
Metabolic Rate and Egg Production in Japanese Quails Can Be Predicted by Assessing Growth Parameters of Laying Hens
 , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
- (1)
- To test the hypothesis that there is a relationship between egg productivity and the metabolism level in Japanese quails of various breeds such that M can be predicted through the accurate assessment of S and V of hens’ bodies;
- (2)
- To develop a robust theoretical and methodological basis for determining V and S in Japanese quail hens such that egg productivity can be accurately predicted.
2. Materials and Methods
2.1. Experimental Birds
2.2. Use of Mathematical Similarity Principles
- i.
- Among body measurements, we focused on the bird’s body, leaving out parts such as the head, legs, and unfolded wings. This constitutes about 95% of the total bird’s V and contains the main internal organs responsible for metabolic processes. The commercial management of quails prevents them from actively using their wings and, consequently, hampers their effect on H.
- ii.
- The calculation of the S and V values were performed according to a single mathematical principle that made it possible to level out any possible errors in their measurement.
- iii.
- For calculations, we used the principle of mathematical similarity.
2.3. Determination of S and V of the Quail Body
2.4. Measured Traits and Statistics
3. Results and Discussion
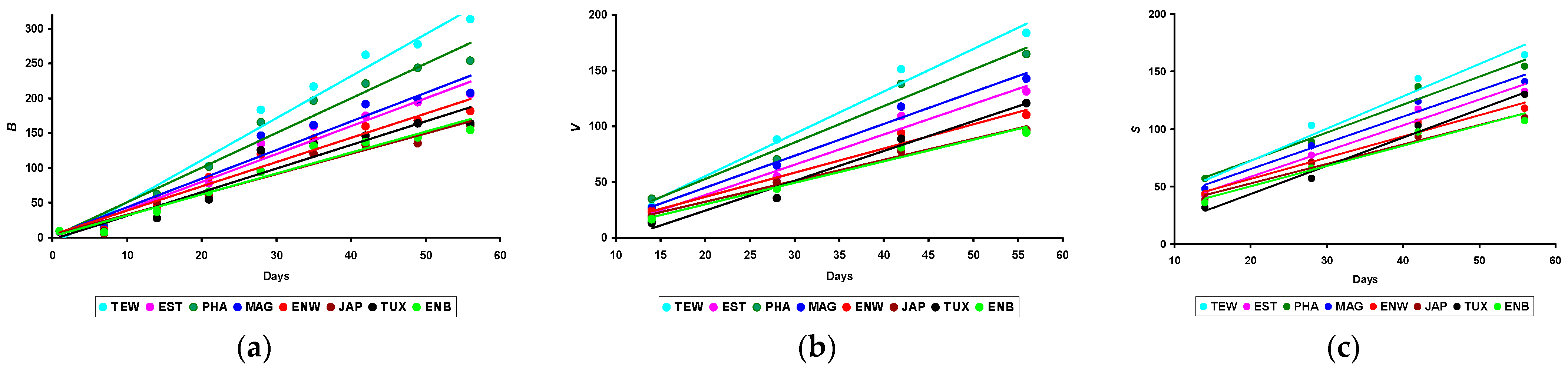
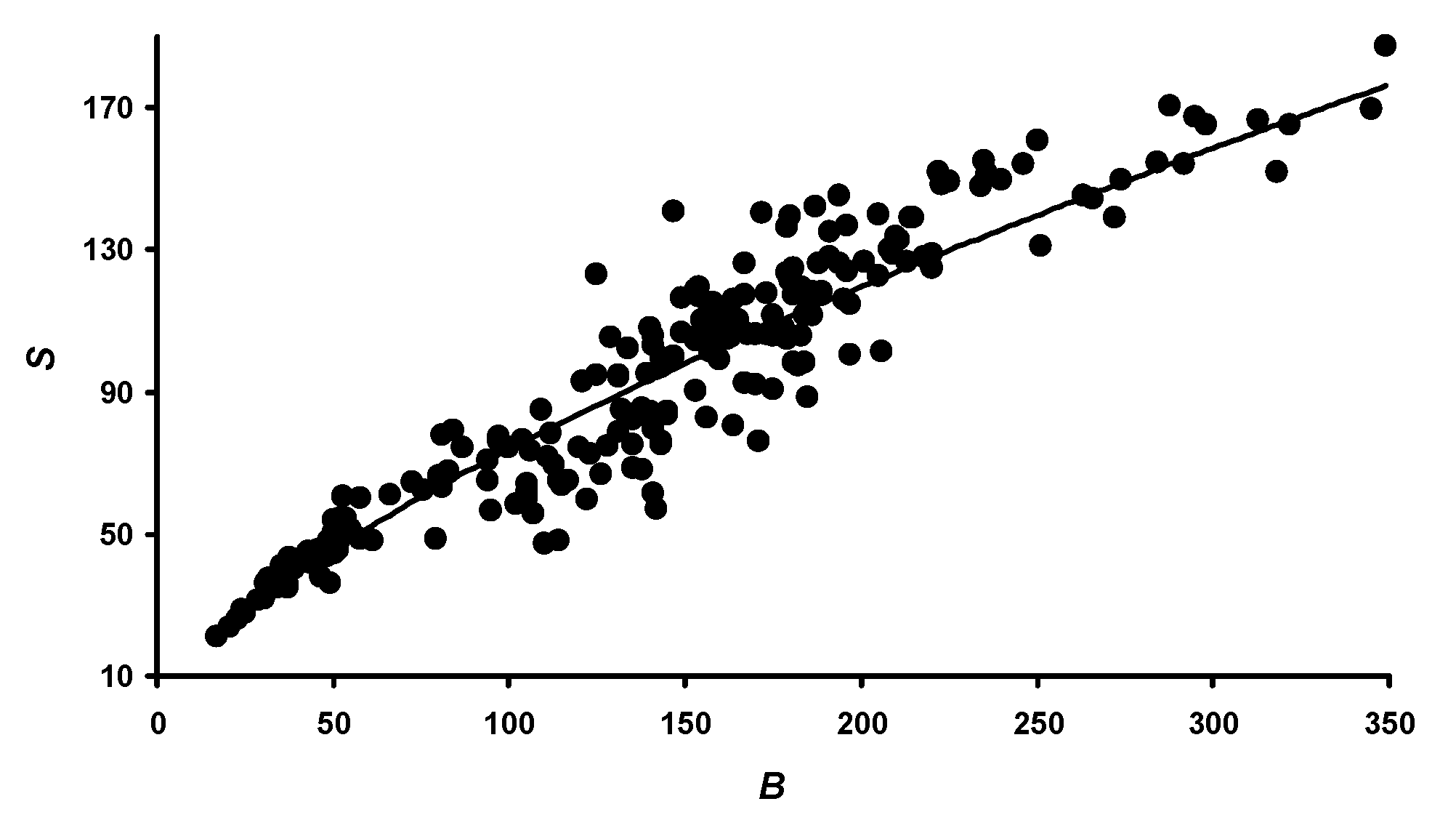
3.1. Changes in B, V, and S
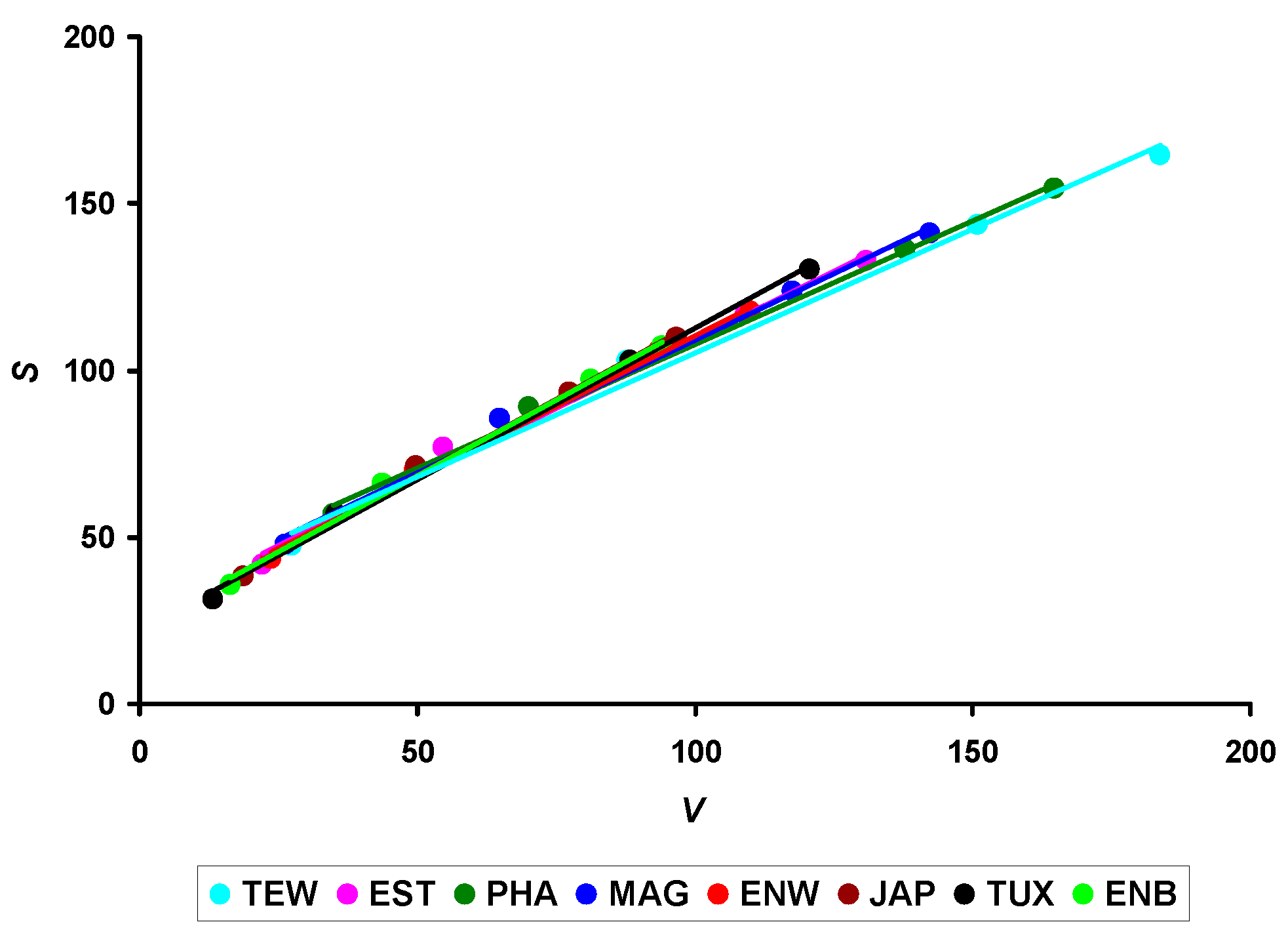
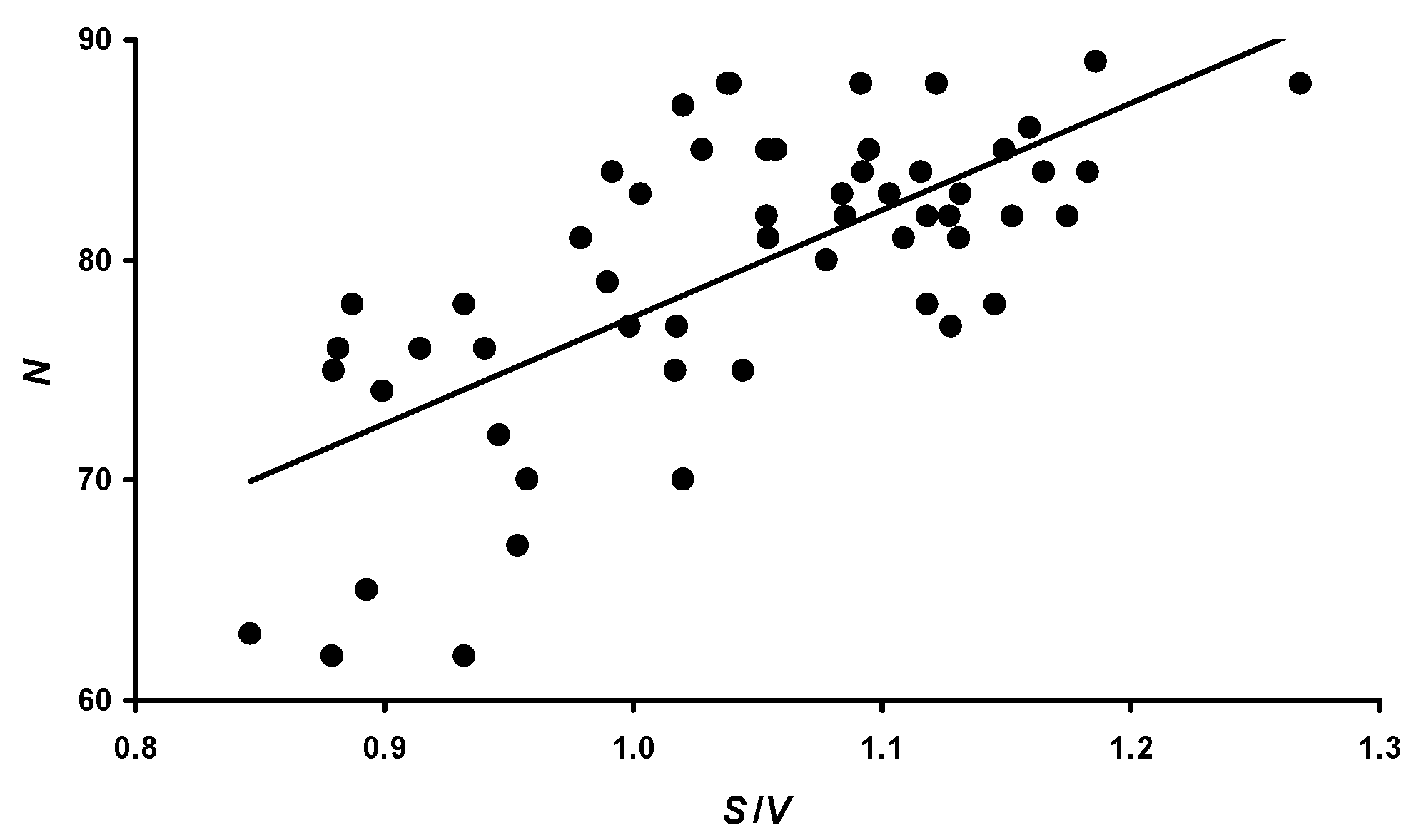
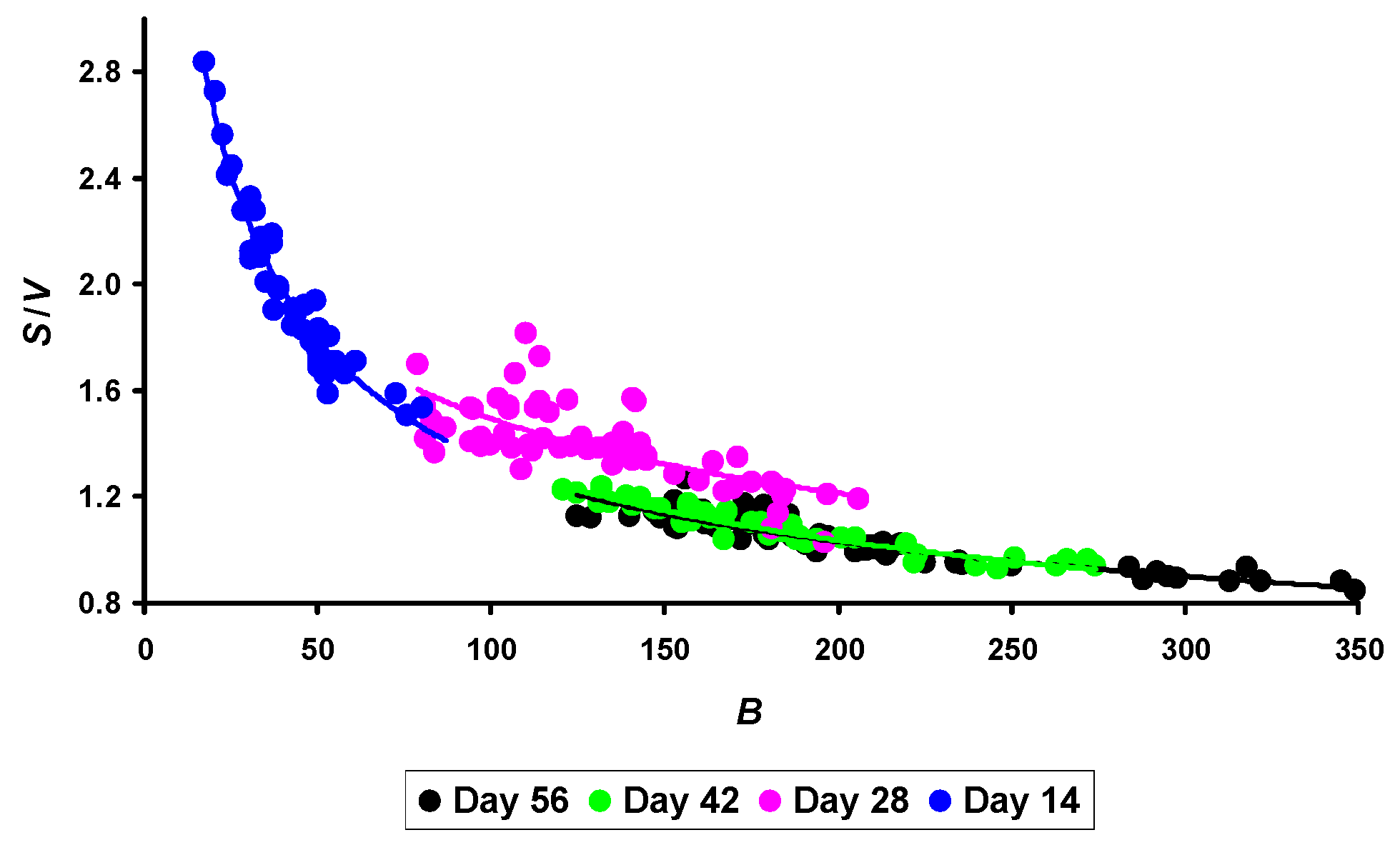
3.2. The S/V Ratio and Metabolism Level
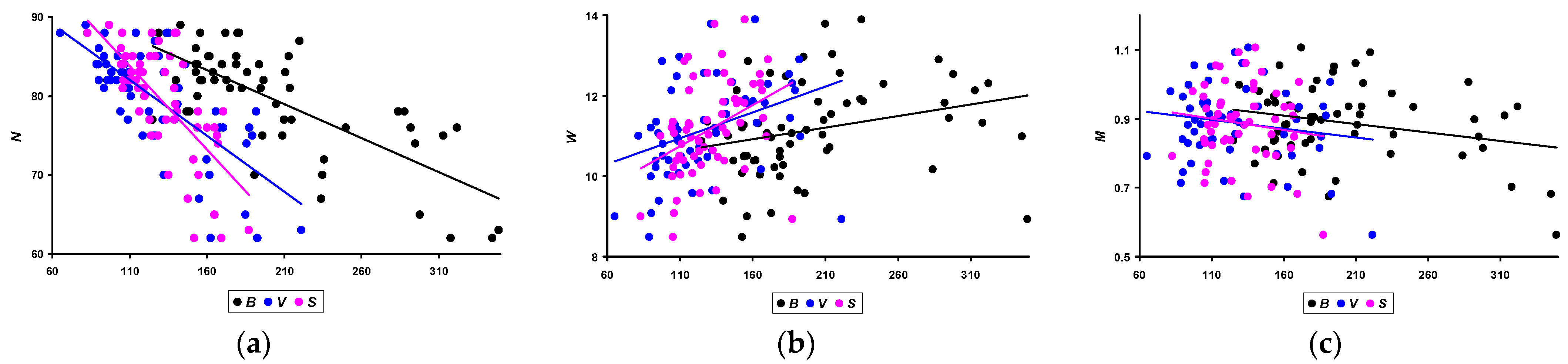
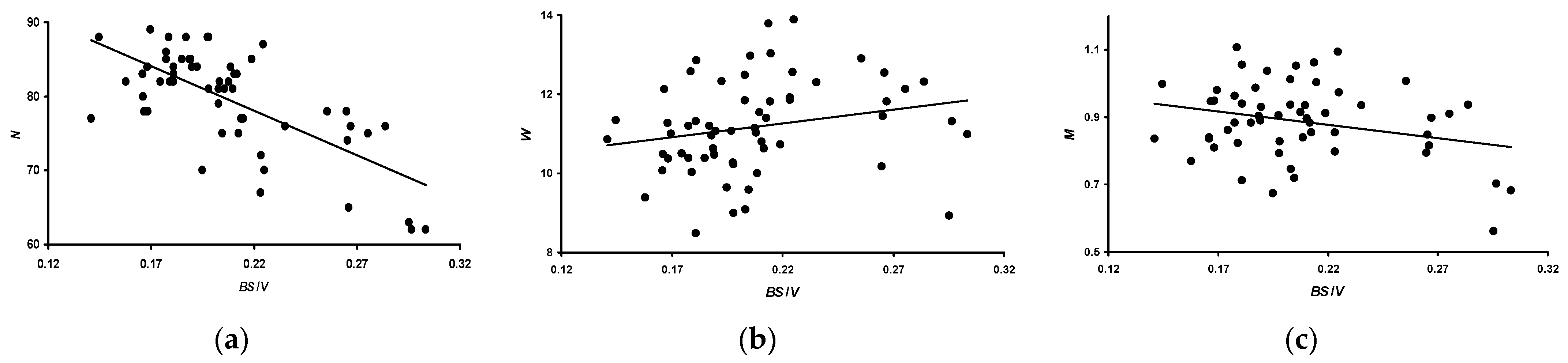
3.3. Correlation between Performance Traits and Morphometric Characteristics
- N: −0.730 (p < 0.05) for B; −0.724 (p < 0.05) for V; and −0.708 (p < 0.05) for S.
- W: 0.271 (p < 0.05) for B; 0.370 (p < 0.05) for V; and 0.384 (p < 0.05) for S.
- M: −0.254 (p < 0.05) for B; −0.160 (insignificant) for V; and −0.135 (insignificant) for S.
3.4. Inferring the Metabolic Index B·S/V
3.5. Correlation between the Metabolic Index B·S/V and Performance Traits
- N: −0.690 (p < 0.05).
- W: 0.219 (insignificant).
- M: −0.275 (p < 0.05).
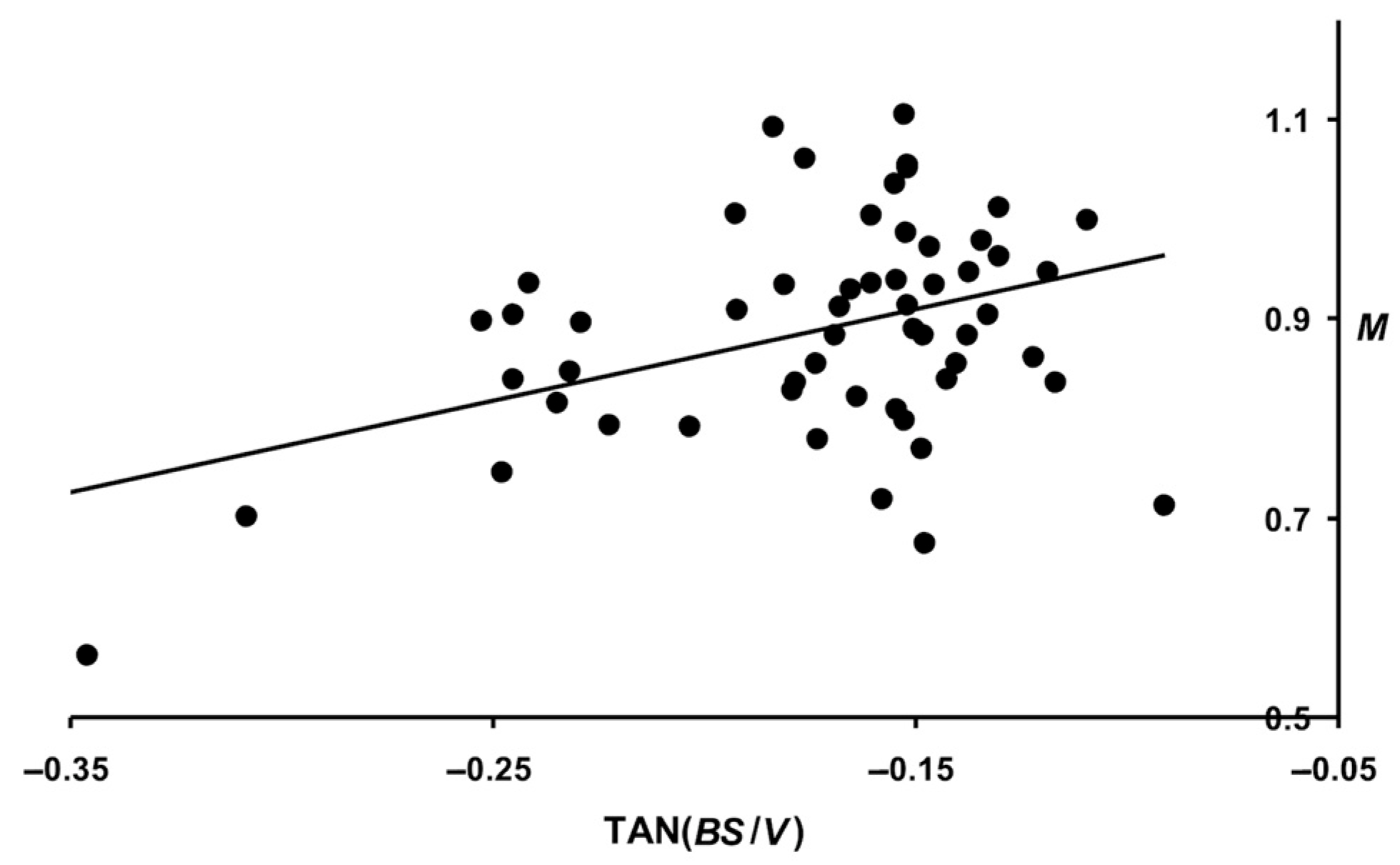
3.6. Examining Slope Angles for B, V, and S Changes
- W: 0.399 (p < 0.05) for TANB; 0.495 (p < 0.05) for TANV; and 0.455 (p < 0.05) for TANS.
- M: 0.388 (p < 0.05) for TANB; 0.374 (p < 0.05) for TANV; and 0.368 (p < 0.05) for TANS.
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nager, R.G. The challenges of making eggs. Ardea 2006, 94, 323–346. [Google Scholar]
- Rahn, H.; Paganelli, C.V.; Ar, A. Relation of avian egg weight to body weight. Auk 1975, 92, 750–765. [Google Scholar] [CrossRef]
- Vézina, F.; Williams, T.D. Plasticity in body composition in breeding birds: What drives the metabolic costs of egg production? Physiol. Biochem. Zool. 2003, 76, 716–730. [Google Scholar] [CrossRef] [PubMed]
- Vézina, F.; Williams, T.D. The metabolic cost of egg production is repeatable. J. Exp. Biol. 2005, 208, 2533–2538. [Google Scholar] [CrossRef] [PubMed]
- Sockman, K.W.; Sharp, P.J.; Schwabl, H. Orchestration of avian reproductive effort: An integration of the ultimate and proximate bases for flexibility in clutch size, incubation behaviour, and yolk androgen deposition. Biol. Rev. 2006, 81, 629–666. [Google Scholar] [CrossRef] [PubMed]
- Fathi, M.M.; Al-Homidan, I.; Ebeid, T.A.; Galal, A.; Abou-Emera, O.K. Assessment of residual feed intake and its relevant measurements in two varieties of japanese quails (Coturnixcoturnix japonica) under high environmental temperature. Animals 2019, 9, 299. [Google Scholar] [CrossRef] [PubMed]
- Rahn, H.; Sotherland, P.R.; Paganelli, C.V. Interrelationships between egg mass and adult body mass and metabolism among passerine birds. J. Ornithol. 1985, 126, 263–271. [Google Scholar] [CrossRef]
- Shingleton, A.W. Allometry: The study of biological scaling. Nat. Sci. Educ. 2010, 3, 2. Available online: https://www.nature.com/scitable/knowledge/library/allometry-the-study-of-biological-scaling-13228439/ (accessed on 1 November 2023).
- Glazier, D.S. Effects of metabolic level on the body size scaling of metabolic rate in birds and mammals. Proc. R. Soc. B 2008, 275, 1405–1410. [Google Scholar] [CrossRef]
- Price, C.A.; Weitz, J.S.; Savage, V.M.; Stegen, J.; Clarke, A.; Coomes, D.A.; Dodds, P.S.; Etienne, R.S.; Kerkhoff, A.J.; McCulloh, K.; et al. Testing the metabolic theory of ecology. Ecol. Lett. 2012, 15, 1465–1474. [Google Scholar] [CrossRef]
- Giancarli, S.M.; Dunham, A.E.; O’Connor, M.P. Clade-specific allometries in avian basal metabolic rate demand a broader theory of allometry. Physiol. Biochem. Zool. 2023, 96, 216–232. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. A unifying explanation for diverse metabolic scaling in animals and plants. Biol. Rev. 2010, 85, 111–138. [Google Scholar] [CrossRef] [PubMed]
- James, W.P.T.; Ferro-Luzzi, A. Metabolic rate. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Trugo, L.C., Finglas, P.M., Eds.; Academic Press: Cambridge, MA, USA; Elsevier Science Ltd.: Amsterdam, The Netherlands; New York, NY, USA, 2003; pp. 3863–3868. [Google Scholar] [CrossRef]
- Sahni, R.; Schulze, K. 58—Temperature control in newborn infants. In Fetal and Neonatal Physiology, 4th ed.; Polin, R.A., Fox, W.W., Abman, S.H., Eds.; Saunders, Elsevier Inc.: Philadelphia, PA, USA, 2011; Volume 1, pp. 624–648. [Google Scholar] [CrossRef]
- Lee, J.K.; Kung, H.H.; Mockros, L.F. Microchannel technologies for artificial lungs: (1) Theory. ASAIO J. 2008, 54, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Planinšič, G.; Vollmer, M. The surface-to-volume ratio in thermal physics: From cheese cube physics to animal metabolism. Eur. J. Phys. 2008, 29, 369–384. [Google Scholar] [CrossRef]
- Mitchell, H.H. The surface area of Single Comb White Leghorn chickens. J. Nutr. 1930, 2, 443–449. [Google Scholar] [CrossRef]
- Perez, C.R.; Moye, J.K.; Pritsos, C.A. Estimating the surface area of birds: Using the homing pigeon (Columba livia) as a model. Biol. Open 2014, 3, 486–488. [Google Scholar] [CrossRef] [PubMed]
- Volkovoy, S.; Bondarenko, Y. [Japanese quail plumage rainbow]. Allot. Husb. 1989, 5, 14–15. Available online: https://yablonka.net/world/zh/686-raduga-opereniya-yaponskogo-perepela.html (accessed on 1 November 2023). (In Russian).
- Ryabokon, Y.O.; Pabat, V.O.; Mykytyuk, D.M.; Frolov, V.V.; Katerynych, O.O.; Bondarenko, Y.V.; Mosyakina, T.V.; Gadyuchko, O.T.; Kovalenko, G.T.; Gritsenko, D.M.; et al. (Eds.) Catalog of Poultry Breeding Resources of Ukraine; Poultry Research Institute: Kharkiv, Ukraine, 2005; Available online: http://avianua.com/archiv/plevreestr/per.pdf (accessed on 1 November 2023). (In Ukrainian)
- Genofond. Catalogue of Breeds: Quails; Official Site of the Company Genofond LLC.: Sergiev Posad, Russia, 2015; Available online: http://www.genofond-sp.ru/quail.html (accessed on 1 November 2023). (In Russian)
- Roiter, Y.S.; Degtyareva, T.N.; Degtyareva, O.N.; Anshakov, D.V. Gene pool of quail breeds: Present condition and prospects for practical application. Ptitsevodstvo 2017, 6, 7–11. Available online: https://www.elibrary.ru/item.asp?id=29444424 (accessed on 1 November 2023). (In Russian with English Summary).
- Domesticfutures. Quail Breeds: Characteristics with Photos. domesticfutures.com. 2021. Available online: https://domesticfutures.com/porody-perepelov-harakteristiki-s-fotografiyami-4457 (accessed on 1 November 2023).
- German, N.Y.; Vetokh, A.N.; Dzhagaev, A.Y.; Ilyina, E.R.; Kotova, T.O. Morphometric parameters of eggs from breeds quail for meat. Vet. Kormlenie 2023, 2, 20–23, (In Russian with English Summary). [Google Scholar] [CrossRef]
- Bachinina, K.N.; Shcherbatov, V.I. Morphological indicators and quality of quail eggs of different breeds. Ptitsevodstvo 2021, 6, 69–72, (In Russian with English Summary). [Google Scholar] [CrossRef]
- Dymkov, A.B.; Fisinin, V.I. Differentiation of quail (Coturnix japonica) breeds based on the morphological parameters of eggs. Agric. Biol. 2022, 57, 694–705. [Google Scholar] [CrossRef]
- Romanov, M.N.; Wezyk, S.; Cywa-Benko, K.; Sakhatsky, N.I. Poultry genetic resources in the countries of Eastern Europe—History and current state. Poult. Avian Biol. Rev. 1996, 7, 1–29. Available online: https://www.researchgate.net/publication/255710929_Poultry_genetic_resources_in_the_countries_of_Eastern_Europe_-_history_and_current_state (accessed on 1 November 2023).
- Prituzhalova, A.O.; Volkova, N.A.; Kuzmina, T.I.; Vetokh, A.N.; Dzhagaev, A.Y. Monitoring of indicators of chromatin status in quails ovarian follicles granulosa cells of different directions of productivity. Agrar. Nauka 2023, 368, 53–57, (In Russian with English Summary). [Google Scholar] [CrossRef]
- Mills, A.D.; Crawford, L.L.; Domjan, M.; Faure, J.M. The behavior of the Japanese or domestic quail Coturnix japonica. Neurosci. Biobehav. Rev. 1997, 21, 261–281. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.B.; Chang, H.; Liu, X.P.; Xu, W.; Wang, H.Y.; Zhao, W.M.; Olowofeso, O. Developmental research on the origin and phylogeny of quails. Worlds Poult. Sci. J. 2005, 61, 105–112. [Google Scholar] [CrossRef]
- Genchev, A. Egg production potential of Manchurian Golden quail breeders. Agric. Sci. Technol. 2011, 3, 73–80. Available online: http://agriscitech.eu/wp-content/uploads/2014/05/GB_02.pdf (accessed on 1 November 2023).
- Purely Poultry. Gold Coturnix Quail Set. Purely Poultry, Durand, WI, USA. 2023. Available online: https://www.purelypoultry.com/index.php?main_page=product_info&products_id=1267 (accessed on 1 November 2023).
- Narushin, V.G.; Lu, G.; Cugley, J.; Romanov, M.N.; Griffin, D.K. A 2-D imaging-assisted geometrical transformation method for non-destructive evaluation of the volume and surface area of avian eggs. Food Control 2020, 112, 107112. [Google Scholar] [CrossRef]
- Eichenwald, A.J.; Reed, M. Biased assessment of thermal properties of birds from estimated body density. J. Therm. Biol. 2023, 112, 103472. [Google Scholar] [CrossRef]
- Vieira Filho, J.A.; Garcia, E.A.; de Molino, A.B.; dos Santos, T.A.; de Almeida Paz, I.C.L.; de Baldo, G.A.A. Productivity of japanese quails in relation to body weight at the end of the rearing phase. Acta Sci. Anim. Sci. 2016, 38, 213–217. [Google Scholar] [CrossRef]
- Lukanov, H.; Genchev, A.; Kolev, P. Comparative investigation of egg production in WG, GG and GL Japanese quail populations. Trakia J. Sci. 2018, 4, 334–343. [Google Scholar] [CrossRef]
- Kleiber, M. Body size and metabolism. Hilgardia 1932, 6, 315–353. [Google Scholar] [CrossRef]
- Schmidt-Nielsen, K. Scaling: Why Is Animal Size So Important? Cambridge University Press: Cambridge, UK, 1984. [Google Scholar]
- Glazier, D.S. Beyond the ‘3/4-power law’: Variation in the intra- and interspecific scaling of metabolic rate in animals. Biol. Rev. Camb. Philos. Soc. 2005, 80, 611–662. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. The 3/4-power law is not universal: Evolution of isometric, ontogenetic metabolic scaling in pelagic animals. BioScience 2006, 56, 325–332. [Google Scholar] [CrossRef]
- Hamershock, D.M.; Seamans, T.W.; Bernhardt, G.E. Determination of body density for twelve bird species. IBIS 1995, 137, 424–428. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | Code | n 1 | Origin | Performance 2 | Refs |
|---|---|---|---|---|---|
Pharaoh | PHA | 7 | The USA; wild type color; a French fattening line that was imported and used in this study | Meat type N = 250 eggs; W = 13.8 g; Bm = 200.2 g, Bf = 260.5 g | [19,20,21,22,23,24] |
Texas White (or Texas Pharaoh, White Pharaoh, Snowy) | TEW | 8 | Texas, USA; from a cross between PHA and ENW | Meat type N = 243 eggs; W = 13.9 g; Bm = 240.0 g, Bf = 280.0 g | [22,23,24,25,26] |
Estonian | EST | 9 | Estonia, 1988; from a cross between JAP (a Moscow line), ENW and Pharaoh | Dual purpose (or universal) N = 263 eggs; W = 13.2 g; Bm = 187.8 g, Bf = 239.5 g | [21,22,23,27] |
English White | ENW | 12 | England; a mutant from JAP quails; brought from Hungary to the USSR in 1987 | Egg type N = 261 eggs; W = 12.0 g; Bm = 155.2 g, Bf = 188.0 g | [19,21,22,23,28] |
English Black | ENB | 2 | England; a mutant from JAP quails; brought from Hungary to the USSR in 1971 | Egg type N = 261 eggs; W = 12.1 g; Bm = 160.0 g, Bf = 190.5 g | [21,22,23] |
Tuxedo | TUX | 7 | from a cross between ENW and ENB | Egg type N = 257 eggs; W = 12.0 g; Bm = 148.2 g, Bf = 180.5 g | [19,21,22,23] |
Japanese | JAP | 15 | Japan; domesticated in Japan and China around 12th century or earlier; selected in the first half of the 20th century, imported to the USSR from Japan in the middle of the 20th century and/or from Yugoslavia in 1964 | Egg type N = 288 eggs; W = 12.0 g; Bm = 150.0 g, Bf = 180.0 g | [19,21,22,23,27,29,30] |
Manchurian Golden | MAG | 4 | Marsh Farms, CA, USA, 1960s; selected by Albert Marsh in a flock of brown quails as a natural mutant | Egg type N = 267 eggs; W = 12.4 g; Bm = 158.2 g, Bf = 189.3 g | [19,21,22,23,31,32] |
| Parameters | Quail Breeds | |||||||
|---|---|---|---|---|---|---|---|---|
| TEW (a) | EST (b) | PHA (c) | MAG (d) | ENW (e) | JAP (f) | TUX (g) | ENB (h) | |
| Body measurements | ||||||||
| Body weight in g (B) | ||||||||
| B1 | 9.6 ± 0.2 e,g | 9.1 ± 1.1 | 9.4 ± 0.3 e | 9.2 ± 0.6 | 8.3 ± 0.5 a,c,d | 9.0 ± 1.6 | 8.8 ± 0.7 a | 9.1 ± 0.7 |
| B7 | 18.0 ± 5.1 e,f,g | 17.0 ± 2.3 e,f,g,h | 15.6 ± 6.8 f,g,h | 13.6 ± 4.2 f,h | 11.1 ± 4.3 a,b,f,h | 6.0 ± 1.3 a,b,c,d,e,g,h | 8.4 ± 1.5 a,b,c,d,e,f | 7.6 ± 0.4 a,b |
| B14 | 61.4 ± 15.5 b,e,g,h | 42.2 ± 8.3 a,c,f,g | 62.2 ± 16.7 b,e,g,h | 47.8 ± 10.5 g | 44.4 ± 12.2 a,c,g | 51.7 ± 8.3 b,g,h | 27.6 ± 5.2 a,b,c,d,e,f,h | 37.0 ± 0.1 a,f,g |
| B21 | 102.4 ± 6.1 b,e,f,g,h | 78.0 ± 14.1 a,c,f,g,h | 102.0 ± 22.2 b,f,g,h | 86.8 ± 15.3 f,g,h | 86.5 ± 14.5 a,f,g,h | 60.5 ± 3.5 a,b,c,d,e,g | 54.4 ± 6.4 a,b,c,d,e,h | 65.5 ± 3.5 a,e |
| B28 | 183.3 ± 9.1 b,d,e,f,g,h | 134.1 ± 13.4 a,c,f,g,h | 165.4 ± 33.1 e,f,h | 146.0 ± 22.5 a,f,h | 119.3 ± 17.9 a,c,f,g | 94.6 ± 10.1 a,b,c,d,e,g | 124.9 ± 15.2 a,b,c,f,h | 94.0 ± 15.6 a,g |
| B35 | 216.4 ± 8.0 b,d,e,f,g,h | 159.6 ± 10.8 a,c,e,f,g,h | 196.6 ± 28.9 b,e,f,g,h | 161.5 ± 39.3 a | 141.5 ± 18.6 a,b,c,f | 120.5 ± 13.4 a,b,c,e,h | 135.5 ± 15.4 a,b,c | 131.0 ± 0 a,b,c |
| B42 | 261.8 ± 12.6 b,c,d,e,f,g,h | 175.1 ± 13.9 a,c,d,f,g,h | 221.1 ± 27.4 a,b,d,e,f,g,h | 191.7 ± 8.1 a,e,f,g,h | 159.6 ± 22.0 a,c,d,f,h | 133.0 ± 1.4 a,b,c,d,e,g | 146.1 ± 16.2 a,b,c,d | 137.0 ± 8.5 a,b,c,d |
| B49 | 277.0 ± 18.4 b,c,d,e,f,g,h | 193.7 ± 15.7 a,c,e,f,g,h | 243.6 ± 32.0 a,b,d,e,f,g,h | 198.3 ± 7.6 a,c,e,f,g,h | 167.7 ± 16.1 a,b,c,d,f,h | 135.5 ± 3.5 a,b,c,d,e,g | 164.0 ± 13.6 a,b,c,d,f | 144.0 ± 9.9 a,b,c,d |
| B56 | 312.9 ± 24.7 b,c,d,e,f,g,h | 205.4 ± 13.5 a,c,e,f,g,h | 253.4 ± 34.2 a,b,d,e,f,g,h | 207.7 ± 11.8 a,c,e,f,g,h | 181.0 ± 21.1 a,b,c,d,f,h | 160.1 ± 16.8 a,b,c,d,e | 164.1 ± 21.9 a,b,c,d | 154.5 ± 2.1 a,b,c,d |
| Body length in cm (l) | ||||||||
| l14 | 6.0 ± 0.8 c | 5.8 ± 0.6 c | 7.0 ± 0.6 a,b,e,f,g,h | 6.7 ± 0.8 | 6.1 ± 0.7 c | 6.0 ± 0.4 c | 5.8 ± 0.7 c | 6.0 ± 0.4 |
| l28 | 8.9 ± 0.8 e,f,g,h | 8.1 ± 0.7 c | 8.2 ± 0.4 e,f,g,h | 8.4 ± 0.9 g | 7.2 ± 0.7 a,b,c,d | 7.5 ± 0.6 a,c,g | 6.7 ± 0.4 a,b,c,d,e,f,h | 7.4 ± 0.1 a,c |
| l42 | 9.5 ± 0.8 e,f,g,h | 8.9 ± 0.4 d,e,f | 9.5 ± 0.8 e,f,g,h | 9.3 ± 0.3 e,f | 8.5 ± 0.4 a,b,c,d | 8.1 ± 1.1 a,b,c,d,e | 8.7 ± 0.6 a,c,d | 8.4 ± 0.5 |
| l56 | 10.3 ± 0.5 b,d,e,f,h | 9.6 ± 0.4 a,e,g,h | 10.3 ± 0.6 b,d,e,f,h | 10.0 ± 0.5 h | 8.9 ± 0.4 a,b,c,g | 9.1 ± 1.2 a,c,g | 10.5 ± 0.6 a,b,d,e,f,h | 8.8 ± 0.0 a,b,c,g |
| Body circumference in cm (c) | ||||||||
| c14 | 9.2 ± 0.6 b,f,g,h | 8.4 ± 0.6 a,c,f,g,h | 9.6 ± 1.1 b,f,g,h | 8.5 ± 0.9 g | 8.4 ± 1.2 c,g,h | 7.7 ± 0.2 a,b,c,e,g,h | 6.5 ± 0.5 a,b,c,d,e,f,h | 7.2 ± 0.1 b,c |
| c28 | 13.5 ± 1.3 b,e,f,g,h | 11.3 ± 0.7 a,c,g,h | 12.6 ± 1.0 b,e,f,g,h | 12.0 ± 1.2 g | 11.3 ± 0.8 a,c,g,h | 11.1 ± 0.6 a,c,g,h | 9.9 ± 0.8 a,b,c,d,e,f | 10.6 ± 0.2 a,c |
| c42 | 17.3 ± 0.2 b,d,e,f,g,h | 15.2 ± 0.6 a,c,f,g,h | 16.5 ± 1.0 b,e,f,g,h | 15.4 ± 0.2 a,e,f,g,h | 14.4 ± 0.8 a,b,c,d,f,g,h | 13.5 ± 0.2 a,b,c,d,e,g | 13.8 ± 0.4 a,b,c,d,f | 13.6 ± 0.2 a,b,c,d |
| c56 | 18.3 ± 0.6 b,d,e,f,g,h | 16.0 ± 0.7 a,c,e,f,g,h | 17.3 ± 1.0 b,d,e,f,g,h | 16.4 ± 0.3 a,e,f,g,h | 15.2 ± 0.7 a,b,c,d,f,h | 14.2 ± 0.6 a,b,c,d,e | 14.7 ± 0.6 a,b,c,d | 14.2 ± 0.4 a,b,c,d |
| Body volume in cm3 (V) | ||||||||
| V14 | 27.3 ± 7.1 f,g,h | 22.1 ± 4.8 c,g,h | 34.9 ± 9.1 b,f,g,h | 26.3 ± 8.3 g | 23.6 ± 7.6 c,g,h | 18.7 ± 2.4 a,c,e,g,h | 13.3 ± 3.2 a,b,c,d,e,f,h | 16.3 ± 0.3 a,c |
| V28 | 87.9 ± 23.7 b,e,f,g,h | 54.9 ± 8.8 c,g,h | 70.1 ± 11.6 b,e,f,g,h | 65.0 ± 16.5 g | 49.5 ± 9.8 a,c,g | 49.7 ± 8.1 a,c,g,h | 35.5 ± 7.2 a,b,c,d,e,f,h | 43.7 ± 2.6 a,c |
| V42 | 151.9 ± 13.8 b,d,e,f,g,h | 111.2 ± 14.6 a,c,d,e,f,g,h | 140.0 ± 26.4 b,e,f,g,h | 117.6 ± 6.3 a,e,f,g,h | 93.6 ± 11.7 a,b,c,d,f,h | 77.4± 12.6 a,b,c,d,e,g | 88.3 ± 9.6 a,b,c,d | 81.3 ± 2.3 a,b,c,d |
| V56 | 183.7 ± 19.0 b,d,e,f,g,h | 128.5 ± 10.2 a,c,e,f,h | 161.3 ± 20.5 b,d,e,f,g,h | 142.5 ± 3.5 a,e,f,g,h | 110.1 ± 9.9 a,b,c,d,f,h | 96.9 ± 14.1 a,b,c,d,e,g | 120.7 ± 16.3 a,c,f,h | 94.2 ± 5.6 a,b,c,d |
| Body surface area in cm2 (S) | ||||||||
| S14 | 47.2 ± 8.8 f,g,h | 41.5 ± 6.4 c,g,h | 56.7 ± 10.1 b,f,g,h | 47.7 ± 10.6 g | 43.5 ± 10.0 c,g,h | 38.3 ± 3.6 a,c,g,h | 31.2 ± 5.4 a,b,c,d,e,f | 35.5 ± 1.2 c |
| S28 | 102.9 ± 18.1 b,e,f,g,h | 76.9 ± 8.4 c,g,h | 88.8 ± 9.1 b,e,f,g,h | 85.3 ± 15.0 g | 70.0 ± 9.8 a,c,g | 71.1 ± 8.1 a,c,g | 56.8 ± 7.3 a,b,c,d,e,f,h | 65.9 ± 2.6 a,c |
| S42 | 144.4 ± 10.3 b,d,e,f,g,h | 118.9 ± 11.0 a,c,d,e,f,g,h | 137.9± 18.3 b,e,f,g,h | 123.7 ± 4.8 a,e,f,g,h | 105.7 ± 8.2 a,b,c,d,f,h | 93.5 ± 12.1 a,b,c,d,e,g | 102.9 ± 8.3 a,b,c,d | 97.1 ± 3.2 a,b,c,d |
| S56 | 164.4 ± 11.5 b,d,e,f,g,h | 130.7 ± 6.0 a,c,e,f,h | 151.5 ± 12.6 b,d,e,f,g,h | 141.1 ± 3.4 a,e,f,g,h | 117.7 ± 6.7 a,b,c,d,h | 109.9 ± 13.0 a,b,c,d,g | 130.1 ± 12.2 a,c,e,f,h | 107.2 ± 3.7 a,b,c,d,g |
| Metabolic level (S/V) | ||||||||
| S14/V14 | 1.76 ± 0.13 f,g,h | 1.91 ± 0.14 a,c,f,g,h | 1.67 ± 0.21 b,f,g,h | 1.87 ± 0.22 g | 1.94 ± 0.35 g,h | 2.06 ± 0.07 a,b,c,g,h | 2.39 ± 0.19 a,b,c,d,e,f,h | 2.17 ± 0.03 a,b,f |
| S28/V28 | 1.19 ± 0.11 b,e,f,g,h | 1.41 ± 0.09 a,c,f,g,h | 1.28 ± 0.09 b,e,f,g,h | 1.34 ± 0.13 g | 1.43 ± 0.11 a,c,g | 1.44 ± 0.08 a,c,g,h | 1.62 ± 0.12 a,b,c,d,e,f | 1.51 ± 0.03 a,c |
| S42/V42 | 0.95 ± 0.02 b,d,e,f,g,h | 1.07 ± 0.04 a,c,e,f,g,h | 0.99 ± 0.06 b,e,f,g,h | 1.05 ± 0.02 a,e,f,g,h | 1.14 ± 0.05 a,b,c,d,f,h | 1.21 ± 0.04 a,b,c,d,e,g | 1.17 ± 0.03 a,b,c,d,f | 1.19 ± 0.01 a,b,c,d |
| S56/V56 | 0.90 ± 0.03 b,c,d,e,f,g,h | 1.02 ± 0.04 a,c,e,f,g,h | 0.94 ± 0.05 b,d,e,f,g,h | 0.99 ± 0.01 a,e,f,g,h | 1.07 ± 0.04 a,b,c,d,f,h | 1.14 ± 0.05 a,b,c,d,e,g | 1.08 ± 0.05 a,b,c,d,f | 1.14 ± 0.03 a,b,c,d |
| Quail productivity for 3 months: egg production (N); average egg weight in g (W); and total egg mass in kg (M) | ||||||||
| N | 69.5 ± 7.1 b,d,e,f,g,h | 78.9 ± 5.6 a,c,f | 73.0 ± 4.1 b,d,e,f,g,h | 80.7 ± 3.5 a,c | 81.7 ± 3.8 a,c | 83.9 ± 3.2 a,b,c | 83.0 ± 4.8 a,c | 84.0 ± 2.8 a,c |
| W | 11.2 ± 1.2 | 11.5 ± 1.4 f | 12.5 ± 0.8 e,f,h | 12.3 ± 0.6 e,f,h | 11.0 ± 0.8 c,d,f | 10.1 ± 0.8 a,b,c,d,e,g | 11.6 ± 0.9 f | 10.9 ± 0.5 |
| M | 0.78 ± 0.12 c,d,e,g | 0.91 ± 0.16 | 0.91 ± 0.08 a | 0.99 ± 0.05 a,f | 0.90 ± 0.07 a,d | 0.85 ± 0.08 d,g | 0.97 ± 0.10 a,f | 0.91 ± 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Narushin, V.G.; Volkova, N.A.; Vetokh, A.N.; Dzhagaev, A.Y.; Volkova, L.A.; Griffin, D.K.; Romanov, M.N.; Zinovieva, N.A. Metabolic Rate and Egg Production in Japanese Quails Can Be Predicted by Assessing Growth Parameters of Laying Hens. Animals 2024, 14, 258. https://doi.org/10.3390/ani14020258
Narushin VG, Volkova NA, Vetokh AN, Dzhagaev AY, Volkova LA, Griffin DK, Romanov MN, Zinovieva NA. Metabolic Rate and Egg Production in Japanese Quails Can Be Predicted by Assessing Growth Parameters of Laying Hens. Animals. 2024; 14(2):258. https://doi.org/10.3390/ani14020258
Chicago/Turabian StyleNarushin, Valeriy G., Natalia A. Volkova, Anastasia N. Vetokh, Alan Yu. Dzhagaev, Ludmila A. Volkova, Darren K. Griffin, Michael N. Romanov, and Natalia A. Zinovieva. 2024. "Metabolic Rate and Egg Production in Japanese Quails Can Be Predicted by Assessing Growth Parameters of Laying Hens" Animals 14, no. 2: 258. https://doi.org/10.3390/ani14020258