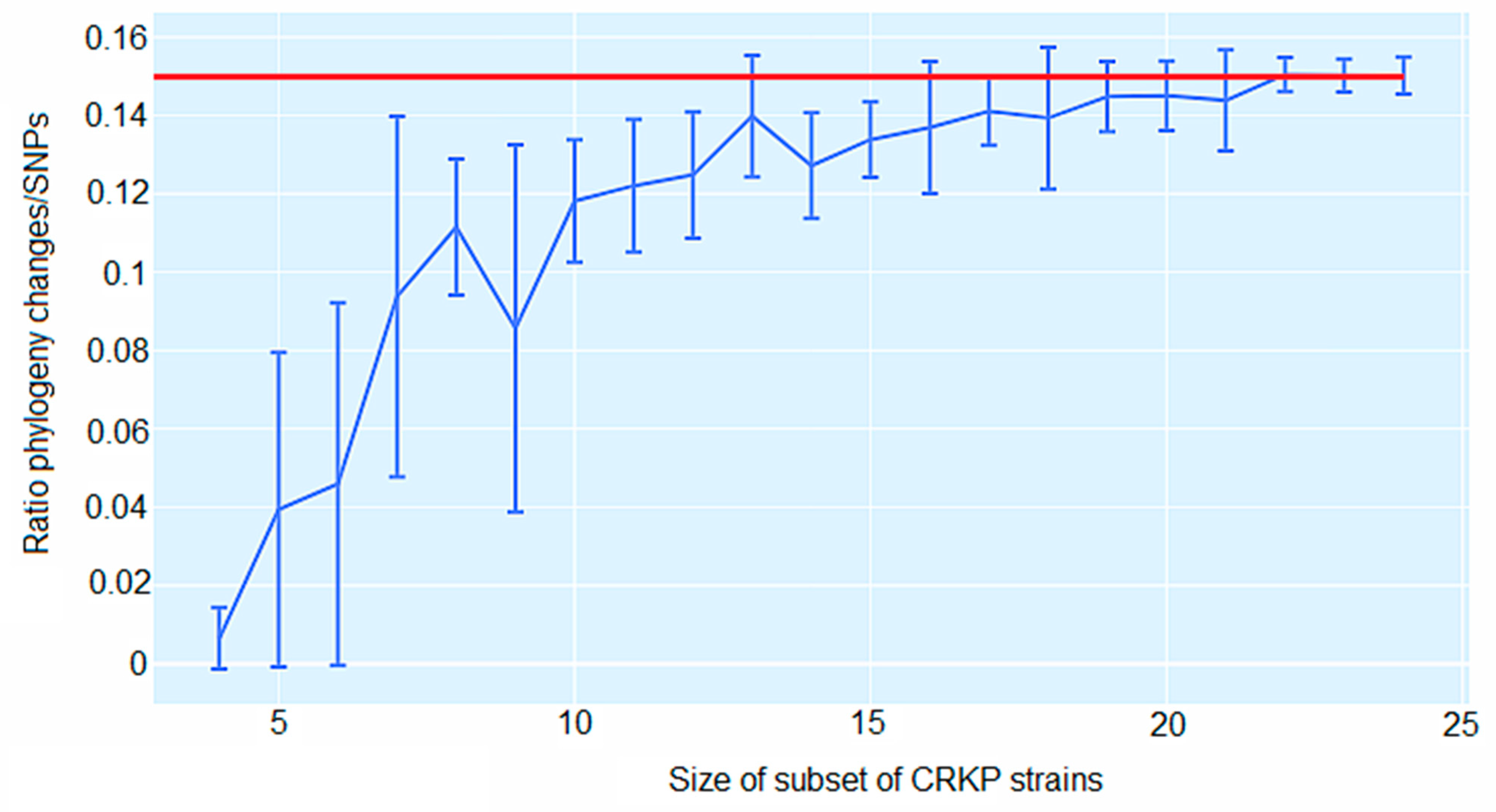
2.4. Genomic Comparison among CRKP Strains
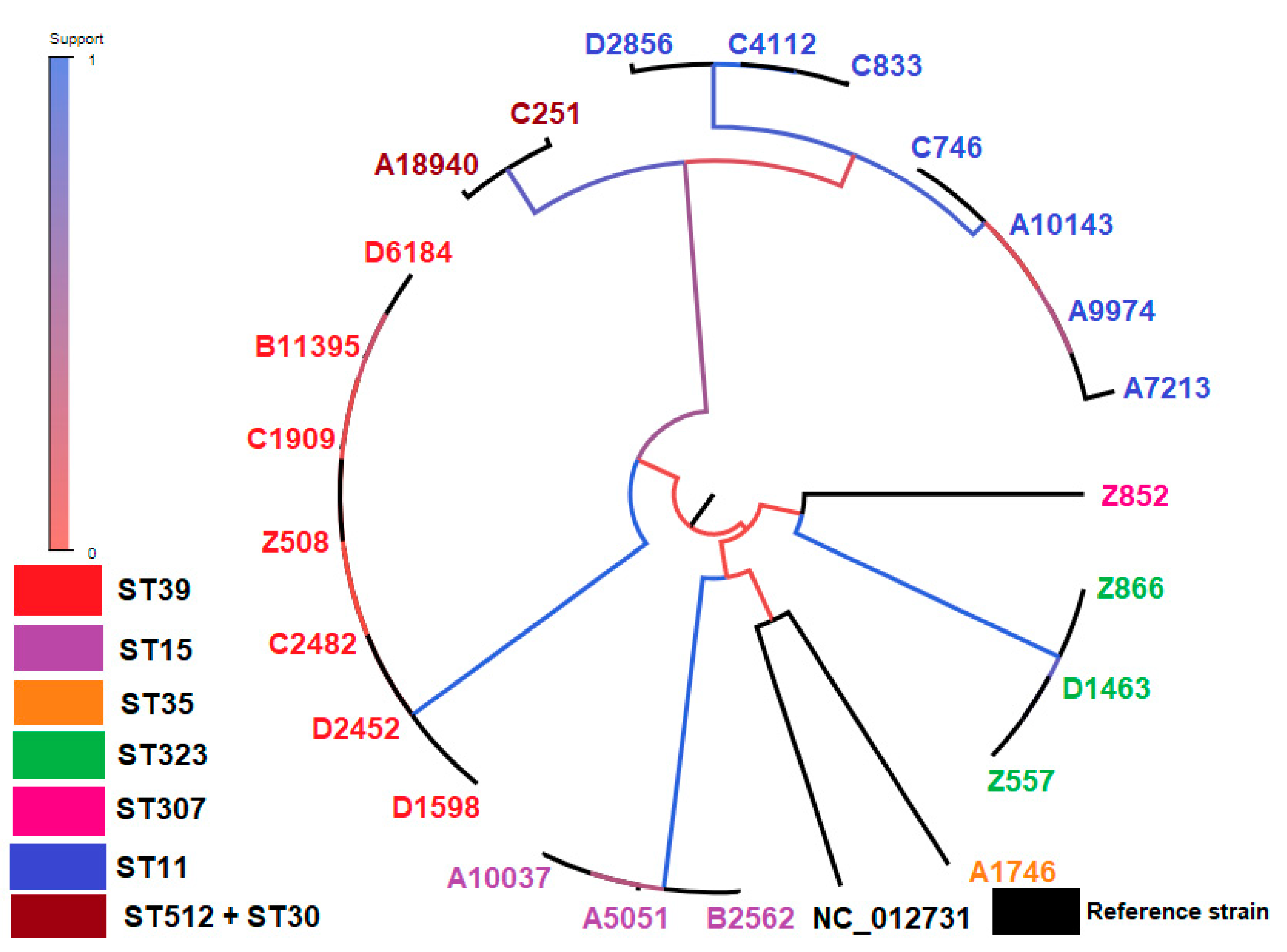
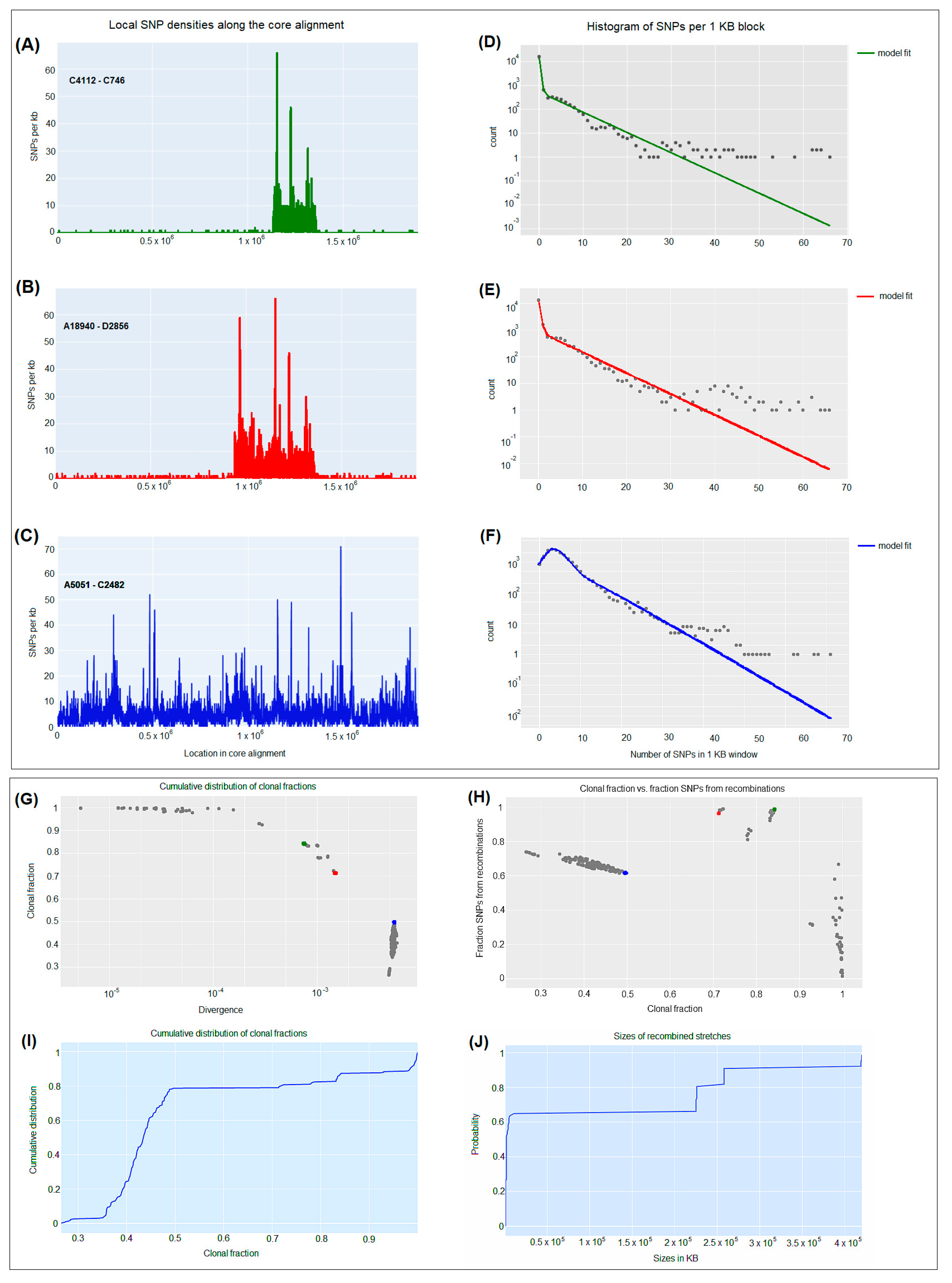
2.5. CgSNP-Based Phylogenetic Analysis
3. Materials and Methods
3.1. Clinical Isolates-Setting
3.2. Ethics Approval
3.3. Microbiological Methods-Antimicrobial Susceptibility Testing
3.4. DNA Extraction and Whole Genome Sequencing
3.5. Assembly Assessment and Genome Annotation
3.6. MLST and Detection of Antimicrobial Resistance Genes and Plasmids
3.7. Genomic Comparison–Core Genome Single-Nucleotide Polymorphism (cgSNP)-Based Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Juan, C.H.; Fang, S.Y.; Chou, C.H.; Tsai, T.Y.; Lin, Y.T. Clinical characteristics of patients with pneumonia caused by Klebsiella pneumoniae in Taiwan and prevalence of antimicrobial-resistant and hypervirulent strains: A retrospective study. Antimicrob. Resist. Infect. Control 2020, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.M.; Bachman, M.A. Colonization, Infection, and the Accessory Genome of Klebsiella pneumoniae. Front. Cell. Infect. Microbiol. 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Wang, W.; Liu, S.; Zhang, Z.; Jiang, M.; Zhang, F. Molecular Epidemiology and Drug Resistant Mechanism of Carbapenem-Resistant Klebsiella pneumoniae in Elderly Patients With Lower Respiratory Tract Infection. Front. Public Health 2021, 9, 669173. [Google Scholar] [CrossRef]
- Gupta, A.; Bhatti, S.; Leytin, A.; Epelbaum, O. Novel complication of an emerging disease: Invasive Klebsiella pneumoniae liver abscess syndrome as a cause of acute respiratory distress syndrome. Clin. Pract. 2018, 8, 1021. [Google Scholar] [CrossRef] [PubMed]
- Ferri, M.; Ranucci, E.; Romagnoli, P.; Giaccone, V. Antimicrobial resistance: A global emerging threat to public health systems. Crit. Rev. Food Sci. Nutr. 2017, 57, 2857–2876. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.; Hemarajata, P.; Sun, D.; Rubio-Aparicio, D.; Tsivkovski, R.; Yang, S.; Sebra, R.; Kasarskis, A.; Nguyen, H.; Hanson, B.M.; et al. Resistance to Ceftazidime-Avibactam is Due to Transposition of KPC in a Porin-Deficient Strain of Klebsiella pneumoniae with Increased Efflux Activity. Antimicrob. Agents Chemother. 2017, 61, e00989-17. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Pan, F.; Wang, C.; Zhao, W.; Sun, Y.; Zhang, T.; Shi, Y.; Zhang, H. Molecular epidemiology of Carbapenem-resistant Klebsiella pneumoniae in a paediatric hospital in China. Int. J. Infect. Dis. 2020, 93, 311–319. [Google Scholar] [CrossRef]
- Galani, I.; Karaiskos, I.; Angelidis, E.; Papoutsaki, V.; Galani, L.; Souli, M.; Antoniadou, A.; Giamarellou, H. Emergence of ceftazidime-avibactam resistance through distinct genomic adaptations in KPC-2-producing Klebsiella pneumoniae of sequence type 39 during treatment. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 219–224. [Google Scholar] [CrossRef]
- Galani, I.; Nafplioti, K.; Adamou, P.; Karaiskos, I.; Giamarellou, H.; Souli, M. Study Collaborators Nationwide epidemiology of carbapenem resistant Klebsiella pneumoniae isolates from Greek hospitals, with regards to plazomicin and aminoglycoside resistance. BMC Infect. Dis. 2019, 19, 167. [Google Scholar]
- Karampatakis, T.; Tsergouli, K.; Politi, L.; Diamantopoulou, G.; Iosifidis, E.; Antachopoulos, C.; Karyoti, A.; Mouloudi, E.; Tsakris, A.; Roilides, E. Molecular Epidemiology of Endemic Carbapenem-Resistant Gram-Negative Bacteria in an Intensive Care Unit. Microb. Drug Resist. 2019, 25, 712–716. [Google Scholar] [CrossRef]
- Zarras, C.; Pappa, S.; Zarras, K.; Karampatakis, T.; Vagdatli, E.; Mouloudi, E.; Iosifidis, E.; Roilides, E.; Papa, A. Changes in molecular epidemiology of carbapenem-resistant Klebsiella pneumoniae in the intensive care units of a Greek hospital, 2018–2021. Acta Microbiol. Immunol. Hung. 2022, 69, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Karampatakis, T.; Antachopoulos, C.; Iosifidis, E.; Tsakris, A.; Roilides, E. Molecular epidemiology of carbapenem-resistant Klebsiella pneumoniae in Greece. Future Microbiol. 2016, 11, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Protonotariou, E.; Meletis, G.; Pilalas, D.; Mantzana, P.; Tychala, A.; Kotzamanidis, C.; Papadopoulou, D.; Papadopoulos, T.; Polemis, M.; Metallidis, S.; et al. Polyclonal Endemicity of Carbapenemase-Producing Klebsiella pneumoniae in ICUs of a Greek Tertiary Care Hospital. Antibiotics 2022, 11, 149. [Google Scholar] [CrossRef] [PubMed]
- Surveillance Atlas of Infectious Diseases. 2020. Available online: https://atlas.Ecdc.Europa.Eu/public/index.Aspx (accessed on 5 December 2022).
- Hatrongjit, R.; Kerdsin, A.; Akeda, Y.; Hamada, S. Detection of plasmid-mediated colistin-resistant and carbapenem-resistant genes by multiplex PCR. MethodsX 2018, 5, 532–536. [Google Scholar] [CrossRef]
- Mari-Almirall, M.; Ferrando, N.; Fernandez, M.J.; Cosgaya, C.; Vines, J.; Rubio, E.; Cuscó, A.; Muñoz, L.; Pellice, M.; Vergara, A.; et al. Clonal Spread and Intra- and Inter-Species Plasmid Dissemination Associated with Klebsiella pneumoniae Carbapenemase-Producing Enterobacterales during a Hospital Outbreak in Barcelona, Spain. Front. Microbiol. 2021, 12, 781127. [Google Scholar] [CrossRef]
- Yanat, B.; Rodriguez-Martinez, J.M.; Touati, A. Plasmid-mediated quinolone resistance in Enterobacteriaceae: A systematic review with a focus on Mediterranean countries. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 421–435. [Google Scholar] [CrossRef]
- Bosch, T.; Lutgens, S.P.M.; Hermans, M.H.A.; Wever, P.C.; Schneeberger, P.M.; Renders, N.H.M.; Leenders, A.C.A.P.; Kluytmans, J.A.J.W.; Schoffelen, A.; Notermans, D.; et al. Outbreak of NDM-1-Producing Klebsiella pneumoniae in a Dutch Hospital, with Interspecies Transfer of the Resistance Plasmid and Unexpected Occurrence in Unrelated Health Care Centers. J. Clin. Microbiol. 2017, 55, 2380–2390. [Google Scholar] [CrossRef]
- Silva, D.D.C.; Rampelotto, R.F.; Lorenzoni, V.V.; Santos, S.O.D.; Damer, J.; Horner, M.; Horner, R. Phenotypic methods for screening carbapenem-resistant Enterobacteriaceae and assessment of their antimicrobial susceptibility profile. Rev. Soc. Bras. Med. Trop. 2017, 50, 173–178. [Google Scholar] [CrossRef]
- Tzouvelekis, L.S.; Miriagou, V.; Kotsakis, S.D.; Spyridopoulou, K.; Athanasiou, E.; Karagouni, E.; Tzelepi, E.; Daikos, G.L. KPC-producing, multidrug-resistant Klebsiella pneumoniae sequence type 258 as a typical opportunistic pathogen. Antimicrob. Agents Chemother. 2013, 57, 5144–5146. [Google Scholar] [CrossRef]
- Karampatakis, T.; Zarras, C.; Pappa, S.; Vagdatli, E.; Iosifidis, E.; Roilides, E.; Papa, A. Emergence of ST39 carbapenem-resistant Klebsiella pneumoniae producing VIM-1 and KPC-2. Microb. Pathog. 2022, 162, 105373. [Google Scholar] [CrossRef]
- Protonotariou, E.; Meletis, G.; Chatzopoulou, F.; Malousi, A.; Chatzidimitriou, D.; Skoura, L. Emergence of Klebsiella pneumoniae ST11 co-producing NDM-1 and OXA-48 carbapenemases in Greece. J. Glob. Antimicrob. Resist. 2019, 19, 81–82. [Google Scholar] [CrossRef]
- Partridge, S.R.; Ginn, A.N.; Wiklendt, A.M.; Ellem, J.; Wong, J.S.; Ingram, P.; Guy, S.; Garner, S.; Iredell, J.R. Emergence of blaKPC carbapenemase genes in Australia. Int. J. Antimicrob. Agents 2015, 45, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.J.; Ku, Y.H.; Lee, M.F.; Chuang, Y.C.; Yu, W.L. In Vitro Activity of Imipenem and Colistin against a Carbapenem-Resistant Klebsiella pneumoniae Isolate Coproducing SHV-31, CMY-2, and DHA-1. BioMed Res. Int. 2015, 2015, 568079. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, F.M.; Dib-Hajj, F.; Shang, W.; Gootz, T.D. High-level carbapenem resistance in a Klebsiella pneumoniae clinical isolate is due to the combination of bla(ACT-1) beta-lactamase production, porin OmpK35/36 insertional inactivation, and down-regulation of the phosphate transport porin phoe. Antimicrob. Agents Chemother. 2006, 50, 3396–3406. [Google Scholar] [CrossRef] [PubMed]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the Offense with a Strong Defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef]
- Brhelova, E.; Antonova, M.; Pardy, F.; Kocmanova, I.; Mayer, J.; Racil, Z.; Lengerova, M. Investigation of next-generation sequencing data of Klebsiella pneumoniae using web-based tools. J. Med. Microbiol. 2017, 66, 1673–1683. [Google Scholar] [CrossRef]
- Enany, S.; Zakeer, S.; Diab, A.A.; Bakry, U.; Sayed, A.A. Whole genome sequencing of Klebsiella pneumoniae clinical isolates sequence type 627 isolated from Egyptian patients. PLoS ONE 2022, 17, e0265884. [Google Scholar] [CrossRef]
- CLSI standard M02; Performance Standards for Antimicrobial Susceptibility Testing; 32nd Ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020.
- U.S. FDA. FDA approves new antibacterial drug Avycaz. In FDA News Release; U.S. FDA: Silver Spring, MD, USA, 2015. [Google Scholar]
- Tsakris, A.; Pournaras, S.; Woodford, N.; Palepou, M.F.; Babini, G.S.; Douboyas, J.; Livermore, D.D. Outbreak of infections caused by Pseudomonas aeruginosa producing VIM-1 carbapenemase in Greece. J. Clin. Microbiol. 2000, 38, 1290–1292. [Google Scholar] [CrossRef]
- Geneious Prime 2021.2.1. Available online: https://www.geneious.com (accessed on 1 December 2022).
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef]
- Wu, K.M.; Li, L.H.; Yan, J.J.; Tsao, N.; Liao, T.L.; Tsai, H.C.; Fung, C.P.; Chen, H.J.; Liy, Y.M.; Chen, Y.T.; et al. Genome sequencing and comparative analysis of Klebsiella pneumoniae NTUH-K2044, a strain causing liver abscess and meningitis. J. Bacteriol. 2009, 191, 4492–4501. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Ponten, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017, Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef] [PubMed]
- Bertels, F.; Silander, O.K.; Pachkov, M.; Rainey, P.B.; van Nimwegen, E. Automated reconstruction of whole-genome phylogenies from short-sequence reads. Mol. Biol. Evol. 2014, 31, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Dixit, P.D.; Pang, T.Y.; Studier, F.W.; Maslov, S. Recombinant transfer in the basic genome of Escherichia coli. Proc. Natl. Acad. Sci. USA 2015, 112, 9070–9075. [Google Scholar] [CrossRef]
- Sakoparnig, T.; Field, C.; van Nimwegen, E. Whole genome phylogenies reflect the distributions of recombination rates for many bacterial species. elife 2021, 10, e65366. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, L.; Zhou, H.; Chan, E.W.; Li, J.; Fang, Y.; Li, Y.; Liao, K.; Chen, S. Nationwide Surveillance of Clinical Carbapenem-resistant Enterobacteriaceae (CRE) Strains in China. eBioMedicine 2017, 19, 98–106. [Google Scholar] [CrossRef]
- Voulgari, E.; Gartzonika, C.; Vrioni, G.; Politi, L.; Priavali, E.; Levidiotou-Stefanou, S.; Tsakris, A. The Balkan region: NDM-1-producing Klebsiella pneumoniae ST11 clonal strain causing outbreaks in Greece. J. Antimicrob. Chemother. 2014, 69, 2091–2097. [Google Scholar] [CrossRef]
- Todorova, B.; Sabtcheva, S.; Ivanov, I.N.; Lesseva, M.; Chalashkanov, T.; Ioneva, M.; Bachvarova, A.; Dobreva, E.; Kantardjiev, T. First clinical cases of NDM-1-producing Klebsiella pneumoniae from two hospitals in Bulgaria. J. Infect. Chemother. 2016, 22, 837–840. [Google Scholar] [CrossRef] [PubMed]
- Berglund, B.; Hoang, N.T.B.; Lundberg, L.; Le, N.K.; Tarnberg, M.; Nilsson, M.; Bornefall, E.; Khu, D.T.K.; Welander, J.; Le, H.T.; et al. Clonal spread of carbapenem-resistant Klebsiella pneumoniae among patients at admission and discharge at a Vietnamese neonatal intensive care unit. Antimicrob. Resist. Infect. Control 2021, 10, 162. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wilksch, J.; Li, B.; Du, J.; Cao, J.; Zhang, X.; Zhou, T. Emergence of ST39 and ST656 extensively drug-resistant Klebsiella pneumoniae isolates in Wenzhou, China. Indian. J. Med. Microbiol. 2017, 35, 145–146. [Google Scholar] [CrossRef]
- Cabanel, N.; Rosinski-Chupin, I.; Chiarelli, A.; Botin, T.; Tato, M.; Canton, R.; Glaser, P. Evolution of VIM-1-Producing Klebsiella pneumoniae Isolates from a Hospital Outbreak Reveals the Genetic Bases of the Loss of the Urease-Positive Identification Character. mSystems 2021, 6, e0024421. [Google Scholar] [CrossRef] [PubMed]
- Kuzina, E.S.; Kislichkina, A.A.; Sizova, A.A.; Skryabin, Y.P.; Novikova, T.S.; Ershova, O.N.; Savin, I.A.; Khokhlova, O.E.; Bogun, A.G.; Fursova, N.K. High-Molecular-Weight Plasmids Carrying Carbapenemase Genes bla(NDM-1), bla(KPC-2), and bla(OXA-48) Coexisting in Clinical Klebsiella pneumoniae Strains of ST39. Microorganisms 2023, 11, 459. [Google Scholar] [CrossRef]
- Giakkoupi, P.; Papagiannitsis, C.C.; Miriagou, V.; Pappa, O.; Polemis, M.; Tryfinopoulou, K.; Tzouvelekis, L.S.; Vatopoulos, A.C. An update of the evolving epidemic of blaKPC-2-carrying Klebsiella pneumoniae in Greece (2009-10). J. Antimihcrob. Chemother. 2011, 66, 1510–1513. [Google Scholar] [CrossRef]
- Piekarska, K.; Zacharczuk, K.; Wolkowicz, T.; Wolaniuk, N.; Rzeczkowska, M.; Gierczynski, R. Emergence of Enterobacteriaceae co-producing CTX-M-15, ArmA and PMQR in Poland. Adv. Clin. Exp. Med. 2019, 28, 249–254. [Google Scholar] [CrossRef]
- Mansour, W.; Grami, R.; Ben Haj Khalifa, A.; Dahmen, S.; Chatre, P.; Haenni, M.; Aouni, M.; Madec, J.Y. Dissemination of multidrug-resistant blaCTX-M-15/IncFIIk plasmids in Klebsiella pneumoniae isolates from hospital- and community-acquired human infections in Tunisia. Diagn. Microbiol. Infect. Dis. 2015, 83, 298–304. [Google Scholar] [CrossRef]
- Galani, I.; Karaiskos, I.; Souli, M.; Papoutsaki, V.; Galani, L.; Gkoufa, A.; Antoniadou, A.; Giamarellou, H. Outbreak of KPC-2-producing Klebsiella pneumoniae endowed with ceftazidime-avibactam resistance mediated through a VEB-1-mutant (VEB-25), Greece, September to October 2019. Eurosurveillance 2020, 25, 2000028. [Google Scholar] [CrossRef]
- Voulgari, E.; Kotsakis, S.D.; Giannopoulou, P.; Perivolioti, E.; Tzouvelekis, L.S.; Miriagou, V. Detection in two hospitals of transferable ceftazidime-avibactam resistance in Klebsiella pneumoniae due to a novel VEB beta-lactamase variant with a Lys234Arg substitution, Greece, 2019. Eurosurveillance 2020, 25, 1900766. [Google Scholar] [CrossRef]
- Livermore, D.M.; Struelens, M.; Amorim, J.; Baquero, F.; Bille, J.; Canton, R.; Henning, S.; Gatermann, S.; Marchese, A.; Mittermayer, H.; et al. Multicentre evaluation of the VITEK 2 Advanced Expert System for interpretive reading of antimicrobial resistance tests. J. Antimicrob. Chemother. 2002, 49, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fulgueiras, V.; Magallanes, C.; Reyes, V.; Cayota, C.; Galiana, A.; Vieytes, M.; Vignoli, R.; Márquez, C. In Vivo High Plasticity of Multi-Drug Resistant ST258 Klebsiella pneumoniae. Microb. Drug Resist. 2021, 27, 1126–1130. [Google Scholar] [CrossRef] [PubMed]
- Albarri, O.; AlMatar, M.; Ocal, M.M.; Koksal, F. Overexpression of Efflux Pumps AcrAB and OqxAB Contributes to Ciprofloxacin Resistance in Clinical Isolates of K. pneumonia. Curr. Protein Pept. Sci. 2022, 23, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Diez-Aguilar, M.; Canton, R. New microbiological aspects of fosfomycin. Rev. Esp. Quimioter. 2019, 32 (Suppl. S1), 8–18. [Google Scholar]
- Sugita, K.; Aoki, K.; Komori, K.; Nagasawa, T.; Ishii, Y.; Iwata, S.; Tateda, K. Molecular Analysis of bla(KPC-2)-Harboring Plasmids: Tn4401a Interplasmid Transposition and Tn4401a-Carrying ColRNAI Plasmid Mobilization from Klebsiella pneumoniae to Citrobacter europaeus and Morganella morganii in a Single Patient. mSphere 2021, 6, e0085021. [Google Scholar] [CrossRef]
- Di Martino, P.; Cafferini, N.; Joly, B.; Darfeuille-Michaud, A. Klebsiella pneumoniae type 3 pili facilitate adherence and biofilm formation on abiotic surfaces. Res. Microbiol. 2003, 154, 9–16. [Google Scholar] [CrossRef]
- Lam, M.M.C.; Wick, R.R.; Wyres, K.L.; Gorrie, C.L.; Judd, L.M.; Jenney, A.W.J.; Brisse, S.; Holt, K.E. Genetic diversity, mobilisation and spread of the yersiniabactin-encoding mobile element ICEKp in Klebsiella pneumoniae populations. Microb. Genom. 2018, 4, e000196. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, T.; Chen, L.; Du, H. Virulence Factors in Hypervirulent Klebsiella pneumoniae. Front. Microbiol. 2021, 12, 642484. [Google Scholar] [CrossRef]
- Di Pilato, V.; Henrici De Angelis, L.; Aiezza, N.; Baccani, I.; Niccolai, C.; Parisio, E.M.; Giordano, C.; Camarlinghi, G.; Barnini, S.; Forni, S.; et al. Resistome and virulome accretion in an NDM-1-producing ST147 sublineage of Klebsiella pneumoniae associated with an outbreak in Tuscany, Italy: A genotypic and phenotypic characterisation. Lancet Microbe 2022, 3, e224–e234. [Google Scholar] [CrossRef]
- Miro, E.; Rossen, J.W.A.; Chlebowicz, M.A.; Harmsen, D.; Brisse, S.; Passet, V.; Navarro, F.; Friedrich, A.W.; García-Cobos, S. Core/Whole Genome Multilocus Sequence Typing and Core Genome SNP-Based Typing of OXA-48-Producing Klebsiella pneumoniae Clinical Isolates From Spain. Front. Microbiol. 2019, 10, 2961. [Google Scholar] [CrossRef]
- Snitkin, E.S.; Zelazny, A.M.; Thomas, P.J.; Stock, F.; NISC Comparative Sequencing Program Group; Henderson, D.K.; Palmore, T.N.; Segre, J.A. Tracking a hospital outbreak of carbapenem-resistant Klebsiella pneumoniae with whole-genome sequencing. Sci. Transl. Med. 2012, 4, 148ra116. [Google Scholar] [CrossRef] [PubMed]
- Marsh, J.W.; Krauland, M.G.; Nelson, J.S.; Schlackman, J.L.; Brooks, A.M.; Pasculle, A.W.; Shutt, K.A.; Doi, Y.; Querry, A.M.; Muto, C.A.; et al. Genomic Epidemiology of an Endoscope-Associated Outbreak of Klebsiella pneumoniae Carbapenemase (KPC)-Producing K. pneumoniae. PLoS ONE 2015, 10, e0144310. [Google Scholar] [CrossRef] [PubMed]
- Onori, R.; Gaiarsa, S.; Comandatore, F.; Pongolini, S.; Brisse, S.; Colombo, A.; Cassani, G.; Marone, P.; Grossi, P.; Minoja, G.; et al. Tracking Nosocomial Klebsiella pneumoniae Infections and Outbreaks by Whole-Genome Analysis: Small-Scale Italian Scenario within a Single Hospital. J. Clin. Microbiol. 2015, 53, 2861–2868. [Google Scholar] [CrossRef]
- Arnold, B.J.; Gutmann, M.U.; Grad, Y.H.; Sheppard, S.K.; Corander, J.; Lipsitch, M.; Hanage, W.P. Weak Epistasis May Drive Adaptation in Recombining Bacteria. Genetics 2018, 208, 1247–1260. [Google Scholar] [CrossRef] [PubMed]
- Dallman, T.; Ashton, P.; Schafer, U.; Jironkin, A.; Painset, A.; Shaaban, S.; Hartman, H.; Myers, R.; Underwood, A.; Jenkins, C.; et al. SnapperDB: A database solution for routine sequencing analysis of bacterial isolates. Bioinformatics 2018, 34, 3028–3029. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | ST | Plasmids | Antibiotic Resistance Genes Affecting | Virulence | Efflux/Regulator | Capsule | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Other Beta-Lactams | Carba-Penems | Amino-Glycosides | Quinolones | Other | ||||||
| C2482/18 | 39 | ColRNAI IncA/C2 IncFIB IncFIB(pQil) Col (pHAD28) | blaSHV-79 blaTEM-1B | blaKPC-2 blaVIM-1 | aac(3)-Iid aac(6′)-Ib aac(6′)-Im aant(2″)-Ia aph(2″)-Ib aph(3′)-Ia | oqxA, oqxB | fosA, sul1, Sul3, dfrA1 | fyuA, mrkABCDFHIJ ybtASTX | acrB, marAR, soxSR, ramA, rob sdiA, fis, envR, oqxBR, rarA | wzc, |
| C4112/18 | 11 | ColRNAI IncFIA(HI1) IncFIB(K) IncFII(K) | blaCTX-M-15 blaSHV-11 | blaNDM-1 | aac(3)-Iia, aac(6′)-Ib aph(3′)-Ia | aac(6′)-lb-cr, oqxA, oqxB | fosA, sul2 dfrA14, catB3, tet(D) | iutA, mrkABCDFHIJ | acrABR marAR soxSR ramAR rob sdiA, fis, envR, oqxR, rarA | wzc, wzi |
| B11395/18 | 39 | IncFIB (AP001918) IncFIB(pQil) IncFII(K) | blaSHV-79 blaTEM-1B | blaKPC-2 | aac(3)-Iid, aadA1, aadA2, aph(3′)-Ia | oqxA, oqxB | fosA, sul2, sul3, dfrA12, cmlA1 | fyuA, mrkABCDFHIJ, ybtEQSTU | acrABR marAR soxSR, ramA, sdiA, fis, envR, oqxA, rarA | wzc, wzi |
| D6184/18 | 39 | ColRNAI IncFIB (AP001918) IncFIB(pQil) | blaSHV-11 blaSHV-40 blaΤΕΜ-1Β | blaKPC-2 | aac(3)-Iid, aac(6′)-Ib, aph(3′)-Ia, aadA1, aadA2 | aac(6′)-lb-cr, oqxA, oqxB | fosA, sul2, sul3, tet(M), dfrA12, catA1, atB3, | fyuA, irp1, irp2, mrkABCDFHIJ, ybtAEPQSTUX | acrABR marAR oxSR, ramA, rob, sdiA, fis, envR, qxABR, rarA | wzc, wzi |
| C251/19 | 30 | ColRNAI IncA/C2 IncFIB(K) IncFII(K) IncX3 | blaSHV-12, blaTEM-1A blaOXA-9 | blaKPC-2 blaVIM-1 | aac(6′)-Ib aph(3′)-Ia aadA2 aadA15 | qnrS1, aac(6′)-lb-cr, oqxA, oqxB | fosA, sul1, dfrA1, dfrA12, catA, tet(D) | fyuA, irp1, iutA mrkBCDFIJ, ybtAEPQSTU | acrABR, marAR, soxSR, ramAR, rob, sdiA, envR, oqxABR, rarA | wzc, wzi |
| Ζ557/19 | 323 | IncFIB(K) IncFIB(pQil) IncFII(K) IncFIB(Mar) IncFIB (pKPHS1) IncHI1B | blaSHV-1 blaSHV-99 | blaKPC-2 blaVIM-1 | ant(2″)-Ia, aac(6′)-Il, aadA1 | qnrA1 oqxA, oqxB | fosA, fosA7, sul1, dfrA1 | mrkABCDFIJ | acrAR, marA, soxSR, ramA, rob, sdiA, fis, envR, oqxA, rarA | wzc, wzi |
| C746/19 | 11 | ColRNAI IncFIB(K) IncFIB(pQil) IncFII(K), IncR | blaSHV-1 blaOXA-1 | blaKPC-2 | aac(6′)-lb-cr aph(3′)-Ia aadA2 | aac(6′)-lb-cr, oqxA oqxB | fosA, sul1 | fyuA, irp1, irp2 iutA, mrkABCDFIJ, ybtAEUX | acrAB, marAR, soxSR, ramAR, rob, sdiA, fis, envR, oqxABR, | wzi |
| B2562/19 | 15 | IncFIB(K) IncFII(K) IncFIA(HI1) | blaCTX-M-15 blaTEM-1B blaOXA-1 | blaNDM-1 | aac(3)-Iia aac(6′)-Ib3 aph(3″)-Ib aph(6)-Id | aac(6′)-lb-cr, oqxA, oqxB | fosA, sul2, dfrA14, catB3 | fyuA, irp1, kfuABC, mrkABCDFIJ, ybtAPQSTUX | acrABR, marAR, soxSR, ramA, rob, sdiA, fis, envR, rarA | wzi |
| Ζ508/19 | 39 | ColRNAI IncA/C2 IncFIB IncFIB(pQil) | blaSHV-11 blaSHV-40 blaΤΕΜ-1Β blaOXA-116 blaOXA-366 | blaKPC-2 blaVIM-1 | aac(3)-Iid aac(6′)-Ib3 aac(6′)-Im aph(2″)-Ib aph(3′)-Ia aph(3′)-Via aadA24, armA, aac(6’)-Il | qnrS1 aac(6′)-lb-cr, oqxA, oqxB | fosA, sul1, sul3, frA1, tet(M) | fyuA, irp1, irp2, mrkABCDFHIJ, ybtAEPQSTUX | acrBR, marAR, soxSR, ramA, rob, sdiA, fis, envR, oqxABR, rarA | wzc, wzi |
| D1463/19 | 323 | IncFIB(K) IncFIB(Mar) IncFIB (pKPHS1) IncFIB(pQil) IncFII(K) IncHI1B | blaSHV-99 | blaKPC-2 blaVIM-1 | ant(2″)-Ia aac(6′)-Il | oqxA oqxB | fosA, fosA7, dfrA1 | mrkABCDFHJ, | acrAR, marA, soxSR, ramA, rob, sdiA, fis, envR, oqxA, rarA | wzc, wzi |
| D1598/19 | 39 | ColRNAI IncA/C2 IncFIB IncFIB(pQil) | blaSHV-40 blaTEM-1B | blaKPC-2 blaVIM-1 | aac(3)-Iid aac(6′)-Il aac(6′)-Im ant(3)-Ia aph(2″)-Ib aph(3′)-Ia | qnrS1, aac(6′)-lb-cr, oqxA oqxB | fosA, sul1, sul3, dfrA1 | ND | ND | wzi |
| Z852/19 | 307 | IncA/C2 IncFIB(pQil) | blaSHV-28 | blaKPC-2 blaVIM-1 | aac(6′)-Il aadA24 | oqxA oqxB | fosA, sul1, dfrA1, frA14 | fyuA, irp1, irp2 iutA, mrkABCDFHIJ, ybtAEPQSTX | acrABR, marAR, soxSR, ramAR, rob, sdiA, fis, envR, oqxABR, rarA | wzc, wzi |
| Z866/19 | 323 | IncFIB(K) IncFIB(Mar) IncFIB (pKPHS1) IncFIB(pQil) IncFII(K) IncHI1B | blaSHV-99 | blaKPC-2 blaVIM-1 | aadA1 | oqxA oqxB | fosA, fosA7, Sul1 | mrkABCDFHIJ | acrAR, marA, soxSR, ramA, rob, sdiA, fis, envR, oqxAR, rarA | wzc, wzi |
| D2452/19 | 39 | ColRNAI IncA/C2 IncFIB(pQil) | blaSHV-79 blaTEM-1B | blaKPC-2 blaVIM-1 | aac(3)-Iid aac(6′)-Il aac(6′)-Im ant(3)-Ia aph(2″)-Ib aph(3′)-Ia | qnrS1, aac(6′)-lb-cr, oqxA oqxB | fosA, sul1, Sul4, dfrA1 | fyuA, irp1, irp2 mrkABCDFHIJ, ybtAEPQSTX | acrBR, marAR, soxSR, ramA, rob, sdiA, fis, envR, oqxABR, rarA | wzc, wzi |
| C833/21 | 11 | Col(pHAD2) IncFIA(HI1) IncFIB(K) IncFII(K) | blaSHV-182 | blaOXA-48 blaNDM-1 | aph(3′)-Ia aac(6′)-lb-cr | aac(6′)-lb-cr, oqxA, oqxB | fosA, sul2, dfrA14, catB3 | mrkABCDFHIJ | acrABR, marA, soxSR, ramAR, rob, sdiA, fis, envR, rarA | wzc, wzi |
| A5051/21 | 15 | IncFIA(HI1) IncFIB(K) IncFII(K) | blaSHV-28 blaSHV-106 | blaNDM-1 | aac(6′)-lb-cr | aac(6′)-lb-cr oqxA | fosA catB3 | fyuA, kfuC, mrkABCDFHIJ ybtQU | acrABR, marA, soxSR, ramAR, rob, sdiA, fis, envR, rarA | wzcwzi |
| D2856/21 | 11 | Col440II IncFIA(HI1) IncFIB(K) IncFII(K) IncR | blaSHV-182 | blaOXA-48 blaNDM-1 | aph(3′)-Ia aac(6′)-lb-cr | oqxA | fosA, sul2, dfrA14, catB3, tet(A) | mrkABCDJ, ybtAPQS | acrABR, marAR, soxSR, ramAR, rob, sdiA, fis, envR, oqxAR, rarA | wzcwzi |
| C1909/21 | 39 | ColRNAI IncFII(K) | blaSHV-40, -79, -85, -89 | blaKPC-33 | aac(6′)-Ib aac(6′)-lb-cr | aac(6′)-lb-cr, oqxA, oqxB | fosA, dfrA12 tet(A) | mrkABCDFHIJ ybtEPQSTU | acrA, marAR, soxSR, ramAR, rob, sdiA, fis, envR, oqxAR, rarA | wzcwzi |
| A18940/21 | 512 | IncFIB(K) IncFII(K) IncN IncX3 | blaSHV-182 blaOXA-10 | blaKPC-2, blaNDM-1 | aac(6′)-Ib-cr, aac(6′)-Ib, aph(3′)-Ia, aadA2, aadA16 | qnrS1, aac(6′)-lb-cr, oqxA, oqxB | fosA, sul1, dfrA12, dfrA27 catA1 | iutA, mrkABCHI | acrR, marAR soxRS, ramA, rob, sdiA, fis, envR, oqxR, rarA | wzi |
| A1746/22 | 35 | IncC, IncR, IncFIA(HI1) IncFIB(K) IncFIB (pKPHS1) IncFIB(pQil) IncFII(K) | blaSHV-33 blaOXA-10 blaTEM-1B blaVEB-25 blaDHA-1 | blaKPC-2 | ant(2″)-Ia, aph(3″)-Ib aph(6)-Id, rmtB,aadA1 | qnrB4 oqxA, oqxB | fosA, sul1 sul2, catA1, cmlA1, tet(A), tet(G) | kfuA, mrkAFHI, ybtEQTX | acrR, marAR, soxRS, ramA rob, sdiA, fis, envR, oqxR rarA | wzc, wzi |
| A7213/22 | 11 | ColRNAI IncC IncFIA(HI1) IncFIB(K) | blaOXA-1 blaOXA-10 blaCTX-M -15 blaTEM-1B blaVEB-1 | blaNDM-1 | aac(6′)-Ib, aac(3)-IIa, aac(6′)-Ib-cr, ant(2″)-Ia, aph(3′)-Ia, rmtB | aac(6′)-lb-cr, oqxA, oqxB | fosA, sul2, dfrA14, tet(A) | mrkAC | soxSR, ramAR, sdiA | wzi |
| A9974/22 | 11 | ColRNAI IncC IncFIA(HI1) IncFIB(K) IncFII(K) | blaSHV-159 blaOXA-1 blaOXA-10 blaCTX-M-15 blaTEM-1B blaVEB-1 | blaNDM-1 | aac(6′)-Ib-cr aac(6′)-Ib aac(3)-IIa aph(6)-Id aph(3′)-Ia aph(3″)-Ib ant(2″)-Ia aadA1, rmtB | aac(6′)-lb-cr, oqxA, oqxB | fosA, sul2, dfrA14, tet(A), tet(G) | fyuA, mrkABCDFHIJ, ybtAEPQST | acrA, marAR soxR, ramAR rob, sdiA, fis, envR, oqxAB, rarA | wzi |
| A10037/22 | 15 | IncC IncFIA(HI1) IncFIB(K) IncFII(K) | blaSHV-100 blaOXA-1 blaOXA-10 blaCTX-M-15 blaTEM-1B blaVEB-1 | blaNDM-1 | aac(6′)-Ib-cr aac(6′)-Ib aac(3)-IIa aph(3″)-Ib aph(6)-Id ant(2″)-Ia aadA1, rmtB | aac(6′)-lb-cr, oqxA, oqxB | fosA, sul2 dfrA14, cmlA1, tet(A), tet(G) | fyuA, kfuAC, mrkABCFHJ, ybtAEPQSTU | acrAR, marAR soxSR, rob, sdiA fis, envR, oqxAR rarA | wzi |
| A10143/22 | 11 | ColRNAI IncFII(K) | blaOXA-1 blaOXA-10 blaCTX-M-15 blaTEM-1B blaVEB-1 | blaNDM-1 | aac(6′)-Ib-cr aac(6′)-Ib aac(3)-IIa aph(3″)-Ib aph(6)-Id ant(2″)-Ia aadA1, rmtB | aac(6′)-lb-cr, oqxA, oqxB | fosA, sul1, sul2, dfrA12, dfrA14, cmlA1, tet(A), tet(G) | fyuA, mrkH, ybtEPQSX | marA, soxSR, ramR, sdiA, fis, envR | wzi |
| Strain | Date of Isolation | Sex | Age (Years) | Ward | Biological Sample | Infection/Colonization |
|---|---|---|---|---|---|---|
| C2482/18 | 9 June 2018 | F | 8 | PICU | Rectal swab | Colonization |
| C4112/18 | 7 September 2018 | F | 56 | ICU | Rectal swab | Colonization |
| B11395/18 | 24 October 2018 | F | 1 | PICU | Urine | Infection |
| D6184/18 | 19 November 2018 | F | 8 | PICU | Bronchial aspirate | Infection |
| C251/19 | 17 January 2019 | F | 17 | PICU | Rectal swab | Colonization |
| Z557/19 | 30 January 2019 | M | 57 | ICU | Rectal swab | Colonization |
| C746/19 | 7 February 2019 | M | 73 | ICU | Rectal swab | Colonization |
| B2562/19 | 4 March 2019 | M | 79 | ICU | Urine | Infection |
| Z508/19 | 20 March 2019 | F | 60 | ICU | Rectal swab | Colonization |
| D1463/19 | 22 March 2019 | M | 55 | ICU | Bronchial aspirate | Infection |
| D1598/19 | 1 April 2019 | M | 40 | ICU | Bronchial aspirate | Infection |
| Z852/19 | 12 April 2019 | M | 3 | PICU | Rectal swab | Colonization |
| Z866/19 | 15 April 2019 | M | 49 | ICU | Rectal swab | Colonization |
| D2452/19 | 24 May 2019 | M | 73 | ICU | Bronchial aspirate | Infection |
| C833/21 | 31 March 2021 | F | 57 | ICU | Wound | Infection |
| A5051/21 | 25 April 2021 | F | 69 | ICU | Blood | Infection |
| D2856/21 | 7 July 2021 | F | 54 | ICU | Bronchial aspirate | Infection |
| C1909/21 | 8 July 2021 | M | 85 | ICU | Wound | Infection |
| A18940/21 | 24 December 2021 | F | 58 | ICU | Blood | Infection |
| A1746/22 | 25 February 2022 | M | 0.25 | PICU | Blood | Infection |
| A7213/22 | 3 May 2022 | F | 30 | ICU | Blood | Infection |
| A9974/22 | 16 June 2022 | F | 67 | ICU | Blood | Infection |
| A10037/22 | 16 June 2022 | F | 53 | ICU | Blood | Infection |
| A10143/22 | 19 June 2022 | M | 88 | ICU | Blood | Infection |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarras, C.; Karampatakis, T.; Pappa, S.; Iosifidis, E.; Vagdatli, E.; Roilides, E.; Papa, A. Genetic Characterization of Carbapenem-Resistant Klebsiella pneumoniae Clinical Isolates in a Tertiary Hospital in Greece, 2018–2022. Antibiotics 2023, 12, 976. https://doi.org/10.3390/antibiotics12060976
Zarras C, Karampatakis T, Pappa S, Iosifidis E, Vagdatli E, Roilides E, Papa A. Genetic Characterization of Carbapenem-Resistant Klebsiella pneumoniae Clinical Isolates in a Tertiary Hospital in Greece, 2018–2022. Antibiotics. 2023; 12(6):976. https://doi.org/10.3390/antibiotics12060976
Chicago/Turabian StyleZarras, Charalampos, Theodoros Karampatakis, Styliani Pappa, Elias Iosifidis, Eleni Vagdatli, Emmanuel Roilides, and Anna Papa. 2023. "Genetic Characterization of Carbapenem-Resistant Klebsiella pneumoniae Clinical Isolates in a Tertiary Hospital in Greece, 2018–2022" Antibiotics 12, no. 6: 976. https://doi.org/10.3390/antibiotics12060976