The Mediterranean Mussel Mytilus galloprovincialis (Mollusca: Bivalvia) in Chile: Distribution and Genetic Structure of a Recently Introduced Invasive Marine Species

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling, DNA Extraction, Amplification and Sequencing
2.2. Data Analyses
2.2.1. Population Genetic Structure
2.2.2. Gene Lineages, Genetic Distance and Demographic Analyses
2.2.3. Population Genetic Diversity
2.2.4. Demographic Analyses—Chile
2.2.5. Phylogenetic and Phylogeographic Analyses
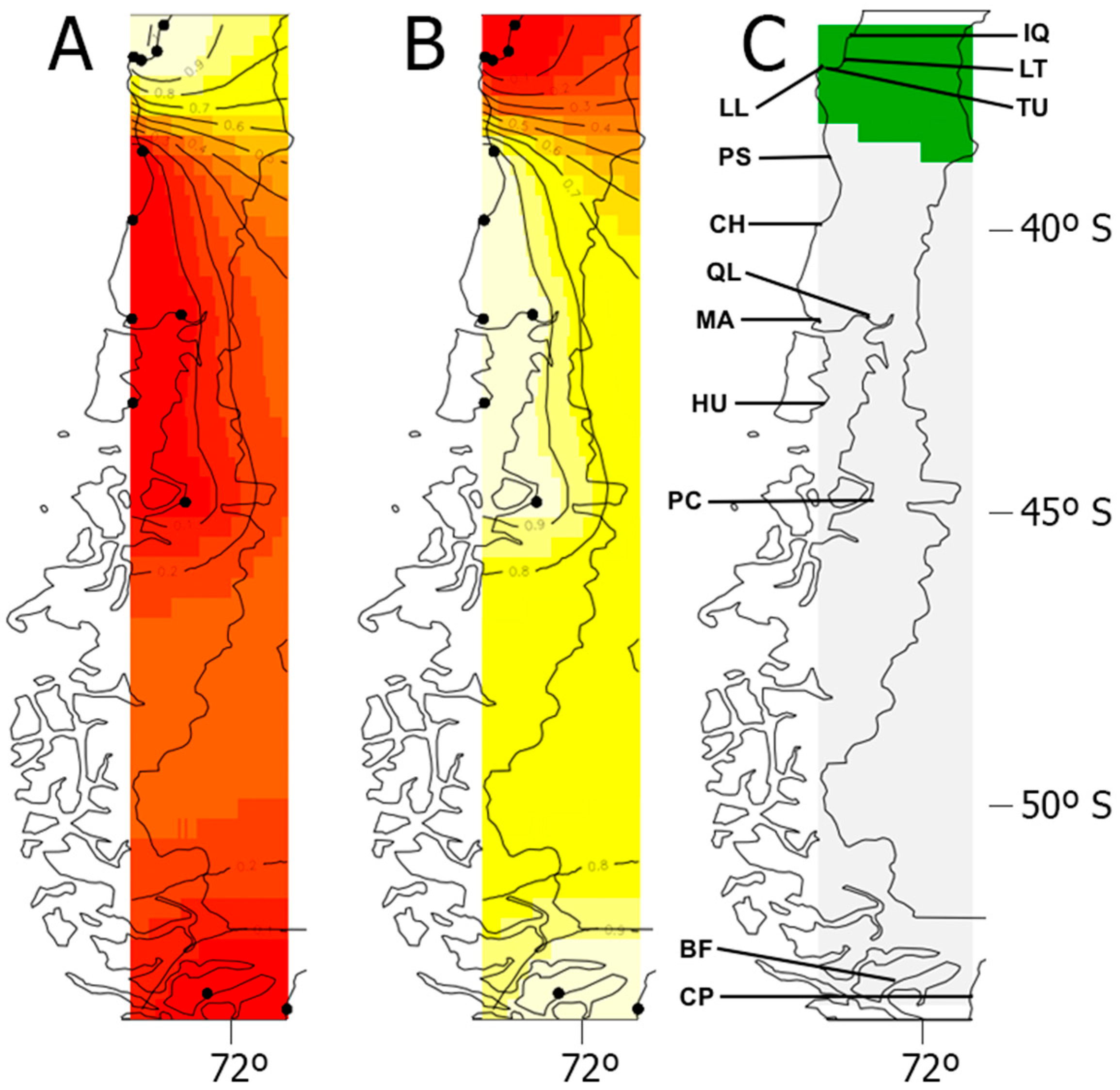
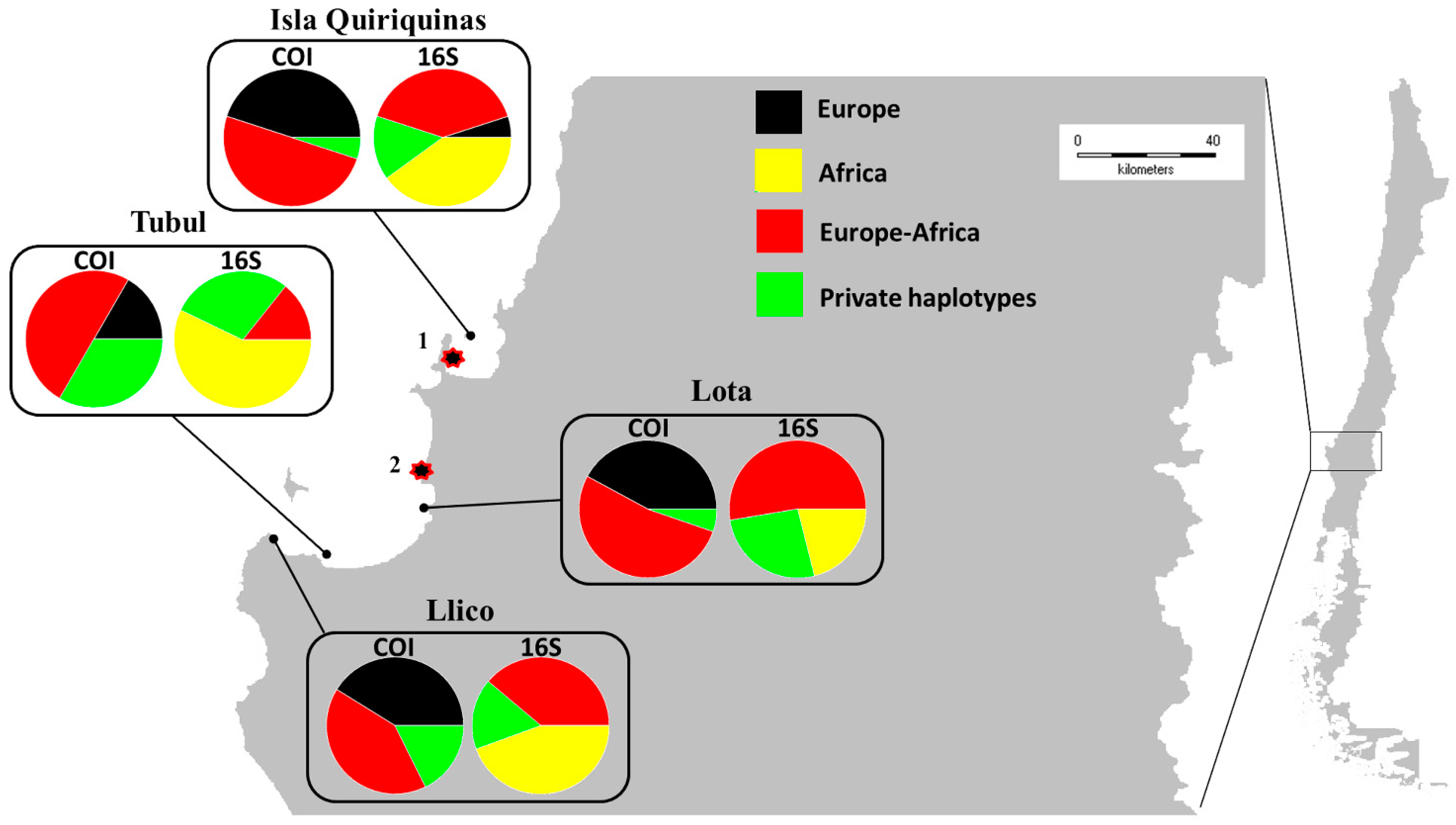
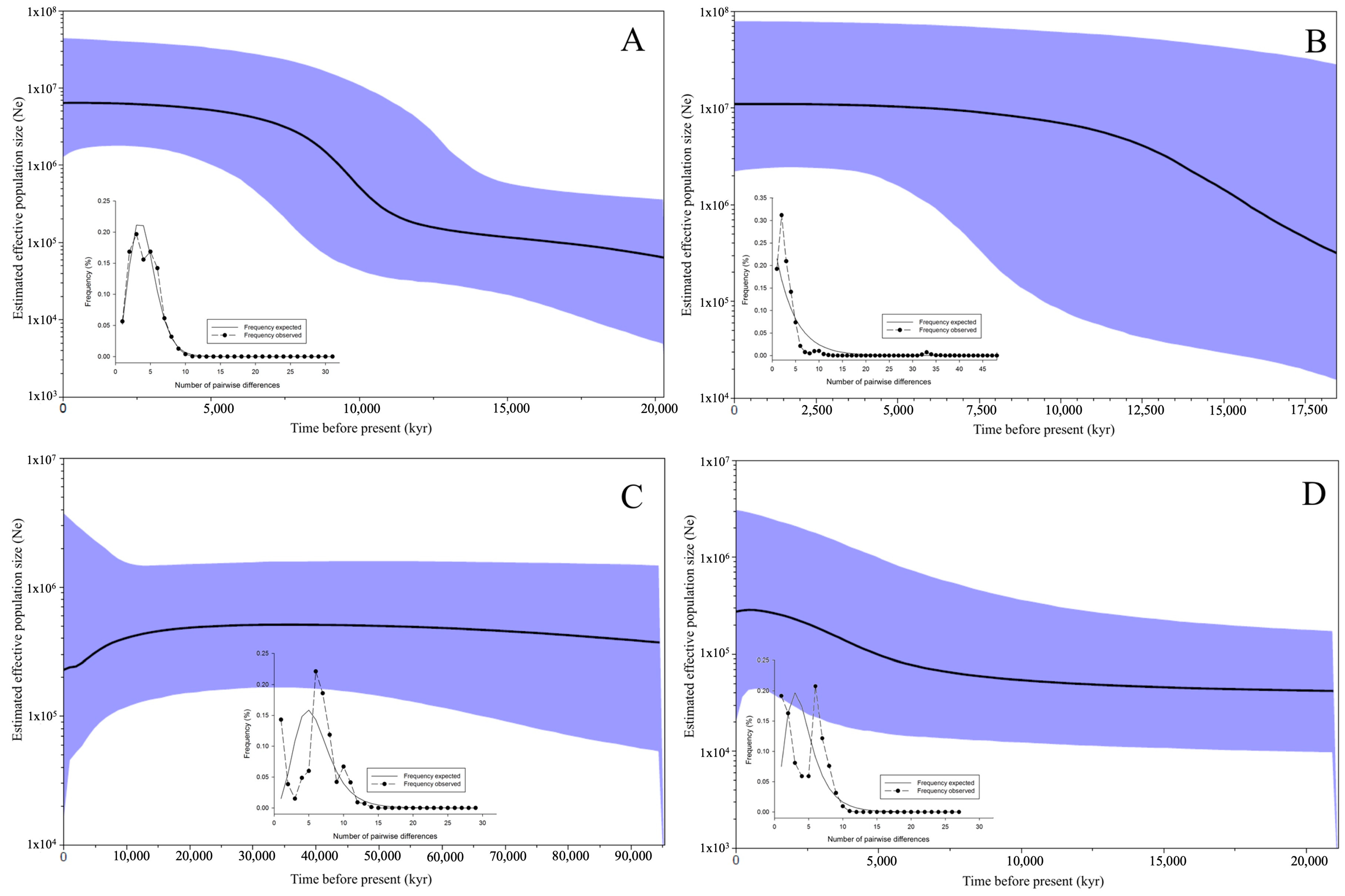
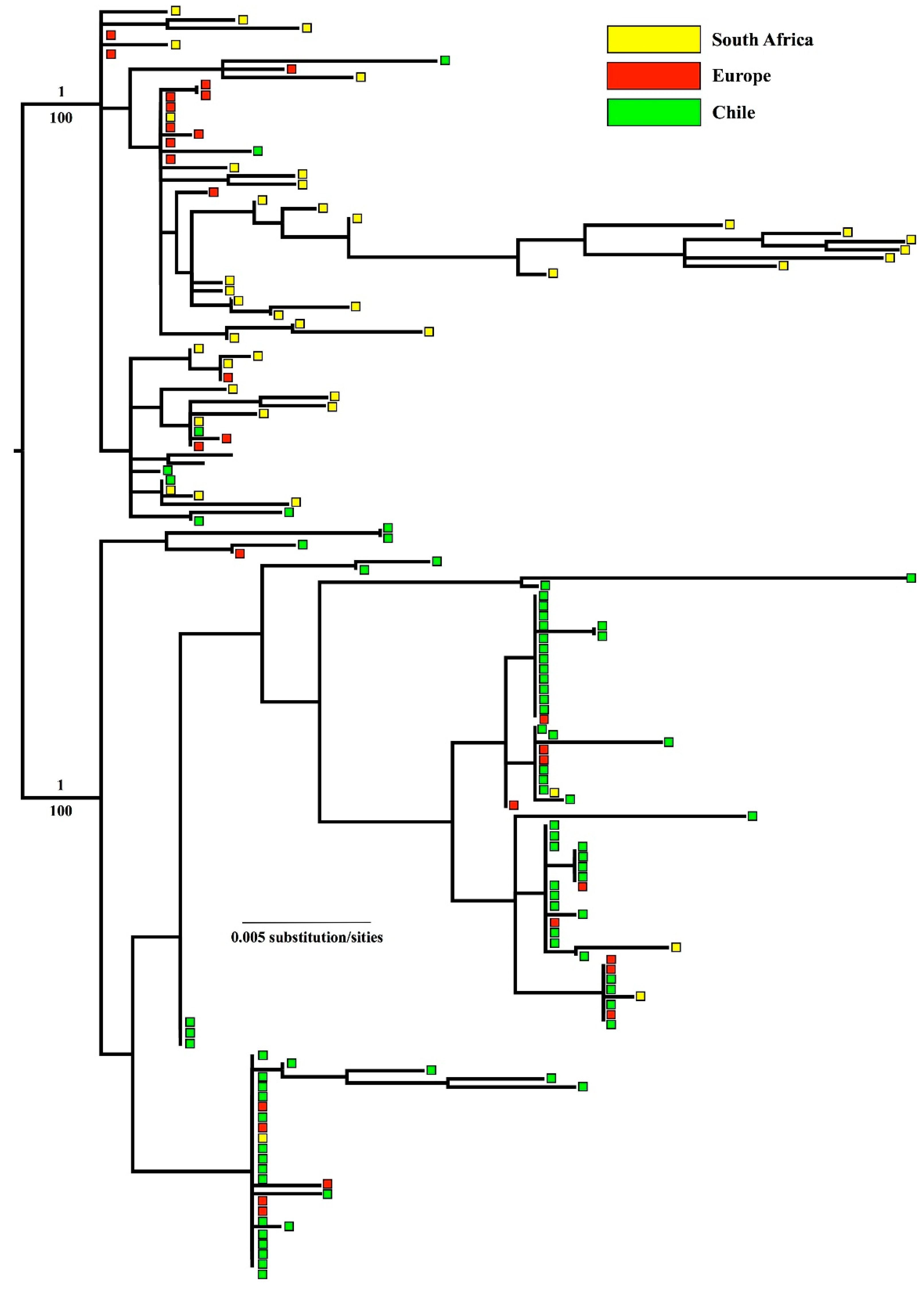
3. Results
3.1. Sequence Characteristics
3.2. Genetic Structure and Gene Lineages
3.3. Genetic Diversity
3.4. Demographic Analyses
3.5. Phylogenetic Relationships
4. Discussion
4.1. Populations of Mytilus spp. in Chile
4.2. Invasion of Chile by Mytilus Galloprovincialis
4.3. Spread of M. galloprovincialis in Chile
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giraud, T. Biological globalisation: Bioinvasions and their impacts on nature, the economy and public health. Environ. Sci. 2008, 5, 214–216. [Google Scholar] [CrossRef]
- Paiva, F.; Barco, A.; Chen, Y.; Mirzajani, A.; Chan, F.T.; Lauringson, V.; Baltazar-Soares, M.; Zhan, A.; Bailey, S.A.; Javidpour, J.; et al. Is salinity an obstacle for biological invasions? Glob. Chang. Biol. 2018, 24, 2708–2720. [Google Scholar] [CrossRef] [PubMed]
- Bax, N.; Williamson, A.; Aguero, M.; Gonzalez, E.; Geeves, W. Marine invasive alien species: A threat to global biodiversity. Mar. Pol. 2003, 27, 313–323. [Google Scholar] [CrossRef]
- Ojaveer, H.; Galil, B.S.; Campbell, M.L.; Carlton, J.T.; Canning-Clode, J.; Cook, E.J. Classification of non-Indigenous species based on their impacts: Considerations for application in marine management. PLoS Biol. 2015, 13, e1002130. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Winter, M. Disentangling the role of environmental and human pressures on biological invasions across Europe. Proc. Natl. Acad. Sci. USA 2010, 107, 12157–12162. [Google Scholar] [CrossRef] [PubMed]
- Cosner, C. Challenges in modeling biological invasions and population distributions in a changing climate. Ecol. Complex. 2014, 20, 258–263. [Google Scholar] [CrossRef]
- Gardner, J.P.A.; Zbawicka, M.; Westfall, K.M.; Wenne, R. Invasive blue mussels threaten regional scale genetic diversity in mainland and remote offshore locations: The need for baseline data and enhanced protection in the Southern Ocean. Glob. Chang. Biol. 2016, 9, 3182–3195. [Google Scholar] [CrossRef]
- Geller, J.B. Decline of a native mussel masked by sibling species invasion. Conserv. Biol. 1999, 13, 661–664. [Google Scholar] [CrossRef]
- Oyarzún, P.A.; Toro, J.E.; Canete, J.I.; Gardner, J.P.A. Bioinvasion threatens the genetic integrity of native diversity and a natural hybrid zone: Smooth-shelled blue mussels (Mytilus spp.) in the Strait of Magellan. Biol. J. Linn. Soc. 2016, 117, 574–585. [Google Scholar] [CrossRef]
- Hockey, P.A.R.; van Erkom Schurink, C. The invasive biology of the mussel Mytilus galloprovincialis on the southern African coast. Trans. R Soc. South Afr. 1992, 48, 123–139. [Google Scholar] [CrossRef]
- Robinson, T.B.; Griffiths, C.L.; McQuaid, C.D.; Rius, M. Marine alien species of South Africa—Status and impacts. Afr. J. Mar. Sci. 2005, 27, 297–306. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The population biology of invasive species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef]
- Anderson, A.S.; Bilodeau, A.L.; Gilg, M.R.; Hilbish, T.J. Routes of introduction of the Mediterranean mussel (Mytilus galloprovincialis) to Puget Sound and Hood Canal. J. Shell Res. 2002, 21, 75–79. [Google Scholar]
- Apte, S.; Holland, B.S.; Godwin, L.S.; Gardner, J.P.A. Jumping ship: A stepping stone event mediating transfer of a non-indigenous species via a potentially unsuitable environment. Biol. Invasions 2000, 2, 75–79. [Google Scholar] [CrossRef]
- Colautti, R.I.; Lau, J.A. Contemporary evolution during invasion: Evidence for differentiation, natural selection, and local adaptation. Mol. Ecol. 2015, 24, 1999–2017. [Google Scholar] [CrossRef]
- Bossdorf, O.; Auge, H.; Lafuma, L.; Rogers, W.E.; Siemann, E.; Prati, D. Phenotypic and genetic differentiation between native and introduced plant populations. Oecologia 2005, 144, 1–11. [Google Scholar] [CrossRef]
- Estoup, A.; Ravigné, V.; Hufbauer, R.; Vitalis, R.; Gautier, M.; Facon, B. Is there a genetic paradox of biological invasion? Annu. Rev. Ecol. Evol. Syst. 2016, 47, 51–72. [Google Scholar] [CrossRef]
- Rollins, L.A.; Moles, A.T.; Lam, S.; Buitenwef, R.; Buswell, J.M.; Brandenburger, C.R.; Flores-Moreno, H.; Nielsen, K.B.; Couchman, K.; Brown, G.S.; et al. High genetic diversity is not essential for successful introduction. Ecol. Evol. 2013, 3, 4501–4517. [Google Scholar] [CrossRef] [PubMed]
- Crawford, K.M.; Whitney, K.D. Population genetic diversity influences colonization success. Mol. Ecol. 2010, 19, 1253–1263. [Google Scholar] [CrossRef]
- Jones, E.I.; Gomulkiewicz, R. Biotic interactions, rapid evolution, and the establishment of introduced species. Am. Nat. 2012, 179, E28–E36. [Google Scholar] [CrossRef] [PubMed]
- Hufbauer, R.A. Biological invasions: Paradox lost and paradise gained. Curr. Biol. 2008, 18, R246–R247. [Google Scholar] [CrossRef]
- Kolbe, J.J.; Glor, R.E.; Rodriguez, L.; Chamizo, A.; Larson, A.; Losos, J.B. Genetic variation increases during biological invasion by a Cuban lizard. Nature 2004, 431, 177–181. [Google Scholar] [CrossRef]
- Roman, J.; Darling, J.A. Paradox lost: Genetic diversity and the success of aquatic invasions. Trends Ecol. Evol. 2007, 22, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Allendorf, F.W.; Luikart, G. Conservation and the Genetics of Populations; Blackwell: Malden, MA, USA, 2007. [Google Scholar]
- Allendorf, F.W.; Lundquist, L.L. Introduction: Population biology, evolution, and control of invasive species. Conserv. Biol. 2003, 17, 24–30. [Google Scholar] [CrossRef]
- Geller, J.B.; Darling, J.A.; Carlton, J.T. Genetic perspectives on marine biological invasions. Ann. Rev. Mar. Sci. 2010, 2, 401–427. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R. Invasion biology—Resolving the genetic paradox in invasive species. Heredity 2005, 94, 385. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Dlugosch, K.M.; Parker, I.M. Founding events in species invasions: Genetic variation, adaptive evolution, and the role of multiple introductions. Mol. Ecol. 2008, 17, 431–449. [Google Scholar] [CrossRef] [PubMed]
- Kinzinger, A.P.; Nakamoto, R.J.; Anderson, E.C.; Harvey, B.C. Small founding number and low genetic diversity in an introduced species exhibiting limited invasion success (speckled dace, Rhinichthys osculus). Ecol. Evol. 2011, 1, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J.P.A.; Oyarzún, P.A.; Toro, J.E.; Wenne, R.; Zbawicka, M. Phylogeography of Southern Hemisphere blue mussels of the genus Mytilus: Evolution, biosecurity, aquaculture and food labelling. Oceanogr. Mar. Biol. Annu. Rev. 2021, 59, 139–228. [Google Scholar]
- Gosling, E.M. Systematics and geographic distribution of Mytilus. In The Mussel Mytilus: Ecology, Physiology, Genetics and Culture; Gosling, E.M., Ed.; Elsevier: Amsterdam, The Netherlands, 1992; pp. 1–20. [Google Scholar]
- Larraín, M.A.; Zbawicka, M.; Araneda, C.; Gardner, J.P.A.; Wenne, R. Native and invasive taxa on the Pacific coast of South America: Impacts on aquaculture, traceability and biodiversity of blue mussels (Mytilus spp.). Evol. Appl. 2018, 11, 298–311. [Google Scholar] [CrossRef]
- McDonald, J.H.; Seed, R.; Koehn, R.K. Allozymes and morphometric characters of three species of Mytilus in the northern and southern hemispheres. Mar. Biol. 1991, 111, 323–333. [Google Scholar] [CrossRef]
- Oyarzún, P.A.; Toro, J.E.; Nuñez, J.J.; Suárez-Villota, E.Y.; Gardner, J.P.A. Blue mussels of the Mytilus edulis species complex from South America: The application of species delimitation models to DNA sequence variation. PLoS ONE 2021, 16, e0256961. [Google Scholar] [CrossRef] [PubMed]
- Zbawicka, M.; Gardner, J.P.A.; Wenne, R. Cryptic diversity in smooth-shelled mussels on Southern Ocean islands: Connectivity, hybridisation and a marine invasion. Front. Zool. 2019, 6, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Zbawicka, M.; Trucco, M.I.; Wenne, R. Single nucleotide polymorphisms in native South American Atlantic coast populations of smooth shelled mussels: Hybridization with invasive European Mytilus galloprovincialis. Genet. Sel. Evol. 2018, 50, 5. [Google Scholar] [CrossRef] [PubMed]
- Zbawicka, M.; Wenne, R.; Dias, P.J.; Gardner, J.P.A. Combined threats to native smooth-shelled mussels (genus Mytilus) in Australian: Bioinvasions and hybridisation. Zool. J. Linn. Soc. 2021, 194, 1194–1211. [Google Scholar] [CrossRef]
- Illesca, A.; Oyarzún, P.A.; Toro, J.E.; Gardner, J.P.A. Morphometric variability of smooth-shelled blue mussels from the Pacific coast of South America. Biol. J. Linn. Soc. 2018, 125, 194–209. [Google Scholar] [CrossRef]
- Hilbish, T.J.; Mullinax, A.; Dolven, S.I.; Meyer, A.; Koehn, R.K.; Rawson, P.D. Origin of the antitropical distribution pattern in marine mussels (Mytilus spp.): Routes and timing of transequatorial migration. Mar. Biol. 2000, 136, 69–77. [Google Scholar] [CrossRef]
- Popovic, I.; Matias, A.M.A.; Bierne, N.; Riginos, C. Twin introductions by independent invader mussel lineages are both associated with recent admixture with a native congener in Australia. Evol. Appl. 2020, 13, 515–532. [Google Scholar] [CrossRef]
- Barsotti, G.; Meluzzi, C. Osservazioni su Mytilus edulis L. e Mytilus Gall. Lamark. Conchiglie 1968, 4, 50–58. [Google Scholar]
- GISD. Global Invasive Species Database. Mytilus galloprovincialis. 2012. Available online: http://www.iucngisd.org/gisd/species.php?sc=102 (accessed on 28 February 2024).
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’S Worst Invasive Alien Species. A Selection from the Global Invasive Species Database (IUCN); Hollands Printing: Auckland, New Zealand, 2000. [Google Scholar]
- Borsa, P.; Rolland, V.; Daguin-Thiebaut, C. Genetics and taxonomy of Chilean smooth-shelled mussels, Mytilus spp. (Bivalvia: Mytilidae). Comptes Rendus Biol. 2012, 335, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Brannock, P.M.; Wethey, D.S.; Hilbish, T.J. Extensive hybridization with minimal introgression in Mytilus galloprovincialis and M-trossulus in Hokkaido, Japan. Mar. Ecol. Prog. Ser. 2009, 383, 161–171. [Google Scholar] [CrossRef]
- Daguin, C.; Borsa, P. Genetic relationships of Mytilus galloprovincialis Lmk. populations worldwide: Evidence from nuclear DNA markers. In Bivalve Systematics and Evolution; Crame, A., Harper, E., Taylor, J.S., Eds.; Geological Society of London Special Publications: London, UK, 2000; pp. 389–397. [Google Scholar]
- Elliott, J.; Holmes, K.; Chambers, R.; Leon, K.; Wimberger, P. Differences in morphology and habitat use among the native mussel Mytilus trossulus, the non-native M-galloprovincialis, and their hybrids in Puget Sound, Washington. Mar. Biol. 2008, 156, 39–53. [Google Scholar] [CrossRef]
- Grant, W.S.; Cherry, M.I. M. galloprovincialis Lmk. in southern Africa. J. Exp. Mar. Biol. Ecol. 1985, 90, 179–191. [Google Scholar] [CrossRef]
- Westfall, K.M.; Gardner, J.P.A. Genetic diversity of Southern hemisphere blue mussels (Bivalvia: Mytilidae) and the identification of non-indigenous taxa. Biol. J. Linn. Soc. 2010, 101, 898–909. [Google Scholar] [CrossRef]
- Westfall, K.M.; Gardner, J.P.A. Interlineage Mytilus galloprovincialis Lmk. 1819 hybridization yields inconsistent genetic outcomes in the Southern hemisphere. Biol. Invasions 2013, 15, 1493–1506. [Google Scholar] [CrossRef]
- Cárdenas, L.; Leclerc, J.-C.; Bruning, P.; Garrido, I.; Détrée, C.; Figueroa, A.; Astorga, M.; Navarro, J.M.; Johnson, L.E.; Carlston, J.T.; et al. First mussel settlement observed in Antarctica reveals the potential for future invasions. Sci. Rep. 2020, 10, 5552. [Google Scholar] [CrossRef]
- Lee, J.E.; Chown, S.L. Mytilus on the move: Transport of an invasive bivalve to the Antarctic. Mar. Ecol. Prog. Ser. 2007, 339, 307–310. [Google Scholar] [CrossRef]
- Murray, C.C.; Pakhomov, E.A.; Therriault, T.W. Recreational boating: A large unregulated vector transporting marine invasive species. Divers. Distrib. 2011, 17, 1161–1172. [Google Scholar] [CrossRef]
- Simon, A.; Arbiol, C.; Nielsen, E.E.; Couteau, J.; Sussarellu, R.; Burgeot, T.; Bernard, I.; Coolen, J.W.P.; Lamy, J.-B.; Robert, S.; et al. Replicated anthropogenic hybridisations reveal parallel patterns of admixture in marine mussels. Evol. Appl. 2019, 13, 575–599. [Google Scholar] [CrossRef]
- Gardner, J.P.A.; Westfall, K.M. Geographic distribution and molecular identification of a metapopulation of blue mussels (Genus Mytilus) in Northeastern New Zealand. J. Moll Stud. 2012, 78, 66–73. [Google Scholar] [CrossRef]
- Robinson, T.; Griffiths, C.L.; Branch, G.M.; Govender, A. The invasion and subsequent die-off of Mytilus galloprovincialis in Langebaan Lagoon, South Africa: Effects on natural communities. Mar. Biol. 2007, 152, 225–232. [Google Scholar] [CrossRef]
- Robinson, T.B.; Branch, G.M.; Griffiths, C.L.; Govender, A.; Hockey, P.A.R. Changes in South African rocky intertidal invertebrate community structure associated with the invasion of the mussel Mytilus galloprovincialis. Mar. Ecol. Prog. Ser. 2007, 340, 163–171. [Google Scholar] [CrossRef]
- Ma, K.C.K.; Gusha, M.N.C.; Zardi, G.I.; Nicastro, K.R.; Monsinjon, J.R.; McQuaid, C.D. Biogeographic drivers of distribution and abundance in an alien ecosystem engineer: Transboundary range expansion, barriers to spread, and spatial structure. J. Biogeogr. 2021, 48, 1941–1959. [Google Scholar] [CrossRef]
- Assis, J.; Zupan, M.; Nicastro, K.R.; Zardi, G.I.; McQuaid, C.D.; Serrão, E.A. Oceanographic conditions limit the spread of a marine invader along southern African shores. PLoS ONE 2015, 10, e0128124. [Google Scholar] [CrossRef] [PubMed]
- Zardi, G.I.; McQuaid, C.D.; Jaciento, R.; Lourenço, C.R.; Serrão, E.A.; Nicastro, K.R. Re-assessing the origins of the invasive mussel Mytilus galloprovincialis in southern Africa. Mar. Fresh Res. 2018, 69, 607–613. [Google Scholar] [CrossRef]
- Oyarzún, P.A.; Toro, J.E.; Garrido, O.; Briones, C.; Guiñez, R. Differences in sperm ultrastructure between Mytilus chilensis and Mytilus galloprovincialis (Bivalvia, Mytilidae): Could be used as a taxonomic trait? Latin. Am. J. Aquac. Res. 2014, 42, 172–179. [Google Scholar] [CrossRef]
- Oyarzún, P.A.; Toro, J.E.; Navarro, J.M. Comparison of the physiological energetics between Mytilus chilensis, Mytilus galloprovincialis and their hybrids, under laboratory conditions. Aquac. Res. 2013, 44, 1805–1814. [Google Scholar] [CrossRef]
- Toro, J.E.; Ojeda, J.A.; Vergara, A.M.; Castro, G.C.; Alcapán, A.C. Molecular characterization of the Chilean blue mussel (Mytilus chilensis Hupe 1854) demonstrates evidence for the occurrence of Mytilus galloprovincialis in southern Chile. J. Shell. Res. 2005, 24, 1117–1121. [Google Scholar]
- Tarifeño, E.; Galleguillos, R.; Llanos-Rivera, A.; Arriagada, D.; Ferrada, S.; Canales-Aguirre, C.B.; Seguel, M. Erroneous identification of the mussel, Mytilus galloprovincialis (Lamarck 1819) as the specie, Mytilus chilensis (Hupé 1854) in the Bay of Concepcion, Chile. Gayana 2012, 76, 167–172. [Google Scholar] [CrossRef]
- Gérard, K.; Bierne, N.; Borsa, P.; Chenuil, A.; Féral, J.-P. Pleistocene separation of mitochondrial lineages of Mytilus spp. mussels from Northern and Southern Hemispheres and strong genetic differentiation among southern populations. Mol. Phy. Evol. 2008, 49, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Toro, J.E.; Oyarzún, P.A.; Nuñez, J.J. Proyecto FIPA Nº 2015-14 Caracterización Genética y Distribución Espacial del Género Mytilus en Chile. 2017. Available online: https://www.subpesca.cl/fipa/613/articles-92071_informe_final.pdf (accessed on 28 February 2024).
- Díaz-Puente, B.; Pita, A.; Uribe, J.; Cuéllar-Pinzón, J.; Guiñez, R.; Presa, P. A biogeography-based management for Mytilus chilensis: The genetic hodgepodge of Los Lagos versus the pristine hybrid zone of the Magellanic ecotone. Aquat. Conserv. Mar. Fresh Ecosyst. 2020, 30, 412–425. [Google Scholar] [CrossRef]
- Toro, J.E.; Oyarzún, P.A.; Peñaloza, C.; Alcapán, A.; Videla, V.; Tilleria, J.; Astorga, M.; Martinez, V. Production and performance of pure species and hybrid larvae and spot of Mytilus chilensis and Mytilus galloprovincialis from laboratory crosses. Lat. Am. J. Aqu. Res. 2012, 40, 243–247. [Google Scholar] [CrossRef]
- CABI. 2021. Available online: https://www.cabi.org/isc/datasheet/73756 (accessed on 28 February 2024).
- Lee, C.E. Evolutionary genetics of invasive species. Trends Ecol. Evol. 2002, 17, 386–391. [Google Scholar] [CrossRef]
- Santaclara, F.J.; Espineira, M.; Cabado, G.; Aldasoro, A.; Gonzalez-Lavín, N.; Vieites, J.M. Development of a method for the genetic identification of mussel species belonging to Mytilus, Perna, Aulacomya, and other genera. J. Agric. Food Chem. 2006, 54, 8461–8470. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 5, 294–299. [Google Scholar]
- Palumbi, S.R.; Martin, A.P.; Kessing, B.D.; McMillan, W.O. Detecting population structure using mitochondrial DNA. In Genetic Ecology of Whales and Dolphins; Hoelzel, A.R., Ed.; International Whaling Commission: Cambridge, UK, 1991; pp. 271–278. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef]
- Guillot, G.; Mortier, F.; Estoup, A. GENELAND: A computer package for landscape genetics. Mol. Ecol. Notes 2005, 5, 712–715. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2010. Available online: http://www.r-project.org/ (accessed on 28 February 2024).
- Lischer, H.E.L.; Excoffier, L. PGDSpider: An automated data conversion tool for connecting population genetics and genomics programs. Bioinformatics 2012, 28, 298–299. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. Partitionfinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Rogers, A.R.; Harpending, H. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A.; Shapiro, B.; Pybus, O.G. Bayesian coalescent inference of past population dynamics from molecular sequences. Mol. Biol. Evol. 2005, 22, 1185–1192. [Google Scholar] [CrossRef]
- Ho, S.Y.W.; Shapiro, B. Skyline-plot methods for estimating demographic history from nucleotide sequences. Mol. Ecol. Resour. 2011, 11, 423–434. [Google Scholar] [CrossRef]
- Wares, J.P.; Cunningham, C.W. Phylogeography and historical ecology of the North Atlantic intertidal. Evolution 2001, 55, 2455–2469. [Google Scholar]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef]
- Bazinet, A.L.; Zwickl, D.J.; Cummings, M.P. A gateway for phylogenetic analysis powered by grid computing featuring GARLI 2.0. Syst. Biol. 2014, 63, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Nylander, J.A.A.; Ronquist, F.; Huelsenbeck, J.P.; Nieves-Aldrey, J. Bayesian phylogenetic analysis of combined data. Syst. Biol. 2004, 53, 47–67. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Rannala, B. Frequentist properties of bayesian posterior probabilities of phylogenetic trees under simple and complex substitution models. Syst. Biol. 2004, 53, 904–913. [Google Scholar] [CrossRef]
- Kingman, J.F.C. The coalescent. Stoch. Process. Their Appl. 1982, 13, 235–248. [Google Scholar] [CrossRef]
- Goodall-Copestake, W.P.; Tarling, G.A.; Murphy, E.J. On the comparison of population-level estimates of haplotype and nucleotide diversity: A case study using the gene cox1 in animals. Heredity 2012, 109, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Camus, P.A. Biogeografía marina de Chile continental. Rev. Chil. Hist. Nat. 2001, 74, 587–617. [Google Scholar] [CrossRef]
- Lancellotti, D.A.; Vásquez, J.A. Biogeographical patterns of benthic macroinvertebrates in the Southeastern Pacific littoral. J. Biogeogr. 1999, 26, 1001–1006. [Google Scholar] [CrossRef]
- Jaramillo, E. The sand beach ecosystem of Chile. In Coastal Marine Ecosystems of Latin America Ecological Studies (Analysis and Synthesis); Seeliger, U.K.B., Ed.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 219–227. [Google Scholar]
- Katolikova, M.; Khaitov, V.; Väinölä, R.; Gantsevich, M.; Strelkov, P. Genetic, ecological and morphological distinctness of the blue mussels Mytilus trossulus Gould and M. edulis L. in the White Sea. PLoS ONE 2016, 11, e0152963. [Google Scholar]
- FAO. CWP Handbook of Fishery Statistical Standards. Section J: Aquaculture; FAO: Roma, Italy, 2017; Available online: http://www.fao.org/fishery/cwp/handbook/J./en (accessed on 28 February 2024).
- SUBPESCA. 2017. Available online: https://www.subpesca.cl/portal/616/w3-article-843.html (accessed on 25 July 2022).
- Astorga, M.P.; Vargas, J.; Valenzuela, A.; Molinet, C.; Marín, S.L. Population genetic structure and differential selection in mussel Mytilus chilensis. Aquac. Res. 2017, 49, 919–927. [Google Scholar] [CrossRef]
- Larraín, M.A.; Diaz, N.F.; Lamas, C.; Uribe, C.; Jilberto, F.; Araneda, C. Heterologous microsatellite-based genetic diversity in blue mussel (Mytilus chilensis) and differentiation among localities in southern Chile. Lat. Am. J. Aquac. Res. 2015, 43, 998–1010. [Google Scholar] [CrossRef]
- Toro, J.; Innes, D.J.; Thompson, R.J. Genetic variation among life-history stages of mussels in a Mytilus edulis-M-trossulus hybrid zone. Mar. Biol. 2004, 145, 713–725. [Google Scholar] [CrossRef]
- Toro, J.E.; Castro, G.C.; Ojeda, J.A.; Vergara, A.M. Allozymic variation and differentiation in the chilean blue mussel, Mytilus chilensis, along its natural distribution. Gen. Mol. Biol. 2006, 29, 174–179. [Google Scholar] [CrossRef]
- Carrasco, A.V.; Astorga, M.; Cisterna, A.; Farias, A.; Espinoza, V.; Uriarte, I. Pre-feasibility Study for the Installation of a Chilean Mussel Mytilus chilensis (Hupé, 1854) Seed Hatchery in the Lakes Region, Chile. Fish. Aquac. J. 2014, 5, 102. [Google Scholar]
- MarineTraffic. 2022. Available online: https://www.marinetraffic.com/en/ais/home/centerx:-73.5/centery:-36.8/zoom:9 (accessed on 28 February 2024).
- Ntuli, N.N.; Nicastro, K.R.; Zardi, G.I.; Assis, J.; McQuaid, C.D.; Teske, P.R. Rejection of the genetic implications of the “Abundant Centre Hypothesis” in marine mussels. Sci. Rep. 2020, 10, 604. [Google Scholar] [CrossRef]
- Forrest, B.M.; Gardner, J.P.A.; Taylor, M. Internal borders for the management of invasive marine species. J. Appl. Ecol. 2009, 46, 46–54. [Google Scholar] [CrossRef]
- Glor, R.E.; Warren, D. Testing ecological explanations for biogeographic boundaries. Evolution 2010, 65, 673–683. [Google Scholar] [CrossRef]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.l.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef]
- MMA. 2022. Available online: https://especies-exoticas.mma.gob.cl/index.php/el-problema-de-eei-en-chile/ (accessed on 25 July 2022).
- Ferguson, R. The Effectiveness of Australia’s Response to the Black Striped Mussel Incursion in Darwin, Australia. A Report of the Marine Pest Incursion Management Workshop, 27–28 August 1999. Report Published by Environment Australia. 2000. Available online: https://www.awe.gov.au/sites/default/files/documents/bsmfinalreport.pdf (accessed on 28 February 2024).
- Hopkins, G.A.; Forrest, B.M.; Jiang, W.M.; Gardner, J.P.A. Successful eradication of a non-indigenous marine bivalve from a subtidal soft-sediment environment. J. Appl. Ecol. 2011, 48, 424–431. [Google Scholar] [CrossRef]
- Riginos, C.; Hickerson, M.J.; Henzler, C.M.; Cunningham, C.W. Differential patterns of male and female mtDNA exchange across the Atlantic Ocean in the blue mussel, Mytilus edulis. Evolution. 2004, 58, 2438–2451. [Google Scholar] [PubMed]
- Zardi, G.I.; McQuaid, C.D.; Teske, P.R.; Barker, N.P. Unexpected genetic structure of mussel populations in South Africa: Indigenous Perna perna and invasive Mytilus galloprovincialis. Mar. Ecol. Prog. Ser. 2007, 337, 135–144. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species and Country/Site | Map Code | Coordinates | n |
|---|---|---|---|
| M. galloprovincialis | |||
| CHILE | |||
| Isla Quiriquinas | IQ | 36°37′49.9″ S; 73°03′09.2″ W | 20 |
| Lota | LT | 37°04′27.1″ S; 73°09′53.4″ W | 20 |
| Tubul | TU | 37°13′37.3″ S; 73°26′02.2″ W | 20 |
| Llico | LL | 37°10′06.9″ S; 73°33′41.5″ W | 20 |
| SPAIN | |||
| Moaña-Pontevedra | MP | 42°16′28.5″ N; 8°43′48.3″ W | 20 |
| SOUTH AFRICA | |||
| Port Elizabeth | PE | 33°58′52.0″ S; 25°39′36.0″ E | 20 |
| M. chilensis | |||
| CHILE | |||
| Puerto Saavedra | PS | 38°46′44.7″ S; 73°24′32.3″ W | 20 |
| Chaihuín | CH | 39°56′40.4″ S; 73°34′40.4″ W | 20 |
| Maullín | MA | 41°37′25.3″ S; 73°35′36.9″ W | 20 |
| Quillaipe | QL | 41°32′59.5″ S; 72°45′14.0″ W | 20 |
| Huildad | HU | 43°03′02.8″ S; 73°34′21.1″ W | 20 |
| Puerto Cisnes | PC | 44°44′11.4″ S; 72°41′07.9″ W | 20 |
| Estero Fanny | BF | 53°05′04.6″ S; 72°18′39.6″ W | 20 |
| Caleta Pescadores | CP | 53°21′06.2″ S; 70°57′27.8″ W | 20 |
| Total number of mussels | 280 | ||
| Species/Locality | K | H (SD) | S | Π | π | Tajima’s D | Fu’s FS | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| COI | 16S | COI | 16S | COI | 16S | COI | 16S | COI | 16S | COI | 16S | COI | 16S | |
| M. galloprovincialis | ||||||||||||||
| CHILE (introduced) | ||||||||||||||
| Isla Quiriquina | 7 | 8 | 0.832 (0.049) | 0.826 (0.061) | 31 | 13 | 10.66 | 3.821 | 0.01669 | 0.00832 | 0.58 | 0.16 | 5.14 | −0.10 |
| Lota | 7 | 8 | 0.865 (0.042) | 0.754 (0.090) | 28 | 11 | 8.392 | 3.094 | 0.01349 | 0.00674 | 0.05 | −0.36 | 3.57 | −0.89 |
| Tubul | 9 | 5 | 0.889 (0.049) | 0.905 (0.103) | 21 | 9 | 6.157 | 4.190 | 0.00980 | 0.00913 | −0.15 | 0.74 | 0.24 | −0.05 |
| Llico | 12 | 6 | 0.906 (0.050) | 0.810 (0.057) | 34 | 10 | 10.60 | 3.007 | 0.01754 | 0.00655 | −0.20 | 0.12 | −0.22 | 0.68 |
| All locations | 14 | 16 | 0.857 (0.020) | 0.809 (0.035) | 28 | 18 | 5.065 | 3.384 | 0.00863 | 0.00737 | −0.45 | −0.48 | 0.50 | −3.28 |
| SPAIN (native) | ||||||||||||||
| Moaña-Pontevedra | 6 | 7 | 0.757 (0.091) | 0.872 (0.067) | 25 | 10 | 7.191 | 3.538 | 0.01124 | 0.00751 | ns | ns | ns | ns |
| SOUTH AFRICA (introduced) | ||||||||||||||
| Port Elizabeth | 20 | 7 | 1.000 (0.016) | 0.742 (0.071) | 50 | 10 | 11.84 | 2.116 | 0.01830 | 0.00449 | ns | ns | ns | ns |
| M. chilensis | ||||||||||||||
| CHILE (native) | ||||||||||||||
| Puerto Saavedra | 14 | 9 | 0.947 (0.038) | 0.678 (0.122) | 20 | 16 | 3.257 | 2.292 | 0.00484 | 0.00488 | −1.87 | −2.11 | −8.43 | −2.93 |
| Chaihuín | 13 | 9 | 0.942 (0.037) | 0.678 (0.122) | 15 | 14 | 2.532 | 1.936 | 0.00375 | 0.00412 | −1.68 | −2.05 | −8.37 | −3.64 |
| Maullín | 14 | 8 | 0.971 (0.032) | 0.860 (0.055) | 19 | 8 | 3.632 | 1.721 | 0.00539 | 0.00366 | −1.39 | −0.97 | −9.04 | −3.23 |
| Quillaipe | 12 | 7 | 0.932 (0.035) | 0.824 (0.064) | 20 | 8 | 3.789 | 1.662 | 0.00561 | 0.00354 | −1.25 | −1.05 | −3.96 | −2.15 |
| Huildad | 10 | 8 | 0.914 (0.056) | 0.867 (0.067) | 14 | 38 | 2.895 | 5.590 | 0.00428 | 0.01189 | −1.30 | −2.28 | −4.31 | −0.14 |
| Puerto Cisnes | 15 | 9 | 0.942 (0.043) | 0.879 (0.043) | 21 | 9 | 3.647 | 1.932 | 0.00540 | 0.00411 | −1.46 | −1.10 | −8.89 | −3.46 |
| Bahía Fanny | 14 | 10 | 0.947 (0.034) | 0.711 (0.113) | 21 | 14 | 3.089 | 1.816 | 0.00457 | 0.00386 | −1.82 | −1.98 | −8.28 | −5.01 |
| Caleta Pescadores | 15 | 11 | 0.921 (0.055) | 0.763 (0.103) | 28 | 10 | 3.326 | 1.442 | 0.00492 | 0.00307 | −2.26 | −1.90 | −9.63 | −8.16 |
| All locations | 69 | 49 | 0.943 (0.011) | 0.808 (0.030) | 76 | 70 | 3.161 | 2.289 | 0.00470 | 0.00488 | −2.48 | −2.64 | −94.83 | −59.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oyarzún, P.A.; Toro, J.E.; Nuñez, J.J.; Ruiz-Tagle, G.; Gardner, J.P.A. The Mediterranean Mussel Mytilus galloprovincialis (Mollusca: Bivalvia) in Chile: Distribution and Genetic Structure of a Recently Introduced Invasive Marine Species. Animals 2024, 14, 823. https://doi.org/10.3390/ani14060823
Oyarzún PA, Toro JE, Nuñez JJ, Ruiz-Tagle G, Gardner JPA. The Mediterranean Mussel Mytilus galloprovincialis (Mollusca: Bivalvia) in Chile: Distribution and Genetic Structure of a Recently Introduced Invasive Marine Species. Animals. 2024; 14(6):823. https://doi.org/10.3390/ani14060823
Chicago/Turabian StyleOyarzún, Pablo A., Jorge E. Toro, José J. Nuñez, Gonzalo Ruiz-Tagle, and Jonathan P. A. Gardner. 2024. "The Mediterranean Mussel Mytilus galloprovincialis (Mollusca: Bivalvia) in Chile: Distribution and Genetic Structure of a Recently Introduced Invasive Marine Species" Animals 14, no. 6: 823. https://doi.org/10.3390/ani14060823