
Acinetobacter baylyi Strain BD413 Can Acquire an Antibiotic Resistance Gene by Natural Transformation on Lettuce Phylloplane and Enter the Endosphere

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
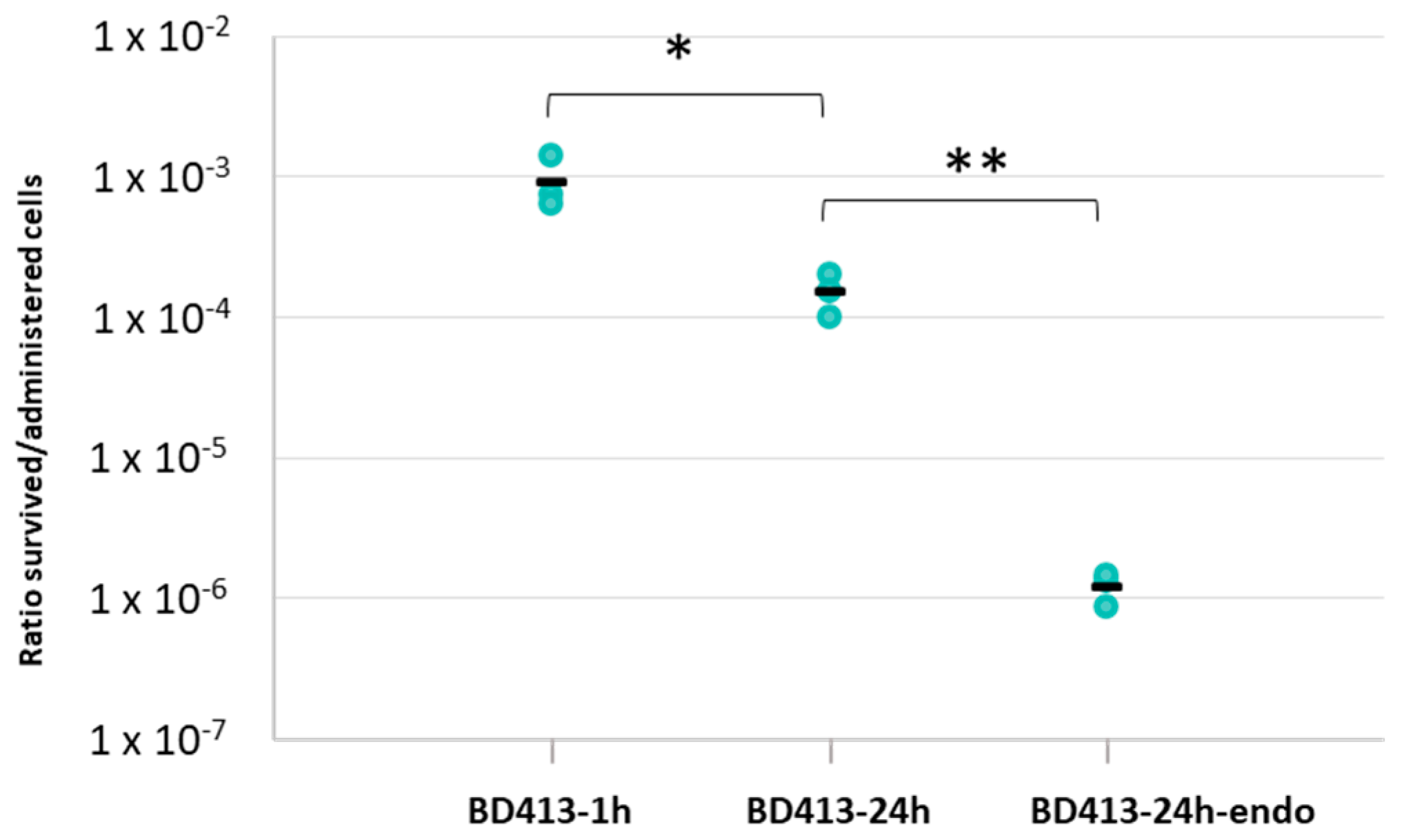
2.1. A. baylyi BD413 Permanence as Viable and Culturable Cells into Lettuce Leaves
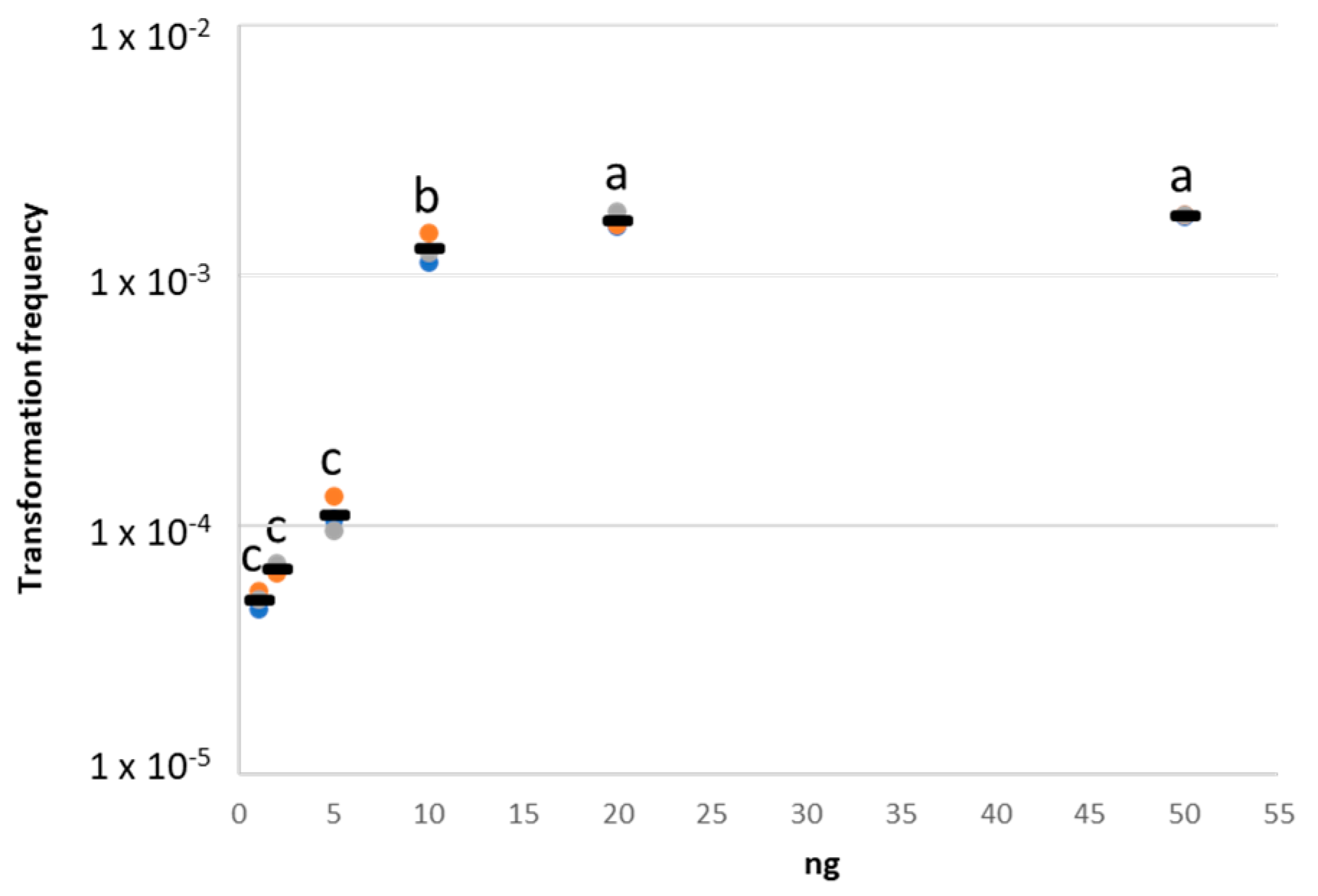
2.2. Natural Transformation on Nitrocellulose Membrane Filters
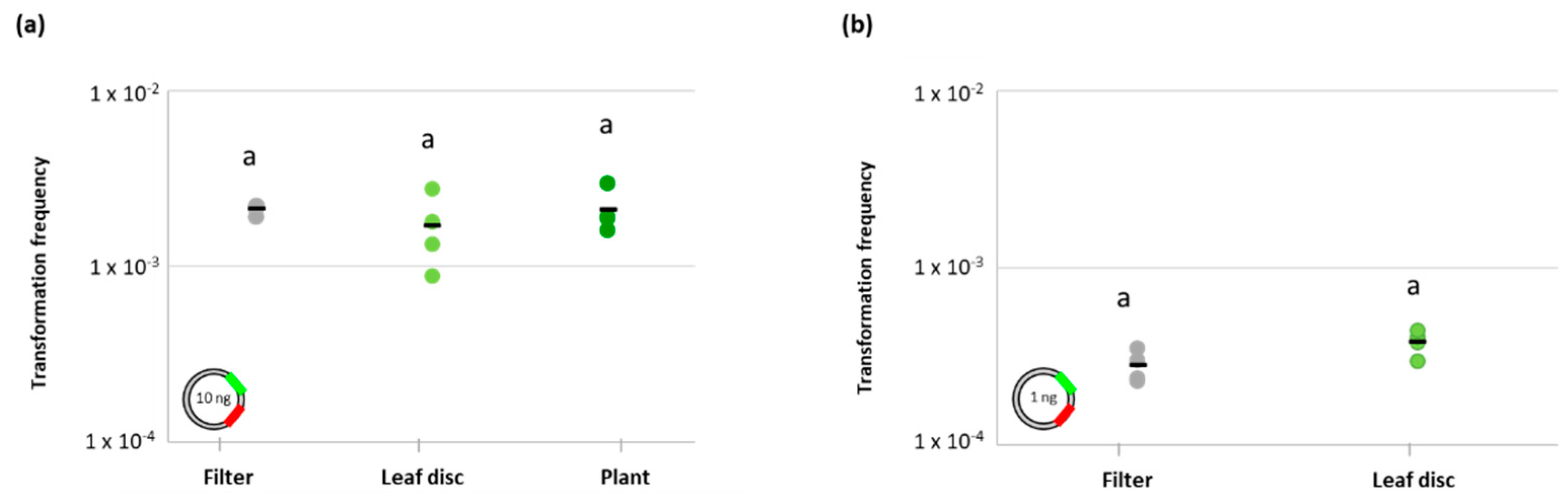
2.3. Natural Transformation on Lettuce Leaves
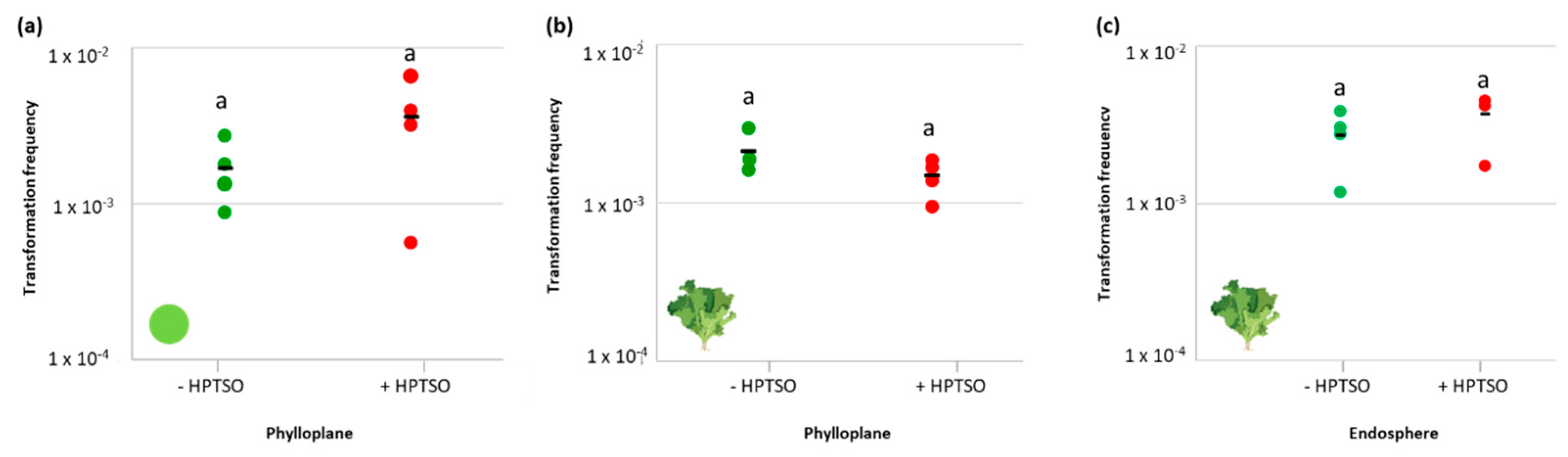
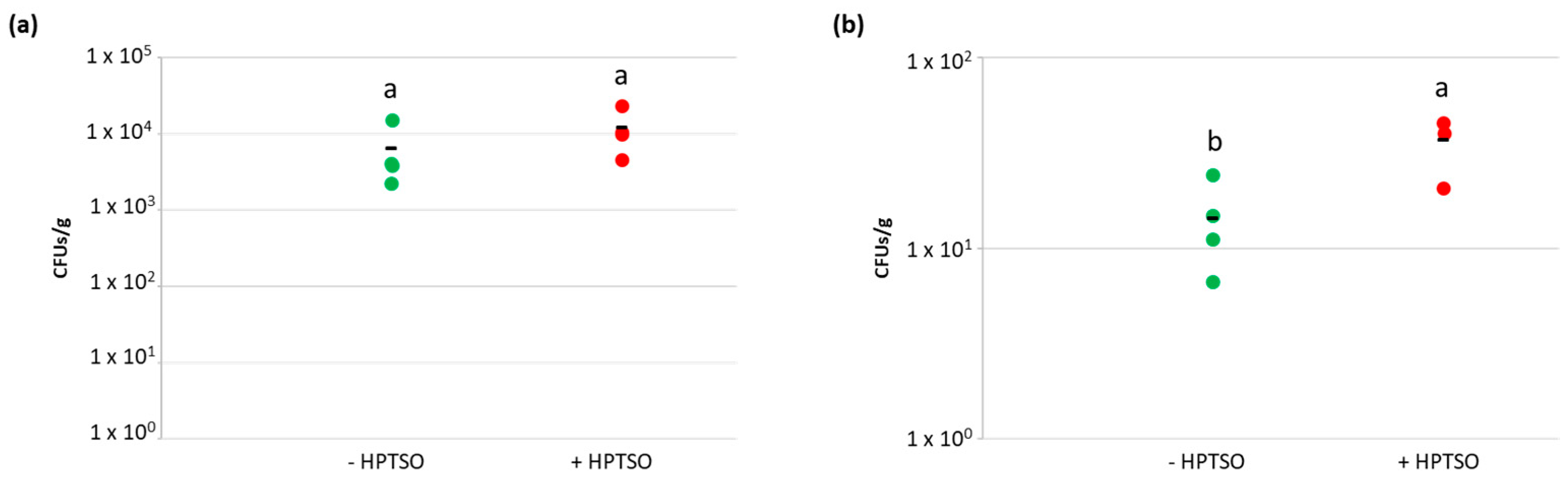
2.4. Effect of Surfactant Molecule on the Ability of A. baylyi BD413 to Acquire exDNA and Enter the Leaf Endosphere
3. Discussion
4. Materials and Methods
4.1. A. baylyi BD413 Survival Assay on Lettuce Phylloplane and into Leaf Endosphere
4.2. Natural Transformation Protocols
4.2.1. Bacterial Culture Preparation
4.2.2. Extracellular DNA (exDNA) Preparation
4.2.3. Selection of the exDNA Quantity for In Vivo Experiments
4.2.4. Natural Transformation on Leaf Disc
4.2.5. Natural Transformation in Planta
4.3. Entry of Total and Transformant A. baylyi BD413 Cells into Leaf Endosphere
4.4. Effect of a Surfactant Molecule on A. baylyi BD413 Transformation and Penetration into the Endosphere
4.5. Bacterial Cell Membrane Permeability Assays
4.6. Lettuce Leaf Acquisition of Extracellular DNA
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Larsson, D.G.J.; Flach, C.F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef]
- Westhoff, S.; van Leeuwe, T.M.; Qachach, O.; Zhang, Z.; van Wezel, G.P.; Rozen, D.E. The evolution of no-cost resistance at sub-MIC concentrations of streptomycin in Streptomyces coelicolor. ISME J. 2017, 11, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Antibiotic resistance in the environment: A link to the clinic? Curr. Opin. Microbiol. 2010, 13, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, J.; Engelstädter, J.; Zhang, S.; Ding, P.; Mao, L.; Yuan, Z.; Bond, P.L.; Guo, J. Non-antibiotic pharmaceuticals enhance the transmission of exogenous antibiotic resistance genes through bacterial transformation. ISME J. 2020, 14, 2179–2196. [Google Scholar] [CrossRef] [PubMed]
- Riva, V.; Riva, F.; Vergani, L.; Crotti, E.; Borin, S.; Mapelli, F. Microbial assisted phytodepuration for water reclamation: Environmental benefits and threats. Chemosphere 2020, 241, 124843. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-L.; Cui, H.-L.; Su, J.-Q.; Penuelas, J.; Zhu, Y.-G. Antibiotic Resistomes in Plant Microbiomes. Trends Plant Sci. 2019, 24, 530–541. [Google Scholar] [CrossRef]
- Scaccia, N.; Vaz-Moreira, I.; Manaia, C.M. The risk of transmitting antibiotic resistance through endophytic bacteria. Trends Plant Sci. 2021, 26, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Remus-Emsermann, M.N.P.; Aicher, D.; Pelludat, C.; Gisler, P.; Drissner, D. Conjugation dynamics of self-transmissible and mobilisable plasmids into e. Coli o157:H7 on Arabidopsis thaliana rosettes. Antibiotics 2021, 10, 1–14. [Google Scholar]
- Deng, B.; Li, W.; Lu, H.; Zhu, L. Film mulching reduces antibiotic resistance genes in the phyllosphere of lettuce. J. Environ. Sci. 2022, 112, 121–128. [Google Scholar] [CrossRef]
- Blau, K.; Bettermann, A.; Jechalke, S.; Fornefeld, E.; Vanrobaeys, Y.; Stalder, T.; Top, E.M.; Smalla, K. The transferable resistome of produce. mBio 2018, 9, e01300-18. [Google Scholar] [CrossRef]
- Von Wintersdorff, C.J.H.; Penders, J.; Van Niekerk, J.M.; Mills, N.D.; Majumder, S.; Van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarei-Baygi, A.; Smith, A.L. Intracellular versus extracellular antibiotic resistance genes in the environment: Prevalence, horizontal transfer, and mitigation strategies. Bioresour. Technol. 2021, 319, 124181. [Google Scholar] [CrossRef] [PubMed]
- de Berardinis, V.; Durot, M.; Weissenbach, J.; Salanoubat, M. Acinetobacter baylyi ADP1 as a model for metabolic system biology. Curr. Opin. Microbiol. 2009, 12, 568–576. [Google Scholar] [CrossRef]
- Borin, S.; Crotti, E.; Mapelli, F.; Tamagnini, I.; Corselli, C.; Daffonchio, D. DNA is preserved and maintains transforming potential after contact with brines of the deep anoxic hypersaline lakes of the Eastern Mediterranean Sea. Saline Syst. 2008, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Pontiroli, A.; Rizzi, A.; Simonet, P.; Daffonchio, D.; Vogel, T.M.; Monier, J.-M. Visual evidence of horizontal gene transfer between plants and bacteria in the phytosphere of transplastomic tobacco. Appl. Environ. Microbiol. 2009, 75, 3314–3322. [Google Scholar] [CrossRef]
- Munoz-Price, L.S.; Weinstein, R.A. Acinetobacter infection. N. Engl. J. Med. 2008, 358, 1271–1281. [Google Scholar] [CrossRef]
- Fulsundar, S.; Harms, K.; Flaten, G.E.; Johnsen, P.J.; Chopade, B.A.; Nielsen, K.M. Gene transfer potential of outer membrane vesicles of Acinetobacter baylyi and effects of stress on vesiculation. Appl. Environ. Microbiol. 2014, 80, 3469–3483. [Google Scholar] [CrossRef]
- Riva, V.; Mapelli, F.; Bagnasco, A.; Mengoni, A.; Borin, S. A meta-analysis approach to defining the culturable core of plant endophytic bacterial communities. Appl. Environ. Microbiol. 2022, 88, 1–10. [Google Scholar] [CrossRef]
- Carvalheira, A.; Silva, J.; Teixeira, P. Lettuce and fruits as a source of multidrug resistant Acinetobacter spp. Food Microbiol. 2017, 64, 119–125. [Google Scholar] [CrossRef]
- Brennan, F.P.; Alsanius, B.W.; Allende, A.; Burgess, C.M.; Moreira, H.; Johannessen, G.S.; Castro, P.M.L.; Uyttendaele, M.; Truchado, P.; Holden, N.J. Harnessing agricultural microbiomes for human pathogen control. ISME Commun. 2022, 2, 1–6. [Google Scholar] [CrossRef]
- Haelewaters, D.; Urbina, H.; Brown, S.; Newerth-Henson, S.; Aime, M. Isolation and molecular characterization of the romaine lettuce phylloplane mycobiome. J. Fungi 2021, 7, 277. [Google Scholar] [CrossRef]
- Maeusli, M.; Lee, B.; Miller, S.; Reyna, Z.; Lu, P.; Yan, J.; Ulhaq, A.; Skandalis, N.; Spellberg, B.; Luna, B. Horizontal gene transfer of antibiotic resistance from Acinetobacter baylyi to Escherichia coli on lettuce and subsequent antibiotic resistance transmission to the gut microbiome. mSphere 2020, 5, 1–7. [Google Scholar] [CrossRef]
- Mantilla-Calderon, D.; Plewa, M.J.; Michoud, G.; Fodelianakis, S.; Daffonchio, D.; Hong, P.Y. Water disinfection byproducts increase natural transformation rates of environmental DNA in Acinetobacter baylyi ADP1. Environ. Sci. Technol. 2019, 53, 6520–6528. [Google Scholar] [CrossRef]
- Zdarta, A.; Pacholak, A.; Smułek, W.; Zgoła-Grześkowiak, A.; Ferlin, N.; Bil, A.; Kovensky, J.; Grand, E.; Kaczorek, E. Biological impact of octyl d-glucopyranoside based surfactants. Chemosphere 2019, 217, 567–575. [Google Scholar] [CrossRef]
- Gu, G.; Hu, J.; Cevallos-Cevallos, J.M.; Richardson, S.M.; Bartz, J.A.; van Bruggen, A.H.C. Internal colonization of salmonella enterica serovar typhimurium in tomato plants. PLoS ONE 2011, 6, e27340. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Franco, D.; van Loosdrecht, M.C.M.; Abeel, T.; Weissbrodt, D.G. Free-floating extracellular DNA: Systematic profiling of mobile genetic elements and antibiotic resistance from wastewater. Water Res. 2021, 189, 116592. [Google Scholar] [CrossRef] [PubMed]
- Seitz, P.; Blokesch, M. Cues and regulatory pathways involved in natural competence and transformation in pathogenic and environmental Gram-negative bacteria. FEMS Microbiol. Rev. 2013, 37, 336–363. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Caparas, M.; Zobel, Z.; Castanho, A.D.A.; Schwalm, C.R. Increasing risks of crop failure and water scarcity in global breadbaskets by 2030. Environ. Res. Lett. 2021, 16, 104013. [Google Scholar] [CrossRef]
- Miłobedzka, A.; Ferreira, C.; Vaz-Moreira, I.; Calderón-Franco, D.; Gorecki, A.; Purkrtova, S.; Bartacek, J.; Dziewit, L.; Singleton, C.M.; Nielsen, P.H.; et al. Monitoring antibiotic resistance genes in wastewater environments: The challenges of filling a gap in the One-Health cycle. J. Hazard. Mater. 2021, 424, 127407. [Google Scholar] [CrossRef]
- Jin, M.; Liu, L.; Wang, D.; Yang, D.; Liu, W.L.; Yin, J.; Yang, Z.W.; Wang, H.R.; Qiu, Z.G.; Shen, Z.Q.; et al. Chlorine disinfection promotes the exchange of antibiotic resistance genes across bacterial genera by natural transformation. ISME J. 2020, 14, 1847–1856. [Google Scholar] [CrossRef] [PubMed]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.; Ryu, C.-M. Plant perceptions of extracellular DNA and RNA. Mol. Plant 2016, 9, 956–958. [Google Scholar] [CrossRef] [PubMed]
- Luziatelli, F.; Ficca, A.G.; Colla, G.; Švecová, E.B.; Ruzzi, M. Foliar application of vegetal-derived bioactive compounds stimulates the growth of beneficial bacteria and enhances microbiome biodiversity in lettuce. Front. Plant Sci. 2019, 10, 1–16. [Google Scholar] [CrossRef]
- Cardinale, M.; Grube, M.; Erlacher, A.; Quehenberger, J.; Berg, G. Bacterial networks and co-occurrence relationships in the lettuce root microbiota. Environ. Microbiol. 2015, 17, 239–252. [Google Scholar]
- Erlacher, A.; Cardinale, M.; Grube, M.; Berg, G. Biotic stress shifted structure and abundance of enterobacteriaceae in the lettuce microbiome. PLoS ONE 2015, 10, e0118068. [Google Scholar] [CrossRef]
- Klerks, M.M.; Franz, E.; Van Gent-Pelzer, M.; Zijlstra, C.; Van Bruggen, A.H.C. Differential interaction of Salmonella enterica serovars with lettuce cultivars and plant-microbe factors influencing the colonization efficiency. ISME J. 2007, 1, 620–631. [Google Scholar]
- Mapelli, F.; Marasco, R.; Rolli, E.; Barbato, M.; Cherif, H.; Guesmi, A.; Ouzari, I.; Daffonchio, D.; Borin, S. Potential for Plant Growth Promotion of Rhizobacteria Associated with Salicornia Growing in Tunisian Hypersaline Soils. BioMed Res. Int. 2013, 2013, 248078. [Google Scholar] [CrossRef]
- Riva, F.; Riva, V.; Eckert, E.M.; Colinas, N.; Di Cesare, A.; Borin, S.; Mapelli, F.; Crotti, E. An environmental Escherichia coli strain is naturally competent to acquire exogenous DNA. Front. Microbiol. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Li, J.; McLellan, S.; Ogawa, S. Accumulation and fate of green fluorescent labeled Escherichia coli in laboratory-scale drinking water biofilters. Water Res. 2006, 40, 3023–3028. [Google Scholar]
- Zhang, D.; Zhu, L.; Li, F. Influences and mechanisms of surfactants on pyrene biodegradation based on interactions of surfactant with a Klebsiella oxytoca strain. Bioresour. Technol. 2013, 142, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Halder, S.K.; Yadav, K.K.; Sarkar, R.; Mukherjee, S.; Saha, P.; Haldar, S.; Karmakar, S.; Sen, T. Alteration of Zeta potential and membrane permeability in bacteria: A study with cationic agents. SpringerPlus 2015, 4, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | 1 h | 24 h |
|---|---|---|
| A. baylyi BD413 | 9.39 × 10−4 ± 4.24 × 10−4 | 1.54 × 10−4 ± 5.11 × 10−5 |
| A. baylyi BD413-endo | - | 1.25 × 10−6 ± 3.29 × 10−7 |
| E. coli DH5α | 5.24 × 10−3 ± 3.87 × 10−3 | 4.34 × 10−5 ± 6.81 × 10−5 |
| K. cowanii VR04 | 2.39 × 10−2 ± 1.19 × 10−3 | 3.01 × 10−3 ± 2.14 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riva, V.; Patania, G.; Riva, F.; Vergani, L.; Crotti, E.; Mapelli, F. Acinetobacter baylyi Strain BD413 Can Acquire an Antibiotic Resistance Gene by Natural Transformation on Lettuce Phylloplane and Enter the Endosphere. Antibiotics 2022, 11, 1231. https://doi.org/10.3390/antibiotics11091231
Riva V, Patania G, Riva F, Vergani L, Crotti E, Mapelli F. Acinetobacter baylyi Strain BD413 Can Acquire an Antibiotic Resistance Gene by Natural Transformation on Lettuce Phylloplane and Enter the Endosphere. Antibiotics. 2022; 11(9):1231. https://doi.org/10.3390/antibiotics11091231
Chicago/Turabian StyleRiva, Valentina, Giovanni Patania, Francesco Riva, Lorenzo Vergani, Elena Crotti, and Francesca Mapelli. 2022. "Acinetobacter baylyi Strain BD413 Can Acquire an Antibiotic Resistance Gene by Natural Transformation on Lettuce Phylloplane and Enter the Endosphere" Antibiotics 11, no. 9: 1231. https://doi.org/10.3390/antibiotics11091231