
Evolutionary Dynamics between Phages and Bacteria as a Possible Approach for Designing Effective Phage Therapies against Antibiotic-Resistant Bacteria



1
Department of Biomedical Science, Kangwon National University, Chuncheon 24341, Gangwon, Korea
2
Institute of Bioscience and Biotechnology, Kangwon National University, Chuncheon 24341, Gangwon, Korea
*
Author to whom correspondence should be addressed.
Antibiotics 2022, 11(7), 915; https://doi.org/10.3390/antibiotics11070915
Submission received: 7 June 2022
/
Revised: 1 July 2022
/
Accepted: 5 July 2022
/
Published: 7 July 2022
(This article belongs to the Special Issue Feature Reviews on Bacteriophages)
Abstract
:With the increasing global threat of antibiotic resistance, there is an urgent need to develop new effective therapies to tackle antibiotic-resistant bacterial infections. Bacteriophage therapy is considered as a possible alternative over antibiotics to treat antibiotic-resistant bacteria. However, bacteria can evolve resistance towards bacteriophages through antiphage defense mechanisms, which is a major limitation of phage therapy. The antiphage mechanisms target the phage life cycle, including adsorption, the injection of DNA, synthesis, the assembly of phage particles, and the release of progeny virions. The non-specific bacterial defense mechanisms include adsorption inhibition, superinfection exclusion, restriction-modification, and abortive infection systems. The antiphage defense mechanism includes a clustered regularly interspaced short palindromic repeats (CRISPR)–CRISPR-associated (Cas) system. At the same time, phages can execute a counterstrategy against antiphage defense mechanisms. However, the antibiotic susceptibility and antibiotic resistance in bacteriophage-resistant bacteria still remain unclear in terms of evolutionary trade-offs and trade-ups between phages and bacteria. Since phage resistance has been a major barrier in phage therapy, the trade-offs can be a possible approach to design effective bacteriophage-mediated intervention strategies. Specifically, the trade-offs between phage resistance and antibiotic resistance can be used as therapeutic models for promoting antibiotic susceptibility and reducing virulence traits, known as bacteriophage steering or evolutionary medicine. Therefore, this review highlights the synergistic application of bacteriophages and antibiotics in association with the pleiotropic trade-offs of bacteriophage resistance.
1. Introduction
Since the discovery of penicillin by Alexander Fleming in 1928, antibiotics have saved millions of lives [1]. However, at the same time, bacteria have evolved antibiotic resistance, exposing the limitation of this magic medicine to treat bacterial infections [2]. The emergence and spread of antibiotic-resistant bacteria have been accelerated due to antibiotic misuse and overuse in medicine and agriculture [3,4]. A large number of antibiotic-resistant bacteria have been identified as high-priority pathogens, including methicillin-resistant Staphylococcus aureus (MRSA), extended-spectrum β-lactamases (ESBL) producing Enterobacteriaceae, vancomycin-resistant enterococci (VRE), and multidrug-resistant Pseudomonas aeruginosa, Streptococcus pneumonia, Mycobacterium tuberculosis, and Acinetobacter baumannii [5,6,7,8,9,10]. Human health care, animal industries, food manufacturing companies, and agricultural sectors are facing the antibiotic resistance threat [11,12]. Along with economic losses, antimicrobial resistance tolls 700,000 lives every year throughout the world due to therapeutic failure [13,14]. Bacteria can rapidly acquire molecular mechanisms to evolve antibiotic resistance through horizontal gene transfer (HGT) from everywhere [15]. The infections caused by multidrug-resistant (MDR) pathogens are difficult to treat because of the limited chemotherapeutic options, which has become a top public health concern worldwide. The frequent antibiotic treatment failures have urged the development of alternative therapeutic agents against MDR pathogens [16].
Bacteriophages (phages) are the most abundant biological entities that specifically infect bacteria [17]. Phages contribute to the diversity of bacterial communities in terms of the coevolutionary fitness dynamics [18,19]. Recently, phages have gained revived attention as alternative antibacterial agents over conventional antibiotics [20,21,22,23]. In Europe, phages have been used for therapeutic and prophylactic purposes, with a less adverse effect on normal microbial flora and no side effects [22,24]. However, the emergence of phage resistance still remains a major drawback in therapeutic applications [25,26,27]. The emergence of phage resistance is a major concern for the use of phage therapy in terms of the coevolutionary arms races between phages and bacteria [28,29]. The competitive interactions between phages and bacteria are evolutionary processes, ongoing defense strategies and counterstrategies, for survival in nature [20]. Bacteria evolve phage resistance under selection pressure, resulting in the coincidental changes in bacterial fitness and virulence [4,30]. In this context, it may be assumed that phage-resistant bacteria induce cross-resistance to different phages and other antibiotics in the same way as antibiotic resistance. However, there is still little information with regard to the evolutionary trade-offs and trade-ups between phage resistance and antibiotic resistance. Here, we review the antiphage defense mechanisms in association with the changes in fitness and virulence, specifically antibiotic resistance, in bacteria.
2. Phage Structure and Life Cycle
Phages are estimated to outnumber bacteria by tenfold [31]. Phages were discovered in 1917 and first used as therapeutic agents in 1919 [32,33]. However, the interest in phage therapy was shifted to antibiotics commonly used to treat bacterial infections from the 1940s onwards. Many phages consist of a head/protein capsid containing the genomic material and tail fibers acting as receptor-binding sites [34]. Most phages have polyhedral capsids, predominantly icosahedral, except for filamentous ones [35]. The structure of tailed phages consists of an icosahedral head and a tail with receptor-binding proteins (RBPs) such as tail spikes and tail fibers at the distal end [36,37]. Phage genomes containing single-stranded or double-stranded RNA or DNA are encased in the protein capsid. RBPs are observed in Myoviridae as long and short fibers attached to the contractile tail, in Podoviridae as spikes or fibers attached to a short non-contractile tail, and in Siphoviridae as baseplates, fibers, spikes, or single straight filaments attached to a long non-contractile tail [38]. The key component of the tail is the contractile sheath. In T4 phages, the contractile sheath contracts to less than half of its original length during the infection to insert the tail tube through the outer membrane for genome delivery [39].
Phages are completely dependent on their hosts for multiplication. Phages have two distinctive life cycles, lytic and lysogenic. In the lytic life cycle, the phages infect the host, multiply inside, and lyse the host cell to release the mature progeny phages. In the lysogenic life cycle, on the contrary, phages integrate their genome into the host’s chromosome, and integrated phage DNA is replicated concurrently with bacterial DNA. In addition, certain temperate phages are maintained as episomal components rather than being incorporated into the host chromosome. The phage DNA integrated into the host cell is termed as the prophage. However, stresses such as UV radiation, antibiotics, pH, temperature, and water activity can activate prophages [32,40]. When prophage activation is triggered, an irreversible transformation from lysogeny to the lytic life cycle takes place and the phage completes the lytic cycle by making copies and lyses the cell to burst them out [41]. The phage attachment to the bacterial host commences with the binding of RBP on the tip of the phage tail to a target receptor on the host cell surface, occurring in three steps—initial contact, reversible adherence and irreversible attachment [42,43]. The initial step is the random collision of the phage with the hosts and the recognition of the receptors on the host surface [42]. After phage receptor recognition, the phage RBP reversibly binds with the receptor [44]. The last step is the irreversible phage binding to the receptor [45]. After the permanent binding with the receptor, the phage ejects the genetic material into the host cytoplasm [46] and the phage genome is expressed to produce virion particles such as head, tail, base plate, and fiber, followed by simultaneous assembly. After the phage assembly, phage-encoding enzymes such as endolysin and holin help release the progenies from the host cells [47].
3. Coevolutionary Dynamics of Phage-Bacteria Interactions
Bacteria evolve phage resistance through bacteria defense systems under phage selection pressure [48,49] (Figure 1). In this context, phages, however, evolve counter adaptations against bacterial antiphage mechanisms [27,50]. Therefore, phages and bacteria can undergo continuous coevolutionary processes involving phage infection and antiphage defense mechanisms [21,27,50,51,52]. The phage resistance mechanisms of bacteria include non-specific adaptations and specific adaptation systems [48,49]. The non-specific bacterial defense mechanisms (innate immune systems) against phages include the inhibition of phage attachment to the host surface receptors, the prevention of phage genome entry into the host cells, the restriction of secondary phage infection (superinfection exclusion), the activation of endonucleases and methyltransferases (restriction-modification system), and the induction of suicide in infected cell (abortive infection system) [48]. The phage-specific bacterial defense mechanisms (adaptive immune systems) are a second line of antiphage defense systems such as clustered regularly interspaced short palindromic repeats (CRISPR)–CRISPR-associated (Cas) proteins [53]. The phage-resistant bacteria result in phenotypic and genotypic changes, including growth rate, membrane permeability, capsular polysaccharide (CPS) production, phage-binding receptor, virulence, and antibiotic susceptibility [54,55]. The antiphage mechanisms in bacteria developed against phage infection stages, including adsorption, penetration, synthesis, assembly, and release [56,57,58,59].
4. Antiphage Defense Mechanisms
4.1. Phage Binding-Based Defense Mechanisms
The first stage of the phage infection process is the attachment to bacterial surface receptors such as lipopolysaccharides (LPSs), pili, outer membrane proteins (OMPs), and efflux pumps [60]. The attachment-blocking mechanism is the predominant defensive strategy for preventing phage infection [59,61].
4.1.1. Prevention of Phage Attachment and Entry
Phage adsorption is blocked by modifying cell surface receptors, producing extracellular polysaccharides, or synthesizing the analogs of receptor-binding proteins, resulting in phage resistance [22,49,62]. To be specific, the mutation in the cell surface receptors, such as pili, flagella, outer membrane proteins, and LPS, is pivotal for the inhibition of phage binding to the host bacteria [63,64,65,66]. The modification of phage-binding receptors occurs in nutrient-rich conditions, while the CRISPR–Cas system is active in nutrient-depleted conditions [66]. The overproduction of glycocalyx, including capsule, slime layer, and exopolysaccharide (EPS), prevents phage adsorption by masking the phage-binding receptors [67,68]. Biofilm cells enclosed in physical barriers, mainly the EPS matrix, confer a high level resistance to phages [69]. An outer membrane lipoprotein (TraT) encoded by the F plasmid overlays phage receptor (OmpA) and prevents the attachment of OmpA-specific phages [70,71]. Mutation in phage-binding receptors can induce bacterial resistance to phage infection. The outer membrane protein OmpA of Escherichia coli K-12 is used as a receptor by T-even-like bacteriophages [72]. Phage resistance is achieved by point mutations, rearrangements, and insertions that modify the OmpA surface protein [73].
Bacteria often use a phase variation mechanism, in which bacteria undergo reversible phenotypic changes through alterations in definite loci of the genome, to evade phage infection [74]. Hemophilus influenzae adopts a phase variation mechanism to alter lipooligosaccharide (LOS) construction by changing the lic2A gene, which affects LOS biosynthesis, to resist HP1c1 phage adsorption [75]. Similarly, Salmonella Typhymurium harbors a gtrABC1 cluster which glycosylates the O12 antigen to evade the SPC35 phage infection. Although BtuB is the receptor of phage SPC35, it is O12 antigen which facilitates the adsorption with BtuB. Upon the selection of phage SPC35, gtrABC1 glycosylates the O12 antigen, which successfully prevents the phage infection [76]. Random variation in hypermutable homonucleotide 7–11G (polyG) tracts causes phase variation in Campylobacter and in Campylobacter jejuni. The polyG tracts were observed in 20–30 genes, which are mostly related to the modification of surface structures [77,78]. The Campylobacter jejuni strain was reported to alter its phase by changing the number of Gs in the hypervariable gene cj1421, which codes for GlafNAc MeOPN transferase, to become resistant to phage F336. The resistant isolate lacked the O-methyl phosphoramidate moiety as it was apparent from the periodate treatment that the carbohydrate moiety was the receptor of phage F336 [78]. The alteration of the number of Gs was also observed in the C. jejuni genes cj1139 and cj0039 [77]. Phase variation also modulates the mutual persistence of phage and bacterial host. When bacteroides intestinalis was selected with crAss-like phage crAss001, the bacterial host developed resistance via a phase variation in CPS [79].
The outer membrane vesicles (OMVs) limit the access of phages to the surface. The OMV successfully protects P. aeruginosa against two phages, KT28 and LUZ7 [80]. Phages also face competition from receptor molecules for binding with bacteria. Microcin J25 (MccJ25), a 21 L-amino acid plasmid-encoded antimicrobial peptide, binds with an outer membrane receptor of T5 phage, FhuA [81]. The antimicrobial peptides, as analogs, competitively bind to phage-binding receptors such as FhuA [62]. Thus, MccJ25 prevents the T5 phage binding with FhuA and infect the host [62].
4.1.2. Superinfection Exclusion Systems
The prophage integrated in a host chromosome blocks the entry of the phage genome, known as superinfection immunity [82]. The entry of the phage genome into the host cell is regulated by superinfection exclusion (Sie) [83]. The pre-existing Sie-related genome encoded in the prophage plays an important role in phage–phage interactions [59]. The Sie system encoded by Imm and Sp prevents phage genome injection into the host cells. Imm blocks the passage of the phage genome through the plasma membrane, and Sp protects the bacterial murein layer which is a target site for tail-associated lysozymes (gp5) [84,85]. Spackle, a periplasmic protein, triggers the Sie system and inactivates gp5 through the formation of a stoichiometric complex [86]. Twitching inhibitory protein (Tip), expressed by the P. aeruginosa prophage D3112, prevents type IV pili expansion by interacting with ATPase PilB, leading to the inhibition of phage infection [87]. Lambdoid phages, including mEp213, mEp237, and mEp410, cannot infect mEp167 lysogenic cells [88,89]. Subcluster B2 Hedgerow and Rosebush phages bind to the receptors in a Wag31 kinase-dependent manner [90]. A Cluster F mycobacteriophage Fuitloop-encoded protein gp52 interacts with Wag31, resulting in the prevention of superinfection [91]. Similarly, Cor protein blocks the phage-binding receptor (FhuA) and mediates superinfection exclusion [89]. The Cor protein also interacts with the outer membrane proteins such as OmpA, OmpC, OmpF, OmpW, LamB, and Slp, which can block the attachment of FhuA-dependent phages [92]. The expression of the cI repressor in lysogenic cells promotes the cleavage of the infecting phage genome, leading to the development of the antiphage defense system [25,93].
4.2. Inhibition of Phage Synthesis and Assembly
After phage adsorption, the phage genome enters the host cell. The injected phage genome is degraded at the levels of replication, transcription, translation, and assembly by restriction-modification (R-M) and CRISPR–Cas systems [94,95].
4.2.1. Restriction-Modification Systems
Restriction-modification (R-M) systems are prokaryotic immune systems used to protect bacteria against foreign DNA such as phages and plasmids. The R-M systems consist of restriction endonucleases (REases) and methyltransferases (MTases), which degrade unmethylated phage genomes and methylate host DNA to protect the self-genome from cleaving by REase, respectively [96,97,98]. The REases are classified into four types according to their subunit structure, recognition site, cofactor requirement, and specific activity [99]. Type I, II, and III endonucleases correspond to the MTases with the same reignition sites. Type I R-M systems are composed of three different subunits required for restriction, methylation, and recognition. Type II and Type III R-M systems are composed of two distinct subunits for restriction and modification. Unlike other R-M systems, Type IV R-M systems are composed of a single unit that can cleave modified sequences such as methylated hydroxymethylcytosine (HMC) and other modified bases [100]. Homing endonucleases are involved in specific DNA modifications [101]. The hemi-methylated molecules are targeted for methylation by MTase, and completely methylated DNA is excluded from both of these functions [102]. REase cleaves the unmethylated DNA into benign segments after the successful attachment of a phage and insertion of DNA into the host cell. Bacteriophage exclusion (BREX) does not degrade unmethylated DNA but inhibits the replication of the phage genome in the host cells [103].
4.2.2. CRISPR–Cas Systems
The CRISPR–Cas systems are responsible for adaptive immunity, which have the ability to effectively destroy the injected phage genome by remembering past infections [59,95]. The CRISPR–Cas locus is composed of short identical repeats interspaced with space sequences which are not identical and flanked by Cas protein genes. The repeats are identical in length and sequence, whereas the spacers are uniform only in length. Repeat lengths vary from 21 bp to 47 bp based on the species. Spacers, or protospacers, are also similar in size, between 20 and 72 bp [104]. Protospacer adjacent motif (PAM) (2–5 bp) is used to identify the target DNA. This short-conserved region is essential to differentiate between self- and non-self-DNA [105]. After the recognition of PAM, protospacer, a segment of downstream DNA, is copied from the foreign DNA and transcribed into CRISPR RNAs (crRNAs). The degradation of foreign DNA or RNA occurs when crRNA with trans-activating crRNA guides the Cas9 endonuclease to the cleaving site of the invading DNA [106,107]. Bacteria evolve CRISPR–Cas-mediated phage resistance through several steps, including the active integration of phage-derived proto-spacers into the CRISPR loci, the transcription of CRISPR loci to express precursor-CRISPR RNA (pre-crRNA), the modification of pre-crRNA into short crRNA, the formation of guide RNA (crRNA and tracrRNA), and the RNA-guided interference of the target phages (sequence-specific immunity) [108,109,110]. The CRISPR–Cas systems are one of the major antiphage defense strategies, showing more than 70% Cas positivity in phage-resistant bacteria [111,112,113].
4.2.3. Abortive Infection Systems
Abortive infection (Abi) systems can block the replication, transcription, and translation in phage-infected cells, which protect adjacent uninfected cells within the population from phage attack [49,114]. Phage infection is very specific; thus, Abi protects only very closely related bacteria by killing only the infected host [115]. The functional modules are required for the Abi system to recognize the phage infection and shut down the metabolism [114]. The cell killing module is initiated by the recognition of phage infection. However, this module is regulated very tightly so that the Abi system does not activate unless the cell is infected and hinders normal cell activity [61].
One of the most studied Abi is the Rex system, which is found in the λ phage of lysogenic E. coli strains [59]. Rex is a two-component system that aborts the lytic growth of phages expressed by λ prophage genes, rexA and rexB [116]. A protein–DNA complex is produced as a result of replication or recombination during the phage infection that activates RexA [117]. One copy of RexB, a membrane-anchored protein with four transmembrane helices, is activated by two copies of RexA, showing that the protein ratio is critical for Abi action [118]. RexB produces an ion channel in the inner membrane, leading to a severe loss of membrane potential and a subsequent drop in cellular ATP levels and resulting in a decrease in the synthesis of macromolecules, consequently inhibiting bacterial growth and aborting phage infection [116]. Although the activation of Rex kills the majority of cells, it protects about 1/100 of the “Rex-activated” population, leading to the theory that the Rex system causes an osmotic shift that induces a stationary phase that deters the propagation of a superinfecting phage such as T4 [119,120].
DNA replication, transcription, translation, the packing and assembly of phage particles, and host lysis are inhibited by Abi systems [121]. Abi systems induce altruistic self-destruction in phage-infected host cells through toxin–antitoxin (TA) systems [122,123]. The replication of phage DNA is inhibited by the phage exclusion system and disruption of replication-related genes [103,124]. The transcription and translation of the phage genome are also inhibited by the disruption of regulatory proteins [124,125]. Abi systems block phage assembly and induces the death of phage-infected host cells through the TA system, leading to the inhibition of phage replication [123,126,127]. Phage packaging interference (Ppi) proteins play an important role in interfering with the phage packaging [128].
4.2.4. Toxin–Antitoxin Systems
TA systems consist of two genes, a toxin gene and toxin-diminishing antitoxin gene [129]. Toxins inhibit major cellular processes, including replication, translation, and cell wall construction, while antitoxins neutralize the cognate toxins [61]. Toxins impact DNase and RNase activities, ATP synthesis, and replication inhibition, as well as cell division inhibition [61]. The TA systems are involved in essential biological functions such as growth arrest, survival, biofilm formation, plasmid integrity, phage resistance, and virulence [96,130]. Eight different types of TA systems (types I-VIII) are classified based on the characteristics of the toxin and antitoxin molecules (protein or RNA) and the toxin neutralization reactions [131]. TA-encoding genes are found in bacterial chromosomes, plasmids, and phage genomes, particularly in prophages [132].
RnlAB is a type II TA system identified in E. coli K12 that provides effective protection against phage T4 with a defective dmd gene [133,134]. The rnlA-encoded endoribonuclease RNase LS was first known as a bacteriophage T4 infection inhibitor, cleaving T4 mRNAs to limit the expression and preventing T4 phage multiplication [133,134]. The toxin-encoding rnlA is responsible for RNase activity and the rnlB encoded downstream of rnlA encodes antitoxin RnlB which neutralizes the toxicity of RnlA [133,134]. The RnlB is readily degraded after infection with a T4 dmd mutant [133]. The host transcription is halted by T4 infection due to rnlAB transcription, leading to the release of active RnlA endoribonuclease [132,133].
5. Phage-Evolving Counterstrategies
Despite the development of bacterial defense mechanisms, phages can evolve counterstrategies to evade bacterial antiphage defense systems through fast adaptive plasticity and replication, including point mutations, genomic rearrangements, and inactivating proteins critical for antiphage defense mechanisms [135].
5.1. Counterstrategies against Receptor Alterations
Phages can evolve new infection mechanisms to evade adsorption-blocking-dependent phage-resistance systems by recognizing new or altered receptors on the host cell surface. Phages can modify their tail fibers (receptor-binding sites) to cope with antiphage system acquired by the conformational changes in the phage-binding receptors [136,137]. The tail fiber protein J of λ phages binds to the outer membrane protein (LamB) of E. coli for infection. The λ phages evolve the J gene that can bind to an alternative receptor (OmpF) instead of modified LamB [136]. Although pathogenic E. coli modifies lipopolysaccharide biosynthesis and/or membrane transporter protein (OmpA) to resist phage infection (ϕHP3), the putative tail spike protein of ϕHP3 is altered to avoid the antiphage defense system [138]. In addition, phages produce enzymes such as depolymerases and hydrolases that degrade exopolysaccharides, leading to an increase in phage adsorption rates [49,139]. The phage-encoding enzymes are classified into lyases and hydrolases based on the cleavage of polysaccharides [59]. A tail spike protein of the Proteus bacteriophage (PmiS_PM-CJR) encodes pectate lyase to degrade the biofilm matrix [140]. Similarly, a tail protein of the phage IME200 degrades the capsule polysaccharide, in Acenetobacter baumanni [141]. Acinetobacter podo phage Petty encodes depolymerase which degrades capsular EPS [142]. Pseudomonas aeruginosa and Bordetella phages create phenotypically diverse phages through hypermutable polyG tracts in order to overcome phage resistance [143,144].
5.2. Counterstrategies against Restriction Modification
Phages also develop counterstrategies against the restriction-modification systems [49,145]. The phage counterstrategies include the alteration of restriction sites within the phage genome, the degradation of cofactors required in the restriction-modification systems, and the methylation of the phage genome [49,59,98]. To evade the R-M systems, phages change the restriction recognition site through point mutation or DNA modification. The T4 phage’s genome contains HMC rather than cytosine so that the REase enzyme may not detect the restriction site of the T4 phage [146]. Similar nucleotide modifications occur in Salmonella phage Vil (thymidine replacement by 5-(2-aminoethoxy)methyluridine), Pseudomonas phage M6 (thymidine replacement by 5-(2-aminoethyl)uridine), Bacillus phage SP8 and SPO1 (5-hydroxymethyl uridine (5hmdU)), and cyaophage S-2L (diaminopurine substituted for adenine) [147,148,149]. Some phages containing MTase can protect the phage genome from bacterial endonuclease cleavage [150]. The phage KP15 contains DNA adenine methyltransferase (Dam) and DNA cytosine methyltransferase (Dcm) [150,151]. Coli phages (T2 and T4) and lactococcal 936-type (Phi93, Phi145, and Phi15) encode the methyltransferase gene [150]. The glycosylated HMC of the phages T4, T2 and T6 is degraded by glucose-modified restriction (GMR) proteins such as GmrS and GmrD in E. coli CT596 [152,153]. However, the GRM enzymes are inactivated by T4 phage internal protein I (IPI*), which is degraded by the GMR fusion protein in uropathogenic E. coli UT189 [154]. The phage P1 has Dar proteins (DarA and DarB) which defend the phage genome from the type I RM system. Structural analogs of specific phage DNA sequences protect the phage genome against R-M systems by sequestering it [155].
5.3. Counterstrategies against CRISPR–Cas
Phages can evade CRISPR–Cas systems through point mutation in the protospacer region and the expression of anti-CRISPR proteins [110,156]. To circumvent CRISPR–Cas-mediated immunity, the point mutations in the CRISPR–Cas-targeted sequence and the formation of nucleus-like structures prevent recognition and the cleavage of target sites [157]. The anti-CRISPR proteins directly inactivate the CRISPR–Cas effector complex. Phages need more than one anti-CRISPR defense mechanism to circumvent a CRISPR–Cas system because of the presence of several different CRISPR spacers [65]. Phages successfully infect P. aeruginosa containing type I-F CRISPR–Cas systems. The anti-CRISPR proteins, including AcrF1, AcrF2, AcrF3, AcrF4, and AcrF5, inhibit CRISPR–Cas-mediated gene editing [158]. The Listeria phage (ϕLS46) encodes anti-CRISPR protein (AcrVIA1) to inhibit the type VI-A CRISPR–Cas system of Listeria seeligeri [159]. The T4 phage can repair its genome cleaved by a CRISP–Cas system by using phage-encoded recombinase UvsX [160]. The temperate phage ΦAP1.1 expresses AcrIIA23, an anti-CRISPR protein, to cease the functionality of Cas9. The inhibition of Cas9 activity allows ΦAP1.1 to integrate into direct repeats of CRISPR and neutralizes the immunity. The loss or alteration of spacers modulates the type II-A CRISPR immune response during integration or excision cycles [161].
5.4. Counterstrategies against Abortive Infection
Interestingly, phages induce mutations in their own genome and produce antitoxin proteins to evade abortive infection systems [127]. The T4 phage interferes with the Rex antiphage mechanism to successfully complete the phage replication cycle and produce phage virions in the presence of ATP-dependent and host inner membrane-associated proteins (RIIA and RIIB) [162,163]. T4 phages form plaques on E. coli K-12 strain with lysogenic λ in the presence of the rII locus encoding rIIA and rIIB genes, whereas T4rII mutants are unable to form plaques [163]. Phages can also evade the Abi caused by the PifA protein through mutations in the phage proteins gp10 and gp1.2 [164]. PifA is a membrane-associated protein encoded by the F plasmid pifA gene which suppresses the transcription of T7 late genes [165]. PifA interacts with the membrane integrity to promote the loss of ATP without lysing and killing the cells [166]. The T7 proteins gp10 and gp1.2 activate the PifA abortive mechanism [167].
6. Coevolutionary Trade-Offs between Phage Resistance and Antibiotic Resistance
The evolutionary interactions between phages and bacteria result in the emergence of phage resistance that positively or negatively compromises the antibiotic resistance [168,169]. Phage therapy can enhance not only the resistance to phages but also the susceptibility to antibiotics [30,170]. The bacteria that evolve phage resistance may make trade-offs with bacterial growth, virulence, antibiotic resistance, nutrient uptake, and biofilm formation [27,171]. The enhanced antibiotic susceptibility, which is the positive effect of phage resistance, is a possible solution to overcome the limitations of phage therapy [15] (Table 1).
6.1. Phage-Binding Receptor-Mediated Trade-Offs
The emergence of phage-resistant bacteria is associated with alterations in phage-binding receptors [184]. The bacterial cell surface components are involved in various cellular processes, including nutrient transport, bacterial motility, biofilm formation, and biosynthesis, which incur fitness costs [27]. Bacteria developing resistance to phages reduce virulence because the conformational changes in phage-binding receptors impose high fitness costs [185]. Klebsiella pneumoniae ST258 evolves resistance to the phages Pharr and KpNIH-2 due to the mutation in galU which regulates cell envelope synthesis, galactose metabolism, and trehalose metabolism. Thus, this phage-resistant K. pneumoniae ST258 results in a significantly low growth rate [172]. The resistance of Acinetobacter baumannii to the phages ΦFG02 and ΦCO01 is due to the deletion of gtr29 and gpi, respectively. Mutations in the genes gtr29 and gpi cause reduced biofilm formation, antibiotic resistance, and fitness because these genes are responsible for the biosynthesis of CPSs [173]. The alterations in virulence factors such as CPS and LPS are caused by mutations in genes (wzc, wbaP, and wcaI) related to the biosynthesis of CPS and LPS, which are responsible for phage resistance and virulence attenuation [25,186,187]. The change in the CPS that mediates protection against β-lactams improves the access to antibiotics, leading to enhanced antibiotic susceptibility [188,189]. The alteration in the LPS that mediates membrane permeability promotes antibiotic susceptibility [4,190]. The fitness loss of the PAO1 mutant P. aeruginosa is attributed to the modification of LPS and T4p receptors, which are responsible for the regulation of cell metabolism [174]. The outer membrane protein (FhuA) is the receptor for phages such as T1, T5, Φ80, N15, and HK022, as well as mEp167, mEp213, and mEp450 coliphages [92,191,192,193]. The alteration in FhuA results in phage-insensitive mutants and an enhanced susceptibility to rifamycin, albomycin, colicin M, and microcin J25 [193,194]. Therefore, the modification or loss of bacterial cell surface molecules promotes phage resistance and induces phenotypic conversion [27]. The phage resistance can trade-off or trade-up against virulence, fitness, and antibiotics resistance [64].
6.2. Efflux Pump-Mediated Trade-Offs
The bacterial cell surface molecules include virulence factors and efflux transporters, which are involved in adherence, secretion, and uptake [195,196]. Bacteria have membrane-associated efflux pumps to secrete antimicrobials, metabolites, and quorum-sensing signal molecules [197,198]. The major efflux system consists of a periplasmic lipoprotein (AcrA), inner membrane transporter (AcrB), and outer membrane protein (TolC) in Gram-negative bacteria [199,200]. The efflux pumps play a major role in antibiotic resistance and bacterial pathogenicity [201]. The tolC-deficient E. coli was more sensitive to antibiotics and bile salts than the wild-type strain [202]. The phage-resistant bacteria are deficient in drug efflux pumps, resulting from the evolutionary trade-offs [30,203]. The phage-binding receptors function as multidrug efflux pumps such as TolC and OprM [189]. Modifications in TolC as a receptor for Escherichia phage (TLS) and Salmonella phages (ST27, ST29, and ST35) can enhance susceptibility to antibiotic substrates [170,204]. Alterations in porins, which are phage-binding receptors, are associated with the reduction in antibiotic resistance in phage-resistant bacteria [30,205]. The genomic deletions in antibiotic biotic-resistant bacteria are induced by PIAS, leading to compromised efflux pump systems [20,206]. The loss of outer membrane- and efflux-associated antibiotic resistance is attributed to the modification of phage-binding receptors such as MexAB, MexCD, MexEF, and MexXY [20]. The Mex efflux pump system consists of an inner membrane transporter (MexB, MexY), periplasmic membrane proteins (MexA, MexX), and an outer membrane protein (OprM) [207,208]. The outer membrane permeability-associated low uptake and efflux pump-associated active transport contribute to antibiotic resistance in bacteria [209,210]. Quorum sensing downregulates the phage-binding receptors (OmpK) [187,211]. The efflux pumps impaired by phage resistance can be applied for phage steering to sensitize phage-resistant bacteria to antibiotics [16,30,212].
6.3. Fitness Trade-Offs of Antibiotic Resistance
Receptor-mediated phage resistance imposes high fitness costs and is compromised with increased phage resistance, resulting in trade-offs between enhanced phage resistance and reduced fitness traits [54,138]. The compensatory mutations under phage selection pressure contribute to attenuated fitness, reduced virulence, and increased antibiotic susceptibility [26,64,213,214,215]. This phenomenon is a double-edged sword in that one trait can be developed for survival but other traits can be lost by way of compensation [212]. The altered fitness traits in phage-resistant bacteria include decreases in growth rate, motility, nutrient transport, virulence, and antibiotic resistance [4,21,27,28]. Phage-binding receptors are involved in bacterial pathogenesis, including attachment, colonization, invasion, and immune evasion [216]. The phage resistance trade-off with virulence is responsible for capsule production and biofilm formation [28,217,218,219]. The lack of modification of phage-binding receptors can cause virulence attenuation in phage-resistant bacteria [64,220]. However, unlike receptor-mediated phage resistance, CRISPR-mediated phage resistance does not induce trade-offs between phage resistance and virulence [219]. The phage-induced deletion of resistance gene clusters including bla, aph, sul-fol, and mph restores the susceptibility of phage-resistant bacteria to β-lactam resistance genes, aminoglycoside, sulphonamide/trimethoprim, and macrolide, respectively [25]. However, phage resistance can pleiotropically not only make trade-offs but also trade-ups with fitness, virulence, and antibiotic resistance [4,30,51,189,215,221,222]. The major negative effects of phage resistance in bacteria include the enhanced production of β-lactamases, resulting in antagonistic pleiotropy [189,215,222]. The pleiotropic trade-ups depend on the structural barriers of OMPs and LPSs, leading to enhanced antibiotic resistance and virulence [23,189]. The substrate-specific uptake channels can enhance the cross-resistance to gyrase inhibitor-associated antibiotics due to the mutations in OmpF and Tsx porins [23,223,224]. The trade-ups between phage resistance and antibiotic resistance are regarded as a main limitation of phage therapy [189,215]. In contrast to the trade-offs, the modification in LPS can trade-up with increased antibiotic resistance in combination with decreased outer membrane permeability [223]. Therefore, the therapeutic application of phages needs to consider both antagonistic and synergistic pleiotropies to effectively control MDR bacterial infections [189].
7. Concluding Remarks
The modification (or loss) of phage-binding receptors on the bacterial cell surface results in resistance towards some phages and susceptibility towards others that have alternative receptors. The trade-offs can sensitize phage-resistant bacteria to antibiotics. Thus, combination chemotherapy can be a potential alternative to treat MDR bacteria and also provide a benefit of reusing conventional antibiotics. The phage cocktails can be used to overcome receptor-mediated phage resistance. The evolved phage-induced antibiotic sensitivity (E-PIAS) can be used to overcome the drawback of phage therapy. The combination therapy of E-PIAS and antibiotics can be an alternative method to effectively inhibit MDR pathogens. In view of evolutionary trade-offs, bacteria developing phage resistance by receptor mutations are re-sensitized to antibiotics due to the gene deletions in the mucoid state. The reduced virulence and enhanced antibiotic susceptibility in phage-resistant bacteria ensure the possibility of using phage–antibiotic combination therapy. Phages need specific receptors such as efflux-, uptake-, permeability-, and maintenance-associated bacterial cell surface molecules on the bacterial cell surface. The modification of phage-binding receptors can lead to the trade-offs with decreased antibiotic resistance. Therefore, the trade-offs in phage resistance play an important role in developing effective phage therapy using positive evolutionary outcomes such as decreased virulence, fitness defects, and enhanced antibiotic susceptibility. Specifically, the trade-off with antibiotic resistance can overcome the negative effect of phage therapy. Phage therapy can be tailored factoring in the consequences of trade-offs, which provides synergy with antibiotics to treat MDR infections. However, the application of phages in the treatment of MDR bacteria has to carefully considered in order to avoid negative evolutionary trade-ups. Future studies need to understand the discrepancy between coevolution dynamics in vitro and in vivo on the interaction of phages and antibiotics.
Author Contributions
M.H. was involved in collecting and updating research and also wrote the manuscript. J.A. drafted and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2016R1D1A3B01008304).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gaynes, R. The discovery of penicillin—New insights after more than 75 years of clinical use. Emerg. Infect. Dis. 2017, 23, 849–853. [Google Scholar] [CrossRef]
- Alos, J.I. Antibiotic resistance: A global crisis. Enferm. Infec. Micribiol. Clin. 2015, 33, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Ojala, V.; Laitalainen, J.; Jalasvuori, M. Fight evolution with evolution: Plasmid-dependent phages with a wide host range prevent the spread of antibiotic resistance. Evol. Appl. 2013, 6, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, A.R.; Fortier, A.; Roush, C.; Lessing, A.J.; Bender, R.G.; Barahman, R.; Grant, R.; Chan, B.K.; Turner, P.E. Pleiotropy complicates a trade-off between phage resistance and antibiotic resistance. Proc. Natl. Acad. Sci. USA 2020, 117, 11207–11216. [Google Scholar] [CrossRef] [PubMed]
- Golkar, Z.; Bagasra, O.; Pace, D.G. Bacteriophage therapy: A potential solution for the antibiotic resistance crisis. J. Infect. Dev. Ctries. 2014, 8, 129–236. [Google Scholar] [CrossRef]
- Grant, G.R.; Lederman, J.A.; Brandstetter, R.D. T.G. Heaton, tuberculosis, and artificial pneumothorax: Once again, back to the future? Chest 1997, 112, 7–8. [Google Scholar] [CrossRef]
- Levin, A.S.; Barone, A.A.; Penco, J.; Santos, M.V.; Marinho, I.S.; Arruda, E.A.; Manrique, E.I.; Costa, S.F. Intravenous colistin as therapy for nosocomial infections caused by multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii. Clin. Infect. Dis. 1999, 28, 1008–1011. [Google Scholar] [CrossRef] [Green Version]
- Nachega, J.B.; Chaisson, R.E. Tuberculosis drug resistance: A global threat. Clin. Infect. Dis. 2003, 36, S24–S30. [Google Scholar] [CrossRef] [Green Version]
- Rossolini, G.M.; Mantengoli, E. Antimicrobial resistance in Europe and its potential impact on empirical therapy. Clin. Microbiol. Infect. 2008, 14, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Whitney, C.G.; Farley, M.M.; Hadler, J.; Harrison, L.H.; Lexau, C.; Reingold, A.; Lefkowitz, L.; Cieslak, P.R.; Cetron, M.; Zell, E.R.; et al. Increasing prevalence of multidrug-resistant Streptococcus pneumoniae in the United States. N. Engl. J. Med. 2000, 343, 1917–1924. [Google Scholar] [CrossRef] [Green Version]
- Morehead, M.S.; Scarbrough, C. Emergence of global antibiotic resistance. Prim. Care 2018, 45, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Thanner, S.; Drissner, D.; Walsh, F. Antimicrobial resistance in agriculture. MBio 2016, 7, e02227-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokharel, S.; Raut, S.; Adhikari, B. Tackling antimicrobial resistance in low-income and middle-income countries. BMJ Glob. Health 2019, 4, e002104. [Google Scholar] [CrossRef]
- Uddin, M.J.; Dawan, J.; Jeon, G.; Yu, T.; He, X.; Ahn, J. The role of bacterial membrane vesicles in the dissemination of antibiotic resistance and as promising carriers for therapeutic agent delivery. Microorganisms 2020, 8, 670. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Loh, B.; Gordillo Altamirano, F.; Yu, Y.; Hua, X.; Leptihn, S. Colistin-phage combinations decrease antibiotic resistance in Acinetobacter baumannii via changes in envelope architecture. Emerg. Microbes Infect. 2021, 10, 2205–2219. [Google Scholar] [CrossRef]
- Gurney, J.; Pradier, L.; Griffin, J.S.; Gougat-Barbera, C.; Chan, B.K.; Turner, P.E.; Kaltz, O.; Hochberg, M.E. Phage steering of antibiotic-resistance evolution in the bacterial pathogen, Pseudomonas aeruginosa. Evol. Med. Public Health 2020, 2020, 148–157. [Google Scholar] [CrossRef]
- Hendrix, R.W.; Smith, M.C.M.; Burns, R.N.; Ford, M.E.; Hatfull, G.F. Evolutionary relationships among diverse bacteriophages and prophages: All the world’s a phage. Proc. Natl. Acad. Sci. USA 1999, 96, 2192–2197. [Google Scholar] [CrossRef] [Green Version]
- Koskella, B.; Brockhurst, M.A. Bacteria-phage coevolution as a driver of ecological and evolutionary processes in microbial communities. FEMS Microbiol. Rev. 2014, 38, 916–931. [Google Scholar] [CrossRef] [Green Version]
- Hall, A.R.; Scanlan, P.D.; Morgan, A.D.; Buckling, A. Host-parasite coevolutionary arms races give way to fluctuating selection. Ecol. Lett. 2011, 14, 635–642. [Google Scholar] [CrossRef]
- Koderi Valappil, S.; Shetty, P.; Deim, Z.; Terhes, G.; Urbán, E.; Váczi, S.; Patai, R.; Polgár, T.; Pertics, B.Z.; Schneider, G.; et al. Survival comes at a cost: A coevolution of phage and its host leads to phage resistance and antibiotic sensitivity of Pseudomonas aeruginosa multidrug resistant strains. Front. Microbiol. 2021, 12, 783722. [Google Scholar] [CrossRef]
- Oechslin, F. Resistance development to bacteriophages occurring during bacteriophage therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.; Mills, S.; Ross, R.P. Phages and antibiotic resistance: Are the most abundant entities on earth ready for a comeback? Future Microbiol. 2018, 13, 711–726. [Google Scholar] [CrossRef] [PubMed]
- Kortright, K.E.; Doss-Gollin, S.; Chan, B.K.; Turner, P.E. Evolution of bacterial cross-resistance to lytic phages and albicidin antibiotic. Front. Microbiol. 2021, 12, 658374. [Google Scholar] [CrossRef]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majkowska-Skrobek, G.; Markwitz, P.; Sosnowska, E.; Lood, C.; Lavigne, R.; Drulis-Kawa, Z. The evolutionary trade-offs in phage-resistant Klebsiella pneumoniae entail cross-phage sensitization and loss of multidrug resistance. Environ. Microbiol. 2021, 23, 7723–7740. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.C.T.; Friman, V.-P.; Smith, M.C.M.; Brockhurst, M.A. Cross-resistance is modular in bacteria-phage interactions. PLoS Biol. 2018, 16, e2006057. [Google Scholar] [CrossRef]
- Mangalea, M.R.; Duerkop, B.A. Fitness trade-offs resulting from bacteriophage resistance potentiate synergistic antibacterial strategies. Infect. Immun. 2020, 88, e00926-19. [Google Scholar] [CrossRef] [Green Version]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage therapy: A renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Fauconnier, A. Regulating phage therapy. EMBO Rep. 2017, 18, 198–200. [Google Scholar] [CrossRef]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef] [Green Version]
- Brussow, H.; Hendrix, R.W. Phage genomics: Small is beautiful. Cell 2002, 108, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Haq, I.; Chaudhry, W.; Akhtar, M.; Andleeb, S.; Qadri, I. Bacteriophages and their implications on future biotechnology: A review. Virol. J. 2012, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermoso, J.A.; García, J.L.; García, P. Taking aim on bacterial pathogens: From phage therapy to enzybiotics. Curr. Opin. Microbiol. 2007, 10, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Wurtz, M. Bacteriophage structure. Electron Microsc. Rev. 1992, 5, 283–309. [Google Scholar] [CrossRef]
- Ackermann, H.W. Tailed bacteriophages: The order caudovirales. Adv. Virus Res. 1998, 51, 135–201. [Google Scholar] [PubMed]
- Walter, M.; Fiedler, C.; Grassl, R.; Biebl, M.; Rachel, R.; Hermo-Parrado, X.L.; Llamas-Saiz, A.L.; Seckler, R.; Miller, S.; van Raaij, M.J. Structure of the receptor-binding protein of bacteriophage det7: A podoviral tail spike in a myovirus. J. Virol. 2008, 82, 2265–2273. [Google Scholar] [CrossRef] [Green Version]
- Chao, K.L.; Shang, X.; Greenfield, J.; Linden, S.B.; Alreja, A.B.; Nelson, D.C.; Herzberg, O. Structure of Escherichia coli O157:H7 bacteriophage CBA120 tailspike protein 4 baseplate anchor and tailspike assembly domains (TSP4-N). Sci. Rep. 2022, 12, 2061. [Google Scholar] [CrossRef]
- Iwasaki, T.; Yamashita, E.; Nakagawa, A.; Enomoto, A.; Tomihara, M.; Takeda, S. Three-dimensional structures of bacteriophage neck subunits are shared in Podoviridae, Siphoviridae and Myoviridae. Genes Cells 2018, 23, 528–536. [Google Scholar] [CrossRef] [Green Version]
- Aksyuk, A.A.; Leiman, P.G.; Kurochkina, L.P.; Shneider, M.M.; Kostyuchenko, V.A.; Mesyanzhinov, V.V.; Rossmann, M.G. The tail sheath structure of bacteriophage T4: A molecular machine for infecting bacteria. EMBO J. 2009, 28, 821–829. [Google Scholar] [CrossRef]
- Czaplewski, L.; Bax, R.; Clokie, M.; Dawson, M.; Fairhead, H.; Fischetti, V.A.; Foster, S.; Gilmore, B.F.; Hancock, R.E.; Harper, D.; et al. Alternatives to antibiotics-a pipeline portfolio review. Lancet Infect. Dis. 2016, 16, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Kiesel, B.; Kallies, R.; Jiang, F.L.; Liu, Y.; Maskow, T. A fast and reliable method for monitoring of prophage-activating chemicals. Microb. Biotechnol. 2018, 11, 1112–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moldovan, R.G.; Chapman-McQuiston, E.; Wu, X.L. On kinetics of phage adsorption. Biophys. J. 2007, 93, 303–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiberoni, A.; Guglielmotti, D.; Binetti, A.; Reinheimer, J. Characterization of three Lactobacillus delbrueckii subsp. bulgaricus phages and the physicochemical analysis of phage adsorption. J. Appl. Microbiol. 2004, 96, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, J.; Takemura, I.; Satoh, M.; Kato, S.; Ujihara, T.; Akechi, K.; Matsuzaki, S.; Daibata, M. Improved adsorption of an Enterococcus faecalis bacteriophage PhiEF24C with a spontaneous point mutation. PLoS ONE 2011, 6, e26648. [Google Scholar] [CrossRef]
- Dowah, A.S.A.; Clokie, M.R.J. Review of the nature, diversity and structure of bacteriophage receptor binding proteins that target Gram-positive bacteria. Biophys. Rev. 2018, 10, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Egido, J.E.; Costa, A.R.; Aparicio-Maldonado, C.; Haas, P.J.; Brouns, S.J.J. Mechanisms and clinical importance of bacteriophage resistance. FEMS Microbiol. Rev. 2022, 46, fuab048. [Google Scholar] [CrossRef]
- Kasman, L.M.; Porter, L.D. Bacteriophages. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Bikard, D.; Marraffini, L.A. Innate and adaptive immunity in bacteria: Mechanisms of programmed genetic variation to fight bacteriophages. Curr. Opin. Immunol. 2012, 24, 15–20. [Google Scholar] [CrossRef]
- Samson, J.E.; Magadán, A.H.; Sabri, M.; Moineau, S. Revenge of the phages: Defeating bacterial defences. Nat. Rev. Microbiol. 2013, 11, 675–687. [Google Scholar] [CrossRef]
- Burmeister, A.R.; Sullivan, R.M.; Gallie, J.; Lenski, R.E. Sustained coevolution of phage Lambda and Escherichia coli involves inner- as well as outer-membrane defences and counter-defences. Microbiology 2021, 167, 1063. [Google Scholar] [CrossRef]
- Scanlan, P.D.; Buckling, A. Co-evolution with lytic phage selects for the mucoid phenotype of Pseudomonas fluorescens SBW25. ISME J. 2012, 6, 1148–1158. [Google Scholar] [CrossRef] [Green Version]
- Scanlan, P.D.; Hall, A.R.; Lopez-Pascua, L.D.; Buckling, A. Genetic basis of infectivity evolution in a bacteriophage. Mol. Ecol. 2011, 20, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Isaev, A.B.; Musharova, O.S.; Severinov, K.V. Microbial arsenal of antiviral defenses—Part I. Biochemistry 2021, 86, 319–337. [Google Scholar] [CrossRef] [PubMed]
- Kortright, K.E.; Done, R.E.; Chan, B.K.; Souza, V.; Turner, P.E.; Vives, M. Selection for phage resistance reduces virulence of Shigella flexneri. Appl. Environ. Microbiol. 2022, 88, e01514-21. [Google Scholar] [CrossRef] [PubMed]
- Capparelli, R.; Nocerino, N.; Lanzetta, R.; Silipo, A.; Amoresano, A.; Giangrande, C.; Becker, K.; Blaiotta, G.; Evidente, A.; Cimmino, A.; et al. Bacteriophage-resistant Staphylococcus aureus mutant confers broad immunity against staphylococcal infection in mice. PLoS ONE 2010, 5, e11720. [Google Scholar] [CrossRef] [Green Version]
- Aminov, R.I. A brief history of the antibiotic era: Lessons learned and challenges for the future. Front. Microbiol. 2010, 1, 134. [Google Scholar] [CrossRef] [Green Version]
- Summers, W.C. The strange history of phage therapy. Bacteriophage 2012, 2, 130–133. [Google Scholar] [CrossRef] [Green Version]
- Hampton, H.G.; Watson, B.N.J.; Fineran, P.C. The arms race between bacteria and their phage foes. Nature 2020, 577, 327–336. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Luong, T.; Salabarria, A.C.; Roach, D.R. Phage therapy in the resistance era: Where do we stand and where are we going? Clin. Ther. 2020, 42, 1659–1680. [Google Scholar] [CrossRef]
- Rostøl, J.T.; Marraffini, L. (Ph)ighting phages: How bacteria resist their parasites. Cell Host Microbe 2019, 25, 184–194. [Google Scholar] [CrossRef] [Green Version]
- Destoumieux-Garzón, D.; Duquesne, S.; Peduzzi, J.; Goulard, C.; Desmadril, M.; Letellier, L.; Rebuffat, S.; Boulanger, P. The iron-siderophore transporter FhuA is the receptor for the antimicrobial peptide microcin J25: Role of the microcin Val11-Pro16 beta-hairpin region in the recognition mechanism. Biochem. J. 2005, 389, 869–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qimron, U.; Marintcheva, B.; Tabor, S.; Richardson, C.C. Genomewide screens for Escherichia coli genes affecting growth of T7 bacteriophage. Proc. Natl. Acad. Sci. USA 2006, 103, 19039–19044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Loessner, M.J. Beyond antibacterials—Exploring bacteriophages as antivirulence agents. Curr. Opin. Biotechnol. 2021, 68, 166–173. [Google Scholar] [CrossRef] [PubMed]
- van Houte, S.; Ekroth, A.K.; Broniewski, J.M.; Chabas, H.; Ashby, B.; Bondy-Denomy, J.; Gandon, S.; Boots, M.; Paterson, S.; Buckling, A.; et al. The diversity-generating benefits of a prokaryotic adaptive immune system. Nature 2016, 532, 385–388. [Google Scholar] [CrossRef] [Green Version]
- Westra, E.R.; van Houte, S.; Oyesiku-Blakemore, S.; Makin, B.; Broniewski, J.M.; Best, A.; Bondy-Denomy, J.; Davidson, A.; Boots, M.; Buckling, A. Parasite exposure drives selective evolution of constitutive versus inducible defense. Curr. Biol. 2015, 25, 1043–1049. [Google Scholar] [CrossRef] [Green Version]
- Bull, J.J.; Vegge, C.S.; Schmerer, M.; Chaudhry, W.N.; Levin, B.R. Phenotypic resistance and the dynamics of bacterial escape from phage control. PLoS ONE 2014, 9, e94690. [Google Scholar]
- Nwodo, U.U.; Green, E.; Okoh, A.I. Bacterial exopolysaccharides: Functionality and prospects. Int. J. Mol. Sci. 2012, 13, 14002–14015. [Google Scholar] [CrossRef] [Green Version]
- Ferriol-González, C.; Domingo-Calap, P. Phages for biofilm removal. Antibiotics 2020, 9, 268. [Google Scholar] [CrossRef]
- Achtman, M.; Kennedy, N.; Skurray, R. Cell-cell interactions in conjugating Escherichia coli: Role of traT protein in surface exclusion. Proc. Natl. Acad. Sci. USA 1977, 74, 5104–5108. [Google Scholar] [CrossRef] [Green Version]
- Riede, I. Receptor specificity of the short tail fibres (gp12) of T-even type Escherichia coli phages. Mol. Gen. Genetic. 1987, 206, 110–115. [Google Scholar] [CrossRef]
- Morona, R.; Klose, M.; Henning, U. Escherichia coli K-12 outer membrane protein (OmpA) as a bacteriophage receptor: Analysis of mutant genes expressing altered proteins. J. Bacteriol. 1984, 159, 570–578. [Google Scholar] [CrossRef] [Green Version]
- Koebnik, R. Structural and functional roles of the surface-exposed loops of the beta-barrel membrane protein OmpA from Escherichia coli. J. Bacteriol. 1999, 181, 3688–3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallet, B. Playing Dr Jekyll and Mr Hyde: Combined mechanisms of phase variation in bacteria. Curr. Opin. Microbiol. 2001, 4, 570–581. [Google Scholar] [CrossRef] [Green Version]
- Zaleski, P.; Wojciechowski, M.; Piekarowicz, A. The role of Dam methylation in phase variation of Haemophilus influenzae genes involved in defence against phage infection. Microbiology 2005, 151, 3361–3369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Ryu, S. Spontaneous and transient defence against bacteriophage by phase-variable glucosylation of O-antigen in Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2012, 86, 411–425. [Google Scholar] [CrossRef]
- Bayliss, C.D.; Bidmos, F.A.; Anjum, A.; Manchev, V.T.; Richards, R.L.; Grossier, J.P.; Wooldridge, K.G.; Ketley, J.M.; Barrow, P.A.; Jones, M.A.; et al. Phase variable genes of Campylobacter jejuni exhibit high mutation rates and specific mutational patterns but mutability is not the major determinant of population structure during host colonization. Nucleic Acids Res. 2012, 40, 5876–5889. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, M.C.; van Alphen, L.B.; Harboe, A.; Li, J.; Christensen, B.B.; Szymanski, C.M.; Brondsted, L. Bacteriophage F336 recognizes the capsular phosphoramidate modification of Campylobacter jejuni NCTC11168. J. Bacteriol. 2011, 193, 6742–6749. [Google Scholar] [CrossRef] [Green Version]
- Shkoporov, A.N.; Khokhlova, E.V.; Stephens, N.; Hueston, C.; Seymour, S.; Hryckowian, A.J.; Scholz, D.; Ross, R.P.; Hill, C. Long-term persistence of crAss-like phage crAss001 is associated with phase variation in Bacteroides intestinalis. BMC Biol. 2021, 19, 163. [Google Scholar] [CrossRef]
- Augustyniak, D.; Olszak, T.; Drulis-Kawa, Z. Outer membrane vesicles (OMVs) of Pseudomonas aeruginosa provide passive resistance but not sensitization to LPS-specific phages. Viruses 2022, 14, 121. [Google Scholar] [CrossRef]
- Vincent, P.A.; Morero, R.D. The structure and biological aspects of peptide antibiotic microcin J25. Curr. Med. Chem. 2009, 16, 538–549. [Google Scholar] [CrossRef]
- Folimonova, S.Y. Superinfection exclusion is an active virus-controlled function that requires a specific viral protein. J. Virol. 2012, 86, 5554–5561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabbir, M.A.B.; Hao, H.; Shabbir, M.Z.; Wu, Q.; Sattar, A.; Yuan, Z. Bacteria vs. bacteriophages: Parallel evolution of immune arsenals. Front. Microbiol. 2016, 7, 1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M.J.; Henning, U. Superinfection exclusion by T-even-type coliphages. Trends Microbiol. 1994, 2, 137–139. [Google Scholar] [CrossRef]
- Nakagawa, H.; Arisaka, F.; Ishii, S. Isolation and characterization of the bacteriophage T4 tail-associated lysozyme. J. Virol. 1985, 54, 460–466. [Google Scholar] [CrossRef] [Green Version]
- Shi, K.; Oakland, J.T.; Kurniawan, F.; Moeller, N.H.; Banerjee, S.; Aihara, H. Structural basis of superinfection exclusion by bacteriophage T4 Spackle. Commun. Biol. 2020, 3, 691. [Google Scholar] [CrossRef]
- Chung, P.Y.; Toh, Y.S. Anti-biofilm agents: Recent breakthrough against multi-drug resistant Staphylococcus aureus. Pathog. Dis. 2014, 70, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Kameyama, L.; Fernandez, L.; Calderon, J.; Ortiz-Rojas, A.; Patterson, T.A. Characterization of wild lambdoid bacteriophages: Detection of a wide distribution of phage immunity groups and identification of a nus-dependent, nonlambdoid phage group. Virology 1999, 263, 100–111. [Google Scholar] [CrossRef] [Green Version]
- Uc-Mass, A.; Loeza, E.J.; de la Garza, M.; Guarneros, G.; Hernandez-Sanchez, J.; Kameyama, L. An orthologue of the cor gene is involved in the exclusion of temperate lambdoid phages. Evidence that Cor inactivates FhuA receptor functions. Virology 2004, 329, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.M.; Nyayapathy, S.; Lee, J.Y.; Suh, J.W.; Husson, R.N. Wag31, a homologue of the cell division protein DivIVA, regulates growth, morphology and polar cell wall synthesis in Mycobacteria. Microbiology 2008, 154, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Ko, C.C.; Hatfull, G.F. Mycobacteriophage Fruitloop gp52 inactivates Wag31 (DivIVA) to prevent heterotypic superinfection. Mol. Microbiol. 2018, 108, 443–460. [Google Scholar] [CrossRef] [Green Version]
- Arguijo-Hernandez, E.S.; Hernandez-Sanchez, J.; Briones-Pena, S.J.; Oviedo, N.; Mendoza-Hernandez, G.; Guarneros, G.; Kameyama, L. Cor interacts with outer membrane proteins to exclude FhuA-dependent phages. Arch. Virol. 2018, 163, 2959–2969. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.; Wolz, C. Phages of Staphylococcus aureus and their impact on host evolution. Infect. Genet. Evol. 2014, 21, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Ofir, G.; Melamed, S.; Sberro, H.; Mukamel, Z.; Silverman, S.; Yaakov, G.; Doron, S.; Sorek, R. DISARM is a widespread bacterial defence system with broad anti-phage activities. Nat. Microbiol. 2018, 3, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Dy, R.L.; Richter, C.; Salmond, G.P.; Fineran, P.C. Remarkable mechanisms in microbes to resist phage infections. Annu. Rev. Virol. 2014, 1, 307–331. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.; Cormican, M. Whole genome sequencing provides possible explanations for the difference in phage susceptibility among two Salmonella Typhimurium phage types (DT8 and DT30) associated with a single foodborne outbreak. BMC Res. Notes 2015, 8, 728. [Google Scholar] [CrossRef] [Green Version]
- Tock, M.R.; Dryden, D.T. The biology of restriction and anti-restriction. Curr. Opin. Microbiol. 2005, 8, 466–472. [Google Scholar] [CrossRef]
- Roberts, R.J.; Belfort, M.; Bestor, T.; Bhagwat, A.S.; Bickle, T.A.; Bitinaite, J.; Blumenthal, R.M.; Degtyarev, S.K.; Dryden, D.T.F.; Dybvig, K.; et al. A nomenclature for restriction enzymes, DNA methyltransferases, homing endonucleases and their genes. Nucleic Acids Res. 2003, 31, 1805–1812. [Google Scholar] [CrossRef] [Green Version]
- Safari, F.; Sharifi, M.; Farajnia, S.; Akbari, B.; Karimi Baba Ahmadi, M.; Negahdaripour, M.; Ghasemi, Y. The interaction of phages and bacteria: The co-evolutionary arms race. Crit. Rev. Biotechnol. 2020, 40, 119–137. [Google Scholar] [CrossRef]
- Oliveira, H.; Thiagarajan, V.; Walmagh, M.; Sillankorva, S.; Lavigne, R.; Neves-Petersen, M.T.; Kluskens, L.D.; Azeredo, J. A thermostable Salmonella phage endolysin, Lys68, with broad bactericidal properties against Gram-negative pathogens in presence of weak acids. PLoS ONE 2014, 9, e108376. [Google Scholar] [CrossRef] [Green Version]
- Vovis, G.F.; Horiuchi, K.; Zinder, N.D. Kinetics of methylation of DNA by a restriction endonuclease from Escherichia coli B. Proc. Natl. Acad. Sci. USA 1974, 71, 3810–3813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldfarb, T.; Sberro, H.; Weinstock, E.; Cohen, O.; Doron, S.; Charpak-Amikam, Y.; Afik, S.; Ofir, G.; Sorek, R. BREX is a novel phage resistance system widespread in microbial genomes. EMBO J. 2015, 34, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Karginov, F.V.; Hannon, G.J. The CRISPR system: Small RNA-guided defense in bacteria and archaea. Mol. Cell 2010, 37, 7–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleditzsch, D.; Pausch, P.; Muller-Esparza, H.; Ozcan, A.; Guo, X.; Bange, G.; Randau, L. PAM identification by CRISPR-Cas effector complexes: Diversified mechanisms and structures. RNA Biol. 2019, 16, 504–517. [Google Scholar] [CrossRef]
- Brouns, S.J.; Jore, M.M.; Lundgren, M.; Westra, E.R.; Slijkhuis, R.J.; Snijders, A.P.; Dickman, M.J.; Makarova, K.S.; Koonin, E.V.; van der Oost, J. Small CRISPR RNAs guide antiviral defense in prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Leon, L.M.; Mendoza, S.D.; Bondy-Denomy, J. How bacteria control the CRISPR-Cas arsenal. Curr. Opin. Microbiol. 2018, 42, 87–95. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Haft, D.H.; et al. An updated evolutionary classification of CRISPR–Cas systems. Nat. Rev. Microbiol. 2015, 13, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Mohanraju, P.; Makarova, K.S.; Zetsche, B.; Zhang, F.; Koonin, E.V.; van der Oost, J. Diverse evolutionary roots and mechanistic variations of the CRISPR-Cas systems. Science 2016, 353, aad5147. [Google Scholar] [CrossRef] [Green Version]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRcompar: A website to compare clustered regularly interspaced short palindromic repeats. Nucleic Acids Res. 2008, 36, W145–W148. [Google Scholar] [CrossRef] [Green Version]
- Leungtongkam, U.; Thummeepak, R.; Kitti, T.; Tasanapak, K.; Wongwigkarn, J.; Styles, K.M.; Wellington, E.M.H.; Millard, A.D.; Sagona, A.P.; Sitthisak, S. Genomic analysis reveals high virulence and antibiotic resistance amongst phage susceptible Acinetobacter baumannii. Sci. Rep. 2020, 10, 16154. [Google Scholar] [CrossRef] [PubMed]
- Wiedenheft, B.; Duijn, E.v.; Bultema, J.B.; Waghmare, S.P.; Zhou, K.; Barendregt, A.; Westphal, W.; Heck, A.J.R.; Boekema, E.J.; Dickman, M.J.; et al. RNA-guided complex from a bacterial immune system enhances target recognition through seed sequence interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 10092–10097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopatina, A.; Tal, N.; Sorek, R. Abortive infection: Bacterial suicide as an antiviral immune strategy. Annu. Rev. Virol. 2020, 7, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Fukuyo, M.; Sasaki, A.; Kobayashi, I. Success of a suicidal defense strategy against infection in a structured habitat. Sci. Rep. 2012, 2, 238. [Google Scholar] [CrossRef] [Green Version]
- Parma, D.H.; Snyder, M.; Sobolevski, S.; Nawroz, M.; Brody, E.; Gold, L. The Rex system of bacteriophage lambda: Tolerance and altruistic cell death. Genes Dev. 1992, 6, 497–510. [Google Scholar] [CrossRef] [Green Version]
- Snyder, L. Phage-exclusion enzymes: A bonanza of biochemical and cell biology reagents? Mol. Microbiol. 1995, 15, 415–420. [Google Scholar] [CrossRef]
- Snyder, L.; McWilliams, K. The rex genes of bacteriophage lambda can inhibit cell function without phage superinfection. Gene 1989, 81, 17–24. [Google Scholar] [CrossRef]
- Slavcev, R.A.; Hayes, S. Rex-centric mutualism. J. Bacteriol. 2002, 184, 857–858. [Google Scholar] [CrossRef] [Green Version]
- Slavcev, R.A.; Hayes, S. Stationary phase-like properties of the bacteriophage lambda Rex exclusion phenotype. Mol. Gene. Genom. 2003, 269, 40–48. [Google Scholar] [CrossRef]
- Domingues, S.; McGovern, S.; Plochocka, D.; Santos, M.A.; Ehrlich, S.D.; Polard, P.; Chopin, M.C. The lactococcal abortive infection protein AbiP is membrane-anchored and binds nucleic acids. Virology 2008, 373, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Blower, T.R.; Short, F.L.; Fineran, P.C.; Salmond, G.P.C. Viral molecular mimicry circumvents abortive infection and suppresses bacterial suicide to make hosts permissive for replication. Bacteriophage 2012, 2, 234–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fineran, P.C.; Blower, T.R.; Foulds, I.J.; Humphreys, D.P.; Lilley, K.S.; Salmond, G.P. The phage abortive infection system, ToxIN, functions as a protein-RNA toxin-antitoxin pair. Proc. Natl. Acad. Sci. USA 2009, 106, 894–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maynard, N.D.; Birch, E.W.; Sanghvi, J.C.; Chen, L.; Gutschow, M.V.; Covert, M.W. A forward-genetic screen and dynamic analysis of lambda phage host-dependencies reveals an extensive interaction network and a new anti-viral strategy. PLoS Genet. 2010, 6, e1001017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, J.; Sun, L.; Yang, X.; Pan, X.; Huang, Z.; Zhang, X.; Gong, M.; Fan, Z.; Li, L.; Cui, X.; et al. Regulatory protein SrpA controls phage infection and core cellular processes in Pseudomonas aeruginosa. Nat. Commun. 2018, 9, 1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidnenko, E.; Chopin, M.C.; Ehrlich, S.D.; Anba, J. Lactococcus lactis AbiD1 abortive infection efficiency is drastically increased by a phage protein. FEMS Microbiol. Lett. 2002, 214, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Stern, A.; Sorek, R. The phage-host arms race: Shaping the evolution of microbes. BioEssays 2011, 33, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Ram, G.; Chen, J.; Kumar, K.; Ross, H.F.; Ubeda, C.; Damle, P.K.; Lane, K.D.; Penadés, J.R.; Christie, G.E.; Novick, R.P. Staphylococcal pathogenicity island interference with helper phage reproduction is a paradigm of molecular parasitism. Proc. Natl. Acad. Sci. USA 2012, 109, 16300–16305. [Google Scholar] [CrossRef] [Green Version]
- Unterholzner, S.J.; Poppenberger, B.; Rozhon, W. Toxin-antitoxin systems: Biology, identification, and application. Mob. Genet. Elements 2013, 3, e26219. [Google Scholar] [CrossRef] [Green Version]
- Page, R.; Peti, W. Toxin-antitoxin systems in bacterial growth arrest and persistence. Nat. Chem. Biol. 2016, 12, 208–214. [Google Scholar] [CrossRef]
- Song, S.; Wood, T.K. A primary physiological role of toxin/antitoxin systems is phage inhibition. Front. Microbiol. 2020, 11, 1895. [Google Scholar] [CrossRef]
- LeRoux, M.; Laub, M.T. Toxin-antitoxin systems as phage defense elements. Ann. Rev. Microbiol. 2022, 76. [Google Scholar] [CrossRef] [PubMed]
- Koga, M.; Otsuka, Y.; Lemire, S.; Yonesaki, T. Escherichia coli rnlA and rnlB compose a novel toxin-antitoxin system. Genetics 2011, 187, 123–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsuka, Y.; Yonesaki, T. A novel endoribonuclease, RNase LS, in Escherichia coli. Genetics 2005, 169, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Pawluk, A.; Davidson, A.R.; Maxwell, K.L. Anti-CRISPR: Discovery, mechanism and function. Nat. Rev. Microbiol. 2018, 16, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.R.; Dobias, D.T.; Weitz, J.S.; Barrick, J.E.; Quick, R.T.; Lenski, R.E. Repeatability and contingency in the evolution of a key innovation in phage lambda. Science 2012, 335, 428–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weitz, J.S.; Hartman, H.; Levin, S.A. Coevolutionary arms races between bacteria and bacteriophage. Proc. Natl. Acad. Sci. USA 2005, 102, 9535–9540. [Google Scholar] [CrossRef] [Green Version]
- Salazar, K.C.; Ma, L.; Green, S.I.; Zulk, J.J.; Trautner, B.W.; Ramig, R.F.; Clark, J.R.; Terwilliger, A.L.; Maresso, A.W.; Goldman, G.H. Antiviral resistance and phage counter adaptation to antibiotic-resistant extraintestinal pathogenic Escherichia coli. MBio 2021, 12, e00211-21. [Google Scholar] [CrossRef]
- Sutherland, I.W. Polysaccharide lyases. FEMS Microbiol. Rev. 1995, 16, 323–347. [Google Scholar] [CrossRef]
- Rice, C.J.; Kelly, S.A.; O’Brien, S.C.; Melaugh, E.M.; Ganacias, J.C.B.; Chai, Z.H.; Gilmore, B.F.; Skvortsov, T. Novel phage-derived depolymerase with activity against Proteus mirabilis biofilms. Microorganisms 2021, 9, 2172. [Google Scholar] [CrossRef]
- Liu, Y.; Mi, Z.; Mi, L.; Huang, Y.; Li, P.; Liu, H.; Yuan, X.; Niu, W.; Jiang, N.; Bai, C.; et al. Identification and characterization of capsule depolymerase Dpo48 from Acinetobacter baumannii phage IME200. PeerJ 2019, 7, e6173. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Morales, A.C.; Lessor, L.L.; Wood, T.L.; Migl, D.; Mijalis, E.M.; Cahill, J.; Russell, W.K.; Young, R.F.; Gill, J.J. Genomic and biochemical characterization of Acinetobacter poodophage Petty reveals a novel lysis mechanism and tail-associated depolymerase activity. J. Virol. 2018, 92, e01064. [Google Scholar] [CrossRef] [Green Version]
- Ceyssens, P.J.; Miroshnikov, K.; Mattheus, W.; Krylov, V.; Robben, J.; Noben, J.P.; Vanderschraeghe, S.; Sykilinda, N.; Kropinski, A.M.; Volckaert, G.; et al. Comparative analysis of the widespread and conserved PB1-like viruses infecting Pseudomonas aeruginosa. Environ. Microbiol. 2009, 11, 2874–2883. [Google Scholar] [CrossRef]
- Sorensen, M.C.H.; Vitt, A.; Neve, H.; Soverini, M.; Ahern, S.J.; Klumpp, J.; Brondsted, L. Campylobacter phages use hypermutable polyG tracts to create phenotypic diversity and evade bacterial resistance. Cell Rep. 2021, 35, 109214. [Google Scholar] [CrossRef]
- Ravin, V.; Räisänen, L.; Alatossava, T. A conserved C-terminal region in Gp71 of the small isometric-head phage LL-H and ORF474 of the prolate-head phage JCL1032 is implicated in specificity of adsorption of phage to its host, Lactobacillus delbrueckii. J. Bacteriol. 2002, 184, 2455–2459. [Google Scholar] [CrossRef] [Green Version]
- O’Farrell, P.H.; Kutter, E.; Nakanishi, M. A restriction map of the bacteriophage T4 genome. Mol. Gen. Genetic. 1980, 179, 421–435. [Google Scholar] [CrossRef] [Green Version]
- Kirnos, M.D.; Khudyakov, I.Y.; Alexandrushkina, N.I.; Vanyushin, B.F. 2-aminoadenine is an adenine substituting for a base in S-2L cyanophage DNA. Nature 1977, 270, 369–370. [Google Scholar] [CrossRef]
- Lee, Y.J.; Dai, N.; Walsh, S.E.; Muller, S.; Fraser, M.E.; Kauffman, K.M.; Guan, C.; Correa, I.R., Jr.; Weigele, P.R. Identification and biosynthesis of thymidine hypermodifications in the genomic DNA of widespread bacterial viruses. Proc. Natl. Acad. Sci. USA 2018, 115, E3116–E3125. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.R.; Casjens, S.R.; Cresawn, S.G.; Houtz, J.M.; Smith, A.L.; Ford, M.E.; Peebles, C.L.; Hatfull, G.F.; Hendrix, R.W.; Huang, W.M.; et al. The genome of Bacillus subtilis bacteriophage SPO1. J. Mol. Biol. 2009, 388, 48–70. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.; Mahony, J.; Ainsworth, S.; Nauta, A.; van Sinderen, D. Bacteriophage orphan DNA methyltransferases: Insights from their bacterial origin, function, and occurrence. Appl. Environ. Microbiol. 2013, 79, 7547–7555. [Google Scholar] [CrossRef] [Green Version]
- Kęsik-Szeloch, A.; Drulis-Kawa, Z.; Weber-Dąbrowska, B.; Kassner, J.; Majkowska-Skrobek, G.; Augustyniak, D.; Łusiak-Szelachowska, M.; Żaczek, M.; Górski, A.; Kropinski, A.M. Characterising the biology of novel lytic bacteriophages infecting multidrug resistant Klebsiella pneumoniae. Virol. J. 2013, 10, 100. [Google Scholar] [CrossRef] [Green Version]
- Bair, C.L.; Black, L.W. A type IV modification dependent restriction nuclease that targets glucosylated hydroxymethyl cytosine modified DNAs. J. Mol. Biol. 2007, 366, 768–778. [Google Scholar] [CrossRef] [Green Version]
- Bair, C.L.; Rifat, D.; Black, L.W. Exclusion of glucosyl-hydroxymethylcytosine DNA containing bacteriophages is overcome by the injected protein inhibitor IPI*. J. Mol. Biol. 2007, 366, 779–789. [Google Scholar] [CrossRef] [Green Version]
- Rifat, D.; Wright, N.T.; Varney, K.M.; Weber, D.J.; Black, L.W. Restriction endonuclease inhibitor IPI* of bacteriophage T4: A novel structure for a dedicated target. J. Mol. Biol. 2008, 375, 720–734. [Google Scholar] [CrossRef] [Green Version]
- Iida, S.; Streiff, M.B.; Bickle, T.A.; Arber, W. Two DNA antirestriction systems of bacteriophage P1, darA, and darB: Characterization of darA-phages. Virology 1987, 157, 156–166. [Google Scholar] [CrossRef]
- Stanley, S.Y.; Borges, A.L.; Chen, K.-H.; Swaney, D.L.; Krogan, N.J.; Bondy-Denomy, J.; Davidson, A.R. Anti-CRISPR-associated proteins are crucial repressors of anti-CRISPR transcription. Cell 2019, 178, 1452–1464. [Google Scholar] [CrossRef]
- Jia, N.; Patel, D.J. Structure-based functional mechanisms and biotechnology applications of anti-CRISPR proteins. Nature Rev. Mol. Cell Biol. 2021, 22, 563–579. [Google Scholar] [CrossRef]
- Bondy-Denomy, J.; Pawluk, A.; Maxwell, K.L.; Davidson, A.R. Bacteriophage genes that inactivate the CRISPR/Cas bacterial immune system. Nature 2013, 493, 429–432. [Google Scholar] [CrossRef] [Green Version]
- Meeske, A.J.; Jia, N.; Cassel, A.K.; Kozlova, A.; Liao, J.; Wiedmann, M.; Patel, D.J.; Marraffini, L.A. A phage-encoded anti-CRISPR enables complete evasion of type VI-A CRISPR-Cas immunity. Science 2020, 369, 54–59. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, J.; Tao, P.; Rao, V.B. Bacteriophage T4 escapes CRISPR attack by minihomology recombination and repair. MBio 2021, 12, e0136121. [Google Scholar] [CrossRef]
- Varble, A.; Campisi, E.; Euler, C.W.; Maguin, P.; Kozlova, A.; Fyodorova, J.; Rostol, J.T.; Fischetti, V.A.; Marraffini, L.A. Prophage integration into CRISPR loci enables evasion of antiviral immunity in Streptococcus pyogenes. Nat. Microbiol. 2021, 6, 1516–1525. [Google Scholar] [CrossRef]
- Shinedling, S.; Parma, D.; Gold, L. Wild-type bacteriophage T4 is restricted by the lambda rex genes. J. Virol. 1987, 61, 3790–3794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.; Alattas, H.; Slavcev, R.A. A snapshot of the lambda T4rII exclusion (Rex) phenotype in Escherichia coli. Curr. Genet. 2021, 67, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Molineux, I.J.; Schmitt, C.K.; Condreay, J.P. Mutants of bacteriophage T7 that escape F restriction. J. Mol. Biol. 1989, 207, 563–574. [Google Scholar] [CrossRef]
- Cheng, X.; Wang, W.; Molineux, I.J. F exclusion of bacteriophage T7 occurs at the cell membrane. Virology 2004, 326, 340–352. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, C.K.; Kemp, P.; Molineux, I.J. Genes 1.2 and 10 of bacteriophages T3 and T7 determine the permeability lesions observed in infected cells of Escherichia coli expressing the F plasmid gene pifA. J. Bacteriol. 1991, 173, 6507–6514. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, C.K.; Molineux, I.J. Expression of gene 1.2 and gene 10 of bacteriophage T7 is lethal to F plasmid-containing Escherichia coli. J. Bacteriol. 1991, 173, 1536–1543. [Google Scholar] [CrossRef] [Green Version]
- Card, K.J.; Thomas, M.D.; Graves, J.L.; Barrick, J.E.; Lenski, R.E. Genomic evolution of antibiotic resistance is contingent on genetic background following a long-term experiment with Escherichia coli. Proc. Natl. Acad. Sci. USA 2021, 118, e2016886118. [Google Scholar] [CrossRef]
- Scanlan, P.D.; Hall, A.R.; Blackshields, G.; Friman, V.P.; Davis, M.R., Jr.; Goldberg, J.B.; Buckling, A. Coevolution with bacteriophages drives genome-wide host evolution and constrains the acquisition of abiotic-beneficial mutations. Mol. Biol. Evol. 2015, 32, 1425–1435. [Google Scholar] [CrossRef] [Green Version]
- German, G.J.; Misra, R. The TolC protein of Escherichia coli serves as a cell-surface receptor for the newly characterized TLS bacteriophage1. J. Mol. Biol. 2001, 308, 579–585. [Google Scholar] [CrossRef]
- Castledine, M.; Padfield, D.; Sierocinski, P.; Soria Pascual, J.; Hughes, A.; Mäkinen, L.; Friman, V.P.; Pirnay, J.P.; Merabishvili, M.; de Vos, D.; et al. Parallel evolution of Pseudomonas aeruginosa phage resistance and virulence loss in response to phage treatment in vivo and in vitro. Elife 2022, 11, e73679. [Google Scholar] [CrossRef]
- Hesse, S.; Rajaure, M.; Wall, E.; Johnson, J.; Bliskovsky, V.; Gottesman, S.; Adhya, S. Phage resistance in multidrug-resistant Klebsiella pneumoniae ST258 evolves via diverse mutations that culminate in impaired adsorption. MBio 2020, 11, e02530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordillo Altamirano, F.; Forsyth, J.H.; Patwa, R.; Kostoulias, X.; Trim, M.; Subedi, D.; Archer, S.K.; Morris, F.C.; Oliveira, C.; Kielty, L.; et al. Bacteriophage-resistant Acinetobacter baumannii are resensitized to antimicrobials. Nat. Microbiol. 2021, 6, 157–161. [Google Scholar] [CrossRef]
- Markwitz, P.; Lood, C.; Olszak, T.; van Noort, V.; Lavigne, R.; Drulis-Kawa, Z. Genome-driven elucidation of phage-host interplay and impact of phage resistance evolution on bacterial fitness. ISME J. 2022, 16, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Braun, V.; Killmann, H.; Herrmann, C. Inactivation of FhuA at the cell surface of Escherichia coli K-12 by a phage T5 lipoprotein at the periplasmic face of the outer membrane. J. Bacteriol. 1994, 176, 4710–4717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siden, I.; Boman, H.G. Escherichia coli mutants with an altered sensitivity to cecropin D. J. Bacteriol. 1983, 154, 170–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zago, M.; Orru, L.; Rossetti, L.; Lamontanara, A.; Fornasari, M.E.; Bonvini, B.; Meucci, A.; Carminati, D.; Cattivelli, L.; Giraffa, G. Survey on the phage resistance mechanisms displayed by a dairy Lactobacillus helveticus strain. Food Microbiol. 2017, 66, 110–116. [Google Scholar] [CrossRef]
- Suarez, V.B.; Maciel, N.; Guglielmotti, D.; Zago, M.; Giraffa, G.; Reinheimer, J. Phage-resistance linked to cell heterogeneity in the commercial strain Lactobacillus delbrueckii subsp. lactis Ab1. Int. J. Food Microbiol. 2008, 128, 401–405. [Google Scholar] [CrossRef]
- Chatterjee, A.; Johnson, C.N.; Luong, P.; Hullahalli, K.; McBride, S.W.; Schubert, A.M.; Palmer, K.L.; Carlson, P.E., Jr.; Duerkop, B.A. Bacteriophage resistance alters Antibiotic-mediated intestinal expansion of Enterococci. Infect. Immun. 2019, 87, e00085-19. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.K.; Fitzpatrick, A.D.; Schwartzkopf, C.M.; Faith, D.R.; Jennings, L.K.; Coluccio, A.; Hunt, D.J.; Michaels, L.A.; Hargil, A.; Chen, Q.; et al. A filamentous bacteriophage protein inhibits type IV pili to prevent superinfection of Pseudomonas aeruginosa. MBio 2022, 13, e02441-21. [Google Scholar] [CrossRef]
- Chung, I.Y.; Jang, H.J.; Bae, H.W.; Cho, Y.H. A phage protein that inhibits the bacterial ATPase required for type IV pilus assembly. Proc. Natl. Acad. Sci. USA 2014, 111, 11503–11508. [Google Scholar] [CrossRef] [Green Version]
- Howlett, R.; Anttonen, K.; Read, N.; Smith, M.C.M. Disruption of the GDP-mannose synthesis pathway in Streptomyces coelicolor results in antibiotic hyper-susceptible phenotypes. Microbiology 2018, 164, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Vale, P.F.; Lafforgue, G.; Gatchitch, F.; Gardan, R.; Moineau, S.; Gandon, S. Costs of CRISPR-Cas-mediated resistance in Streptococcus thermophilus. Proc. Biol. Sci. 2015, 282, 20151270. [Google Scholar] [PubMed] [Green Version]
- Bohannan, B.J.M.; Kerr, B.; Jessup, C.M.; Hughes, J.B.; Sandvik, G. Trade-offs and coexistence in microbial microcosms. Antonie van Leeuwenhoek 2002, 81, 107–115. [Google Scholar] [CrossRef]
- Scott, A.E.; Timms, A.R.; Connerton, P.L.; Loc Carrillo, C.; Adzfa Radzum, K.; Connerton, I.F. Genome dynamics of Campylobacter jejuni in response to bacteriophage predation. PLoS Pathog. 2007, 3, e119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, R.; Wang, G.; Le, S.; Wu, M.; Cheng, M.; Guo, Z.; Ji, Y.; Xi, H.; Zhao, C.; Wang, X.; et al. Three capsular polysaccharide synthesis-related glucosyltransferases, GT-1, GT-2 and WcaJ, are associated with virulence and phage sensitivity of Klebsiella pneumoniae. Front. Microbiol. 2019, 10, 1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.; Zhang, Y.; Qin, J.; Le, S.; Gu, J.; Chen, L.-K.; Guo, X.; Zhu, T. A frameshift mutation in wcaJ associated with phage resistance in Klebsiella pneumoniae. Microorganisms 2020, 8, 378. [Google Scholar] [CrossRef] [Green Version]
- Altamirano, F.G.; Forsyth, J.H.; Patwa, R.; Kostoulias, X.; Trim, M.; Subedi, D.; Archer, S.; Morris, F.C.; Oliveira, C.; Kielty, L.; et al. Bacteriophages targeting Acinetobacter baumannii capsule induce antimicrobial resensitization. Nat. Microbiol. 2020, 6, 157–161. [Google Scholar] [CrossRef]
- Burmeister, A.R.; Turner, P.E. Trading-off and trading-up in the world of bacteria-phage evolution. Curr. Biol. 2020, 30, R1120–R1124. [Google Scholar] [CrossRef]
- Bertozzi Silva, J.; Storms, Z.; Sauvageau, D. Host receptors for bacteriophage adsorption. FEMS Microbiol. Lett. 2016, 363, fnw002. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Sanchez, J.; Bautista-Santos, A.; Fernandez, L.; Bermudez-Cruz, R.M.; Uc-Mass, A.; Martinez-Penafiel, E.; Martinez, M.A.; Garcia-Mena, J.; Guarneros, G.; Kameyama, L. Analysis of some phenotypic traits of feces-borne temperate lambdoid bacteriophages from different immunity groups: A high incidence of cor+, FhuA-dependent phages. Arch. Virol. 2008, 153, 1271–1280. [Google Scholar] [CrossRef]
- Kadner, R.J.; Heller, K.; Coulton, J.W.; Braun, V. Genetic control of hydroxamate-mediated iron uptake in Escherichia coli. J. Bacteriol. 1980, 143, 256–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langenscheid, J.; Killmann, H.; Braun, V. A FhuA mutant of Escherichia coli is infected by phage T1-independent of TonB. FEMS Microbiol. Lett. 2004, 234, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, A.D.; Kodding, J.; Walker, G.; Bos, C.; Coulton, J.W.; Diederichs, K.; Braun, V.; Welte, W. Active transport of an antibiotic rifamycin derivative by the outer-membrane protein FhuA. Structure 2001, 9, 707–716. [Google Scholar] [CrossRef] [Green Version]
- Green, E.R.; Mecsas, J.; Kudva, I.T. Bacterial secretion systems: An overview. Microbiol. Spectr. 2016, 4, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, O.E.; Sauer, K. Sticky situations: Key components that control bacterial surface attachment. J. Bacteriol. 2012, 194, 2413–2425. [Google Scholar] [CrossRef] [Green Version]
- Pearson, J.P.; Van Delden, C.; Iglewski, B.H. Active efflux and diffusion are involved in transport of Pseudomonas aeruginosa cell-to-cell signals. J. Bacteriol. 1999, 181, 1203–1210. [Google Scholar] [CrossRef] [Green Version]
- Piddock, L.J. Multidrug-resistance efflux pumps—Not just for resistance. Nat. Rev. Microbiol. 2006, 4, 629–636. [Google Scholar] [CrossRef]
- Schuster, S.; Vavra, M.; Schweigger, T.M.; Rossen, J.W.A.; Matsumura, Y.; Kern, W.V. Contribution of AcrAB-TolC to multidrug resistance in an Escherichia coli sequence type 131 isolate. Int. J. Antimicrob. Agents 2017, 50, 477–481. [Google Scholar] [CrossRef]
- Zgurskaya, H.I.; Nikaido, H. AcrA is a highly asymmetric protein capable of spanning the periplasm. J. Mol. Biol. 1999, 285, 409–420. [Google Scholar] [CrossRef]
- Nishino, K.; Nikaido, E.; Yamaguchi, A. Regulation and physiological function of multidrug efflux pumps in Escherichia coli and Salmonella. Biochim. Biophys. Acta 2009, 1794, 834–843. [Google Scholar] [CrossRef]
- Tamer, Y.T.; Gaszek, I.; Rodrigues, M.; Coskun, F.S.; Farid, M.; Koh, A.Y.; Russ, W.; Toprak, E. The antibiotic efflux protein TolC is a highly evolvable target under colicin E1 or TLS phage selection. Mol. Biol. Evol. 2021, 38, 4493–4504. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, Y.; Wang, Z.; Wei, J.; Hu, T.; Si, J.; Tao, G.; Zhang, L.; Xie, L.; Abdalla, A.E.; et al. A combination therapy of phages and antibiotics: Two is better than one. Int. J. Biol. Sci. 2021, 17, 3573–3582. [Google Scholar] [CrossRef] [PubMed]
- Ricci, V.; Piddock, L.J.V. Exploiting the role of TolC in pathogenicity: Identification of a bacteriophage for eradication of Salmonella serovars from poultry. Appl. Environ. Microbiol. 2010, 76, 1704–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila, J.; Martí, S.; Sánchez-Céspedes, J. Porins, efflux pumps and multidrug resistance in Acinetobacter baumannii. J. Antimicrob. Chemother. 2007, 59, 1210–1215. [Google Scholar] [CrossRef] [Green Version]
- Hernando-Amado, S.; Sanz-García, F.; Martínez, J.L. Rapid and robust evolution of collateral sensitivity in Pseudomonas aeruginosa antibiotic-resistant mutants. Sci. Adv. 2020, 6, eaba5493. [Google Scholar] [CrossRef]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, N.; Tsujimoto, H.; Nishino, T. Substrate specificities of MexAB-OprM, MexCD-OprJ, and MexXY-oprM efflux pumps in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3322–3327. [Google Scholar] [CrossRef] [Green Version]
- Poole, K. Efflux pumps as antimicrobial resistance mechanisms. Ann. Med. 2007, 39, 162–176. [Google Scholar] [CrossRef]
- Cunrath, O.; Meinel, D.M.; Maturana, P.; Fanous, J.; Buyck, J.M.; Saint Auguste, P.; Seth-Smith, H.M.B.; Körner, J.; Dehio, C.; Trebosc, V.; et al. Quantitative contribution of efflux to multi-drug resistance of clinical Escherichia coli and Pseudomonas aeruginosa strains. EBioMedicine 2019, 41, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Riou, M.; Avrain, L.; Carbonnelle, S.; El Garch, F.; Pirnay, J.-P.; De Vos, D.; Plésiat, P.; Tulkens, P.M.; Van Bambeke, F. Increase of efflux-mediated resistance in Pseudomonas aeruginosa during antibiotic treatment in patients suffering from nosocomial pneumonia. Int. J. Antimicrob. Agents 2016, 47, 77–83. [Google Scholar] [CrossRef]
- Hoque, M.M.; Naser, I.B.; Bari, S.M.N.; Zhu, J.; Mekalanos, J.J.; Faruque, S.M. Quorum regulated resistance of Vibrio cholerae against environmental bacteriophages. Sci. Rep. 2016, 6, 37956. [Google Scholar] [CrossRef]
- Chan, B.K.; Turner, P.E.; Kim, S.; Mojibian, H.R.; Elefteriades, J.A.; Narayan, D. Phage treatment of an aortic graft infected with Pseudomonas aeruginosa. Evol. Med. Public Health 2018, 2018, 60–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernando-Amado, S.; Laborda, P.; Valverde, J.R.; Martínez, J.L. Rapid decline of ceftazidime resistance in antibiotic-free and sublethal environments is contingent on genetic background. Mol. Biol. Evol. 2022, 39, msac049. [Google Scholar] [CrossRef] [PubMed]
- Melnyk, A.H.; Wong, A.; Kassen, R. The fitness costs of antibiotic resistance mutations. Evol. Appl. 2015, 8, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Moulton-Brown, C.E.; Friman, V.-P. Rapid evolution of generalized resistance mechanisms can constrain the efficacy of phage–antibiotic treatments. Evol. Appl. 2018, 11, 1630–1641. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.; Day, C.J.; von Itzstein, M.; Paton, J.C.; Jennings, M.P. Glycointeractions in bacterial pathogenesis. Nat. Rev. Microbiol. 2018, 16, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Alseth, E.O.; Pursey, E.; Luján, A.M.; McLeod, I.; Rollie, C.; Westra, E.R. Bacterial biodiversity drives the evolution of CRISPR-based phage resistance. Nature 2019, 574, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Feng, Y.; McNally, A.; Zong, Z. Characterization of phage resistance and phages capable of intestinal decolonization of carbapenem-resistant Klebsiella pneumoniae in mice. Commun. Biol. 2022, 5, 48. [Google Scholar] [CrossRef]
- León, M.; Bastías, R. Virulence reduction in bacteriophage resistant bacteria. Front. Microbiol. 2015, 6, 343. [Google Scholar] [CrossRef] [Green Version]
- Oechslin, F.; Piccardi, P.; Mancini, S.; Gabard, J.; Moreillon, P.; Entenza, J.M.; Resch, G.; Que, Y.-A. Synergistic interaction between phage therapy and antibiotics clears Pseudomonas aeruginosa infection in endocarditis and reduces virulence. J. Infect. Dis. 2017, 215, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.C.; Pfrunder-Cardozo, K.R.; Meinel, D.; Egli, A.; Hall, A.R. Associations among antibiotic and phage resistance phenotypes in natural and clinical Escherichia coli isolates. MBio 2017, 8, e01341. [Google Scholar] [CrossRef] [Green Version]
- Uddin, M.J.; Dawan, J.; Ahn, J. Assessment of the alteration in phage adsorption rates of antibiotic-resistant Salmonella typhimurium. Arch. Microbiol. 2019, 201, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Broeker, N.K.; Barbirz, S. Not a barrier but a key: How bacteriophages exploit host’s O-antigen as an essential receptor to initiate infection. Mol. Microbiol. 2017, 105, 353–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kortright, K.E.; Chan, B.K.; Turner, P.E. High-throughput discovery of phage receptors using transposon insertion sequencing of bacteria. Proc. Natl. Acad. Sci. USA 2020, 117, 18670–18679. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Overview of bacterial antiphage defense mechanisms. 1. Phage infection: successful phage attachment and infection; 2. competition in receptor binding: blocking of the phage-binding receptor to evade phage attachment; 3. alteration in cell surface receptor: mutation or phase variation in the receptor to avoid phage attachment; 4. hiding phage receptor: production of extracellular polysaccharides to hide phage receptor; 5. superinfection exclusion—A: integration of prophage into host genome; B: expression of protein to block DNA entry; C: exclusion of superinfection; 6. restriction-modification system—A: recognition of restriction site; B: cleaving of inserted phage DNA; 7. CRISPR–Cas immunity; 8. toxin–antitoxin immunity—A: antitoxin neutralizes toxin before phage infection; B: phage infection-mediated liberation of toxin to induce reduced metabolism or cell death; 9. abortive infection—A: phage infection mediates expression of abortive infection mechanism; B: release of unassembled phage particles from the host cell.
Figure 1.
Overview of bacterial antiphage defense mechanisms. 1. Phage infection: successful phage attachment and infection; 2. competition in receptor binding: blocking of the phage-binding receptor to evade phage attachment; 3. alteration in cell surface receptor: mutation or phase variation in the receptor to avoid phage attachment; 4. hiding phage receptor: production of extracellular polysaccharides to hide phage receptor; 5. superinfection exclusion—A: integration of prophage into host genome; B: expression of protein to block DNA entry; C: exclusion of superinfection; 6. restriction-modification system—A: recognition of restriction site; B: cleaving of inserted phage DNA; 7. CRISPR–Cas immunity; 8. toxin–antitoxin immunity—A: antitoxin neutralizes toxin before phage infection; B: phage infection-mediated liberation of toxin to induce reduced metabolism or cell death; 9. abortive infection—A: phage infection mediates expression of abortive infection mechanism; B: release of unassembled phage particles from the host cell.


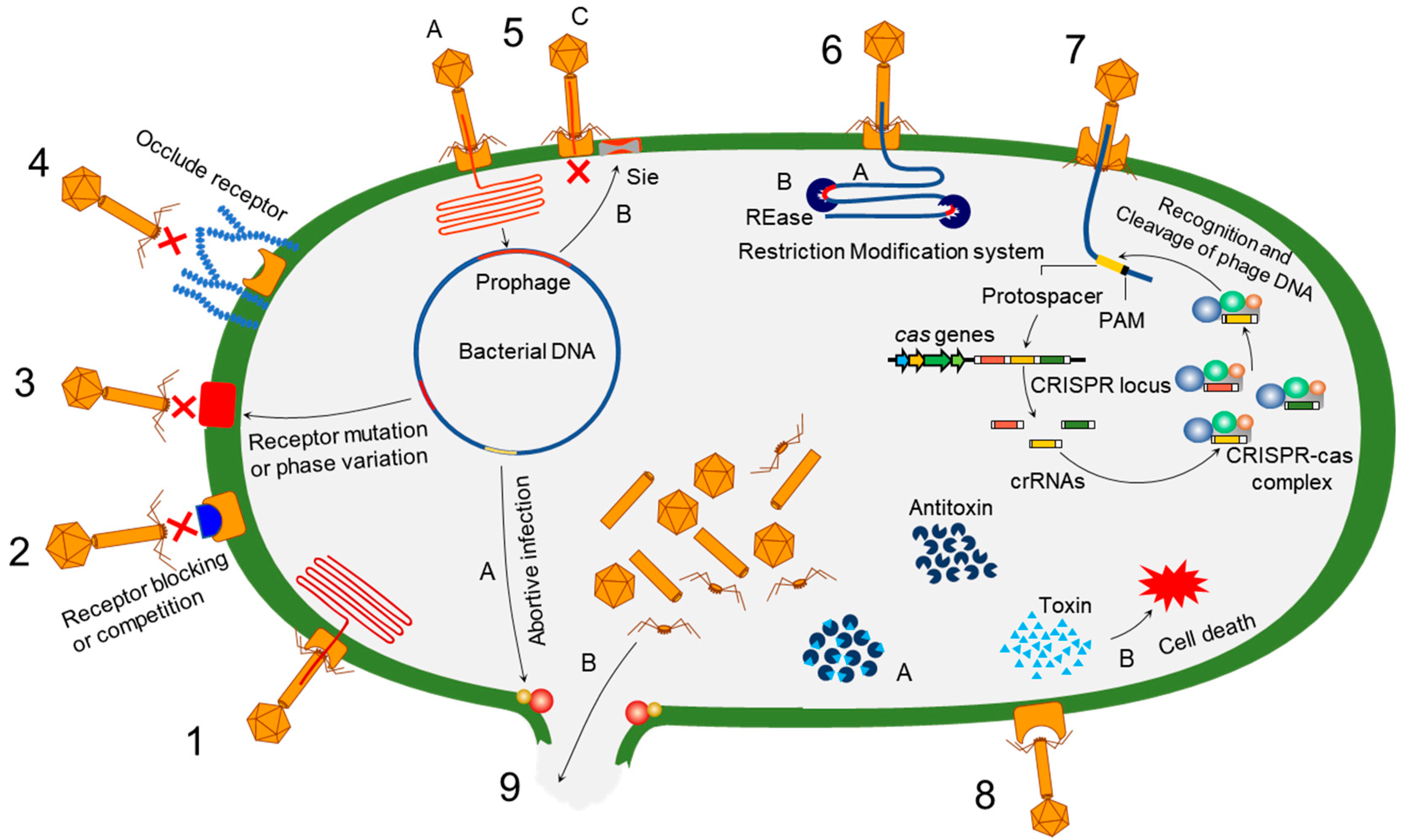{kind=link}
Table 1.
Trade-offs between phages and bacteria.
| Host | Phage | Phage-Binding Receptors | Antiphage Mechanism | Trade-Off | References |
|---|---|---|---|---|---|
| Phage-binding receptor-mediated trade-off | |||||
| Klebsiella pneumonia ST258 | Pharr | Capsular polysaccharide | Mutation in galU | Decrease in growth rate | [172] |
| Klebsiella pneumonia ST258 | ΦKpNIH-2 | LPS, OmpC | Mutation in galU | Decrease in growth rate | [172] |
| Acinetobacter baumannii | ΦFG02 | Capsule | Defective capsule production | Increase in antibiotic susceptibility and, reduction in biofilm formation | [173] |
| Acinetobacter baumannii | ΦCO01 | Capsule | Defective capsule production | Increase in antibiotic susceptibility and reduction in biofilm formation | [173] |
| Pseudomonas aeruginosa PAO1 | φKZ, KTN4, LUZ19 | Type IV pili | Mutation in T4p and global regulatory genes | Decrease in metabolism | [174] |
| Pseudomonas aeruginosa PA14 | Phage DMS3vir | Type IV pili | Loss of pili | Significant competitive cost | [66] |
| Pseudomonas aeruginosa PAO1 | KT28, KTN6, LUZ27 | LPS | Mutations in LPS and global regulatory genes | Reduction in fitness | [174] |
| Escherichia coli | T5 | FhuA | Confrontational changes | Reduction in ferrichrome uptake | [175] |
| Shigella flexneri | A1-1 | OmpA | Mutations in ompA | Increase in vancomycin sensitivity and loss of intracellular movement | [54] |
| Shigella flexneri | A1-1 | LPS | Mutation in genes gmhA and gmhC | Increase in erythromycin sensitivity and loss of intracellular movement | [54] |
| Escherichia coli SB1004 | ΦJE | OmpC | Deficient OmpC | Increase in sensitivity to peptide cecropin D | [176] |
| Lactobacillus helveticus ATCC10386 | ΦLh56 | S-layer | Point mutation | Decrease in S-layer integrity | [177] |
| Lactobacillus delbrueckii subsp. lactis Ab1 T-type | Phage YAB | polysaccharide–peptidoglycan complex | Receptor modification | Decrease in proteolytic and acidifying activities | [43,178] |
| Enterococcus faecalis V583 | Phi4 | Enterococcal polysaccharide antigen (Epa) | Mutation in epaX, epaAC, and epaY | Increase in susceptibility to cell wall-targeting antibiotics | [179] |
| Enterococcus faecalis SF28073 | Phi47 | Mutation in epaR and epaS | Increase in susceptibility to cell wall-targeting antibiotics and deficiency in intestinal colonization | ||
| Acinetobacter baumannii | Phab24 | Capsule | Mutation in gtr9 | Decrease in virulence and increase in colistin susceptibility | [15] |
| Efflux pump-mediated trade-off | |||||
| Pseudomonas aeruginosa PAO1 | OMKO1 | OprM | Suppression of oprM | Increase in antibiotic sensitivity | [30] |
| Pseudomonas aeruginosa MDR | PIAS | MexXY-OprM | Loss or substantial modifications | Increase in antibiotic sensitivity | [20] |
| Escherichia coli | U136B | TolC | Mutation | Increase in sensitivity to tetracycline and colistin | [4] |
| Superinfection exclusion-mediated trade-off | |||||
| Pseudomonas aeruginosa | Pf | Type IV pili | PA0721-PilC complex | Suppression of twitching motility | [180] |
| Pseudomonas aeruginosa | D3112 | Type IV pili | PilB-gp05 complex | Inhibition of twitching motility | [181] |
| Escherichia coli | mEp167 | FhuA | Antiadsorption (Cor-OMP interaction) | Reduction in ferrichorme uptake | [92] |
| Streptomyces coelicolor | φC31 | Glycoproteins | Mutation in ppm1 | Increase in antibiotic susceptibility | [182] |
| CRISPR–Cas-mediated trade-off | |||||
| Streptococcus thermophilus | Phage 2972 | Spacer addition | Reduction in fitness | [183] | |
| Pseudomonas aeruginosa PA14 | Phage DMS3vir | Type IV pili | Spacer addition and loss of pili | Reduction in fitness | [66] |
| R-M system-mediated trade-off | |||||
| Lactobacillus delbrueckii subsp. lactis Ab1 C-type | Phage YAB | Polysaccharide–peptidoglycan complex | Type I R-M system | Decrease in proteolytic and acidifying activity | [43,178] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hasan, M.; Ahn, J. Evolutionary Dynamics between Phages and Bacteria as a Possible Approach for Designing Effective Phage Therapies against Antibiotic-Resistant Bacteria. Antibiotics 2022, 11, 915. https://doi.org/10.3390/antibiotics11070915
AMA Style
Hasan M, Ahn J. Evolutionary Dynamics between Phages and Bacteria as a Possible Approach for Designing Effective Phage Therapies against Antibiotic-Resistant Bacteria. Antibiotics. 2022; 11(7):915. https://doi.org/10.3390/antibiotics11070915
Chicago/Turabian StyleHasan, Mahadi, and Juhee Ahn. 2022. "Evolutionary Dynamics between Phages and Bacteria as a Possible Approach for Designing Effective Phage Therapies against Antibiotic-Resistant Bacteria" Antibiotics 11, no. 7: 915. https://doi.org/10.3390/antibiotics11070915
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.