
Analysis of Plasma-Derived Exosomal MicroRNAs as Potential Biomarkers for Canine Idiopathic Epilepsy
 , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Recruitment, Sample Collection, and Exosome Characterization
2.1.1. Animals and Procedures
2.1.2. Exosome Purification from Plasma
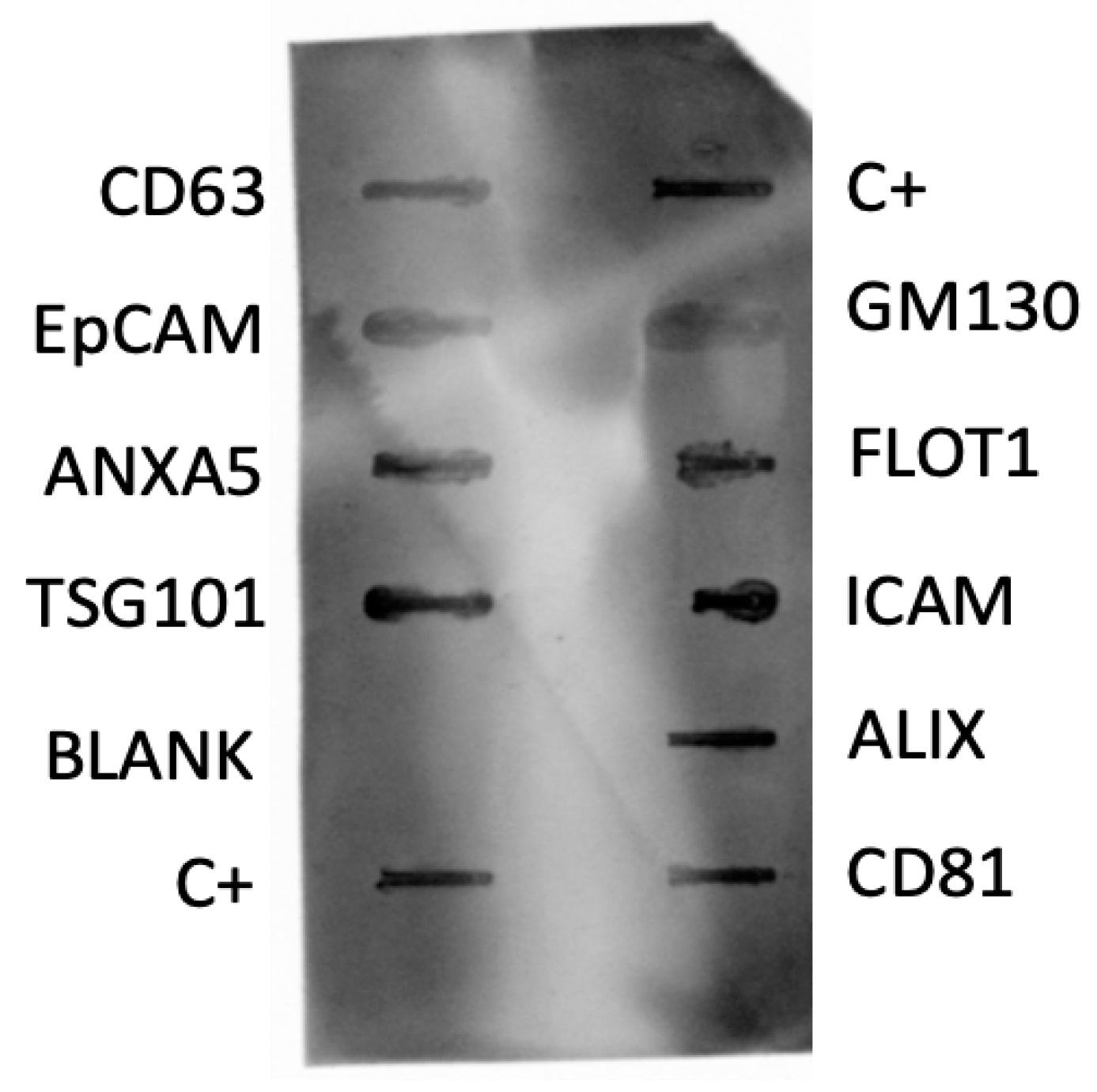
2.1.3. Determination of Exosome Markers by Dot Blotting
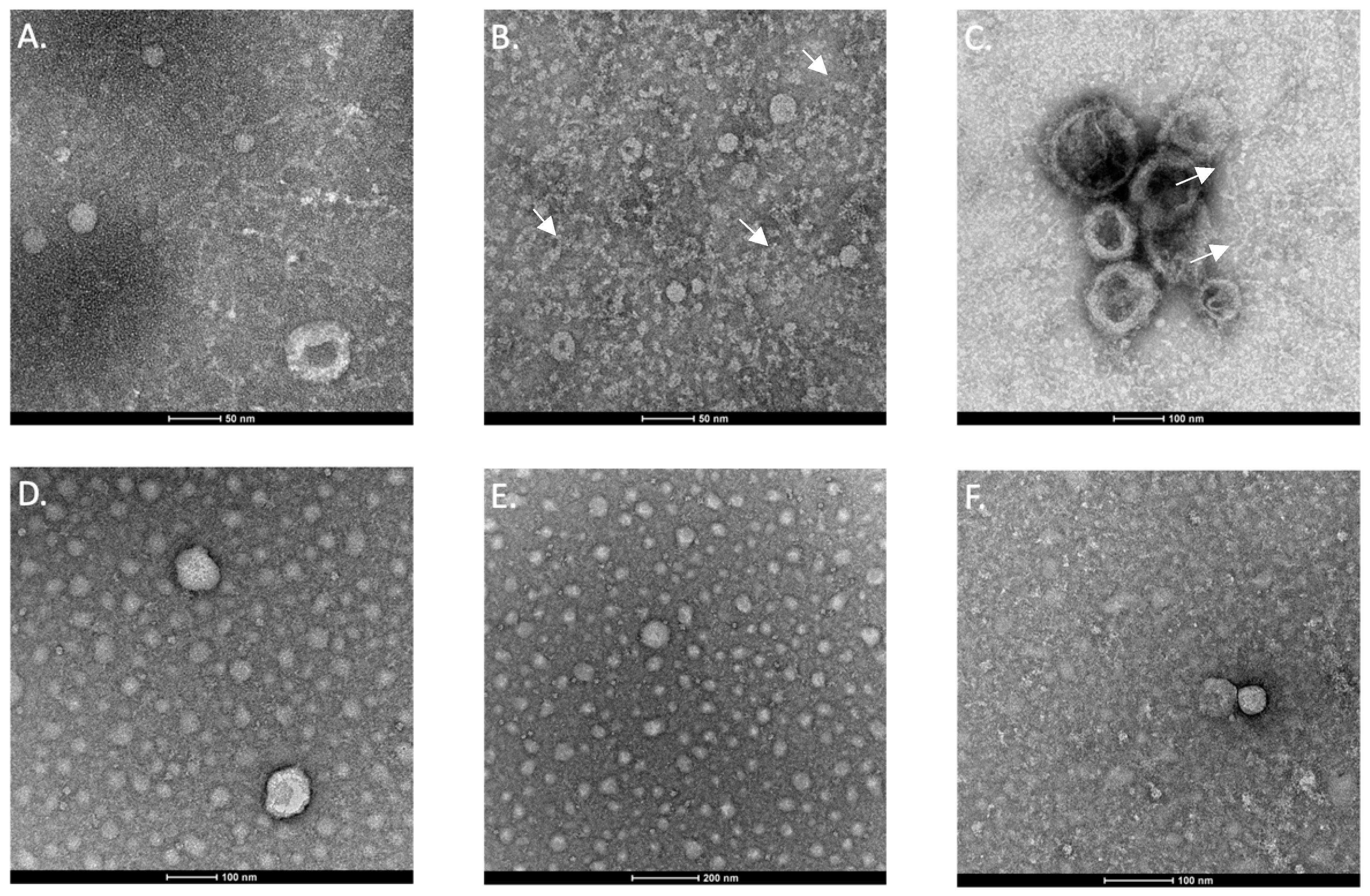
2.1.4. Imaging of Exosomes Using Transmission Electron Microscopy
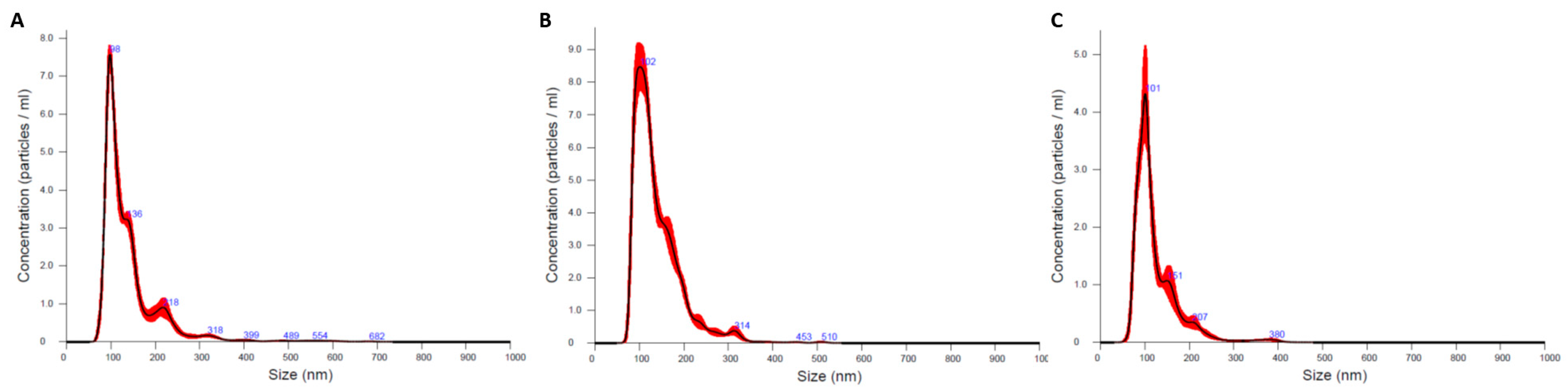
2.1.5. Analysis of Exosome Size and Concentration Using Nano Tracking Analysis
2.2. Exosomal miRNA Analysis
2.2.1. RNA Extraction
2.2.2. Reverse Transcription and Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
2.3. Statistical Analysis
3. Results
3.1. Exosome Characterization
3.1.1. Determination of Exosomal Markers by Dot Blotting
3.1.2. Imaging Exosomes Using Transmission Electron Microscopy (TEM)
3.1.3. Analysis of Extracellular Vesicle Size and Concentration Using Nano Tracking Analysis (NTA)
3.2. The miRNA Analysis
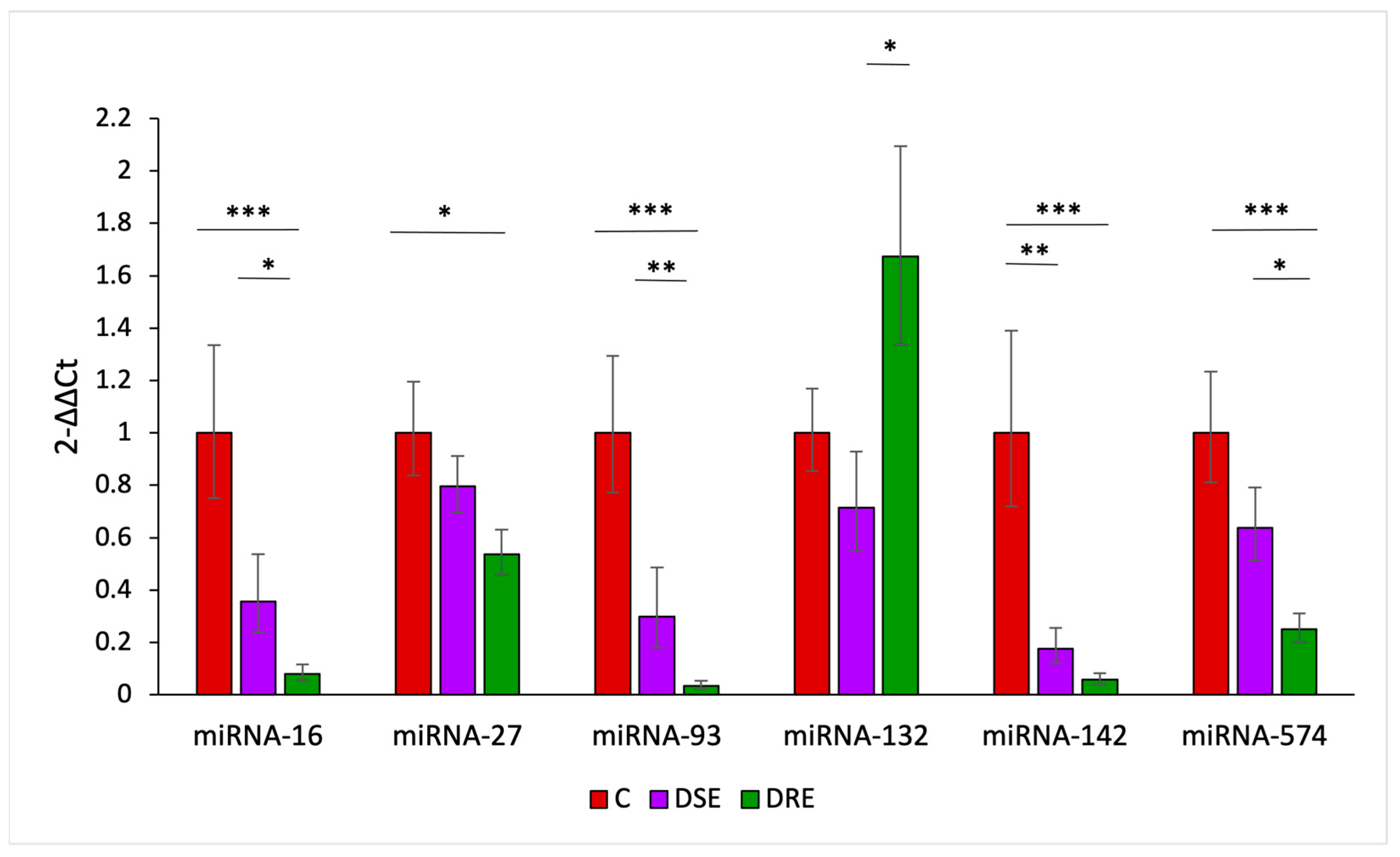
3.2.1. The miRNA Expression Analysis Using RT-qPCR
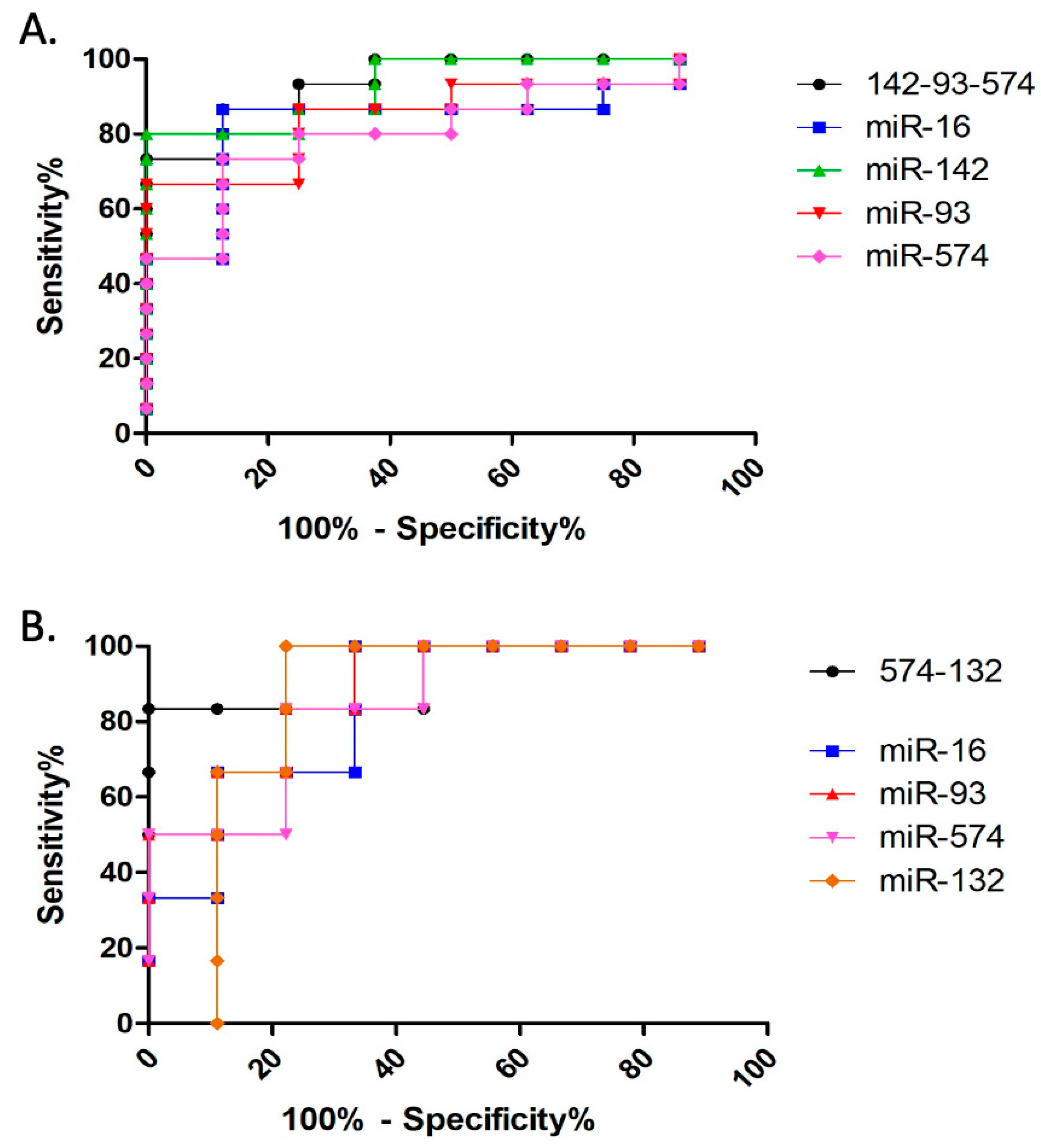
3.2.2. Discriminative Ability Analysis
3.2.3. Target Prediction and Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Löscher, W. Dogs as a natural animal model of epilepsy. Front. Vet. Sci. 2022, 9, 928009. [Google Scholar] [CrossRef] [PubMed]
- Devinsky, O.; Vezzani, A.; O’Brien, T.J.; Jette, N.; Scheffer, I.E.; de Curtis, M.; Perucca, P. Epilepsy. Nat. Rev. Dis. Primers 2018, 4, 18024. [Google Scholar] [CrossRef] [PubMed]
- Berendt, M.; Farquhar, R.G.; Mandigers, P.J.J.; Pakozdy, A.; Bhatti, S.F.M.; De Risio, L.; Fischer, A.; Long, S.; Matiasek, K.; Muñana, K.; et al. International veterinary epilepsy task force consensus report on epilepsy definition, classification and terminology in companion animals. BMC Vet. Res. 2015, 11, 182. [Google Scholar] [CrossRef] [PubMed]
- Bunford, N.; Andics, A.; Kis, A.; Miklósi, Á.; Gácsi, M. Canis familiaris as a model for non-invasive comparative neuroscience. Trends Neurosci. 2017, 40, 438–452. [Google Scholar] [CrossRef] [PubMed]
- Thompkins, A.M.; Deshpande, G.; Waggoner, P.; Katz, J.S. Functional magnetic resonance imaging of the domestic dog: Research, methodology, and conceptual issues. Comp. Cogn. Behav. Rev. 2016, 11, 63–82. [Google Scholar] [CrossRef] [PubMed]
- Nabbout, R.; Kuchenbuch, M. Impact of predictive, preventive, and precision medicine strategies in epilepsy. Nat. Rev. Neurol. 2020, 16, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Bankstahl, M.; Bankstahl, J.P. Recent advances in radiotracer imaging hold potential for future refined evaluation of epilepsy in veterinary neurology. Front. Vet. Sci. 2017, 4, 218. [Google Scholar] [CrossRef]
- Wessmann, A.; Volk, H.A.; Packer, R.M.A.; Ortega, M.; Anderson, T.J. Quality-of-life aspects in idiopathic epilepsy in dogs. Vet. Rec. 2016, 179, 229. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef]
- Kalani, A.; Tyagi, A.; Tyagi, N. Exosomes: Mediators of neurodegeneration, neuroprotection, and therapeutics. Mol. Neurobiol. 2014, 49, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Hessvik, N.P.; Øverbye, A.; Brech, A.; Torgersen, M.L.; Jakobsen, I.S.; Sandvig, K.; Llorente, A. PIKfyve inhibition increases exosome release and induces secretory autophagy. Cell. Mol. Life Sci. 2016, 73, 4717–4737. [Google Scholar] [CrossRef] [PubMed]
- Skotland, T.; Sandvig, K.; Llorente, A. Lipids in exosomes: Current knowledge and the way forward. Prog. Lipid Res. 2017, 66, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Batool, A.; Hill, T.D.M.; Nguyen, N.T.; Langa, E.; Diviney, M.; Mooney, C.; Brennan, G.P.; Connolly, N.M.; Sanz-Rodriguez, A.; Cavanagh, B.L.; et al. Altered biogenesis and MicroRNA content of hippocampal exosomes following experimental status epilepticus. Front. Neurosci. 2019, 13, 1404. [Google Scholar] [CrossRef] [PubMed]
- Mittelbrunn, M.; Gutiérrez-Vázquez, C.; Villarroya-Beltri, C.; González, S.; Sánchez-Cabo, F.; González, M.Á.; Bernad, A.; Sánchez-Madrid, F. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen- presenting cells. Nat. Commun. 2011, 2, 282. [Google Scholar] [CrossRef]
- Harrison, E.B.; Hochfelder, C.G.; Lamberty, B.G.; Meays, B.M.; Morsey, B.M.; Kelso, M.L.; Fox, H.S.; Yelamanchili, S.V. Traumatic brain injury increases levels of miR-21 in extracellular vesicles: Implications for neuroinflammation. FEBS Open Bio. 2016, 6, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Hareendran, S.; Loh, Y.P. Function of exosomes in neurological disorders and brain tumors. Extracell. Vesicles Circ. Nucleic Acids 2021, 2, 55–79. [Google Scholar] [CrossRef]
- De Risio, L.; Bhatti, S.; Muñana, K.; Penderis, J.; Stein, V.; Tipold, A.; Berendt, M.; Farqhuar, R.; Fischer, A.; Long, S.; et al. International veterinary epilepsy task force consensus proposal: Diagnostic approach to epilepsy in dogs. BMC Vet. Res. 2015, 11, 148. [Google Scholar] [CrossRef]
- Ioriatti, E.S.; Cirino, M.L.A.; Lizarte Neto, F.S.; Velasco, T.R.; Sakamoto, A.C.; Freitas-Lima, P.; Tirapelli, D.P.C.; Carlotti, C.G., Jr. Expression of circulating microRNAs as predictors of diagnosis and surgical outcome in patients with mesial temporal lobe epilepsy with hippocampal sclerosis. Epilepsy Res. 2020, 166, 106373. [Google Scholar] [CrossRef]
- Martins-Ferreira, R.; Chaves, J.; Carvalho, C.; Bettencourt, A.; Chorão, R.; Freitas, J.; Samões, R.; Boleixa, D.; Lopes, J.; Ramalheira, J.; et al. Circulating microRNAs as potential biomarkers for genetic generalized epilepsies: A three microRNA panel. Eur. J. Neurol. 2020, 27, 660–666. [Google Scholar] [CrossRef]
- De Benedittis, S.; Fortunato, F.; Cava, C.; Gallivanone, F.; Iaccino, E.; Caligiuri, M.E.; Castiglioni, I.; Bertoli, G.; Manna, I.; Labate, A.; et al. Circulating microRNA: The potential novel diagnostic biomarkers to predict drug resistance in temporal lobe epilepsy, a pilot study. Int. J. Mol. Sci. 2021, 22, 702. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Bin, H.; Chen, W. Inhibition of microRNA-103a inhibits the activation of astrocytes in hippocampus tissues and improves the pathological injury of neurons of epilepsy rats by regulating BDNF. Cancer Cell Int. 2019, 19, 109. [Google Scholar] [CrossRef] [PubMed]
- Brennan, G.P.; Bauer, S.; Engel, T.; Jimenez-Mateos, E.M.; Del Gallo, F.; Hill, T.D.M.; Connolly, N.M.; Costard, L.S.; Neubert, V.; Salvetti, B.; et al. Genome-wide microRNA profiling of plasma from three different animal models identifies biomarkers of temporal lobe epilepsy. Neurobiol. Dis. 2020, 144, 105048. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Wang, J.; Zhang, B. RefFinder: A web-based tool for comprehensively analyzing and identifying reference genes. Funct. Integr. Genom. 2023, 23, 125. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Sourvinou, I.S.; Markou, A.; Lianidou, E.S. Quantification of circulating miRNAs in plasma: Effect of preanalytical and analytical parameters on their isolation and stability. J. Mol. Diagn. 2013, 15, 827–834. [Google Scholar] [CrossRef]
- Schwarzenbach, H.; da Silva, A.M.; Calin, G.; Pantel, K. Data normalization strategies for MicroRNA quantification. Clin. Chem. 2015, 61, 1333–1342. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef]
- Liu, W.; Wang, X. Prediction of functional microRNA targets by integrative modeling of microRNA binding and target expression data. Genome Biol. 2019, 20, 18. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef]
- Jung, M.K.; Mun, J.Y. Sample preparation and imaging of exosomes by transmission electron microscopy. J. Vis. Exp. 2018, 131, e56482. [Google Scholar] [CrossRef]
- Aguilera-Rojas, M.; Badewien-Rentzsch, B.; Plendl, J.; Kohn, B.; Einspanier, R. Exploration of serum- and cell culture-derived exosomes from dogs. BMC Vet. Res. 2018, 14, 179. [Google Scholar] [CrossRef] [PubMed]
- Diomaiuto, E.; Principe, V.; De Luca, A.; Laperuta, F.; Alterisio, C.; Di Loria, A. Exosomes in dogs and cats: An innovative approach to neoplastic and non-neoplastic diseases. Pharmaceuticals 2021, 14, 766. [Google Scholar] [CrossRef] [PubMed]
- Ichii, O.; Ohta, H.; Horino, T.; Nakamura, T.; Hosotani, M.; Mizoguchi, T.; Morishita, K.; Nakamura, K.; Hoshino, Y.; Takagi, S.; et al. Urinary exosome-derived microRNAs reflecting the changes of renal function and histopathology in dogs. Sci. Rep. 2017, 7, 40340. [Google Scholar] [CrossRef]
- Lasko, T.A.; Bhagwat, J.G.; Zou, K.H.; Ohno-Machado, L. The use of receiver operating characteristic curves in biomedical informatics. J. Biomed. Inform. 2005, 38, 404–415. [Google Scholar] [CrossRef]
- Beghi, E. The epidemiology of epilepsy. Neuroepidemiology 2020, 54, 185–191. [Google Scholar] [CrossRef]
- Potschka, H.; Fischer, A.; Löscher, W.; Volk, H.A. Pathophysiology of drug-resistant canine epilepsy. Vet. J. 2023, 296–297, 105990. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Chen, S.-D.; Pan, H.-Y.; Huang, J.-B.; Liu, X.-P.; Li, J.-H.; Ho, C.-J.; Tsai, M.H.; Yang, J.L.; Chen, S.F.; Chen, N.C.; et al. Circulating MicroRNAs from serum exosomes may serve as a putative biomarker in the diagnosis and treatment of patients with focal cortical dysplasia. Cells 2020, 9, 1867. [Google Scholar] [CrossRef] [PubMed]
- Villatoro, A.J.; Martín-Astorga, M.D.C.; Alcoholado, C.; Becerra, J. Canine colostrum exosomes: Characterization and influence on the canine mesenchymal stem cell secretory profile and fibroblast anti-oxidative capacity. BMC Vet. Res. 2020, 16, 417. [Google Scholar] [CrossRef] [PubMed]
- Villatoro, A.J.; Alcoholado, C.; Martín-Astorga, M.D.C.; Rico, G.; Fernández, V.; Becerra, J. Characterization of the secretory profile and exosomes of limbal stem cells in the canine species. PLoS ONE 2020, 15, 0244327. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Sakamaki, Y.; Nakashima, S.; Fukuda, M. Rab39 and its effector UACA regulate basolateral exosome release from polarized epithelial cells. Cell Rep. 2022, 39, 110875. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, L.; Xu, Z.; Xiong, X.; Yu, Y.; Wu, H.; Qiao, H.; Zhong, J.; Zhao, Z.; Dai, J.; et al. A functionalized collagen-I scaffold delivers microRNA 21-loaded exosomes for spinal cord injury repair. Acta Biomater. 2022, 154, 385–400. [Google Scholar] [CrossRef]
- Kugeratski, F.G.; Hodge, K.; Lilla, S.; McAndrews, K.M.; Zhou, X.; Hwang, R.F.; Zanivan, S.; Kalluri, R. Quantitative proteomics identifies the core proteome of exosomes with syntenin-1 as the highest abundant protein and a putative universal biomarker. Nat. Cell Biol. 2021, 23, 631–641. [Google Scholar] [CrossRef]
- Lin, W.-H.; Li, X.-F.; Lin, M.-X.; Zhou, Y.; Huang, H.-P. Novel insights into the effect of paroxetine administration in pilocarpine-induced chronic epileptic rats. Mol. Med. Rep. 2017, 16, 8245–8252. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. mTOR signaling at a glance. J. Cell Sci. 2009, 122, 3589–3594. [Google Scholar] [CrossRef]
- Raoof, R.; Bauer, S.; El Naggar, H.; Connolly, N.M.C.; Brennan, G.P.; Brindley, E.; Hill, T.; McArdle, H.; Spain, E.; Forster, R.J.; et al. Dual-center, dual-platform microRNA profiling identifies potential plasma biomarkers of adult temporal lobe epilepsy. eBioMedicine 2018, 38, 127–141. [Google Scholar] [CrossRef]
- Wu, Z.-D.; Feng, Y.; Ma, Z.-X.; Liu, Z.; Xiong, H.-H.; Zhou, Z.-P.; Ouyang, L.S.; Xie, F.K.; Tang, Y.M.; Wu, Z.D.; et al. MicroRNAs: Protective regulators for neuron growth and development. Neural Regen. Res. 2023, 18, 734. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Xu, J.; Li, L.; Li, H.; Mao, S.; Zhang, F.; Zen, K.; Zhang, C.Y.; Zhang, Q. MicroRNA-23a/b and microRNA-27a/b suppress Apaf-1 protein and alleviate hypoxia-induced neuronal apoptosis. Cell Death Dis. 2014, 5, e1132. [Google Scholar] [CrossRef]
- Xi, T.; Jin, F.; Zhu, Y.; Wang, J.; Tang, L.; Wang, Y.; Liebeskind, D.S.; Scalzo, F.; He, Z. miR-27a-3p protects against blood–brain barrier disruption and brain injury after intracerebral hemorrhage by targeting endothelial aquaporin-11. J. Biol. Chem. 2018, 293, 20041–20050. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Yang, Y.; He, X.; Wang, X. miR-16-1 expression, heat shock protein 70 and inflammatory reactions in astrocytes of mice with epilepsy induced by encephalitis B virus infection. Exp. Ther. Med. 2017, 14, 495–498. [Google Scholar] [CrossRef]
- Fabbri, E.; Montagner, G.; Bianchi, N.; Finotti, A.; Borgatti, M.; Lampronti, I.; Cabrini, G.; Gambari, R. MicroRNA miR-93-5p regulates expression of IL-8 and VEGF in neuroblastoma SK-N-AS cells. Oncol. Rep. 2016, 35, 2866–2872. [Google Scholar] [CrossRef]
- Carroll, R.C.; Beattie, E.C.; von Zastrow, M.; Malenka, R.C. Role of ampa receptor endocytosis in synaptic plasticity. Nat. Rev. Neurosci. 2001, 2, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Blair, R.E.; Sombati, S.; Lawrence, D.C.; McCay, B.D.; DeLorenzo, R.J. Epileptogenesis causes acute and chronic increases in GABAA receptor endocytosis that contributes to the induction and maintenance of seizures in the hippocampal culture model of acquired epilepsy. J. Pharmacol. Exp. Ther. 2004, 310, 871–880. [Google Scholar] [CrossRef]
- Khan, S.; Nobili, L.; Khatami, R.; Loddenkemper, T.; Cajochen, C.; Dijk, D.-J.; Eriksson, S.H. Circadian rhythm and epilepsy. Lancet Neurol. 2018, 17, 1098–1108. [Google Scholar] [CrossRef]
- Moon, J.; Lee, S.-T.; Choi, J.; Jung, K.-H.; Yang, H.; Khalid, A.; Kim, J.M.; Park, K.I.; Shin, J.W.; Ban, J.J.; et al. Unique behavioral characteristics and microRNA signatures in a drug resistant epilepsy model. PLoS ONE. 2014, 9, 85617. [Google Scholar] [CrossRef]
- Mandolesi, G.; De Vito, F.; Musella, A.; Gentile, A.; Bullitta, S.; Fresegna, D.; Sepman, H.; Di Sanza, C.; Haji, N.; Mori, F.; et al. MiR-142-3p is a key regulator of IL-1β-dependent synaptopathy in neuroinflammation. J. Neurosci. 2017, 37, 546–561. [Google Scholar] [CrossRef]
- Holló, A.; Clemens, Z.; Lakatos, P. Epilepsy and vitamin D. Int. J. Neurosci. 2014, 124, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Tong, M.; Fu, Y.; Chen, F.; Zhang, S.; Chen, H.; Ma, X.; Li, D.; Liu, X.; Zhong, Q. Lipids and membrane-associated proteins in autophagy. Protein Cell 2021, 12, 520–544. [Google Scholar] [CrossRef] [PubMed]
- Hahn, T.J.; Birge, S.J.; Scharp, C.R.; Avioli, L.V. Phenobarbital-induced alterations in vitamin D metabolism. J. Clin. Investig. 1972, 51, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Wayman, G.A.; Davare, M.; Ando, H.; Fortin, D.; Varlamova, O.; Cheng, H.-Y.M.; Marks, D.; Obrietan, K.; Soderling, T.R.; Goodman, R.H.; et al. An activity-regulated microRNA controls dendritic plasticity by down-regulating p250GAP. Proc. Natl. Acad. Sci. USA 2008, 105, 9093–9098. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Average Diameter of Particle (nm) | Concentration (Particle/mL) |
|---|---|---|
| DRE1 | 104 ± 4.4 | 6.81 × 1011 ± 6.93 × 109 |
| DRE3 | 102 ± 4.9 | 7.85 × 1011 ± 3.43 × 1010 |
| DSE2 | 89.4 ± 5.5 | 6.94 × 1010 ± 8.45 × 109 |
| DSE5 | 89.3 ± 3.4 | 7.60 × 1011 ± 9.73 × 109 |
| DSE10 | 84.8 ± 2.3 | 4.57 × 1011 ± 1.77 × 1010 |
| C9 | 98.4 ± 1.2 | 4.31 × 1011 ± 3.02 × 109 |
| C10 | 97.5 ± 3.9 | 2.40 × 1011 ± 6.09 × 109 |
| C11 | 132.9 ± 3.4 | 7.61 × 1011 ± 1.84 × 1010 |
| miRNA | Pathway | nGenes | Fold Enrichment | FDR |
|---|---|---|---|---|
| miR-16 | Oocyte meiosis | 7 | 6.8 | 1.7 × 10−2 |
| Cell cycle | 6 | 5.9 | 2.8 × 10−2 | |
| FoxO signaling pathway | 6 | 5.7 | 2.8 × 10−2 | |
| MTOR signaling pathway | 7 | 5.7 | 2.6 × 10−2 | |
| Autophagy | 6 | 5.3 | 3.4 × 10−2 | |
| PI3K-Akt signaling pathway | 10 | 3.5 | 2.8 × 10−2 | |
| miR-27a-3p | EGFR tyrosine kinase inhibitor resistance | 6 | 7.1 | 4.8 × 10−2 |
| miR-93-5p | Circadian rhythm | 4 | 13.7 | 1.9 × 10−2 |
| Endocytosis | 13 | 5.5 | 1.7 × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Gracia, M.; Moreno-Martinez, L.; Hernaiz, A.; Usón, S.; Moral, J.; Sanz-Rubio, D.; Zaragoza, P.; Palacio, J.; Rosado, B.; Osta, R.; et al. Analysis of Plasma-Derived Exosomal MicroRNAs as Potential Biomarkers for Canine Idiopathic Epilepsy. Animals 2024, 14, 252. https://doi.org/10.3390/ani14020252
García-Gracia M, Moreno-Martinez L, Hernaiz A, Usón S, Moral J, Sanz-Rubio D, Zaragoza P, Palacio J, Rosado B, Osta R, et al. Analysis of Plasma-Derived Exosomal MicroRNAs as Potential Biomarkers for Canine Idiopathic Epilepsy. Animals. 2024; 14(2):252. https://doi.org/10.3390/ani14020252
Chicago/Turabian StyleGarcía-Gracia, Mireya, Laura Moreno-Martinez, Adelaida Hernaiz, Sebastián Usón, Jon Moral, David Sanz-Rubio, Pilar Zaragoza, Jorge Palacio, Belén Rosado, Rosario Osta, and et al. 2024. "Analysis of Plasma-Derived Exosomal MicroRNAs as Potential Biomarkers for Canine Idiopathic Epilepsy" Animals 14, no. 2: 252. https://doi.org/10.3390/ani14020252