
A New Integrative and Mobilizable Element Is a Major Contributor to Tetracycline Resistance in Streptococcus dysgalactiae subsp. equisimilis
 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
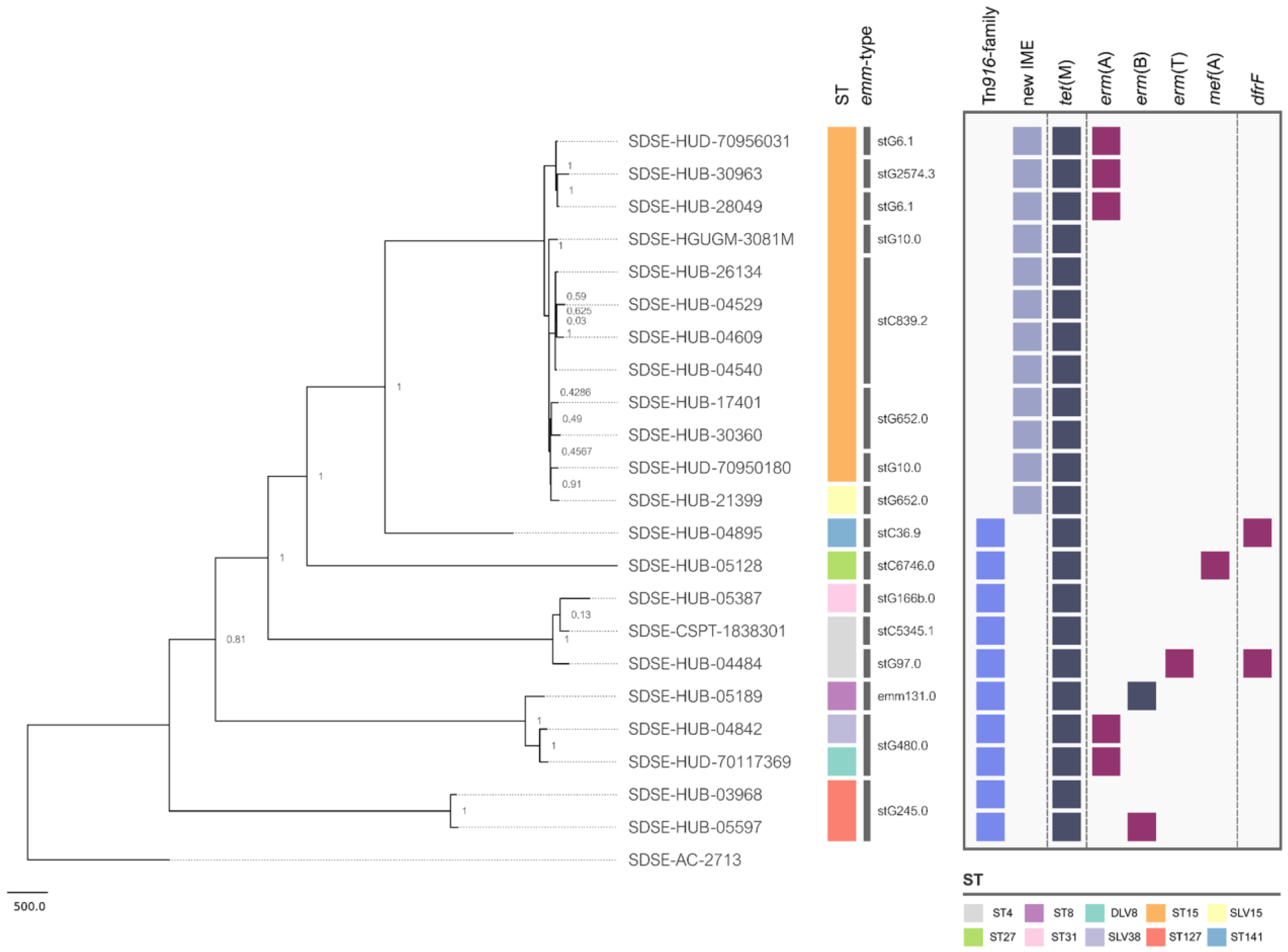
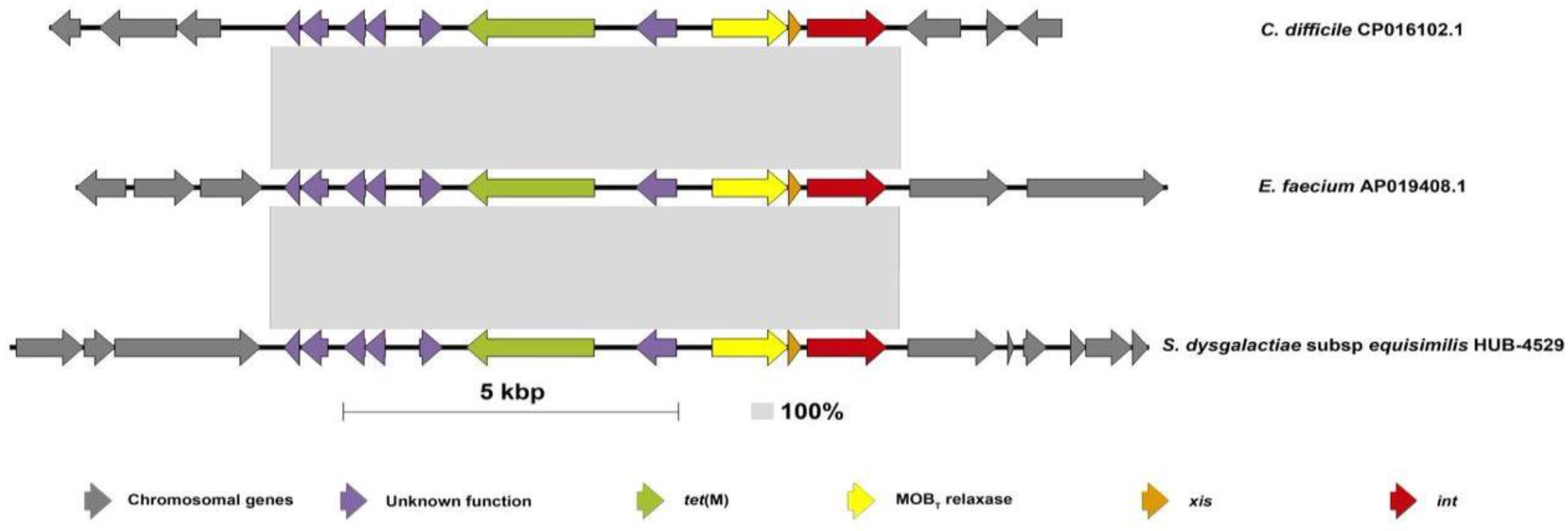
2. Results and Discussion
3. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vandamme, P.; Pot, B.; Falsen, E.; Kersters, K.; Devriese, L.A. Taxonomic Study of Lancefield Streptococcal Groups C, G, and L (Streptococcus dysgalactiae) and Proposal of S. dysgalactiae subsp. equisimilis subsp. Nov. Int. J. Syst. Bacteriol. 1996, 46, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Efstratiou, A. Pyogenic Streptococci of Lancefield Groups C and G as Pathogens in Man. J. Appl. Microbiol. 1997, 83, 72S–79S. [Google Scholar] [CrossRef]
- Brandt, C.M.; Spellerberg, B. Human Infections Due to Streptococcus dysgalactiae subspecies equisimilis. Clin. Infect. Dis. 2009, 49, 766–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, A.; Kilian, M. Delineation of Streptococcus dysgalactiae, Its Subspecies, and Its Clinical and Phylogenetic Relationship to Streptococcus pyogenes. J. Clin. Microbiol. 2012, 50, 113–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambertsen, L.M.; Ingels, H.; Schønheyder, H.C.; Hoffmann, S.; Hoffmann, S.; Ingels, H.; Lambertsen, L.; Christensen, J.J.; Dessau, R.; Lomborg, S.; et al. Nationwide Laboratory-Based Surveillance of Invasive Beta-Haemolytic Streptococci in Denmark from 2005 to 2011. Clin. Microbiol. Infect. 2014, 20, O216–O223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anonymous. Voluntary Surveillance of Pyogenic and Non-Pyogenic Streptococcal Bacteraemia in England, Wales and Northern Ireland: 2020. Health Prot. Rep. 2021, 15, 1–23. [Google Scholar]
- Wajima, T.; Morozumi, M.; Hanada, S.; Sunaoshi, K.; Chiba, N.; Iwata, S.; Ubukata, K. Molecular Characterization of Invasive Streptococcus dysgalactiae subsp. equisimilis, Japan. Emerg. Infect. Dis. 2016, 22, 247–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rantala, S.; Vuopio-Varkila, J.; Vuento, R.; Huhtala, H.; Syrjänen, J. Clinical Presentations and Epidemiology of β-Haemolytic Streptococcal Bacteraemia: A Population-Based Study. Clin. Microbiol. Infect. 2009, 15, 286–288. [Google Scholar] [CrossRef] [Green Version]
- Beres, S.B.; Zhu, L.; Pruitt, L.; Olsen, R.J.; Faili, A.; Kayal, S.; Musser, J.M. Integrative Reverse Genetic Analysis Identifies Polymorphisms Contributing to Decreased Antimicrobial Agent Susceptibility in Streptococcus pyogenes. mBio 2022, 13, e0361821. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline Antibiotics: Mode of Action, Applications, Molecular Biology, and Epidemiology of Bacterial Resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [Green Version]
- Guédon, G.; Libante, V.; Coluzzi, C.; Payot, S.; Leblond-Bourget, N. The Obscure World of Integrative and Mobilizable Elements, Highly Widespread Elements That Pirate Bacterial Conjugative Systems. Genes 2017, 8, 337. [Google Scholar] [CrossRef] [Green Version]
- Libante, V.; Nombre, Y.; Coluzzi, C.; Staub, J.; Guédon, G.; Gottschalk, M.; Teatero, S.; Fittipaldi, N.; Leblond-bourget, N.; Payot, S. Chromosomal Conjugative and Mobilizable Elements in Streptococcus suis: Major Actors in the Spreading of Antimicrobial Resistance and Bacteriocin Synthesis Genes. Pathogens 2019, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Liang, P.; Wang, M.; Gottschalk, M.; Vela, A.I.; Estrada, A.A.; Wang, J.; Du, P.; Luo, M.; Zheng, H.; Wu, Z. Genomic and Pathogenic Investigations of Streptococcus suis Serotype 7 Population Derived from a Human Patient and Pigs. Emerg. Microbes. Infect. 2021, 10, 1960–1974. [Google Scholar] [CrossRef]
- Wang, J.; Qi, K.; Bai, X.; Wu, Z.; Kang, W.; Liang, P.; Zheng, H.; Xu, J. Characterization of Integrative and Conjugative Elements Carrying Antibiotic Resistance Genes of Streptococcus suis Isolated in China. Front. Microbiol. 2022, 13, 1074844. [Google Scholar]
- Lu, B.; Fang, Y.; Huang, L.; Diao, B.; Du, X.; Kan, B.; Cui, Y.; Zhu, F.; Li, D.; Wang, D. Molecular Characterization and Antibiotic Resistance of Clinical Streptococcus dysgalactiae subsp. equisimilis in Beijing, China. Infect. Genet. Evol. 2016, 40, 119–125. [Google Scholar] [CrossRef]
- Rojo-Bezares, B.; Toca, L.; Manuel Azcona-Gutiérrez, J.; Ortega-Unanue, N.; Toledano, P.; Sáenz, Y. Streptococcus dysgalactiae subsp. equisimilis from Invasive and Non-Invasive Infections in Spain: Combining Epidemiology, Molecular Characterization, and Genetic Diversity. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1013–1021. [Google Scholar]
- Mcguinness, W.A.; Malachowa, N.; Deleo, F.R. Vancomycin Resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar]
- Gherardi, G.; Imperi, M.; Palmieri, C.; Magi, G.; Facinelli, B.; Baldassarri, L.; Pataracchia, M.; Creti, R. Genetic Diversity and Virulence Properties of Streptococcus dysgalactiae subsp. equisimilis from Different Sources. J. Med. Microbiol. 2013, 63, 90–98. [Google Scholar]
- de Souza, J.P.; Santos, A.R.; de Paula, G.R.; Barros, R.R. Antimicrobial Susceptibility and Genetic Relationships among Streptococcus dysgalactiae subsp. equisimilis Isolates in Rio de Janeiro. Infect. Dis. 2016, 48, 676–681. [Google Scholar] [CrossRef]
- Leitner, E.; Zollner-Schwetz, I.; Zarfel, G.; Masoud-Landgraf, L.; Gehrer, M.; Wagner-Eibel, U.; Grisold, A.J.; Feierl, G. Prevalence of Emm Types and Antimicrobial Susceptibility of Streptococcus dysgalactiae subsp. equisimilis in Austria. Int. J. Med. Microbiol. 2015, 305, 918–924. [Google Scholar] [CrossRef]
- Traverso, F.; Blanco, A.; Villalón, P.; Beratz, N.; Sáez Nieto, J.A.; Lopardo, H. Molecular Characterization of Invasive Streptococcus dysgalactiae subsp. equisimilis. Multicenter Study: Argentina 2011–2012. Rev. Argent. Microbiol. 2016, 48, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Loubinoux, J.; Plainvert, C.; Collobert, G.; Touak, G.; Bouvet, A.; Poyart, C. Adult Invasive and Noninvasive Infections Due to Streptococcus dysgalactiae subsp. equisimilis in France from 2006 to 2010. J. Clin. Microbiol. 2013, 51, 2724–2727. [Google Scholar] [CrossRef] [Green Version]
- Ambroset, C.; Coluzzi, C.; Guédon, G.; Devignes, M.D.; Loux, V.; Lacroix, T.; Payot, S.; Leblond-Bourget, N. New Insights into the Classification and Integration Specificity of Streptococcus Integrative Conjugative Elements through Extensive Genome Exploration. Front. Microbiol. 2016, 6, 1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, L.B. Tn916 Family Conjugative Transposons and Dissemination of Antimicrobial Resistance Determinants. Antimicrob. Agents Chemother. 1998, 42, 1871–1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Croucher, N.J.; Page, A.J.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid Phylogenetic Analysis of Large Samples of Recombinant Bacterial Whole Genome Sequences Using Gubbins. Nucleic Acids Res. 2015, 43, e15. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A Fast, Scalable and User-friendly Tool for Maximum Likelihood Phylogenetic Inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Antibiotic | % Susceptible | MIC (50) [mg/L] | MIC (90) [mg/L] |
|---|---|---|---|
| Penicillin | 100 | <0.03 | <0.03 |
| Cefotaxime | 100 | <0.06 | <0.03 |
| Erythromycin | 54.2 | <0.25 | >4 |
| Clindamycin | 58.3 | <0.25 | >0.5 |
| Linezolid | 100 | <1 | - |
| Tetracycline | 0 | - | >4 |
| Chloramphenicol | 100 | 2 | 2 |
| Levofloxacin | 100 * | <1 | <1 |
| Vancomycin | 100 | <0.5 | <0.5 |
| Trimethoprim-Sulfamethoxazole | 91.7 | <0.5/9.5 | <0.5/9.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López de Egea, G.; González-Díaz, A.; Guédon, G.; Lao, J.; Berbel, D.; Casabella, A.; Marimón, J.M.; Cercenado, E.; Fernández-Delgado, L.; Chiapello, H.; et al. A New Integrative and Mobilizable Element Is a Major Contributor to Tetracycline Resistance in Streptococcus dysgalactiae subsp. equisimilis. Antibiotics 2023, 12, 579. https://doi.org/10.3390/antibiotics12030579
López de Egea G, González-Díaz A, Guédon G, Lao J, Berbel D, Casabella A, Marimón JM, Cercenado E, Fernández-Delgado L, Chiapello H, et al. A New Integrative and Mobilizable Element Is a Major Contributor to Tetracycline Resistance in Streptococcus dysgalactiae subsp. equisimilis. Antibiotics. 2023; 12(3):579. https://doi.org/10.3390/antibiotics12030579
Chicago/Turabian StyleLópez de Egea, Guillem, Aida González-Díaz, Gérard Guédon, Julie Lao, Dàmaris Berbel, Antonio Casabella, José María Marimón, Emilia Cercenado, Lucía Fernández-Delgado, Hélène Chiapello, and et al. 2023. "A New Integrative and Mobilizable Element Is a Major Contributor to Tetracycline Resistance in Streptococcus dysgalactiae subsp. equisimilis" Antibiotics 12, no. 3: 579. https://doi.org/10.3390/antibiotics12030579