
Prevalence of Gastrointestinal Parasites in Zoo Animals and Phylogenetic Characterization of Toxascaris leonina (Linstow, 1902) and Baylisascaris transfuga (Rudolphi, 1819) in Jiangsu Province, Eastern China
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
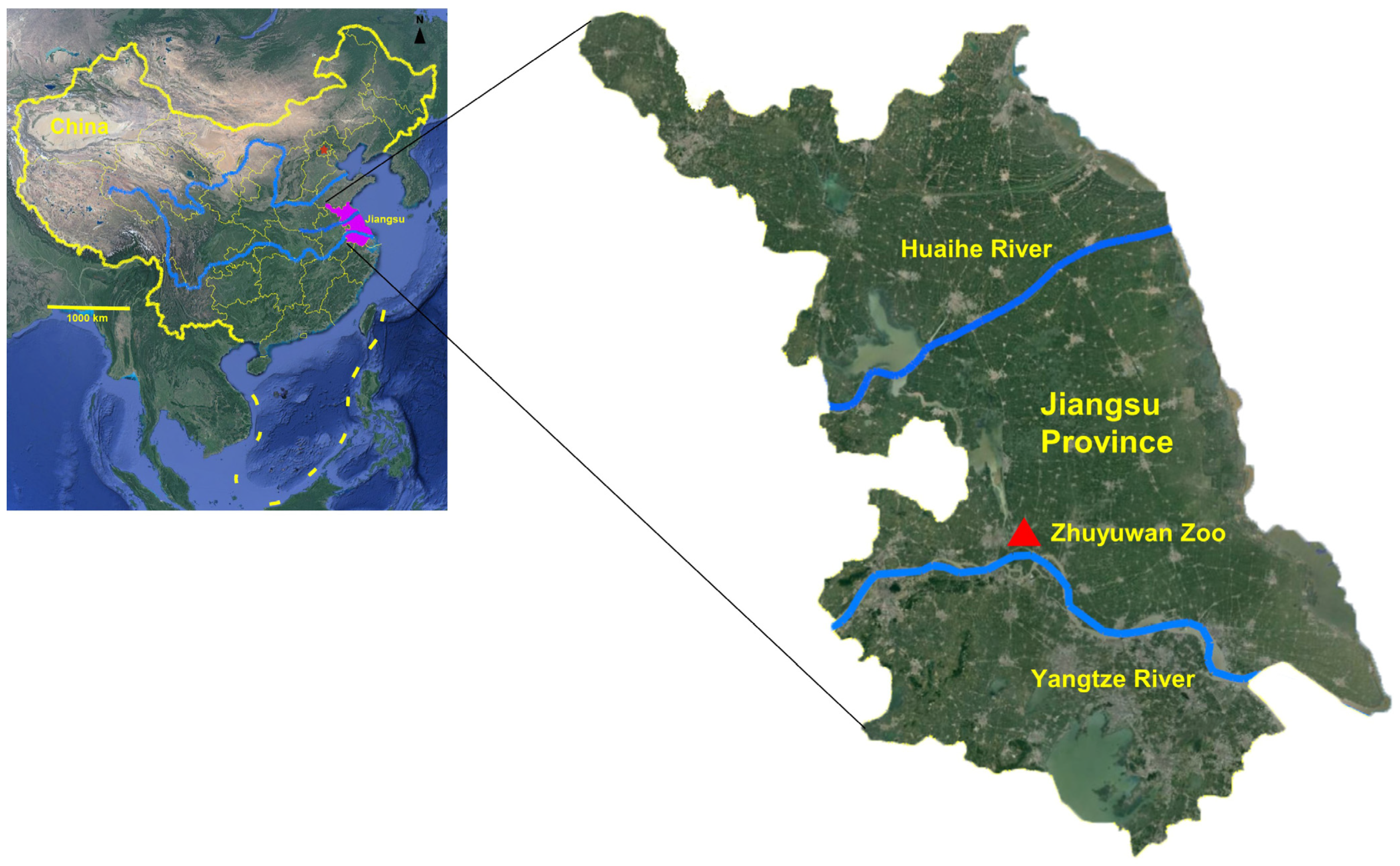
2.1. Study Site
2.2. Sample Collection
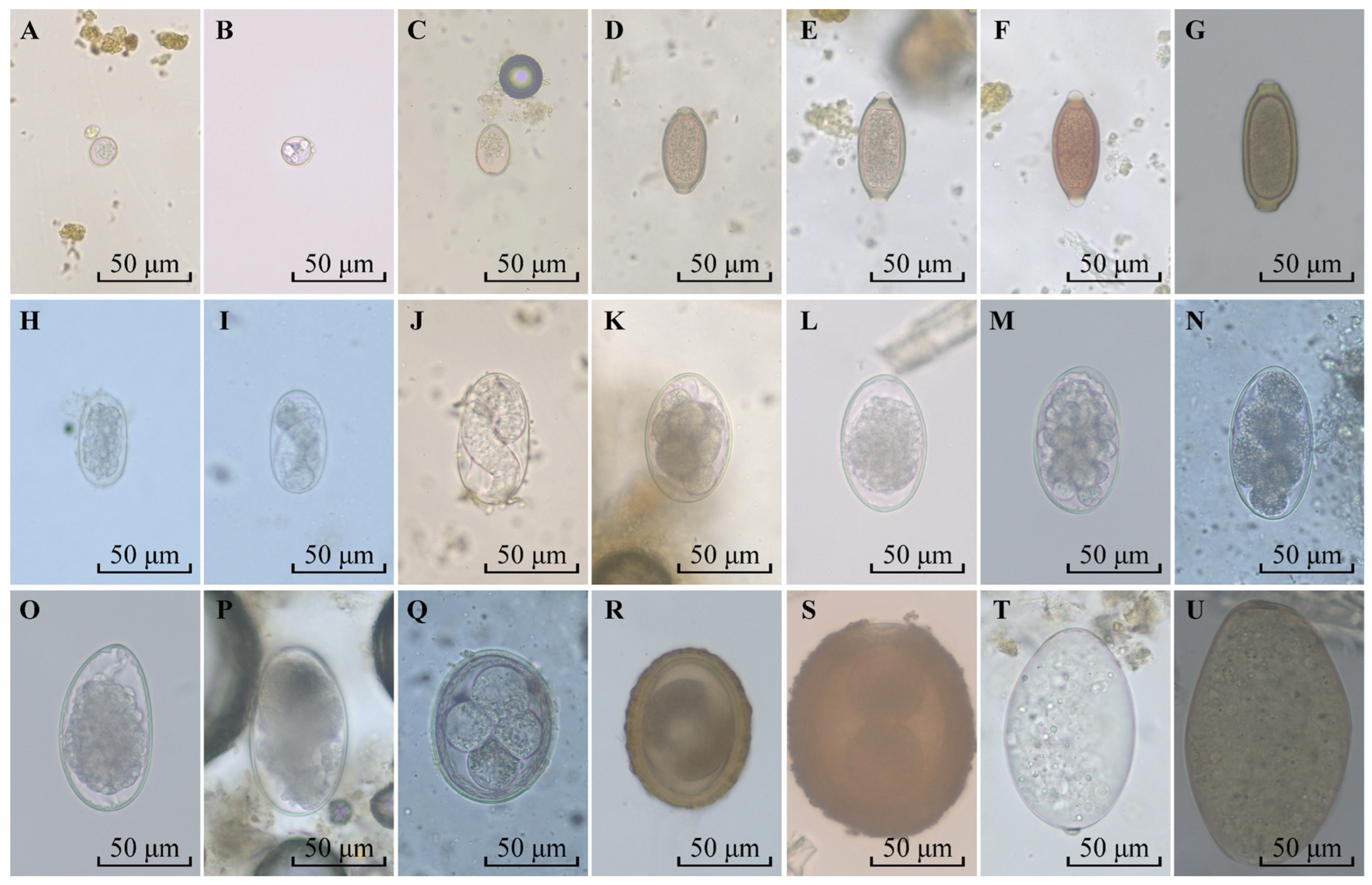
2.3. Microscopy
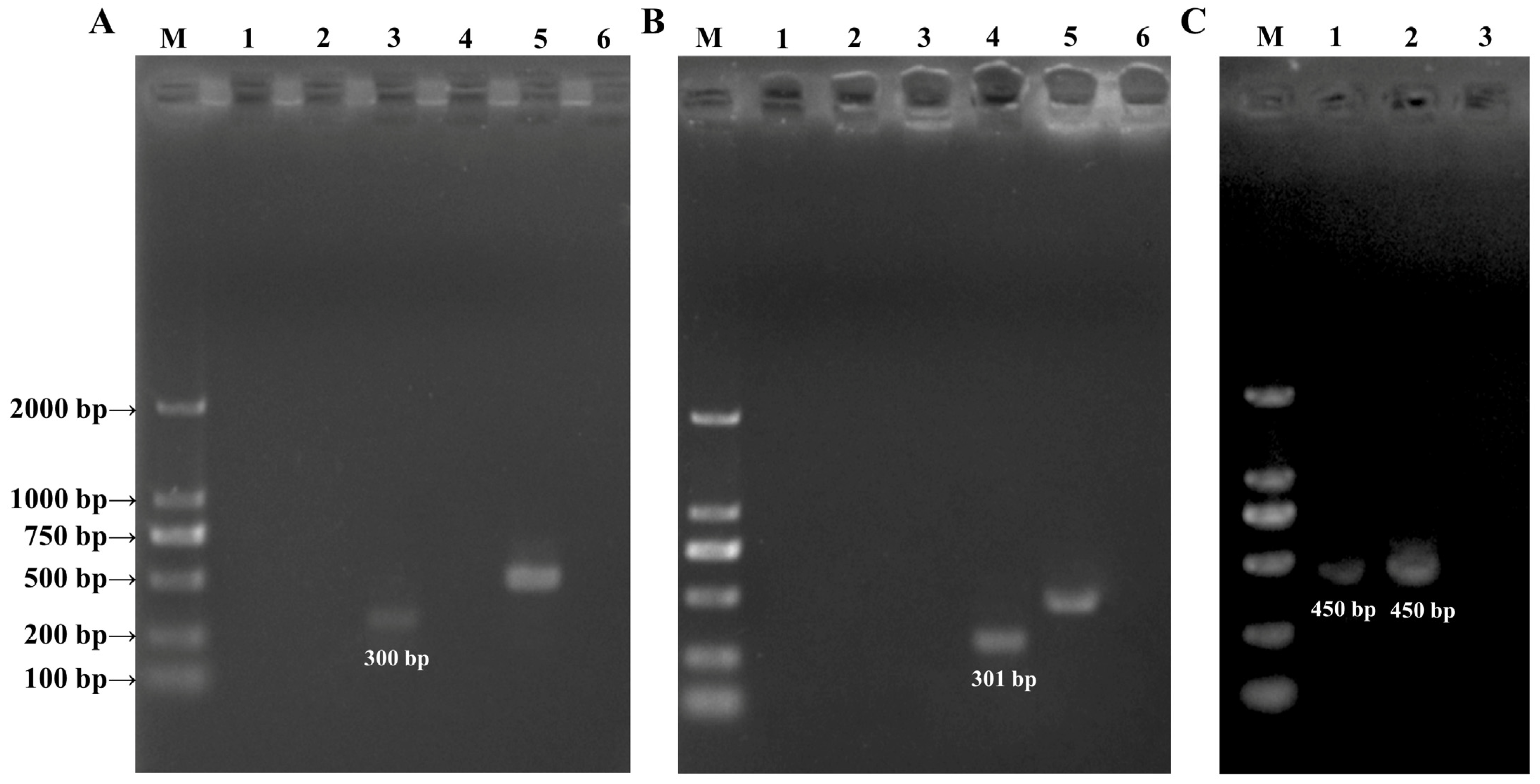
2.4. DNA Isolation and PCR Amplification
2.5. Sequence and Phylogenetic Analysis
3. Results
3.1. Occurrence of Intestinal Parasites
3.2. PCR Amplification Analysis
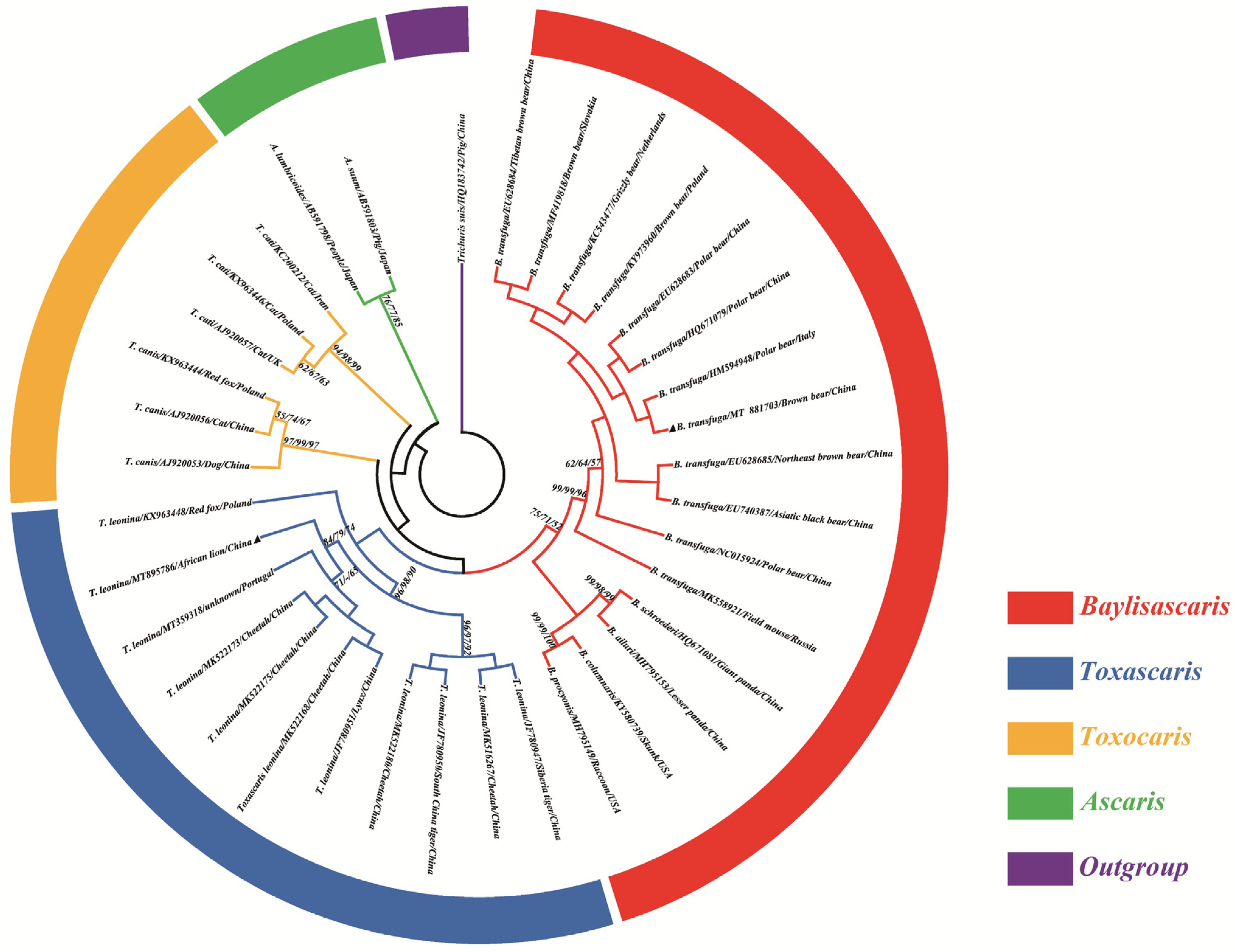
3.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christe, P. Loss of biodiversity: A one-way ticket to new emerging diseases? Rev. Med. Suisse 2023, 19, 842–844. [Google Scholar] [PubMed]
- Edwards, K.L.; Edes, A.N.; Brown, J.L. Stress, well-being and reproductive success. Adv. Exp. Med. Biol. 2019, 1200, 91–162. [Google Scholar] [PubMed]
- Serota, M.W.; Barker, K.J.; Gigliotti, L.C.; Maher, S.M.L.; Shawler, A.L.; Zuckerman, G.R.; Xu, W.; Verta, G.; Templin, E.; Andreozzi, C.L.; et al. Incorporating human dimensions is associated with better wildlife translocation outcomes. Nat. Commun. 2023, 14, 2119. [Google Scholar] [CrossRef] [PubMed]
- Booth, M. Climate change and the neglected tropical diseases. Adv. Parasitol. 2018, 100, 39–126. [Google Scholar] [PubMed]
- Locklear, T.R.; Videla, R.; Breuer, R.M.; Mulon, P.Y.; Passmore, M.; Mochel, J.P.; Gerhold, R.; Schaefer, J.J.; Smith, J.S. Presentation, clinical pathology abnormalities, and identification of gastrointestinal parasites in camels (Camelus bactrianus and Camelus dromedarius) presenting to two north American veterinary teaching hospitals. A retrospective study: 1980–2020. Front. Vet. Sci. 2021, 8, 651672. [Google Scholar] [CrossRef] [PubMed]
- Trogu, T.; Formenti, N.; Marangi, M.; Viganò, R.; Bionda, R.; Giangaspero, A.; Lanfranchi, P.; Ferrari, N. Detection of zoonotic Cryptosporidium ubiquitum in alpine wild ruminants. Pathogens 2021, 10, 655. [Google Scholar] [CrossRef]
- Espinosa García-San Román, J.; Quesada-Canales, Ó.; Arbelo Hernández, M.; Déniz Suárez, S.; Castro-Alonso, A. Veterinary Education and Training on Non-Traditional Companion Animals, Exotic, Zoo, and Wild Animals: Concepts Review and Challenging Perspective on Zoological Medicine. Vet. Sci. 2023, 10, 357. [Google Scholar] [CrossRef]
- Bedoya-Pérez, M.A.; Ward, M.P.; Loomes, M.; McGregor, I.S.; Crowther, M.S. The effect of COVID-19 pandemic restrictions on an urban rodent population. Sci. Rep. 2021, 11, 12957. [Google Scholar] [CrossRef]
- Aguirre, A.A. Essential veterinary education in zoological and wildlife medicine: A global perspective. Rev. Sci. Tech. 2009, 28, 605–610. [Google Scholar] [CrossRef]
- Vlaminck, J.; Levecke, B.; Vercruysse, J.; Geldhof, P. Advances in the diagnosis of Ascaris suum infections in pigs and their possible applications in humans. Parasitology 2014, 141, 1904–1911. [Google Scholar] [CrossRef]
- Jourdan, P.M.; Lamberton, P.H.L.; Fenwick, A.; Addiss, D.G. Soil-transmitted helminth infections. Lancet 2018, 391, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Lamberton, P.H.; Jourdan, P.M. Human Ascariasis: Diagnostics Update. Curr. Trop. Med. Rep. 2015, 2, 189–200. [Google Scholar] [CrossRef]
- Li, M.W.; Lin, R.Q.; Chen, H.H.; Sani, R.A.; Song, H.Q.; Zhu, X.Q. PCR tools for the verification of the specific identity of ascaridoid nematodes from dogs and cats. Mol. Cell Probes 2007, 21, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Fogt-Wyrwas, R.; Mizgajska-Wiktor, H.; Pacoń, J.; Jarosz, W. Intraspecific variation between the ITS sequences of Toxocara canis, Toxocara cati and Toxascaris leonina from different host species in south-western Poland. J. Helminthol. 2013, 87, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Li, Y.; Gu, X.; Liu, Y.; Zhou, X.; Wang, L.; He, R.; Peng, X.; Yang, G. Molecular characterization of ascaridoid parasites from captive wild carnivores in China using ribosomal and mitochondrial sequences. Parasit. Vectors 2020, 13, 382. [Google Scholar] [CrossRef] [PubMed]
- Blouin, M.S. Molecular prospecting for cryptic species of nematodes: Mitochondrial DNA versus internal transcribed spacer. Int. J. Parasitol. 2002, 32, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.A.; Coop, R.L.; Wall, R.L. Veterinary Parasitology; Blackwell Publishing: Oxford, UK, 2007. [Google Scholar]
- Ministry of Agriculture, Fisheries and Food. Manual of Veterinary Parasitological Laboratory Techniques, 2nd ed.; Technical Bulletin No.18.; Her Majesty’s Stationary Office: London, YK, 1977. [Google Scholar]
- Gasser, R.B.; Zhu, X.; McManus, D.P. NADH dehydrogenase subunit 1 and cytochrome c oxidase subunit I sequences compared for members of the genus Taenia (Cestoda). Int. J. Parasitol. 1999, 29, 1965–1970. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, X. Investigation of intestinal parasites in wild animals at Xining Wildlife Park. Heilongjiang Anim. Sci. Vet. Med. 2014, 2, 139–140. [Google Scholar]
- Kvapil, P.; Kastelic, M.; Dovc, A.; Bartova, E.; Cizek, P.; Lima, N.; Strus, S. An eight-year survey of the intestinal parasites of carnivores, hoofed mammals, primates, ratites and reptiles in the Ljubljana zoo in Slovenia. Folia Parasitol. 2017, 64, 13. [Google Scholar] [CrossRef]
- Maesano, G.; Capasso, M.; Ianniello, D.; Cringoli, G.; Rinaldi, L. Parasitic infections detected by FLOTAC in zoo mammals from Warsaw, Poland. Acta Parasitol. 2014, 59, 343–353. [Google Scholar] [CrossRef]
- Li, M.; Zhao, B.; Li, B.; Wang, Q.; Niu, L.; Deng, J.; Gu, X.; Peng, X.; Wang, T.; Yang, G. Prevalence of gastrointestinal parasites in captive non-human primates of twenty-four zoological gardens in China. J. Med. Primatol. 2015, 44, 168–173. [Google Scholar] [CrossRef]
- Dhakal, P.; Sharma, H.P.; Shah, R.; Thapa, P.J.; Pokheral, C.P. Copromicroscopic study of gastrointestinal parasites in captive mammals at Central Zoo, Lalitpur, Nepal. Vet. Med. Sci. 2023, 9, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Cian, A.; El Safadi, D.; Osman, M.; Moriniere, R.; Gantois, N.; Benamrouz-Vanneste, S.; Delgado-Viscogliosi, P.; Guyot, K.; Li, L.L.; Monchy, S.; et al. Molecular Epidemiology of Blastocystis sp. in Various Animal Groups from Two French Zoos and Evaluation of Potential Zoonotic Risk. PLoS ONE 2017, 12, e0169659. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Dong, H.; Hu, L.; Peng, G.; Chen, Z.; Zhou, Z.; Wan, J.; Jian, F.; Zhang, L. Investigation on the Prevalence of Intestinal Parasites in Wild Animal Zoo. China Anim. Husb. Vet. Med. 2011, 38, 188–190. [Google Scholar]
- Capasso, M.; Maurelli, M.P.; Ianniello, D.; Alves, L.C.; Amadesi, A.; Laricchiuta, P.; Silvestre, P.; Campolo, M.; Cringoli, G.; Rinaldi, L. Use of Mini-FLOTAC and Fill-FLOTAC for rapidly diagnosing parasitic infections in zoo mammals. Rev. Bras. Parasitol. Vet. 2019, 28, 168–171. [Google Scholar] [CrossRef]
- Holsback, L.; Cardoso, M.J.; Fagnani, R.; Patelli, T.H. Natural infection by endoparasites among free-living wild animals. Rev. Bras. Parasitol. Vet. 2013, 22, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Munene, E.; Otsyula, M.; Mbaabu, D.A.; Mutahi, W.T.; Muriuki, S.M.; Muchemi, G.M. Helminth and protozoan gastrointestinal tract parasites in captive and wild-trapped African non-human primates. Vet. Parasitol. 1998, 78, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Albuixech-Martí, S.; Lynch, S.A.; Culloty, S.C. Connectivity dynamics in Irish mudflats between microorganisms including Vibrio spp., common cockles Cerastoderma edule, and shorebirds. Sci. Rep. 2021, 11, 22159. [Google Scholar] [CrossRef]
- Parr, N.A.; Fedigan, L.M.; Kutz, S.J. A coprological survey of parasites in white-faced capuchins (Cebus capucinus) from Sector Santa Rosa, ACG, Costa Rica. Folia Primatol. 2013, 84, 102–114. [Google Scholar] [CrossRef]
- Barutzki, D.; Schaper, R. Results of parasitological examinations of faecal samples from cats and dogs in Germany between 2003 and 2010. Parasitol. Res. 2011, 109 (Suppl. S1), S45–S60. [Google Scholar] [CrossRef]
- Rondon, S.; Ortiz, M.; Leon, C.; Galvis, N.; Link, A.; Gonzalez, C. Seasonality, richness and prevalence of intestinal parasites of three neotropical primates (Alouatta seniculus, Ateles hybridus and Cebus versicolor) in a fragmented forest in Colombia. Int. J. Parasitol. Parasites Wildl. 2017, 6, 202–208. [Google Scholar] [CrossRef] [PubMed]
- García-Sánchez, A.M.; Miller, A.Z.; Caldeira, A.T.; Cutillas, C. Bacterial communities from Trichuris spp. A contribution to deciphering the role of parasitic nematodes as vector of pathogens. Acta Trop. 2022, 226, 106277. [Google Scholar] [CrossRef] [PubMed]
- Brodie, J.F.; Mohd-Azlan, J.; Chen, C.; Wearn, O.R.; Deith, M.C.M.; Ball, J.G.C.; Slade, E.M.; Burslem, D.; Teoh, S.W.; Williams, P.J.; et al. Landscape-scale benefits of protected areas for tropical biodiversity. Nature 2023, 620, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: Major findings, mechanisms and recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Polakovicova, N.; Adji, A.V.; Myhill, L.J.; Williams, A.R. Whipworm Infection in Mice Increases Coinfection of Enteric Pathogens but Promotes Clearance of Ascaris Larvae from the Lungs. J. Infect. Dis. 2023, 227, 1428–1432. [Google Scholar] [CrossRef] [PubMed]
- Elaphurus Davidianus. The IUCN Red List of Threatened Species. Available online: https://dx.doi.org/10.2305/IUCN.UK.2016-2.RLTS.T7121A22159785.en (accessed on 1 March 2021).
- Gu, X.H.; Guo, N.; Chen, H.X.; Sitko, J.; Li, L.W.; Guo, B.Q.; Li, L. Mitogenomic phylogenies suggest the resurrection of the subfamily Porrocaecinae and provide insights into the systematics of the superfamily Ascaridoidea (Nematoda: Ascaridomorpha), with the description of a new species of Porrocaecum. Parasit. Vectors 2023, 16, 275. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Chen, L.; Chen, Y.; Shen, N.; Hua, R.; Yang, G. Evaluation of the immunoprotective effects of eight recombinant proteins from Baylisascaris schroederi in mice model. Parasit. Vectors 2023, 16, 254. [Google Scholar] [CrossRef]
- Li, Y.; Niu, L.; Wang, Q.; Zhang, Z.; Chen, Z.; Gu, X.; Xie, Y.; Yan, N.; Wang, S.; Peng, X.; et al. Molecular characterization and phylogenetic analysis of ascarid nematodes from twenty-one species of captive wild mammals based on mitochondrial and nuclear sequences. Parasitology 2012, 139, 1329–1338. [Google Scholar] [CrossRef]
- Testini, G.; Papini, R.; Lia, R.P.; Parisi, A.; Dantas-Torres, F.; Traversa, D.; Otranto, D. New insights into the morphology, molecular characterization and identification of Baylisascaris transfuga (Ascaridida, Ascarididae). Vet. Parasitol. 2011, 175, 97–102. [Google Scholar] [CrossRef]
- Okulewicz, A.; Lonc, E.; Borgsteede, F.H. Ascarid nematodes in domestic and wild terrestrial mammals. Pol. J. Vet. Sci. 2002, 5, 277–281. [Google Scholar]
- Miller, A.D. Pathology of larvae and adults in dogs and cats. Adv. Parasitol. 2020, 109, 537–544. [Google Scholar]
- Macpherson, C.N. The epidemiology and public health importance of toxocariasis: A zoonosis of global importance. Int. J. Parasitol. 2013, 43, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, E.J. Toxocara spp. in dogs and cats in Canada. Adv. Parasitol. 2020, 109, 641–653. [Google Scholar] [PubMed]
- Figueroa, J. New records of parasites in free-ranging Andean bears from Peru. Ursus 2015, 26, 21–27. [Google Scholar] [CrossRef]
- Sapp, S.G.H.; Gupta, P.; Martin, M.K.; Murray, M.H.; Niedringhaus, K.D.; Pfaff, M.A.; Yabsley, M.J. Beyond the raccoon roundworm: The natural history of non-raccoon Baylisascaris species in the New World. Int. J. Parasitol. Parasites Wildl. 2017, 6, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Bauer, C. Baylisascariosis—Infections of animals and humans with ’unusual’ roundworms. Vet. Parasitol. 2013, 193, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Papini, R.; Cavicchio, P.; Casarosa, L. Experimental infection in chickens with larvae of Baylisascaris transfuga (Nematoda: Ascaridoidea). Folia Parasitol. 1993, 40, 141–143. [Google Scholar]
- Papini, R.; Demi, S.; Croce, G. Observations on the migratory behaviour of Baylisascaris transfuga larvae in rabbits. Rev. Med. Vet. 1996, 147, 893–896. [Google Scholar]
- Juránková, J.; Hofmannová, L.; Frgelecová, L.; Daněk, O.; Modrý, D. Baylisascaris transfuga (Ascaridoidea, Nematoda) from European brown bear (Ursus arctos) causing larva migrans in laboratory mice with clinical manifestation. Parasitol. Res. 2022, 121, 645–651. [Google Scholar] [CrossRef]
- Cho, S.; Egami, M.; Ohnuki, H.; Saito, Y.; Chinone, S.; Shichinohe, K.; Suganuma, M.; Akao, N. Migration behaviour and pathogenesis of five ascarid nematode species in the Mongolian gerbil Meriones unguiculatus. J. Helminthol. 2007, 81, 43–47. [Google Scholar] [CrossRef]
- Moser, W.; Schindler, C.; Keiser, J. Efficacy of recommended drugs against soil transmitted helminths: Systematic review and network meta-analysis. BMJ 2017, 358, j4307. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Primer | Sequence (5′ to 3′) |
|---|---|
| For ITS-2 | |
| TcanF | AGTATGATGGGCGCCAAT |
| NC2R | TTAGTTTCTTTTCCTCCGCT |
| TcatF | GGAGAAGTAAGATCGTGGCACGCGT |
| NC2R | TTAGTTTCTTTTCCTCCGCT |
| TaleoF | CGAACGCTCATATAACGGCATACTC |
| NC2R | TTAGTTTCTTTTCCTCCGCT |
| NITSF | TTATGAATTTTCAACATGGC |
| NITSR | GTTAGATGCTTAAATTCAGC |
| For cox1 | |
| JB3 | TTTTTTGGGCATCCTGAGGTTTAT |
| JB4.5 | TAAAGAAAGAACATAATGAAAATG |
| Species | n | No. (%) of Positive Samples for Parasite Species | ||
|---|---|---|---|---|
| Capillaria sp. | Amidostomum sp. | Eimeria spp. | ||
| Birds | 19 | 1 (5.3) | 1 (5.3) | 3 (15.8) |
| Family Dromaiidae | ||||
| Emu (Dromaius novaehollandiae) | 1 | – | – | – |
| Family Struthioidae | ||||
| Common ostrich (Struthio camelus) | 2 | – | – | – |
| Family Phoenicopteridae | ||||
| Greater flamingo (Phoenicopterus roseus) | 1 | – | – | – |
| Family Gruidae | ||||
| Manchurian crane (Grus japonensis) | 2 | – | – | – |
| Hooded crane (Grus monacha) | 1 | – | – | 1 (100) |
| White-naped crane (Grus vipio) | 1 | – | – | – |
| Siberian crane (Grus leucogeranus) | 1 | – | – | – |
| Black crowned-crane (Balearica pavonina) | 2 | 1 (50) | – | – |
| Black-necked crane (Grus nigricollis) | 1 | 1 (100) | ||
| Demoiselle crane (Anthropoides virgo) | 1 | – | – | – |
| Common crane (Grus grus) | 1 | – | – | – |
| Family Ciconiidae | ||||
| Oriental white stork (Ciconia boyciana) | 1 | – | 1 (100) | – |
| Family Phasianidae | ||||
| Green peafowl (Pavo muticus) | 1 | – | – | 1 (100) |
| Family Anatidae | ||||
| Northern pintail (Anas acuta) | 2 | – | – | – |
| Family Accipitridae | ||||
| Black vulture (Aegypius monachus) | 1 | – | – | – |
| Species | n | No. (%) of Positive Samples for Parasite Species | |
|---|---|---|---|
| Trichuris spp. | Strongyloides spp. | ||
| Primates | 19 | 3 (15.8) | 3 (15.8) |
| Family Cebidae | |||
| Black-capped capuchins (Cebus apella) | 2 | – | – |
| Squirrel monkey (Saimiri sciureus) | 3 | – | – |
| Family Lemuridae | |||
| Ring-tailed lemur (Lemur catta) | 4 | – | 3 (75) |
| Family Cercopithecidae | |||
| François’s leaf monkey (Trachypithecus francoisi) | 2 | – | – |
| Golden monkey (Rhinopithecus) | 2 | 2 (100) | – |
| Patas monkey (Erythrocebus patas) | 1 | 1 (100) | – |
| Family Hylobatidae | |||
| Lar gibbon (Hylobates lar) | 3 | – | – |
| Family Hominidae | |||
| Chimpanzee (Pan troglodytes) | 2 | – | – |
| Species | n | No. (%) of Positive Samples for Parasite Species | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Trichostrongylidae | Trichuris spp. | Strongyloides spp. | Toxascaris leonina | Baylisascaris transfuga | Parascaris equorum | Paramphistomum spp. | Fasiola spp. | Eimeria spp. | ||
| Non-primate mammals | 66 | 23 (34.8) | 3 (10.7) | 1 (3.6) | 1 (3.6) | 1 (3.6) | 1 (3.6) | 1 (1.5) | 1 (1.5) | 3 (4.5) |
| Family Camelidae | ||||||||||
| Llama (Lama glama) | 2 | 1 (50) | – | – | – | – | – | – | – | – |
| Alpaca (Vicugna pacos) | 1 | 1 (100) | 1 (100) | – | – | – | – | – | – | – |
| Family Bovidae | ||||||||||
| Blue wildebeest (Connochaetes taurinus) | 11 | 4 (36.4) | – | – | – | – | – | – | – | – |
| Gemsbok (Oryx gazella) | 10 | 10 (100) | 2 (100) | – | – | – | – | – | – | 2 (100) |
| Scimitar-horned oryx (Oryx dammah) | 4 | 4 (100) | – | – | – | – | – | – | – | – |
| Family Equidae | ||||||||||
| Pony (Equus spp., pony) | 2 | – | – | – | – | – | – | – | – | |
| Common zebra (Equus quagga) | 10 | 3 (30) | – | – | – | – | 1 (10) | – | – | – |
| Family Cervidae | ||||||||||
| Père David’s deer (Elaphurus davidianus) | 1 | – | – | – | – | – | – | 1 (100) | – | – |
| Sika deer (Cervus nippon) | 10 | – | – | – | – | – | – | – | 1 (10) | 1 (10) |
| Family Ursidae | ||||||||||
| Giant panda (Ailuropoda melanoleuca) | 2 | – | – | – | – | – | – | – | – | – |
| Brown bear (Ursus arctos) | 2 | – | – | – | – | 1 (50) | – | – | – | – |
| Family Ailuridae | ||||||||||
| Red panda (Ailurus fulgens) | 2 | – | – | 1 (50) | – | – | – | – | – | – |
| Family Felidae | ||||||||||
| Siberian tiger (Panthera tigris ssp. altaica) | 2 | – | – | – | – | – | – | – | – | – |
| African lion (Panthera leo) | 2 | – | – | – | 1 (50) | – | – | – | – | – |
| Leopard (Panthera pardus) | 1 | – | – | – | – | – | – | – | – | – |
| Family Canidae | ||||||||||
| Common wolf (Canis lupus) | 2 | – | – | – | – | – | – | – | – | – |
| Family Macropodidae | ||||||||||
| Gray kangaroo (Macropus giganteus) | 2 | – | – | – | – | – | – | – | – | – |
| Animals | Samples | Helminth Positive (%) | Protozoan Positive (%) | Total |
|---|---|---|---|---|
| Birds | 19 | 2 (10.5) | 3 (15.8) | 5 (26.3) |
| Primates | 19 | 6 (31.6) | 0 (0) | 6 (31.6) |
| Mammals | 66 | 33 (50.0) | 3 (4.5) | 33 (50.0) |
| Total | 104 | 41 (39.4) | 6 (5.8) | 44 (42.3) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, W.; Zhu, Y.; Wang, F.; Feng, Q.; Zhang, Z.; Xue, N.; Xu, X.; Hou, Z.; Liu, D.; Xu, J.; et al. Prevalence of Gastrointestinal Parasites in Zoo Animals and Phylogenetic Characterization of Toxascaris leonina (Linstow, 1902) and Baylisascaris transfuga (Rudolphi, 1819) in Jiangsu Province, Eastern China. Animals 2024, 14, 375. https://doi.org/10.3390/ani14030375
Cai W, Zhu Y, Wang F, Feng Q, Zhang Z, Xue N, Xu X, Hou Z, Liu D, Xu J, et al. Prevalence of Gastrointestinal Parasites in Zoo Animals and Phylogenetic Characterization of Toxascaris leonina (Linstow, 1902) and Baylisascaris transfuga (Rudolphi, 1819) in Jiangsu Province, Eastern China. Animals. 2024; 14(3):375. https://doi.org/10.3390/ani14030375
Chicago/Turabian StyleCai, Weimin, Yu Zhu, Feiyan Wang, Qianqian Feng, Zhizhi Zhang, Nianyu Xue, Xun Xu, Zhaofeng Hou, Dandan Liu, Jinjun Xu, and et al. 2024. "Prevalence of Gastrointestinal Parasites in Zoo Animals and Phylogenetic Characterization of Toxascaris leonina (Linstow, 1902) and Baylisascaris transfuga (Rudolphi, 1819) in Jiangsu Province, Eastern China" Animals 14, no. 3: 375. https://doi.org/10.3390/ani14030375