

A Bioluminescence-Based Drug Screen Identifies Activities of Fexinidazole and Its Metabolites against Helicobacter pylori
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Viability Assay for H. pylori
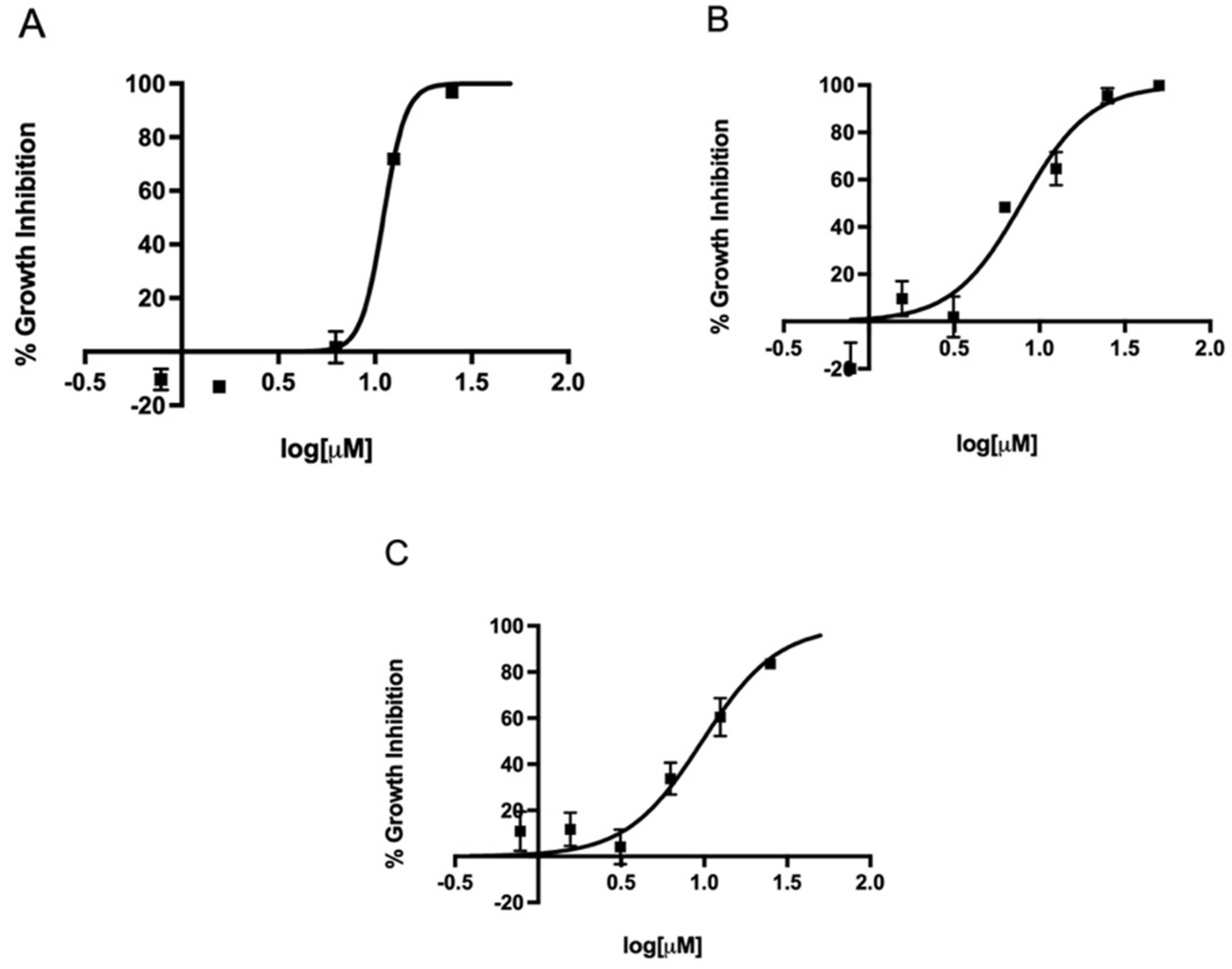
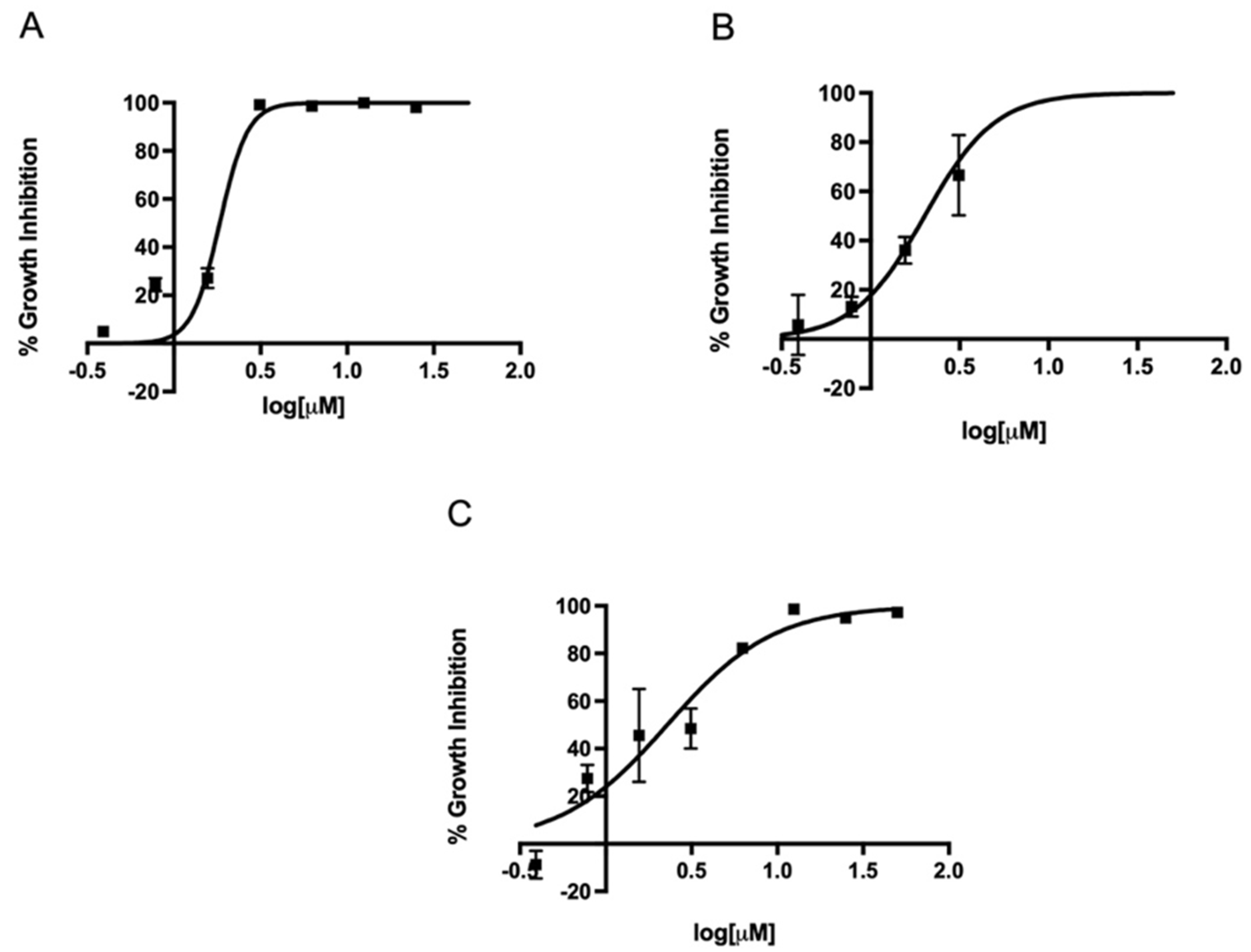
2.2. In Vitro Activity of Metronidazole, Fexinidazole, Fexinidazole Sulfoxide, and Fexinidazole Sulfone against Various H. pylori Strains
2.3. MIC Determination of Metronidazole, Fexinidazole, Fexinidazole Sulfoxide, and Fexinidazole Sulfone against H. pylori Strains
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. H. pylori Bacterial Culture
4.3. H. pylori Strain Viability Assay
4.4. In Vitro Activity of Metronidazole, Fexinidazole, Fexinidazole Sulfoxide, and Fexinidazole Sulfone against Various H. pylori Strains
4.5. MIC Determination of Metronidazole, Fexinidazole, Fexinidazole Sulfoxide, and Fexinidazole Sulfone against Various H. pylori Strains
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peek, R.M., Jr.; Blaser, M.J. Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat. Rev. Cancer 2002, 2, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.G.; Wang, T.C. Inflammation, atrophy, and gastric cancer. J. Clin. Investig. 2007, 117, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Peterson, W.L. Helicobacter pylori and peptic ulcer disease. N. Engl. J. Med. 1991, 324, 1043–1048. [Google Scholar] [CrossRef]
- Suerbaum, S.; Michetti, P. Helicobacter pylori infection. N. Engl. J. Med. 2002, 347, 1175–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wotherspoon, A.C.; Doglioni, C.; Diss, T.C.; Pan, L.; Moschini, A.; de Boni, M.; Isaacson, P.G. Regression of primary low-grade B-cell gastric lymphoma of mucosa-associated lymphoid tissue type after eradication of Helicobacter pylori. Lancet 1993, 342, 575–577. [Google Scholar] [CrossRef]
- McBee, M.E.; Zheng, P.Z.; Rogers, A.B.; Fox, J.G.; Schauer, D.B. Modulation of acute diarrheal illness by persistent bacterial infection. Infect. Immun. 2008, 76, 4851–4858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zajacova, A.; Dowd, J.B.; Aiello, A.E. Socioeconomic and race/ethnic patterns in persistent infection burden among U.S. adults. J. Gerontol. A Biol. Sci. Med. Sci. 2009, 64, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Furuta, T.; Graham, D.Y. Pharmacologic aspects of eradication therapy for Helicobacter pylori Infection. Gastroenterol. Clin. N. Am. 2010, 39, 465–480. [Google Scholar] [CrossRef]
- Gisbert, J.P.; Pajares, J.M. Treatment of Helicobacter pylori infection: The past and the future. Eur. J. Intern. Med. 2010, 21, 357–359. [Google Scholar] [CrossRef]
- Gisbert, J.P.; Pajares, R.; Pajares, J.M. Evolution of Helicobacter pylori therapy from a meta-analytical perspective. Helicobacter 2007, 12 (Suppl. 2), 50–58. [Google Scholar] [CrossRef]
- Bruce, M.G.; Maaroos, H.I. Epidemiology of Helicobacter pylori infection. Helicobacter 2008, 13 (Suppl. 1), 1–6. [Google Scholar] [CrossRef] [PubMed]
- D’Elios, M.M.; Andersen, L.P. Helicobacter pylori inflammation, immunity, and vaccines. Helicobacter 2007, 12 (Suppl. 1), 15–19. [Google Scholar] [CrossRef] [PubMed]
- McMahon, B.J.; Hennessy, T.W.; Bensler, J.M.; Bruden, D.L.; Parkinson, A.J.; Morris, J.M.; Reasonover, A.L.; Hurlburt, D.A.; Bruce, M.G.; Sacco, F.; et al. The relationship among previous antimicrobial use, antimicrobial resistance, and treatment outcomes for Helicobacter pylori infections. Ann. Intern. Med. 2003, 139, 463–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frenck, R.W., Jr.; Clemens, J. Helicobacter in the developing world. Microbes Infect. 2003, 5, 705–713. [Google Scholar] [CrossRef]
- Gisbert, J.P. The recurrence of Helicobacter pylori infection: Incidence and variables influencing it. A critical review. Am. J. Gastroenterol. 2005, 100, 2083–2099. [Google Scholar] [CrossRef]
- Parsonnet, J. What is the Helicobacter pylori global reinfection rate? Can. J. Gastroenterol. 2003, 17 (Suppl. B), 46B–48B. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Sampaio, P.S.; Calado, C.R.C. Potential of FTIR-Spectroscopy for Drugs Screening against Helicobacter pylori. Antibiotics 2020, 9, 897. [Google Scholar] [CrossRef]
- Sousa Sampaio, P.N.; Calado, C.R.C. Antimicrobial evaluation of the Cynara cardunculus extract in Helicobacter pylori cells using mid-infrared spectroscopy and chemometric methods. J. Appl. Microbiol. 2022, 133, 1743–1756. [Google Scholar] [CrossRef]
- Debnath, A.; Parsonage, D.; Andrade, R.M.; He, C.; Cobo, E.R.; Hirata, K.; Chen, S.; Garcia-Rivera, G.; Orozco, E.; Martinez, M.B.; et al. A high-throughput drug screen for Entamoeba histolytica identifies a new lead and target. Nat. Med. 2012, 18, 956–960. [Google Scholar] [CrossRef]
- Mullard, A. FDA approves first all-oral sleeping sickness drug. Nat. Rev. Drug Discov. 2021, 20, 658. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.Y. Efficient identification and evaluation of effective Helicobacter pylori therapies. Clin. Gastroenterol. Hepatol. 2009, 7, 145–148. [Google Scholar] [CrossRef] [Green Version]
- Laheij, R.J.; Rossum, L.G.; Jansen, J.B.; Straatman, H.; Verbeek, A.L. Evaluation of treatment regimens to cure Helicobacter pylori infection—A meta-analysis. Aliment. Pharmacol. Ther. 1999, 13, 857–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laine, L.; Fennerty, M.B.; Osato, M.; Sugg, J.; Suchower, L.; Probst, P.; Levine, J.G. Esomeprazole-based Helicobacter pylori eradication therapy and the effect of antibiotic resistance: Results of three US multicenter, double-blind trials. Am. J. Gastroenterol. 2000, 95, 3393–3398. [Google Scholar] [CrossRef] [PubMed]
- Megraud, F. H. pylori antibiotic resistance: Prevalence, importance, and advances in testing. Gut 2004, 53, 1374–1384. [Google Scholar] [CrossRef] [Green Version]
- Paoluzi, O.A.; Visconti, E.; Andrei, F.; Tosti, C.; Lionetti, R.; Grasso, E.; Ranaldi, R.; Stroppa, I.; Pallone, F. Ten and eight-day sequential therapy in comparison to standard triple therapy for eradicating Helicobacter pylori infection: A randomized controlled study on efficacy and tolerability. J. Clin. Gastroenterol. 2010, 44, 261–266. [Google Scholar] [CrossRef]
- Vakil, N.; Lanza, F.; Schwartz, H.; Barth, J. Seven-day therapy for Helicobacter pylori in the United States. Aliment. Pharmacol. Ther. 2004, 20, 99–107. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Graham, D.Y.; Lu, H. Should triple therapy for Helicobacter pylori infection be abandoned as no longer effective? US Gastroenterol. 2008, 4, 65–67. [Google Scholar]
- Yamaoka, Y.; Graham, D.Y. Empiric H. pylori therapy-10-day concomitant, bismuth quadruple or 14-day triple therapy: None is best. Transl. Cancer Res. 2016, 5, S1476–S1478. [Google Scholar] [CrossRef] [Green Version]
- Debnath, A.; Shahinas, D.; Bryant, C.; Hirata, K.; Miyamoto, Y.; Hwang, G.; Gut, J.; Renslo, A.R.; Pillai, D.R.; Eckmann, L.; et al. Hsp90 inhibitors as new leads to target parasitic diarrheal diseases. Antimicrob. Agents ChemoTher. 2014, 58, 4138–4144. [Google Scholar] [CrossRef] [Green Version]
- Mao, W.J.; Lv, P.C.; Shi, L.; Li, H.Q.; Zhu, H.L. Synthesis, molecular docking and biological evaluation of metronidazole derivatives as potent Helicobacter pylori urease inhibitors. Bioorg. Med. Chem. 2009, 17, 7531–7536. [Google Scholar] [CrossRef] [PubMed]
- Jarrad, A.M.; Ang, C.W.; Debnath, A.; Hahn, H.J.; Woods, K.; Tan, L.; Sykes, M.L.; Jones, A.J.; Pelingon, R.; Butler, M.S.; et al. Design, Synthesis, and Biological Evaluation of 2-Nitroimidazopyrazin-one/-es with Antitubercular and Antiparasitic Activity. J. Med. Chem. 2018, 61, 11349–11371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarrad, A.M.; Debnath, A.; Miyamoto, Y.; Hansford, K.A.; Pelingon, R.; Butler, M.S.; Bains, T.; Karoli, T.; Blaskovich, M.A.; Eckmann, L.; et al. Nitroimidazole carboxamides as antiparasitic agents targeting Giardia lamblia, Entamoeba histolytica and Trichomonas vaginalis. Eur. J. Med. Chem. 2016, 120, 353–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarrad, A.M.; Karoli, T.; Debnath, A.; Tay, C.Y.; Huang, J.X.; Kaeslin, G.; Elliott, A.G.; Miyamoto, Y.; Ramu, S.; Kavanagh, A.M.; et al. Metronidazole-triazole conjugates: Activity against Clostridium difficile and parasites. Eur. J. Med. Chem. 2015, 101, 96–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, Y.; Kalisiak, J.; Korthals, K.; Lauwaet, T.; Cheung, D.Y.; Lozano, R.; Cobo, E.R.; Upcroft, P.; Upcroft, J.A.; Berg, D.E.; et al. Expanded therapeutic potential in activity space of next-generation 5-nitroimidazole antimicrobials with broad structural diversity. Proc. Natl. Acad. Sci. USA 2013, 110, 17564–17569. [Google Scholar] [CrossRef] [Green Version]
- Torreele, E.; Bourdin Trunz, B.; Tweats, D.; Kaiser, M.; Brun, R.; Mazue, G.; Bray, M.A.; Pecoul, B. Fexinidazole--a new oral nitroimidazole drug candidate entering clinical development for the treatment of sleeping sickness. PLoS Negl. Trop. Dis. 2010, 4, e923. [Google Scholar] [CrossRef] [Green Version]
- Tarral, A.; Blesson, S.; Mordt, O.V.; Torreele, E.; Sassella, D.; Bray, M.A.; Hovsepian, L.; Evene, E.; Gualano, V.; Felices, M.; et al. Determination of an optimal dosing regimen for fexinidazole, a novel oral drug for the treatment of human African trypanosomiasis: First-in-human studies. Clin. Pharmacokinet. 2014, 53, 565–580. [Google Scholar] [CrossRef] [Green Version]
- Bkhaitan, M.M.; Alarjah, M.; Mirza, A.Z.; Abdalla, A.N.; El-Said, H.M.; Faidah, H.S. Preparation and biological evaluation of metronidazole derivatives with monoterpenes and eugenol. Chem. Biol. Drug Des. 2018, 92, 1954–1962. [Google Scholar] [CrossRef]
- Moshafi, M.H.; Sorkhi, M.; Emami, S.; Nakhjiri, M.; Yahya-Meymandi, A.; Negahbani, A.S.; Siavoshi, F.; Omrani, M.; Alipour, E.; Vosooghi, M.; et al. 5-Nitroimidazole-based 1,3,4-thiadiazoles: Heterocyclic analogs of metronidazole as anti-Helicobacter pylori agents. Arch. Pharm. 2011, 344, 178–183. [Google Scholar] [CrossRef]
- Sasahara, G.L.; Gouveia Junior, F.S.; Rodrigues, R.O.; Zampieri, D.S.; Fonseca, S.; Goncalves, R.C.R.; Athaydes, B.R.; Kitagawa, R.R.; Santos, F.A.; Sousa, E.H.S.; et al. Nitro-imidazole-based ruthenium complexes with antioxidant and anti-inflammatory activities. J. Inorg. Biochem. 2020, 206, 111048. [Google Scholar] [CrossRef]
- Lee, A.; O’Rourke, J.; De Ungria, M.C.; Robertson, B.; Daskalopoulos, G.; Dixon, M.F. A standardized mouse model of Helicobacter pylori infection: Introducing the Sydney strain. Gastroenterology 1997, 112, 1386–1397. [Google Scholar] [CrossRef]
- Baltrus, D.A.; Amieva, M.R.; Covacci, A.; Lowe, T.M.; Merrell, D.S.; Ottemann, K.M.; Stein, M.; Salama, N.R.; Guillemin, K. The complete genome sequence of Helicobacter pylori strain G27. J. Bacteriol. 2009, 191, 447–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peek, R.M., Jr.; Blaser, M.J.; Mays, D.J.; Forsyth, M.H.; Cover, T.L.; Song, S.Y.; Krishna, U.; Pietenpol, J.A. Helicobacter pylori strain-specific genotypes and modulation of the gastric epithelial cell cycle. Cancer Res. 1999, 59, 6124–6131. [Google Scholar] [PubMed]
- Jung, S.W.; Thamphiwatana, S.; Zhang, L.; Obonyo, M. Mechanism of antibacterial activity of liposomal linolenic acid against Helicobacter pylori. PLoS ONE 2015, 10, e0116519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obonyo, M.; Sabet, M.; Cole, S.P.; Ebmeyer, J.; Uematsu, S.; Akira, S.; Guiney, D.G. Deficiencies of myeloid differentiation factor 88, Toll-like receptor 2 (TLR2), or TLR4 produce specific defects in macrophage cytokine secretion induced by Helicobacter pylori. Infect. Immun. 2007, 75, 2408–2414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obonyo, M.; Zhang, L.; Thamphiwatana, S.; Pornpattananangkul, D.; Fu, V.; Zhang, L. Antibacterial activities of liposomal linolenic acids against antibiotic-resistant Helicobacter pylori. Mol. Pharm. 2012, 9, 2677–2685. [Google Scholar] [CrossRef] [Green Version]
- Shing, B.; Singh, S.; Podust, L.M.; McKerrow, J.H.; Debnath, A. The Antifungal Drug Isavuconazole Is both Amebicidal and Cysticidal against Acanthamoeba castellanii. Antimicrob. Agents ChemoTher. 2020, 64, e02223-19. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Strain | Mean (µM) | 95% Lower CL (µM) 1 | 95% Upper CL (µM) 1 |
|---|---|---|---|---|
| Fexinidazole | SS1 | 2 | 1.2 | 1.9 |
| 60190 | 2.9 | 2.1 | 3.4 | |
| G27 | 2.4 | 2 | 3.6 | |
| Fexinidazole sulfoxide | SS1 | 1.5 | 1.1 | 1.8 |
| 60190 | 1.9 | 1.8 | 2.5 | |
| G27 | 1.7 | 1.6 | 2.4 | |
| Fexinidazole sulfone | SS1 | 1.7 | 1.6 | 2.1 |
| 60190 | 1.8 | 1.6 | 2.5 | |
| G27 | 2.1 | 1.7 | 3.1 | |
| Metronidazole | SS1 | 11.1 | 6.1 | 19.9 |
| 60190 | 7.8 | 6.1 | 10.1 | |
| G27 | 9.3 | 6.1 | 15.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed, A.; Chilingerian, J.N.; Bali, P.; Obonyo, M.; Debnath, A. A Bioluminescence-Based Drug Screen Identifies Activities of Fexinidazole and Its Metabolites against Helicobacter pylori. Antibiotics 2022, 11, 1605. https://doi.org/10.3390/antibiotics11111605
Mohamed A, Chilingerian JN, Bali P, Obonyo M, Debnath A. A Bioluminescence-Based Drug Screen Identifies Activities of Fexinidazole and Its Metabolites against Helicobacter pylori. Antibiotics. 2022; 11(11):1605. https://doi.org/10.3390/antibiotics11111605
Chicago/Turabian StyleMohamed, Abdolhakim, John N Chilingerian, Prerna Bali, Marygorret Obonyo, and Anjan Debnath. 2022. "A Bioluminescence-Based Drug Screen Identifies Activities of Fexinidazole and Its Metabolites against Helicobacter pylori" Antibiotics 11, no. 11: 1605. https://doi.org/10.3390/antibiotics11111605