
Carbohydrases and Phytase in Poultry and Pig Nutrition: A Review beyond the Nutrients and Energy Matrix

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Exogenous Enzymes and Their Functional Mechanisms of Action
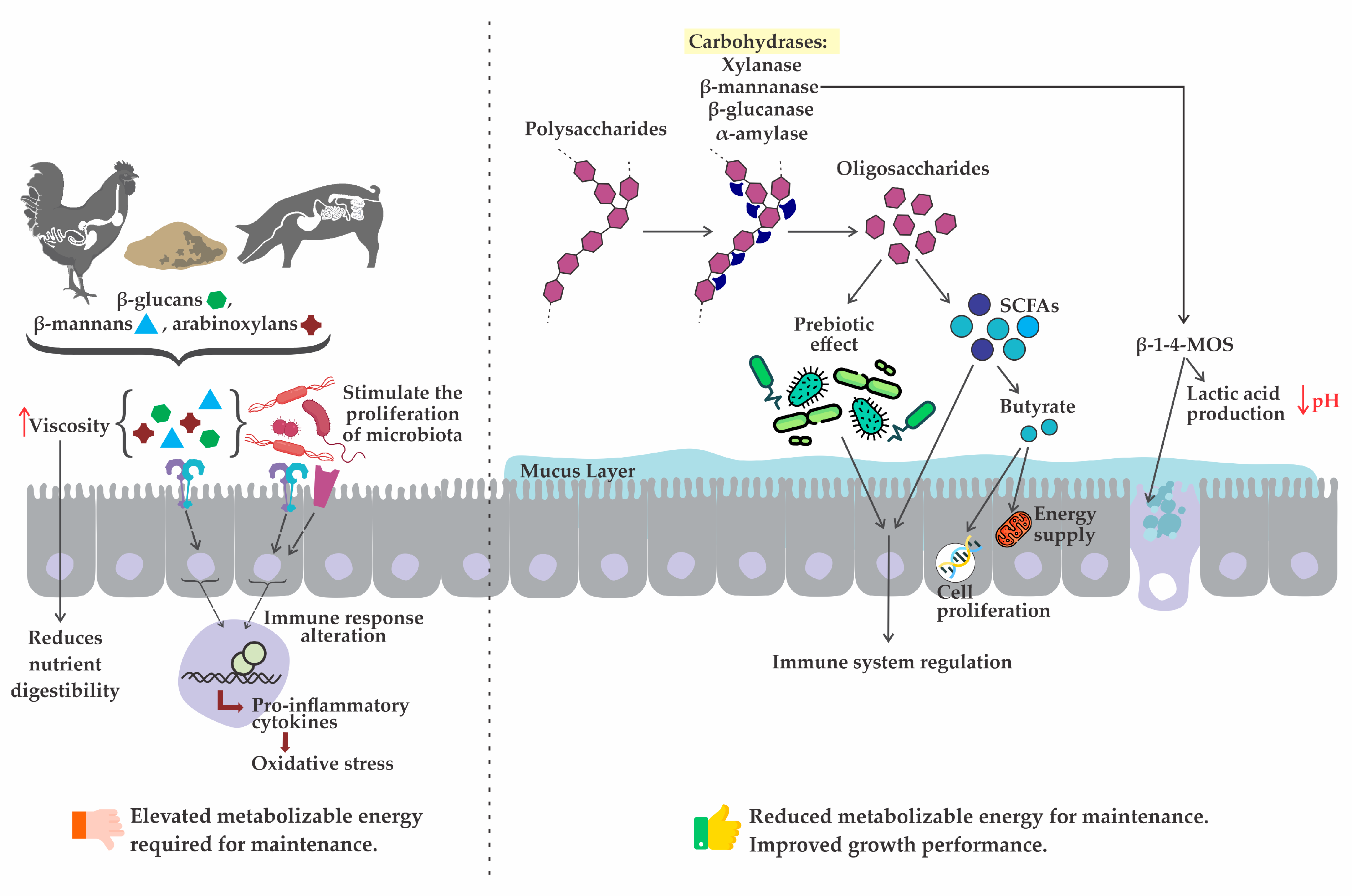
2.1. Carbohydrases
2.1.1. Xylanase
2.1.2. β-Mannanase
2.1.3. β-Glucanase
2.1.4. α-Amylase
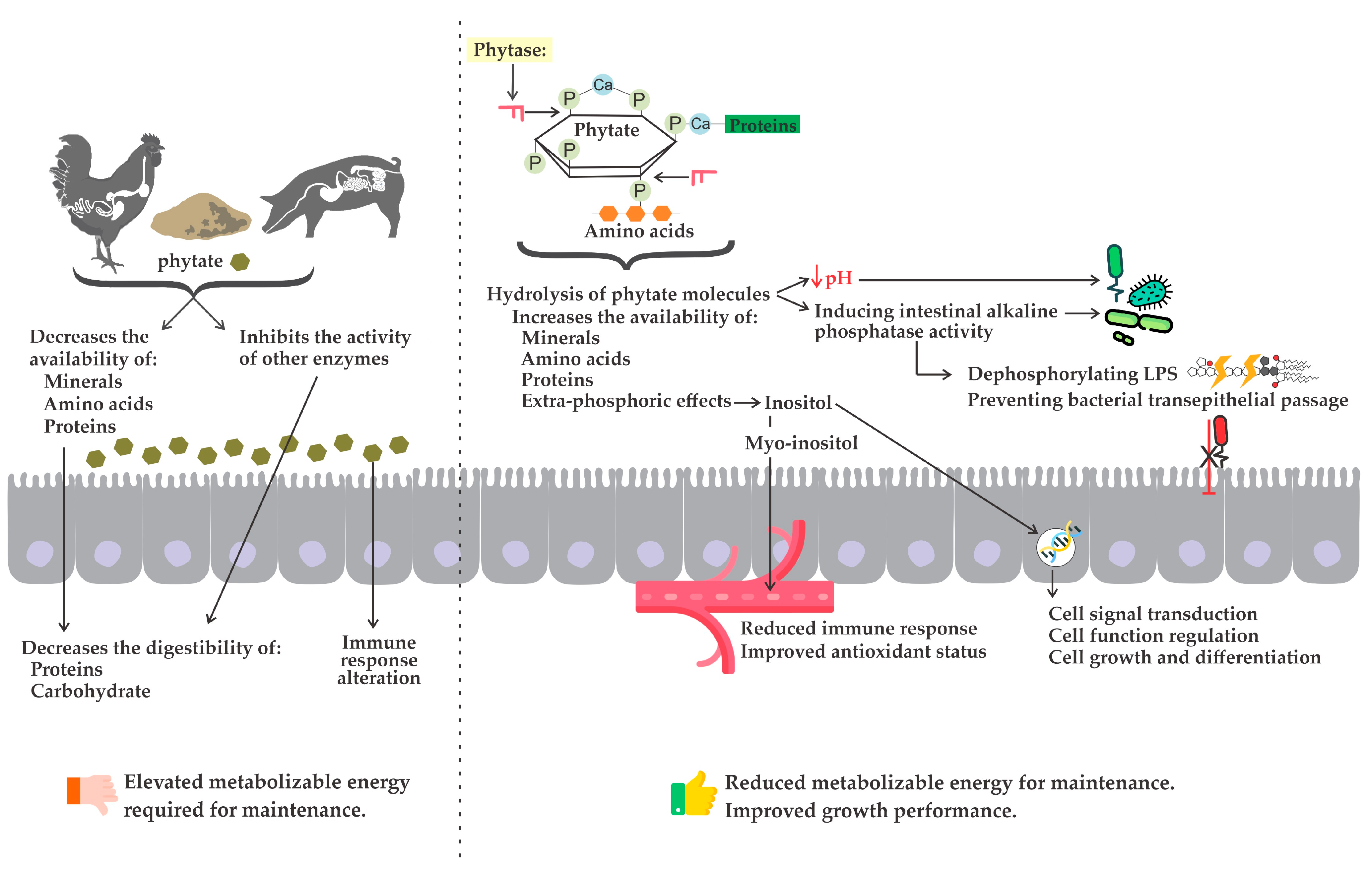
2.2. Phytase
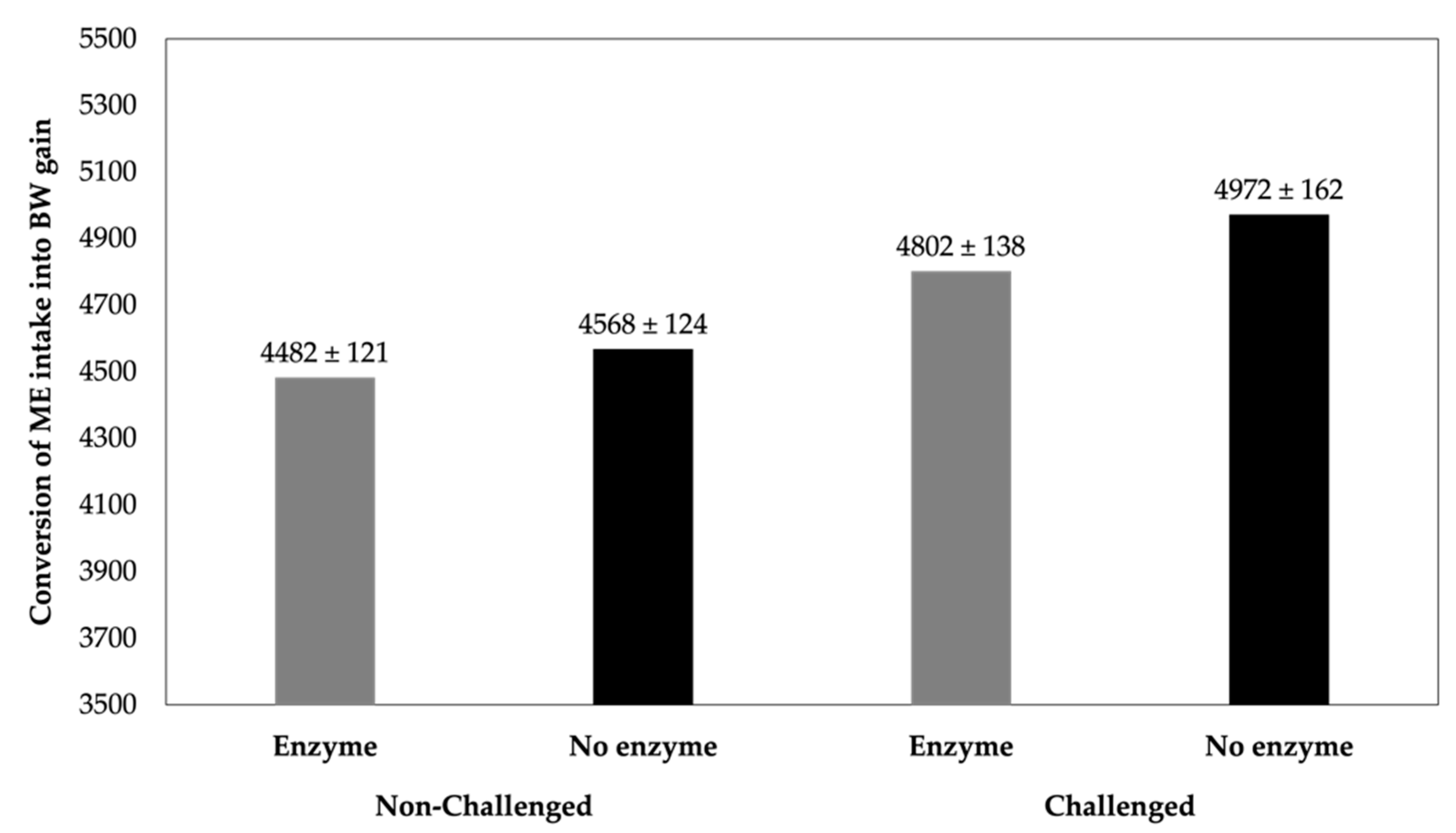
3. The Role of Dietary Exogenous Enzymes in Poultry and Pig under Challenging Conditions
4. Conclusions and Future Directions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kiarie, E.; Walsh, M.C.; Nyachoti, C.M. Performance, digestive function, and mucosal responses to selected feed additives for pigs. J. Anim. Sci. 2016, 94, 169–180. [Google Scholar] [CrossRef]
- Kiarie, E.G.; Steelman, S.; Martinez, M.; Livingston, K. Significance of single β-mannanase supplementation on performance and energy utilization in broiler chickens, laying hens, turkeys, sows, and nursery-finish pigs: A meta-analysis and systematic review. Transl. Anim. Sci. 2021, 5, txab160. [Google Scholar] [CrossRef] [PubMed]
- Dersjant-Li, Y.; Awati, A.; Schulze, H.; Partridge, G. Phytase in non-ruminant animal nutrition: A critical review on phytase activities in the gastrointestinal tract and influencing factors. J. Sci. Food Agric. 2015, 95, 878–896. [Google Scholar] [CrossRef] [PubMed]
- Dersjant-Li, Y.; Dusel, G. Increasing the dosing of a Buttiauxella phytase improves phytate degradation, mineral, energy, and amino acid digestibility in weaned pigs fed a complex diet based on wheat, corn, soybean meal, barley, and rapeseed meal. J. Anim. Sci. 2019, 97, 2524–2533. [Google Scholar] [CrossRef]
- Dal Pont, G.C.; Eyng, C.; Bortoluzzi, C.; Kogut, M.H. Enzymes and Gut Health in Monogastric Animals: Effects beyond Digestibility. In Gut Microbiota, Immunity, and Health in Production Animals; Kogut, M.H., Zhang, G., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 33–55. [Google Scholar]
- Aranda-Aguirre, E.; Robles-Jimenez, L.E.; Osorio-Avalos, J.; Vargas-Bello-Pérez, E.; Gonzalez-Ronquillo, M. A systematic-review on the role of exogenous enzymes on the productive performance at weaning, growing and finishing in pigs. Vet. Anim. Sci. 2021, 14, 100195. [Google Scholar] [CrossRef] [PubMed]
- Petry, A.L.; Patience, J.F. Xylanase supplementation in corn-based swine diets: A review with emphasis on potential mechanisms of action. J. Anim. Sci. 2020, 98, skaa318. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.K.; Ingale, S.L.; Kim, J.S.; Kim, Y.W.; Kim, K.H.; Lohakare, J.D.; Lee, J.H.; Chae, B.J. Effects of exogenous enzyme supplementation to corn- and soybean meal-based or complex diets on growth performance, nutrient digestibility, and blood metabolites in growing pigs. J. Anim. Sci. 2012, 90, 3041–3048. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.E.; Zhou, F.X.; Dutra, W.M.; Kim, S.W. Dietary supplementation of xylanase and protease on growth performance, digesta viscosity, nutrient digestibility, immune and oxidative stress status, and gut health of newly weaned pigs. Anim. Nutr. 2019, 5, 351–358. [Google Scholar] [CrossRef]
- He, X.; Yu, B.; He, J.; Huang, Z.; Mao, X.; Zheng, P.; Luo, Y.; Luo, J.; Wang, Q.; Wang, H.; et al. Effects of xylanase on growth performance, nutrients digestibility and intestinal health in weaned piglets. Livest. Sci. 2020, 233, 103940. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, S.; Kim, S.W. Effects of supplemental xylanase on health of the small intestine in nursery pigs fed diets with corn distillers’ dried grains with solubles. J. Anim. Sci. 2020, 98, skaa185. [Google Scholar] [CrossRef]
- Aderibigbe, A.; Cowieson, A.; Sorbara, J.O.; Adeola, O. Intestinal starch and energy digestibility in broiler chickens fed diets supplemented with α-amylase. Poult. Sci. 2020, 99, 5907–5914. [Google Scholar] [CrossRef] [PubMed]
- Genova, J.L.; Rupolo, P.E.; de Azevedo, L.B.; Henz, D.; Carvalho, S.T.; Kipper, M.; Gonçalves, G.A.C.; Vilela, H.L.O.; Pasquetti, T.J.; de Oliveira, N.T.E.; et al. β-mannanase supplementation in diets reduced in 85 kcal metabolizable energy/kg containing xylanase-phytase improves gain to feed ratio, nutrient usage, and backfat thickness in finisher pigs. Front. Vet. Sci. 2023, 10, 1144692. [Google Scholar] [CrossRef] [PubMed]
- Stefanello, C.; Vieira, S.L.; Rios, H.V.; Simões, C.T.; Ferzola, P.H.; Sorbara, J.O.B.; Cowieson, A.J. Effects of energy, α-amylase, and β-xylanase on growth performance of broiler chickens. Anim. Feed Sci. Technol. 2017, 225, 205–212. [Google Scholar] [CrossRef]
- Kim, J.S.; Ingale, S.L.; Lee, S.H.; Kim, K.H.; Kim, J.S.; Lee, J.H.; Chae, B.J. Effects of energy levels of diet and β-mannanase supplementation on growth performance, apparent total tract digestibility and blood metabolites in growing pigs. Anim. Feed Sci. Technol. 2013, 186, 64–70. [Google Scholar] [CrossRef]
- Scapini, L.B.; de Cristo, A.B.; Schmidt, J.M.; Buzim, R.; Nogueira, L.K.; Palma, S.C.; Fernandes, J.I.M. Effect of β-Mannanase Supplementation in Conventional Diets on the Performance, Immune Competence and Intestinal Quality of Broilers Challenged with Eimeria sp. J. Appl. Poult. Res. 2019, 28, 1048–1057. [Google Scholar] [CrossRef]
- Mejicanos, G.A.; González-Ortiz, G.; Nyachoti, C.M. Effect of dietary supplementation of xylanase in a wheat-based diet containing canola meal on growth performance, nutrient digestibility, organ weight, and short-chain fatty acid concentration in digesta when fed to weaned pigs. J. Anim. Sci. 2020, 98, skaa064. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.C.; Sweeney, T.; Curley, E.; Gath, V.; Duffy, S.K.; Vigors, S.; Rajauria, G.; O’Doherty, J.V. Effect of β-glucanase and β-xylanase enzyme supplemented barley diets on nutrient digestibility, growth performance and expression of intestinal nutrient transporter genes in finisher pigs. Anim. Feed Sci. Technol. 2018, 238, 98–110. [Google Scholar] [CrossRef]
- Jang, J.-C.; Kim, D.H.; Hong, J.S.; Jang, Y.D.; Kim, Y.Y. Effects of Copra Meal Inclusion Level in Growing-Finishing Pig Diets Containing β-Mannanase on Growth Performance, Apparent Total Tract Digestibility, Blood Urea Nitrogen Concentrations and Pork Quality. Animals 2020, 10, 1840. [Google Scholar] [CrossRef]
- Torres-Pitarch, A.; Manzanilla, E.G.; Gardiner, G.E.; O’Doherty, J.V.; Lawlor, P.G. Systematic review and meta-analysis of the effect of feed enzymes on growth and nutrient digestibility in grow-finisher pigs: Effect of enzyme type and cereal source. Anim. Feed Sci. Technol. 2019, 251, 153–165. [Google Scholar] [CrossRef]
- Long, S.; Hu, J.; Mahfuz, S.; Ma, H.; Piao, X. Effects of dietary supplementation of compound enzymes on performance, nutrient digestibility, serum antioxidant status, immunoglobulins, intestinal morphology and microbiota community in weaned pigs. Arch. Anim. Nutr. 2021, 75, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, U.P.; Chen, H.; Kim, S.W.; Jha, R. Supplemental effect of xylanase and mannanase on nutrient digestibility and gut health of nursery pigs studied using both in vivo and in vitro models. Anim. Feed Sci. Technol. 2018, 245, 77–90. [Google Scholar] [CrossRef]
- Karimi, K.; Zhandi, M. The effect of β-mannanase and β-glucanase on small intestine morphology in male broilers fed diets containing various levels of metabolizable energy. J. Appl. Anim. Res. 2015, 43, 324–329. [Google Scholar] [CrossRef]
- Nari, N.; Ghasemi, H.A.; Hajkhodadadi, I.; Farahani, A.H.K. Intestinal microbial ecology, immune response, stress indicators, and gut morphology of male broiler chickens fed low-phosphorus diets supplemented with phytase, butyric acid, or Saccharomyces boulardii. Livest. Sci. 2020, 234, 103975. [Google Scholar] [CrossRef]
- Liu, F.; Li, J.; Ni, H.; Azad, M.A.K.; Mo, K.; Yin, Y. The effects of phytase and non-starch polysaccharide-hydrolyzing enzymes on trace element deposition, intestinal morphology, and cecal microbiota of growing-finishing pigs. Animals 2023, 13, 549. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.M.; González-Ortiz, G.; Melo-Durán, D.; Heo, J.M.; Cordero, G.; Bedford, M.R.; Kim, J.C. Stimbiotic supplementation improved performance and reduced inflammatory response via stimulating fiber fermenting microbiome in weaner pigs housed in a poor sanitary environment and fed an antibiotic-free low zinc oxide diet. PLoS ONE 2020, 15, e0240264. [Google Scholar] [CrossRef]
- Madrid, J.; Catalá-Gregori, P.; García, V.; Hernández, F. Effect of a multienzyme complex in wheat-soybean meal diet on digestibility of broiler chickens under different rearing conditions. Ital. J. Anim. Sci. 2010, 9, e1. [Google Scholar] [CrossRef]
- Adeola, O.; Cowieson, A.J. BOARD-INVITED REVIEW: Opportunities and challenges in using exogenous enzymes to improve nonruminant animal production. J. Anim. Sci. 2011, 89, 3189–3218. [Google Scholar] [CrossRef] [PubMed]
- Bach Knudsen, K.E.; Nørskov, N.P.; Bolvig, A.K.; Hedemann, M.S.; Lærke, H.N. Dietary fibers and associated phytochemicals in cereals. Mol. Nutr. Food Res. 2017, 61, 1600518. [Google Scholar] [CrossRef] [PubMed]
- Adebowale, T.O.; Yao, K.; Oso, A.O. Major cereal carbohydrates in relation to intestinal health of monogastric animals: A review. Anim. Nutr. 2019, 5, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.E.; Sparks, C.; Kim, S.W. Modulation of jejunal mucosa-associated microbiota in relation to intestinal health and nutrient digestibility in pigs by supplementation of β-glucanase to corn–soybean meal-based diets with xylanase. J. Anim. Sci. 2021, 99, skab190. [Google Scholar] [CrossRef]
- Dikeman, C.L.; Fahey, G.C. Viscosity as Related to Dietary Fiber: A Review. Crit. Rev. Food Sci. Nutr. 2006, 46, 649–663. [Google Scholar] [CrossRef] [PubMed]
- Bedford, M.R.; Cowieson, A.J. Exogenous enzymes and their effects on intestinal microbiology. Anim. Feed Sci. Technol. 2012, 173, 76–85. [Google Scholar] [CrossRef]
- Tiwari, U.P.; Fleming, S.A.; Abdul Rasheed, M.S.; Jha, R.; Dilger, R.N. The role of oligosaccharides and polysaccharides of xylan and mannan in gut health of monogastric animals. J. Nutr. Sci. 2020, 9, e21. [Google Scholar] [CrossRef] [PubMed]
- Huntley, N.F.; Nyachoti, C.M.; Patience, J.F. Lipopolysaccharide immune stimulation but not β-mannanase supplementation affects maintenance energy requirements in young weaned pigs. J. Anim. Sci. Biotechnol. 2018, 9, 47. [Google Scholar] [CrossRef]
- Yi, J.Q.; Piao, X.S.; Li, Z.C.; Zhang, H.Y.; Chen, Y.; Li, Q.Y.; Liu, J.D.; Zhang, Q.; Ru, Y.J.; Dong, B. The effects of enzyme complex on performance, intestinal health and nutrient digestibility of weaned pigs. Asian-Australas. J. Anim. Sci. 2013, 26, 1181–1188. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Kumar, B.; Verma, P. A detailed overview of xylanases: An emerging biomolecule for current and future prospective. Bioresour. Bioprocess. 2019, 6, 40. [Google Scholar] [CrossRef]
- Passos, A.A.; Park, I.; Ferket, P.; von Heimendahl, E.; Kim, S.W. Effect of dietary supplementation of xylanase on apparent ileal digestibility of nutrients, viscosity of digesta, and intestinal morphology of growing pigs fed corn and soybean meal based diet. Anim. Nutr. 2015, 1, 19–23. [Google Scholar] [CrossRef]
- Boontiam, W.; Phaenghairee, P.; Van Hoeck, V.; Vasanthakumari, B.L.; Somers, I.; Wealleans, A. Xylanase Impact beyond Performance: Effects on Gut Structure, Faecal Volatile Fatty Acid Content and Ammonia Emissions in Weaned Piglets Fed Diets Containing Fibrous Ingredients. Animals 2022, 12, 3043. [Google Scholar] [CrossRef] [PubMed]
- Petry, A.L.; Huntley, N.F.; Bedford, M.R.; Patience, J.F. Xylanase increased the energetic contribution of fiber and improved the oxidative status, gut barrier integrity, and growth performance of growing pigs fed insoluble corn-based fiber. J. Anim. Sci. 2020, 98, skaa233. [Google Scholar] [CrossRef]
- Petry, A.L.; Patience, J.F.; Koester, L.R.; Huntley, N.F.; Bedford, M.R.; Schmitz-Esser, S. Xylanase modulates the microbiota of ileal mucosa and digesta of pigs fed corn-based arabinoxylans likely through both a stimbiotic and prebiotic mechanism. PLoS ONE 2021, 16, e0246144. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, C.; Sun, Y.; Wang, G.; Chen, H.; Li, D.; Yu, X.; Chen, G. Xylanase and Fermented Polysaccharide of Hericium caputmedusae Reduce Pathogenic Infection of Broilers by Improving Antioxidant and Anti-Inflammatory Properties. Oxidative Med. Cell. Longev. 2018, 2018, 4296985. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cao, H.; Bao, C.; Liu, Y.; Dong, B.; Wang, C.; Shang, Z.; Cao, Y.; Liu, S. Effects of Xylanase in Corn- or Wheat-Based Diets on Cecal Microbiota of Broilers. Front. Microbiol. 2021, 12, 757066. [Google Scholar] [CrossRef]
- Dong, B.; Liu, S.; Wang, C.; Cao, Y. Effects of xylanase supplementation to wheat-based diets on growth performance, nutrient digestibility and gut microbes in weanling pigs. Asian-Australas. J. Anim. Sci. 2018, 31, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.E.; Tyus, J.; Kim, S.W. Synbiotic Effects of Enzyme and Probiotics on Intestinal Health and Growth of Newly Weaned Pigs Challenged with Enterotoxigenic F18 + Escherichia coli. Front. Vet. Sci. 2020, 7, 573. [Google Scholar] [CrossRef]
- Baker, J.T.; Duarte, M.E.; Holanda, D.M.; Kim, S.W. Friend or Foe? Impacts of Dietary Xylans, Xylooligosaccharides, and Xylanases on Intestinal Health and Growth Performance of Monogastric Animals. Animals 2021, 11, 609. [Google Scholar] [CrossRef] [PubMed]
- Bedford, M.R. The evolution and application of enzymes in the animal feed industry: The role of data interpretation. Br. Poult. Sci. 2018, 59, 486–493. [Google Scholar] [CrossRef]
- Bautil, A.; Verspreet, J.; Buyse, J.; Goos, P.; Bedford, M.R.; Courtin, C.M. Arabinoxylan-oligosaccharides kick-start arabinoxylan digestion in the aging broiler. Poult. Sci. 2020, 99, 2555–2565. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Zhang, Z.; Wang, S.; Cao, L.; Zhou, L.; Sun, A.; Zhong, Z.; Nabben, M. Microbial-Driven Butyrate Regulates Jejunal Homeostasis in Piglets During the Weaning Stage. Front. Microbiol. 2019, 9, 3335. [Google Scholar] [CrossRef]
- Diao, H.; Jiao, A.R.; Yu, B.; Mao, X.B.; Chen, D.W. Gastric infusion of short-chain fatty acids can improve intestinal barrier function in weaned piglets. Genes Nutr. 2019, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Thomson, A.; Hemphill, D.; Jeejeebhoy, K.N. Oxidative Stress and Antioxidants in Intestinal Disease. Dig. Dis. 1998, 16, 152–158. [Google Scholar] [CrossRef]
- Pirgozliev, V.R.; Mansbridge, S.C.; Whiting, I.M.; Abdulla, J.M.; Rose, S.P.; Kljak, K.; Johnson, A.; Drijfhout, F.; Atanasov, A.G. The Benefits of Exogenous Xylanase in Wheat—Soy Based Broiler Chicken Diets, Consisting of Different Soluble Non-Starch Polysaccharides Content. Poultry 2023, 2, 123–133. [Google Scholar] [CrossRef]
- Liu, D.; Guo, S.; Guo, Y. Xylanase supplementation to a wheat-based diet alleviated the intestinal mucosal barrier impairment of broiler chickens challenged by Clostridium perfringens. Avian Pathol. 2012, 41, 291–298. [Google Scholar] [CrossRef]
- McCleary, B.V.; Matheson, N.K. Enzymic analysis of polysaccharide structure. Adv. Carbohydr. Chem. Biochem. 1986, 44, 147–176. [Google Scholar]
- Dhawan, S.; Kaur, J. Microbial Mannanases: An Overview of Production and Applications. Crit. Rev. Biotechnol. 2007, 27, 197–216. [Google Scholar] [CrossRef]
- Knudsen, K.E.B. Fiber and nonstarch polysaccharide content and variation in common crops used in broiler diets. Poult. Sci. 2014, 93, 2380–2393. [Google Scholar] [CrossRef]
- Kim, J.S.; Ingale, S.L.; Hosseindoust, A.R.; Lee, S.H.; Lee, J.H.; Chae, B.J. Effects of mannan level and β-mannanase supplementation on growth performance, apparent total tract digestibility and blood metabolites of growing pigs. Animal 2017, 11, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, B.; Ingale, S.L.; Park, J.H.; Rathi, P.C.; Shanmugam, S.; Kim, I.H. Inclusion of dietary β-mannanase improves performance and ileal digestibility and reduces ileal digesta viscosity of broilers fed corn-soybean meal based diet. Poult. Sci. 2018, 97, 3097–3101. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.; Bailey, C.A.; Cartwright, A.L. β-Mannanase ameliorates viscosity-associated depression of growth in broiler chickens fed guar germ and hull fractions. Poult. Sci. 2003, 82, 1925–1931. [Google Scholar] [CrossRef] [PubMed]
- Okubo, T.; Ishihara, N.; Takahashi, H.; Fujisawa, T.; Kim, M.; Yamamoto, T.; Mitsuoka, T. Effects of Partially Hydrolyzed Guar Gum Intake on Human Intestinal Microflora and Its Metabolism. Biosci. Biotechnol. Biochem. 1994, 58, 1364–1369. [Google Scholar] [CrossRef]
- Mudgil, D.; Barak, S.; Patel, A.; Shah, N. Partially hydrolyzed guar gum as a potential prebiotic source. Int. J. Biol. Macromol. 2018, 112, 207–210. [Google Scholar] [CrossRef]
- Mary, P.R.; Prashanth, K.V.H.; Vasu, P.; Kapoor, M. Structural diversity and prebiotic potential of short chain β-manno-oligosaccharides generated from guar gum by endo-β-mannanase (ManB-1601). Carbohydr. Res. 2019, 486, 107822. [Google Scholar] [CrossRef]
- Kiarie, E.G.; Steelman, S.; Martinez, M. Does supplementing β-mannanase modulate the feed-induced immune response and gastrointestinal ecology in poultry and pigs? An appraisal. Front. Anim. Sci. 2022, 3, 875095. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Scapini, L.B.; Ribeiro, M.V.; Pivetta, M.R.; Buzim, R.; Fernandes, J.I.M. Effects of β-mannanase supplementation on the intestinal microbiota composition of broiler chickens challenged with a coccidiosis vaccine. Livest. Sci. 2019, 228, 187–194. [Google Scholar] [CrossRef]
- Arsenault, R.J.; Lee, J.T.; Latham, R.; Carter, B.; Kogut, M.H. Changes in immune and metabolic gut response in broilers fed β-mannanase in β-mannan-containing diets. Poult. Sci. 2017, 96, 4307–4316. [Google Scholar] [CrossRef]
- Chen, W.L.; Chen, H.L.; Guo, G.W.; Huang, Y.C.; Chen, C.Y.; Tsai, Y.; Huang, K.F.; Yang, C.H. Locust bean gum galactomannan hydrolyzed by thermostable β-d-mannanase may reduce the secretion of pro-inflammatory factors and the release of granule constituents. Int. J. Biol. Macromol. 2018, 114, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.; Hsiao, H. Effect of β--mannan (Hemicell® feed enzyme) on acute phase protein levels in chickens and turkeys. Abstract 231. In Proceedings of the Poultry Science Association, 95th Annual Meeting, Edmonton, AB, Canada, 16–19 July 2006; pp. 16–19. [Google Scholar]
- Jang, J.-C.; Kim, K.H.; Jang, Y.D.; Kim, Y.Y. Effects of Dietary β-Mannanase Supplementation on Growth Performance, Apparent Total Tract Digestibility, Intestinal Integrity, and Immune Responses in Weaning Pigs. Animals 2020, 10, 703. [Google Scholar] [CrossRef]
- Huntley, N.F.; Gould, S.A.; Patience, J.F. Evaluation of the effect of β-mannanase supplementation and mannans on nursery pig growth performance and serum acute-phase protein concentrations. Can. J. Anim. Sci. 2020, 100, 111–118. [Google Scholar] [CrossRef]
- Karunaratne, N.; Classen, H. Chapter 10 Beta-glucans and beta-glucanase in animal nutrition, do we understand their full effects? In The Value of Fibre: Engaging the Second Brain for Animal Nutrition; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; pp. 433–438. [Google Scholar]
- Le Gall, M.; Serena, A.; Jørgensen, H.; Theil, P.K.; Bach Knudsen, K.E. The role of whole-wheat grain and wheat and rye ingredients on the digestion and fermentation processes in the gut—A model experiment with pigs. Br. J. Nutr. 2009, 102, 1590–1600. [Google Scholar] [CrossRef] [PubMed]
- Hooda, S.; Matte, J.J.; Vasanthan, T.; Zijlstra, R.T. Dietary oat β-glucan reduces peak net glucose flux and insulin production and modulates plasma incretin in portal-vein catheterized grower pigs. J. Nutr. 2010, 140, 1564–1569. [Google Scholar] [CrossRef]
- Estrada, A.; Yun, C.H.; Van Kessel, A.; Li, B.; Hauta, S.; Laarveld, B. Immunomodulatory Activities of Oat β-Glucan In Vitro and In Vivo. Microbiol. Immunol. 1997, 41, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.; Dove, C.R.; Cline, P.M.; Owusu-Asiedu, A.; Walsh, M.C.; Azain, M. The effect of adding xylanase or β-glucanase to diets with corn distillers dried grains with solubles (CDDGS) on growth performance and nutrient digestibility in nursery pigs. Livest. Sci. 2017, 197, 46–52. [Google Scholar] [CrossRef]
- Sun, H.Y.; Ingale, S.L.; Rathi, P.; Kim, I.H. Influence of β-glucanase supplementation on growth performance, nutrient digestibility, blood parameters, and meat quality in broilers fed wheat–barley–soybean diet. Can. J. Anim. Sci. 2019, 99, 384–391. [Google Scholar] [CrossRef]
- Munyaka, P.M.; Nandha, N.K.; Kiarie, E.; Nyachoti, C.M.; Khafipour, E. Impact of combined β-glucanase and xylanase enzymes on growth performance, nutrients utilization and gut microbiota in broiler chickens fed corn or wheat-based diets. Poult. Sci. 2016, 95, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Kanjan, P.; Sahasrabudhe, N.M.; de Haan, B.J.; de Vos, P. Immune effects of β-glucan are determined by combined effects on Dectin-1, TLR2, 4 and 5. J. Funct. Foods 2017, 37, 433–440. [Google Scholar] [CrossRef]
- Sahasrabudhe, N.M.; Tian, L.; van den Berg, M.; Bruggeman, G.; Bruininx, E.; Schols, H.A.; Faas, M.M.; de Vos, P. Endo-glucanase digestion of oat β-Glucan enhances Dectin-1 activation in human dendritic cells. J. Funct. Foods 2016, 21, 104–112. [Google Scholar] [CrossRef]
- Chen, W.; Wang, S.; Xu, R.; Xia, W.; Ruan, D.; Zhang, Y.; Mohammed, K.A.F.; Azzam, M.M.M.; Fouad, A.M.; Li, K.; et al. Effects of dietary barley inclusion and glucanase supplementation on the production performance, egg quality and digestive functions in laying ducks. Anim. Nutr. 2021, 7, 176–184. [Google Scholar] [CrossRef]
- Cowieson, A.J.; Vieira, S.L.; Stefanello, C. Exogenous Microbial Amylase in the Diets of Poultry: What do We Know? J. Appl. Poult. Res. 2019, 28, 556–565. [Google Scholar] [CrossRef]
- Kumar, S.; Chakravarty, S. 8.1 The importance of carbohydrates and amylase in human nutrition. In Enzymes in Human and Animal Nutrition: Principles and Perspectives; Academic Press: Cambridge, MA, USA, 2018; p. 163. [Google Scholar]
- Moran, E.T., Jr. Digestion and Absorption of Carbohydrates in Fowl and Events through Perinatal Development. J. Nutr. 1985, 115, 665–674. [Google Scholar] [CrossRef]
- Bach Knudsen, K.E.; Hedemann, M.S.; Lærke, H.N. The role of carbohydrates in intestinal health of pigs. Anim. Feed Sci. Technol. 2012, 173, 41–53. [Google Scholar] [CrossRef]
- Giuberti, G.; Gallo, A.; Masoero, F.; Ferraretto, L.F.; Hoffman, P.C.; Shaver, R.D. Factors affecting starch utilization in large animal food production system: A review. Starch-Stärke 2014, 66, 72–90. [Google Scholar] [CrossRef]
- Stefanello, C.; Vieira, S.L.; Soster, P.; Santos, B.M.D.; Dalmoro, Y.K.; Favero, A.; Cowieson, A.J. Utilization of corn-based diets supplemented with an exogenous α-amylase for broilers. Poult. Sci. 2019, 98, 5862–5869. [Google Scholar] [CrossRef] [PubMed]
- Córdova-Noboa, H.A.; Oviedo-Rondón, E.O.; Ortiz, A.; Matta, Y.; Hoyos, S.; Buitrago, G.D.; Martinez, J.D.; Yanquen, J.; Peñuela, L.; Sorbara, J.O.B.; et al. Corn drying temperature, particle size, and amylase supplementation influence growth performance, digestive tract development, and nutrient utilization of broilers. Poult. Sci. 2020, 99, 5681–5696. [Google Scholar] [CrossRef] [PubMed]
- Woyengo, T.A.; Bogota, K.J.; Noll, S.L.; Wilson, J. Enhancing nutrient utilization of broiler chickens through supplemental enzymes. Poult. Sci. 2019, 98, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Dzierva, L.; dos Santos, M.C.; de Almeida, L.M.; da Rocha, C.; Bassi, L.S.; Muramatsu, K.; Maiorka, A. Feed particle size and amylase supplementation in diets for nursery piglets: Digestibility and growth performance. Livest. Sci. 2023, 271, 105211. [Google Scholar] [CrossRef]
- Tester, R.F.; Karkalas, J.; Qi, X. Starch—Composition, fine structure and architecture. J. Cereal Sci. 2004, 39, 151–165. [Google Scholar] [CrossRef]
- Schramm, V.G.; Massuquetto, A.; Bassi, L.S.; Zavelinski, V.A.B.; Sorbara, J.O.B.; Cowieson, A.J.; Félix, A.P.; Maiorka, A. Exogenous α-amylase improves the digestibility of corn and corn–soybean meal diets for broilers. Poult. Sci. 2021, 100, 101019. [Google Scholar] [CrossRef] [PubMed]
- Rupolo, P.E.; Monteiro, D.P.; Ribeiro, T.P.; de Azevedo, L.B.; Gregory, C.R.; Careli, P.S.; Carvalho, S.T.; Paiano, D.; Hannas, M.I.; Nunes, R.V.; et al. Effects of supplementation of α-amylase alone in a model of growing and finishing pigs fed metabolizable energy-reduced diets. Livest. Sci. 2023, 278, 105361. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhou, Y.; Lu, F.; Han, Z.; Wang, T. Effects of Different Levels of Supplementary Alpha-amylase on Digestive Enzyme Activities and Pancreatic Amylase mRNA Expression of Young Broilers. Asian-Australas. J. Anim. Sci. 2008, 21, 97–102. [Google Scholar] [CrossRef]
- Yin, D.; Yin, X.; Wang, X.; Lei, Z.; Wang, M.; Guo, Y.; Aggrey, S.E.; Nie, W.; Yuan, J. Supplementation of amylase combined with glucoamylase or protease changes intestinal microbiota diversity and benefits for broilers fed a diet of newly harvested corn. J. Anim. Sci. Biotechnol. 2018, 9, 24. [Google Scholar] [CrossRef]
- Tan, F.P.Y.; Beltranena, E.; Zijlstra, R.T. Resistant starch: Implications of dietary inclusion on gut health and growth in pigs: A review. J. Anim. Sci. Biotechnol. 2021, 12, 124. [Google Scholar] [CrossRef]
- Saraiva, A.; Donzele, J.L.; de Oliveira, R.F.M.; de Abreu, M.L.T.; Silva, F.C.O.; Haese, D. Available phosphorus levels in diets for swine with high genetic potential for meat deposition from 30 to 60 kg. Braz. J. Anim. Sci. 2009, 38, 1279–1285. [Google Scholar] [CrossRef]
- Pallauf, J.; Rimbach, G. Nutritional significance of phytic acid and phytase. Arch. Tierernahr. 1997, 50, 301–319. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.S.; De Boeck, G.; Becker, K. Phytate and phytase in fish nutrition. J. Anim. Physiol. Anim. Nutr. 2012, 96, 335–364. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Satyanarayana, T. Fungal phytases: Characteristics and amelioration of nutritional quality and growth of non-ruminants. J. Anim. Physiol. Anim. Nutr. 2015, 99, 646–660. [Google Scholar] [CrossRef]
- Singh, M.; Krikorian, A.D. Inhibition of trypsin activity in vitro by phytate. J. Agric. Food Chem. 1982, 30, 799–800. [Google Scholar] [CrossRef]
- Deshpande, S.S.; Cheryan, M. Effects of Phytic Acid, Divalent Cations, and Their Interactions on α-Amylase Activity. J. Food Sci. 1984, 49, 516–519. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V. Phytate-degrading enzymes in pig nutrition. Livest. Sci. 2008, 113, 99–122. [Google Scholar] [CrossRef]
- Lei, X.G.; Porres, J.M. Phytase enzymology, applications, and biotechnology. Biotechnol. Lett. 2003, 25, 1787–1794. [Google Scholar] [CrossRef]
- Moita, V.H.C.; Kim, S.W. Nutritional and Functional Roles of Phytase and Xylanase Enhancing the Intestinal Health and Growth of Nursery Pigs and Broiler Chickens. Animals 2022, 12, 3322. [Google Scholar] [CrossRef]
- Cowieson, A.J.; Ruckebusch, J.P.; Knap, I.; Guggenbuhl, P.; Fru-Nji, F. Phytate-free nutrition: A new paradigm in monogastric animal production. Anim. Feed Sci. Technol. 2016, 222, 180–189. [Google Scholar] [CrossRef]
- Arredondo, M.A.; Casas, G.A.; Stein, H.H. Increasing levels of microbial phytase increases the digestibility of energy and minerals in diets fed to pigs. Anim. Feed Sci. Technol. 2019, 248, 27–36. [Google Scholar] [CrossRef]
- Zeng, Z.K.; Wang, D.; Piao, X.S.; Li, P.F.; Zhang, H.Y.; Shi, C.X.; Yu, S.K. Effects of Adding Super Dose Phytase to the Phosphorus-deficient Diets of Young Pigs on Growth Performance, Bone Quality, Minerals and Amino Acids Digestibilities. Asian-Australas. J. Anim. Sci. 2014, 27, 237–246. [Google Scholar] [CrossRef]
- Zouaoui, M.; Létourneau-Montminy, M.P.; Guay, F. Effect of phytase on amino acid digestibility in pig: A meta-analysis. Anim. Feed Sci. Technol. 2018, 238, 18–28. [Google Scholar] [CrossRef]
- Moran, K.; Wilcock, P.; Elsbernd, A.; Zier-Rush, C.; Boyd, R.D.; van Heugten, E. Effects of super-dosing phytase and inositol on growth performance and blood metabolites of weaned pigs housed under commercial conditions. J. Anim. Sci. 2019, 97, 3007–3015. [Google Scholar] [CrossRef] [PubMed]
- Gehring, C.K.; Bedford, M.R.; Dozier, W.A. Extra-phosphoric effects of phytase with and without xylanase in corn-soybean meal-based diets fed to broilers. Poult. Sci. 2013, 92, 979–991. [Google Scholar] [CrossRef]
- Lu, H.; Cowieson, A.J.; Wilson, J.W.; Ajuwon, K.M.; Adeola, O. Extra-phosphoric effects of super dosing phytase on growth performance of pigs is not solely due to release of myo-inositol. J. Anim. Sci. 2019, 97, 3898–3906. [Google Scholar] [CrossRef]
- Zhang, L.H.; Liu, H.S.; Liu, S.J.; Piao, X.S. Dietary supplementation with 25-hydroxycholecalciferol and phytase in growing-finishing pigs: II. Effects on intestinal antioxidant status, immunity and bone quality. Anim. Feed Sci. Technol. 2021, 280, 115065. [Google Scholar] [CrossRef]
- Adedokun, S.A.; Adeola, O. The response in jejunal and ileal nutrient and energy digestibility and the expression of markers of intestinal inflammation in broiler chickens to coccidial vaccine challenge and phytase supplementation. Can. J. Anim. Sci. 2017, 97, 258–267. [Google Scholar] [CrossRef]
- Derakhshan, M.; Ghasemian, S.O.; Gholami-Ahangaran, M. The effects of probiotic and phytase on growth performance, biochemical parameters and antioxidant capacity in broiler chickens. Vet. Med. Sci. 2023, 9, 860–866. [Google Scholar] [CrossRef]
- Ren, P.; Chen, J.; Wedekind, K.; Hancock, D.; Vázquez-Añón, M. Interactive effects of zinc and copper sources and phytase on growth performance, mineral digestibility, bone mineral concentrations, oxidative status, and gut morphology in nursery pigs. Transl. Anim. Sci. 2020, 4, 783–798. [Google Scholar] [CrossRef]
- Amiri, M.Y.A.; Jafari, M.A.; Irani, M. Growth performance, internal organ traits, intestinal morphology, and microbial population of broiler chickens fed quinoa seed–based diets with phytase or protease supplements and their combination. Trop. Anim. Health Prod. 2021, 53, 535. [Google Scholar] [CrossRef]
- Moita, V.H.C.; Duarte, M.E.; Kim, S.W. Supplemental Effects of Phytase on Modulation of Mucosa-Associated Microbiota in the Jejunum and the Impacts on Nutrient Digestibility, Intestinal Morphology, and Bone Parameters in Broiler Chickens. Animals 2021, 11, 3351. [Google Scholar] [CrossRef] [PubMed]
- Zárate, G.; Ambrosini, V.M.D.; Chaia, A.P.; González, S. Some factors affecting the adherence of probiotic Propionibacterium acidipropionici CRL 1198 to intestinal epithelial cells. Can. J. Microbiol. 2002, 48, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Ru, Y.J.; Li, F.D. Effect of dietary phytate and phytase on metabolic change of blood and intestinal mucosa in chickens. J. Anim. Physiol. Anim. Nutr. 2010, 94, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Lallès, J.-P. Intestinal alkaline phosphatase: Multiple biological roles in maintenance of intestinal homeostasis and modulation by diet. Nutr. Rev. 2010, 68, 323–332. [Google Scholar] [CrossRef]
- Genova, L.J.; Melo, B.A.D.; Rupolo, E.P.; Carvalho, T.S.; Costa, B.L.; Carvalho, O.P.L. A summary of feed additives, intestinal health and intestinal alkaline phosphatase in piglet nutrition. Czech J. Anim. Sci. 2020, 65, 281–294. [Google Scholar] [CrossRef]
- Ptak, A.; Bedford, M.R.; Świątkiewicz, S.; Żyła, K.; Józefiak, D. Phytase Modulates Ileal Microbiota and Enhances Growth Performance of the Broiler Chickens. PLoS ONE 2015, 10, e0119770. [Google Scholar] [CrossRef]
- Gupta, R.; Mahmood, S.; Adeolu, M. A phylogenomic and molecular signature based approach for characterization of the phylum Spirochaetes and its major clades: Proposal for a taxonomic revision of the phylum. Front. Microbiol. 2013, 4, 217. [Google Scholar] [CrossRef]
- Moita, V.H.C.; Kim, S.W. Efficacy of a bacterial 6-phytase supplemented beyond traditional dose levels on jejunal mucosa-associated microbiota, ileal nutrient digestibility, bone parameters, and intestinal health, and growth performance of nursery pigs. J. Anim. Sci. 2023, 101, skad134. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M. Erratum: Enterotypes of the human gut microbiome. Nature 2011, 474, 666. [Google Scholar] [CrossRef]
- Sauer, K.; Cooke, M.P. Regulation of immune cell development through soluble inositol-1,3,4,5-tetrakisphosphate. Nat. Rev. Immunol. 2010, 10, 257–271. [Google Scholar] [CrossRef]
- Gonzalez-Uarquin, F.; Rodehutscord, M.; Huber, K. Myo-inositol: Its metabolism and potential implications for poultry nutrition—A review. Poult. Sci. 2020, 99, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Cowieson, A.J.; Roos, F.F.; Ruckebusch, J.-P.; Wilson, J.W.; Guggenbuhl, P.; Lu, H.; Ajuwon, K.M.; Adeola, O. Time-series responses of swine plasma metabolites to ingestion of diets containing myo-inositol or phytase. Br. J. Nutr. 2017, 118, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Kühn, I.; Bedford, M.R.; Whitfield, H.; Brearley, C.; Adeola, O.; Ajuwon, K.M. Effect of phytase on intestinal phytate breakdown, plasma inositol concentrations, and glucose transporter type 4 abundance in muscle membranes of weanling pigs. J. Anim. Sci. 2019, 97, 3907–3919. [Google Scholar] [CrossRef] [PubMed]
- Ajuwon, K.M.; Sommerfeld, V.; Paul, V.; Däuber, M.; Schollenberger, M.; Kühn, I.; Adeola, O.; Rodehutscord, M. Phytase dosing affects phytate degradation and Muc2 transporter gene expression in broiler starters. Poult. Sci. 2020, 99, 981–991. [Google Scholar] [CrossRef]
- Sens, R.F.; Bassi, L.S.; Almeida, L.M.; Rosso, D.F.; Teixeira, L.V.; Maiorka, A. Effect of different doses of phytase and protein content of soybean meal on growth performance, nutrient digestibility, and bone characteristics of broilers. Poult. Sci. 2021, 100, 100917. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Shin, S.; Kuehn, I.; Bedford, M.; Rodehutscord, M.; Adeola, O.; Ajuwon, K.M. Effect of phytase on nutrient digestibility and expression of intestinal tight junction and nutrient transporter genes in pigs. J. Anim. Sci. 2020, 98, skaa206. [Google Scholar] [CrossRef]
- Khodambashi Emami, N.; Zafari Naeini, S.; Ruiz-Feria, C.A. Growth performance, digestibility, immune response and intestinal morphology of male broilers fed phosphorus deficient diets supplemented with microbial phytase and organic acids. Livest. Sci. 2013, 157, 506–513. [Google Scholar] [CrossRef]
- Kipper, M.; Andretta, I.; Lehnen, C.R.; Lovatto, P.A.; Monteiro, S.G. Meta-analysis of the performance variation in broilers experimentally challenged by Eimeria spp. Vet. Parasitol. 2013, 196, 77–84. [Google Scholar] [CrossRef]
- Pastorelli, H.; van Milgen, J.; Lovatto, P.; Montagne, L. Meta-analysis of feed intake and growth responses of growing pigs after a sanitary challenge. Animal 2012, 6, 952–961. [Google Scholar] [CrossRef]
- Campos, P.H.R.F.; Labussière, E.; Hernández-García, J.; Dubois, S.; Renaudeau, D.; Noblet, J. Effects of ambient temperature on energy and nitrogen utilization in lipopolysaccharide-challenged growing pigs. J. Anim. Sci. 2014, 92, 4909–4920. [Google Scholar] [CrossRef]
- Le Floc’h, N.; Melchior, D.; Obled, C. Modifications of protein and amino acid metabolism during inflammation and immune system activation. Livest. Prod. Sci. 2004, 87, 37–45. [Google Scholar] [CrossRef]
- Gabler, N.K.; Spurlock, M.E. Integrating the immune system with the regulation of growth and efficiency. J. Anim. Sci. 2008, 86, E64–E74. [Google Scholar] [CrossRef] [PubMed]
- Dersjant-Li, Y.; Gibbs, K.; Awati, A.; Klasing, K.C. The effects of enzymes and direct fed microbial combination on performance and immune response of broilers under a coccidia challenge. J. Appl. Anim. Nutr. 2016, 4, e6. [Google Scholar] [CrossRef]
- Dunaway, A.; Adedokun, S.A. Coccidia Vaccine Challenge and Exogenous Enzyme Supplementation in Broiler Chicken 1. Effect on Digesta Viscosity, Diet Energy Utilization, and Apparent Metabolizable Energy Value of Wheat. Animals 2021, 11, 641. [Google Scholar] [CrossRef] [PubMed]
- Pastorelli, H.; Le Floc’h, N.; Merlot, E.; Meunier-Salaün, M.C.; van Milgen, J.; Montagne, L. Feed restriction applied after weaning has different effects on pig performance and health depending on the sanitary conditions. J. Anim. Sci. 2012, 90, 4866–4875. [Google Scholar] [CrossRef] [PubMed]
- Barekatain, M.R.; Antipatis, C.; Rodgers, N.; Walkden-Brown, S.W.; Iji, P.A.; Choct, M. Evaluation of high dietary inclusion of distillers dried grains with solubles and supplementation of protease and xylanase in the diets of broiler chickens under necrotic enteritis challenge. Poult. Sci. 2013, 92, 1579–1594. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.D.; Khattak, F.; Hastie, P.; Bedford, M.R.; Olukosi, O.A. The similarity of the effect of carbohydrase or prebiotic supplementation in broilers aged 21 days, fed mixed cereal diets and challenged with coccidiosis infection. PLoS ONE 2020, 15, e0229281. [Google Scholar] [CrossRef]
- Jia, W.; Slominski, B.A.; Bruce, H.L.; Blank, G.; Crow, G.; Jones, O. Effects of diet type and enzyme addition on growth performance and gut health of broiler chickens during subclinical Clostridium perfringens challenge. Poult. Sci. 2009, 88, 132–140. [Google Scholar] [CrossRef]
- Kim, E.; Moss, A.F.; Morgan, N.K.; Gharib-Naseri, K.; Ader, P.; Choct, M. Non-starch polysaccharide-degrading enzymes may improve performance when included in wheat- but not maize-based diets fed to broiler chickens under subclinical necrotic enteritis challenge. Anim. Nutr. 2022, 10, 54–67. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, B.; Rousseau, X.; Gomes, G.A.; Oh, H.J.; Kim, Y.J.; Chang, S.Y.; An, J.W.; Go, Y.B.; Song, D.C.; et al. Stimbiotic supplementation modulated intestinal inflammatory response and improved broilers performance in an experimentally-induced necrotic enteritis infection model. J. Anim. Sci. Biotechnol. 2022, 13, 100. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lourenco, J.M.; Olukosi, O.A. The effects of protease, xylanase, and xylo-oligosaccharides on growth performance, nutrient utilization, short-chain fatty acids, and microbiota in Eimeria-challenged broiler chickens fed low protein diet. Poult. Sci. 2023, 102, 102789. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Liu, D.; Guo, S.; Chen, Y.; Guo, Y. Effects of dietary essential oil and enzyme supplementation on growth performance and gut health of broilers challenged by Clostridium perfringens. Anim. Feed Sci. Technol. 2015, 207, 234–244. [Google Scholar] [CrossRef]
- Watson, B.C.; Matthews, J.O.; Southern, L.L.; Shelton, J.L. The interactive effects of Eimeria acervulina infection and phytase for broiler chicks. Poult. Sci. 2005, 84, 910–913. [Google Scholar] [CrossRef] [PubMed]
- Zanu, H.K.; Kheravii, S.K.; Morgan, N.K.; Bedford, M.R.; Swick, R.A. Over-processed meat and bone meal and phytase effects on broilers challenged with subclinical necrotic enteritis: Part 1. Performance, intestinal lesions and pH, bacterial counts and apparent ileal digestibility. Anim. Nutr. 2020, 6, 313–324. [Google Scholar] [CrossRef]
- Zanu, H.K.; Kheravii, S.K.; Morgan, N.K.; Bedford, M.R.; Swick, R.A. Interactive effect of 2 dietary calcium and phytase levels on broilers challenged with subclinical necrotic enteritis: Part 1—Broiler performance, gut lesions and pH, bacterial counts, and apparent ileal digestibility. Poult. Sci. 2020, 99, 4861–4873. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.S.; Velayudhan, D.E.; Li, A.; Feng, Z.; Liu, D.; Yin, Y.L.; Nyachoti, C.M. Growth performance, gastrointestinal microbial activity, and immunological response of piglets receiving microencapsulated Enterococcus faecalis CG1.0007 and enzyme complex after an oral challenge with Escherichia coli (K88). Can. J. Anim. Sci. 2016, 96, 609–618. [Google Scholar] [CrossRef]
- Li, Q.; Burrough, E.R.; Gabler, N.K.; Loving, C.L.; Sahin, O.; Gould, S.A.; Patience, J.F. A soluble and highly fermentable dietary fiber with carbohydrases improved gut barrier integrity markers and growth performance in F18 ETEC challenged pigs. J. Anim. Sci. 2019, 97, 2139–2153. [Google Scholar] [CrossRef]
- Song, D.; Lee, J.; Kwak, W.; Song, M.; Oh, H.; Kim, Y.; An, J.; Chang, S.; Go, Y.; Cho, H.; et al. Stimbiotic Supplementation Alleviates Poor Performance and Gut Integrity in Weaned Piglets Induced by Challenge with E. coli. Animals 2022, 12, 1799. [Google Scholar] [CrossRef]
- Waititu, S.M.; Yin, F.; Patterson, R.; Rodriguez-Lecompte, J.C.; Nyachoti, C.M. Short-term effect of supplemental yeast extract without or with feed enzymes on growth performance, immune status and gut structure of weaned pigs challenged with Escherichia coli lipopolysaccharide. J. Anim. Sci. Biotechnol. 2016, 7, 64. [Google Scholar] [CrossRef]
- Kiarie, E.G.; Slominski, B.A.; Krause, D.O.; Nyachoti, C.M. Nonstarch Polysaccharide Hydrolysis Products of Soybean and Canola Meal Protect against Enterotoxigenic Escherichia coli in Piglets. J. Nutr. 2008, 138, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Slominski, B.A.; Bruce, H.L.; Nyachoti, C.M.; Jones, R.O. Enzyme addition facilitates the post-disease compensatory growth of broiler chickens challenged with Clostridium perfringens. Can. J. Anim. Sci. 2009, 89, 369–381. [Google Scholar] [CrossRef]
- Moran, K.; Boyd, R.D.; Zier-Rush, C.; Wilcock, P.; Bajjalieh, N.; van Heugten, E. Effects of high inclusion of soybean meal and a phytase superdose on growth performance of weaned pigs housed under the rigors of commercial conditions. J. Anim. Sci. 2017, 95, 5455–5465. [Google Scholar] [CrossRef]
- Bedford, M.R.; Cowieson, A.J. Matrix values for exogenous enzymes and their application in the real world. J. Appl. Poult. Res. 2020, 29, 15–22. [Google Scholar] [CrossRef]
- Metwally, A.E.; Abdel-Wareth, A.A.A.; Saleh, A.A.; Amer, S.A. Are the energy matrix values of the different feed additives in broiler chicken diets could be summed? BMC Vet. Res. 2020, 16, 391. [Google Scholar] [CrossRef]
- Klein, J.; Williams, M.; Brown, B.; Rao, S.; Lee, J.T. Effects of dietary inclusion of a cocktail NSPase and β-mannanase separately and in combination in low energy diets on broiler performance and processing parameters. J. Appl. Poult. Res. 2015, 24, 489–501. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, G.-M.; Wang, W.-W.; Liu, G.-H.; Cai, H.-Y.; Purba, A.; Zheng, A.-J. Compound non-starch polysaccharide enzymes improve growth performance, slaughter performance, immune function, and apparent utilization rate of nutrients in broiler chickens fed a low-metabolizable energy diet. Front. Vet. Sci. 2023, 10, 1162811. [Google Scholar] [CrossRef]
- Vieira, S.L.; Stefanello, C.; Rios, H.V.; Serafini, N.; Hermes, R.G.I.; Sorbara, J.O.B. Efficacy and Metabolizable Energy Equivalence of an α-Amylase-β-Glucanase Complex for Broilers. Braz. J. Poult. Sci. 2015, 17, 227–236. [Google Scholar] [CrossRef]
- Attia, Y.A.; Bovera, F.; Al-Harthi, M.A.; El-Din, A.E.-R.E.T.; Said Selim, W. Supplementation of Microbial and Fungal Phytases to Low Protein and Energy Diets: Effects on Productive Performance, Nutrient Digestibility, and Blood Profiles of Broilers. Agriculture 2021, 11, 414. [Google Scholar] [CrossRef]
- Jang, Y.D.; Wilcock, P.; Boyd, R.D.; Lindemann, M.D. Effect of combined xylanase and phytase on growth performance, apparent total tract digestibility, and carcass characteristics in growing pigs fed corn-based diets containing high-fiber coproducts. J. Anim. Sci. 2017, 95, 4005–4017. [Google Scholar] [CrossRef]
- de França, T.P.; Ferreira, R.d.S.; Leo, R.A.R.; de Oliveira, C.H.; Dias, K.M.M.; Gomes, K.M.; Costa, L.S.; Albino, L.F.T. Effects of carbohydrase and phytase enzymes supplementation within low energy diets on performance and energy utilization of broiler chickens. Livest. Sci. 2023, 274, 105271. [Google Scholar] [CrossRef]
- Nusairat, B.; Wang, J.-J. The Effect of a Modified GH11 Xylanase on Live Performance, Gut Health, and Clostridium perfringens Excretion of Broilers Fed Corn-Soy Diets. Front. Vet. Sci. 2021, 8, 678536. [Google Scholar] [CrossRef] [PubMed]
- Bello, A.; Dersjant-Li, Y.; van Eerden, E.; Kwakernaak, C.; Marchal, L. Supplementation of an all-plant-based inorganic phosphate-free diet with a novel phytase maintained tibia ash and performance in broilers under a commercial production setting. J. Appl. Poult. Res. 2022, 31, 100253. [Google Scholar] [CrossRef]
- Ennis, C.E.; Jackson, M.; Gutierrez, O.; Cantley, S.; Wamsley, K.G.S. Phytase and carbohydrase inclusion strategies to explore synergy within low-energy diets to optimize 56-day male broiler performance and processing. J. Appl. Poult. Res. 2020, 29, 1045–1067. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; van de Belt, K.; van der Klis, J.D.; Kettunen, H.; Rinttilä, T.; Awati, A. Effect of multi-enzymes in combination with a direct-fed microbial on performance and welfare parameters in broilers under commercial production settings. J. Appl. Poult. Res. 2015, 24, 80–90. [Google Scholar] [CrossRef]
- Bavaresco, C.; Krabbe, E.; Gopinger, E.; Sandi, A.J.; Martinez, F.N.; Wernik, B.; Roll, V.F.B. Hybrid Phytase and Carbohydrases in Corn and Soybean Meal-Based Diets for Broiler Chickens: Performance and Production Costs. Braz. J. Poult. Sci. 2020, 22, 1–8. [Google Scholar] [CrossRef]
- Ribeiro, T.; Dal Pont, G.; Dahlke, F.; da Rocha, C.; Sorbara, J.O.; Maiorka, A. Available Phosphorus and Calcium Reduction in the Finisher Phase and Phytase Utilization on Broilers. J. Appl. Poult. Res. 2019, 28, 263–270. [Google Scholar] [CrossRef]
- da Silva, C.A.; Callegari, M.A.; Dias, C.P.; Bridi, A.M.; Pierozan, C.R.; Foppa, L.; Martins, C.C.d.S.; Dias, F.T.F.; Passos, A.; Hermes, R. Increasing doses of phytase from Citrobacter braakii in diets with reduced inorganic phosphorus and calcium improve growth performance and lean meat of growing and finishing pigs. PLoS ONE 2019, 14, e0217490. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Van de Belt, K.; Kwakernaak, C.; Marchal, L. Buttiauxella phytase maintains growth performance in broilers fed diets with reduced nutrients under a commercial setting. J. Appl. Anim. Nutr. 2020, 8, 49–59. [Google Scholar] [CrossRef]
- Cambra-López, M.; Cerisuelo, A.; Ferrer, P.; Ródenas, L.; Aligué, R.; Moset, V.; Pascual, J.J. Age influence on effectiveness of a novel 3-phytase in barley-wheat based diets for pigs from 12 to 108 kg under commercial conditions. Anim. Feed Sci. Technol. 2020, 267, 114549. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Plumstead, P.; Awati, A.; Remus, J. Productive performance of commercial growing and finishing pigs supplemented with a Buttiauxella phytase as a total replacement of inorganic phosphate. Anim. Nutr. 2018, 4, 351–357. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| References | Species | Diet Type | Evaluation Period | Enzymes | Challenge Type |
|---|---|---|---|---|---|
| [112] | Broiler | Corn/SBM | 49 to 55 d | Phytase 1000 FTU | Coccidial vaccine |
| [112] | Broiler | Corn/SBM | 49 to 55 d | Phytase 5000 FTU | Coccidial vaccine |
| [141] | Broiler | Wheat/Barley/DDGS | 1 to 35 d | Xylanase + protease | Necrotic enteritis |
| [142] | Broiler | Wheat/Corn/Barley/SBM | 14 to 21 d | Xylanase | Coccidiosis vaccine |
| [142] | Broiler | Wheat/Corn/Barley/SBM | 14 to 21 d | Xylanase + β-glucanase | Coccidiosis vaccine |
| [138] | Broiler | Corn/Wheat/Rye/SBM | 1 to 21 d | Xylanase + amylase + protease + Bacillus spp. | Oral coccidia |
| [143] | Broiler | Corn/SBM | 1 to 40 d | Cellulase + pectinase + xylanase + glucanase + mannanase + galactanase | Necrotic enteritis |
| [143] | Broiler | Wheat/SBM | 1 to 40 d | Cellulase + pectinase + xylanase + glucanase + mannanase + galactanase | Necrotic enteritis |
| [144] | Broiler | Wheat/SBM | 1 to 21 d | Xylanase | Necrotic enteritis |
| [144] | Broiler | Maize/SBM | 1 to 21 d | Xylanase | Necrotic enteritis |
| [144] | Broiler | Wheat/SBM | 1 to 21 d | β-Mannanase | Necrotic enteritis |
| [144] | Broiler | Maize/SBM | 1 to 21 d | β-Mannanase | Necrotic enteritis |
| [145] | Broiler | Corn/Wheat/SBM | 1 to30 d | Xylanase + xylo-oligosaccharides | Necrotic enteritis |
| [146] | Broiler | Corn/SBM | 1 to 21 d | Xylanase + protease | Coccidiosis |
| [16] | Broiler | Corn/SBM | 1 to 42 d | β-Mannanase | Coccidiosis |
| [147] | Broiler | Wheat/SBM | 1 to 35 d | Xylanase + β-glucanase + β-Mannanase | Necrotic enteritis |
| [148] | Broiler | Corn/SBM (adequate Ca and P) | 5 to 15 d | Phytase 600 FTU | Coccidiosis |
| [148] | Broiler | Corn/SBM (lower Ca and P) | 5 to 15 d | Phytase 600 FTU | Coccidiosis |
| [149] | Broiler | Wheat/SBM | 1 to 28 d | Phytase 5000 FTU | Necrotic enteritis |
| [150] | Broiler | Wheat/SBM (high Ca) | 1 to 42 d | Phytase 1500 FTU | Necrotic enteritis |
| [150] | Broiler | Wheat/SBM (low Ca) | 1 to 42 d | Phytase 1500 FTU | Necrotic enteritis |
| [151] | Pig | Barley/Wheat/SBM | 21 to 35 d | Carbohydrases | E. coli (K88) |
| [26] | Pig | Wheat/Corn/SBM | 21 to 63 d | Xylanase + xylo-oligosaccharides | Poor sanitary condition |
| [45] | Pig | Corn/SBM/DDGS | 21 to 41 d | Xylanase + Bacillus sp. | E. coli (F18+) |
| [152] | Pig | Corn/SBM/DDGS | 23 to 37 d | Xylanase + β-glucanase + pectinase | E. coli (F18+) |
| [152] | Pig | Corn/SBM/Sugar beet pulp | 23 to 37 d | Xylanase + β-glucanase + pectinase | E. coli (F18+) |
| [153] | Pig | Corn/SBM | 28 to 42 d | Xylanase + xylo-oligosaccharides | E. coli |
| [154] | Pig | Corn/Wheat/SBM | 21 to 32 d | Cellulase + β-mannanase + galactanase + xylanase + β-glucanase + amylase + protease | E. coli (LPS) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valente Junior, D.T.; Genova, J.L.; Kim, S.W.; Saraiva, A.; Rocha, G.C. Carbohydrases and Phytase in Poultry and Pig Nutrition: A Review beyond the Nutrients and Energy Matrix. Animals 2024, 14, 226. https://doi.org/10.3390/ani14020226
Valente Junior DT, Genova JL, Kim SW, Saraiva A, Rocha GC. Carbohydrases and Phytase in Poultry and Pig Nutrition: A Review beyond the Nutrients and Energy Matrix. Animals. 2024; 14(2):226. https://doi.org/10.3390/ani14020226
Chicago/Turabian StyleValente Junior, Dante Teixeira, Jansller Luiz Genova, Sung Woo Kim, Alysson Saraiva, and Gabriel Cipriano Rocha. 2024. "Carbohydrases and Phytase in Poultry and Pig Nutrition: A Review beyond the Nutrients and Energy Matrix" Animals 14, no. 2: 226. https://doi.org/10.3390/ani14020226