
Evaluation of In Vitro Protein Hydrolysis in Seven Insects Approved by the EU for Use as a Protein Alternative in Aquaculture
 , , , and
, , , and
Abstract
:Simple Summary
Abstract

1. Introduction
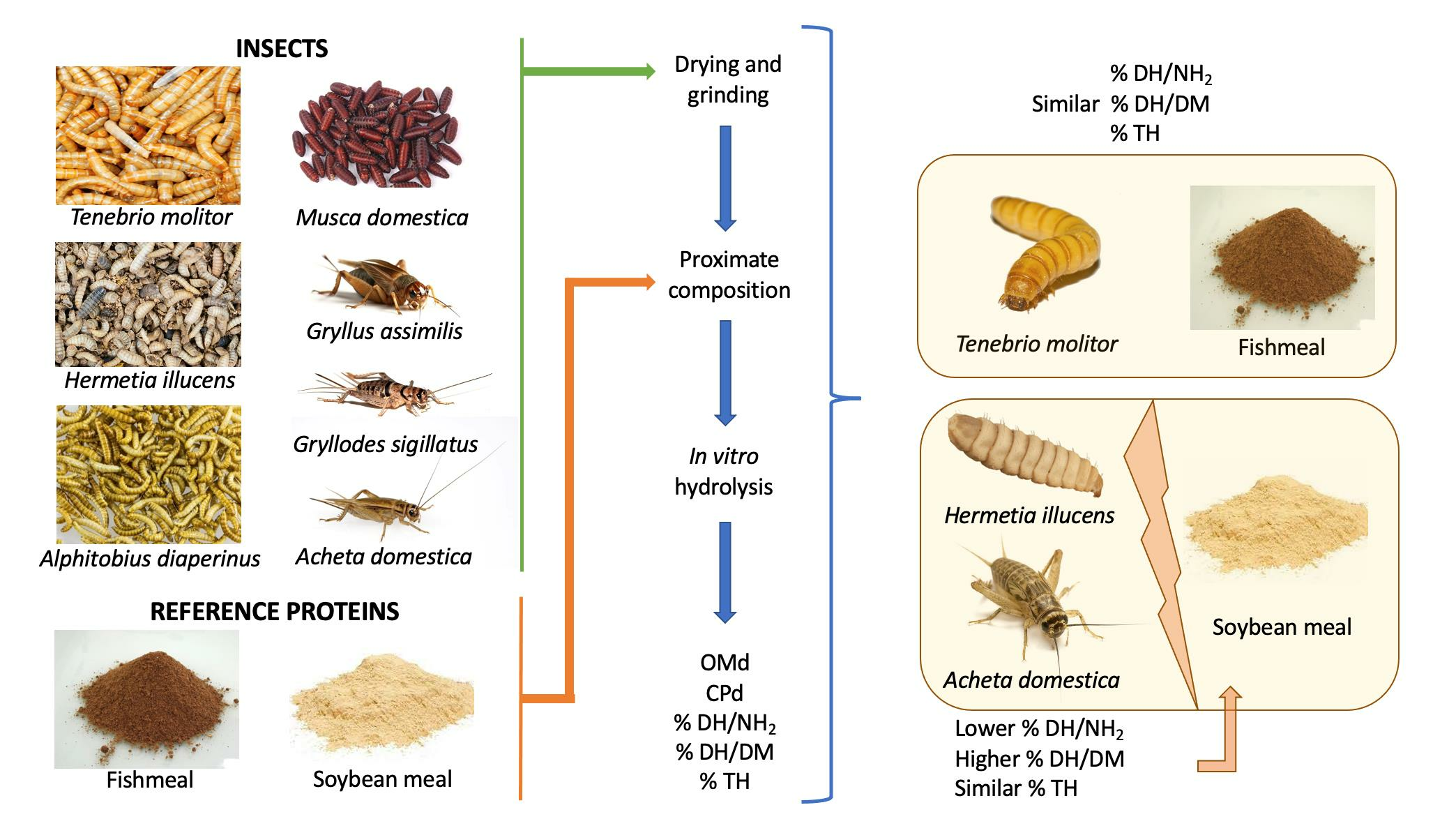
2. Materials and Methods
2.1. Proximate Composition
2.2. In Vitro Protein Hydrolysis
2.3. Organic Matter Digestibility (OMd)
2.4. Crude Protein Digestibility (CPd)
2.5. O-Phthaldialdehyde (OPA) Method
2.6. Total Hydrolysis (TH)
2.7. Statistical Analysis
3. Results
3.1. Proximate Composition
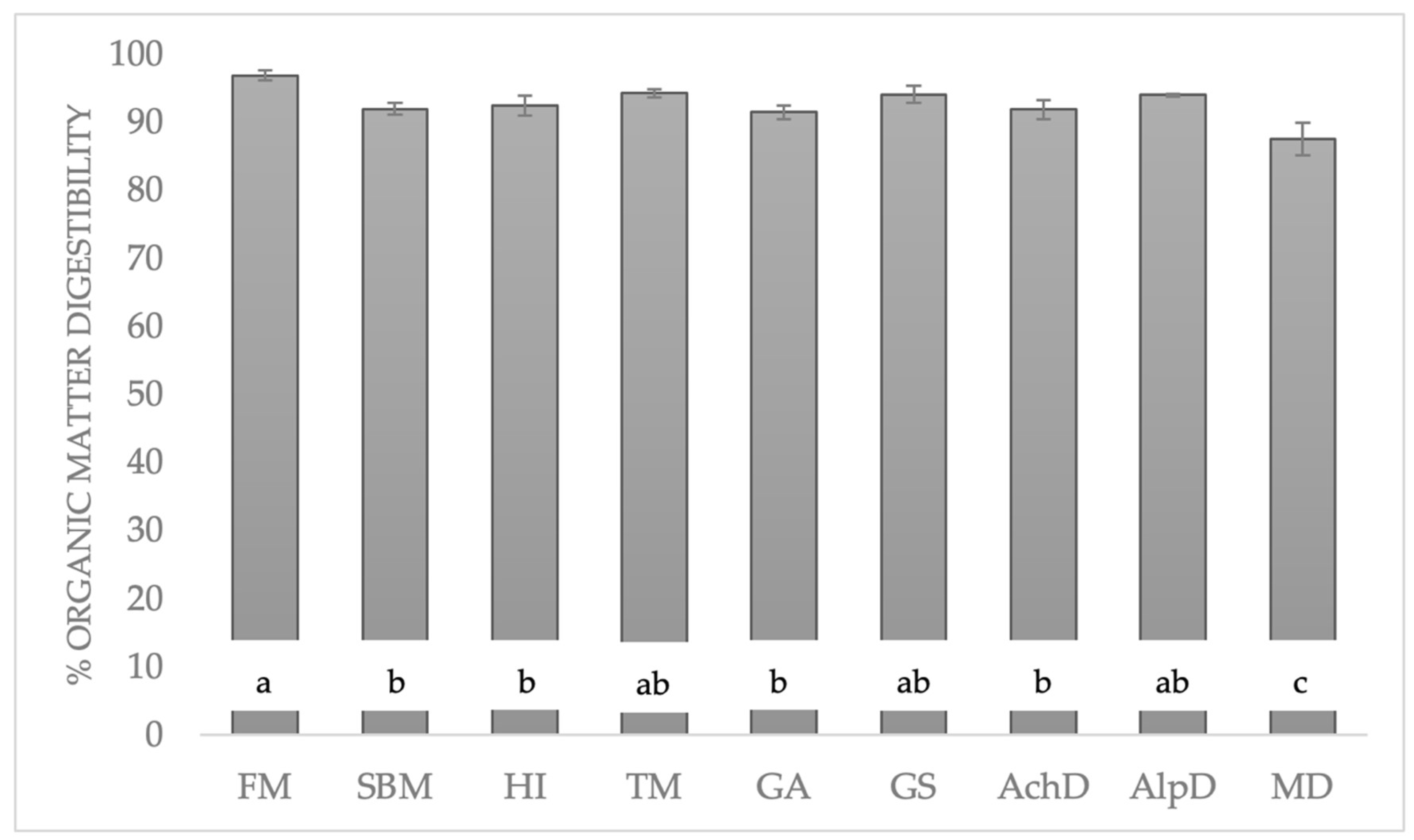
3.2. Organic Matter Digestibility (OMd)
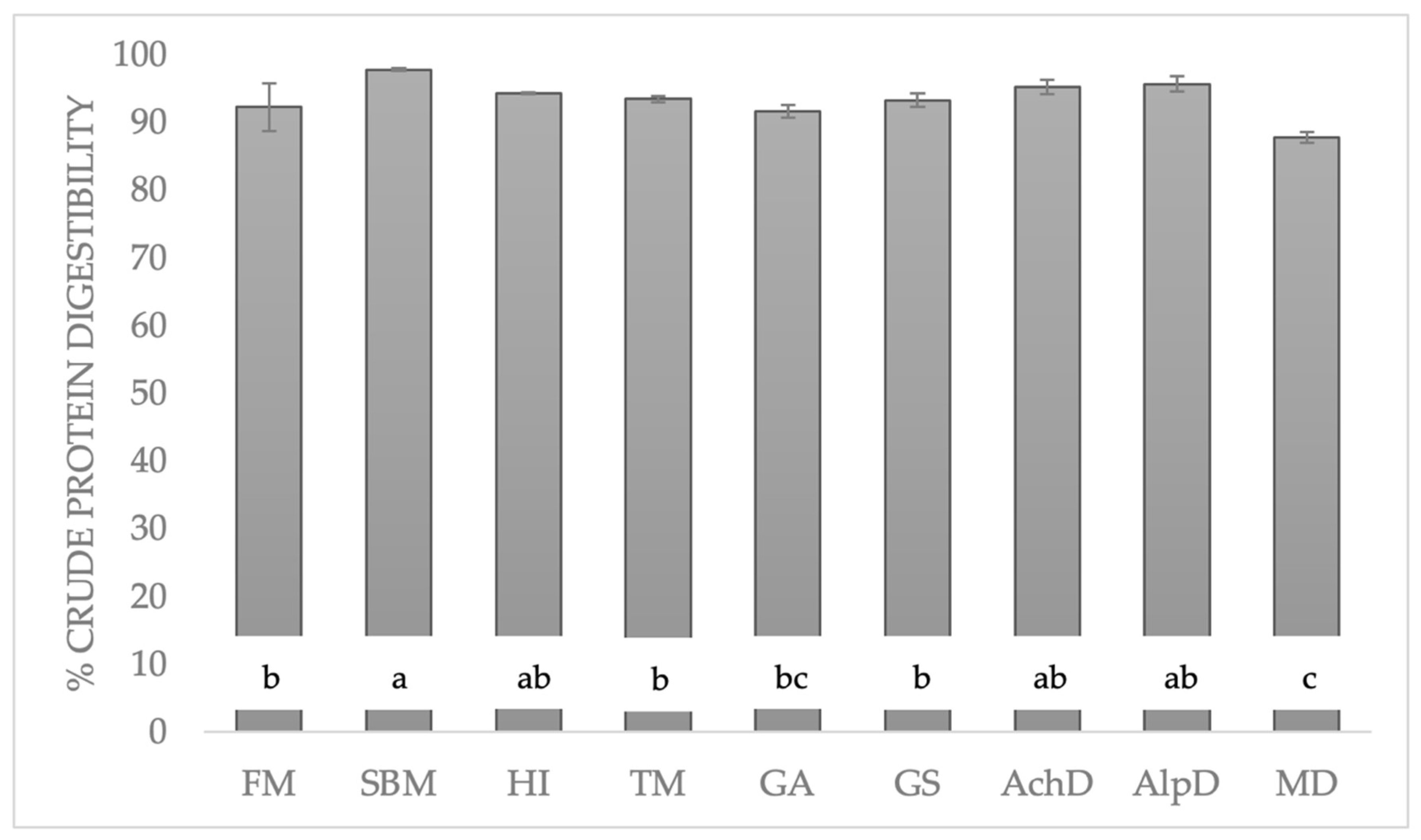
3.3. Crude Protein Digestibility (CPd)
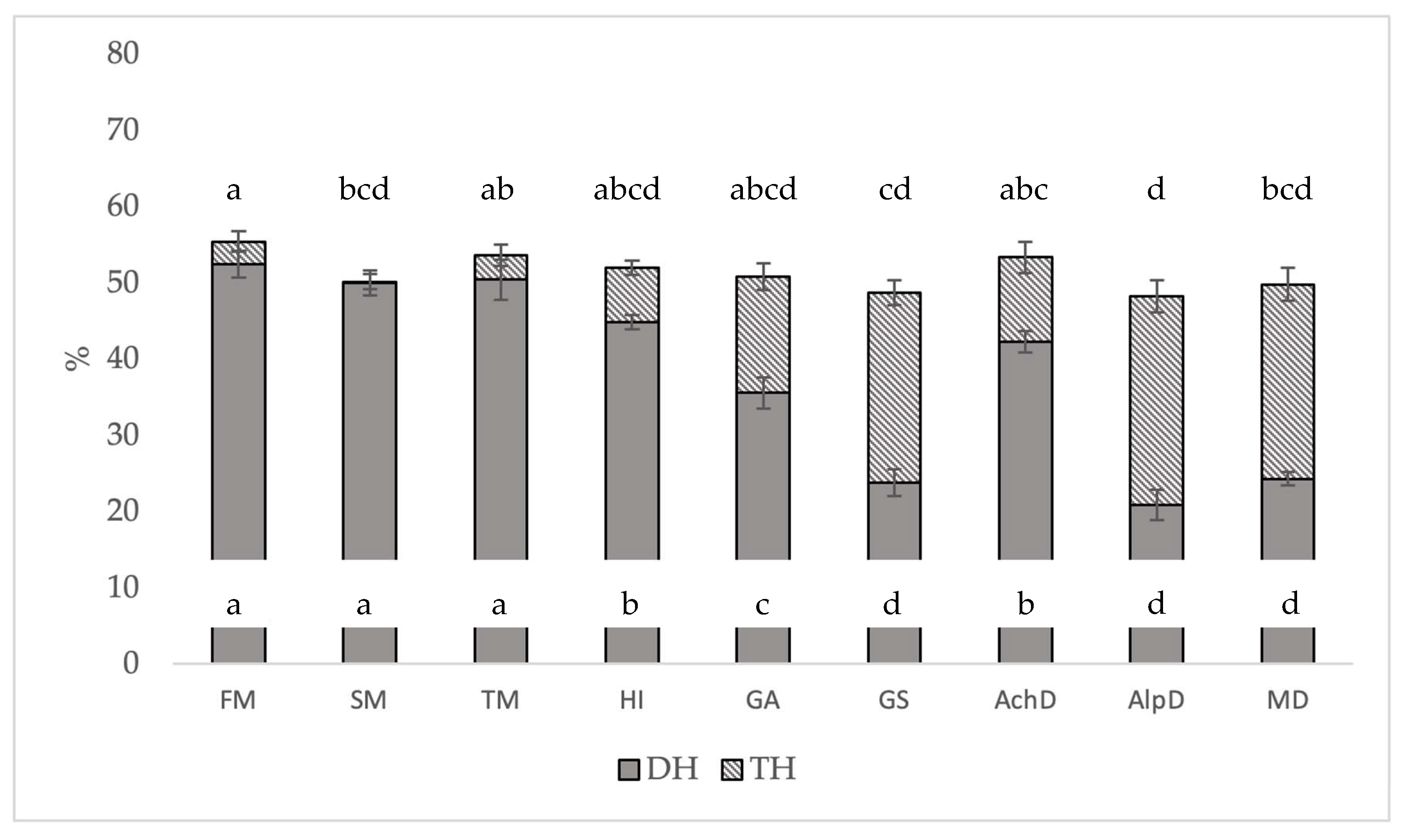
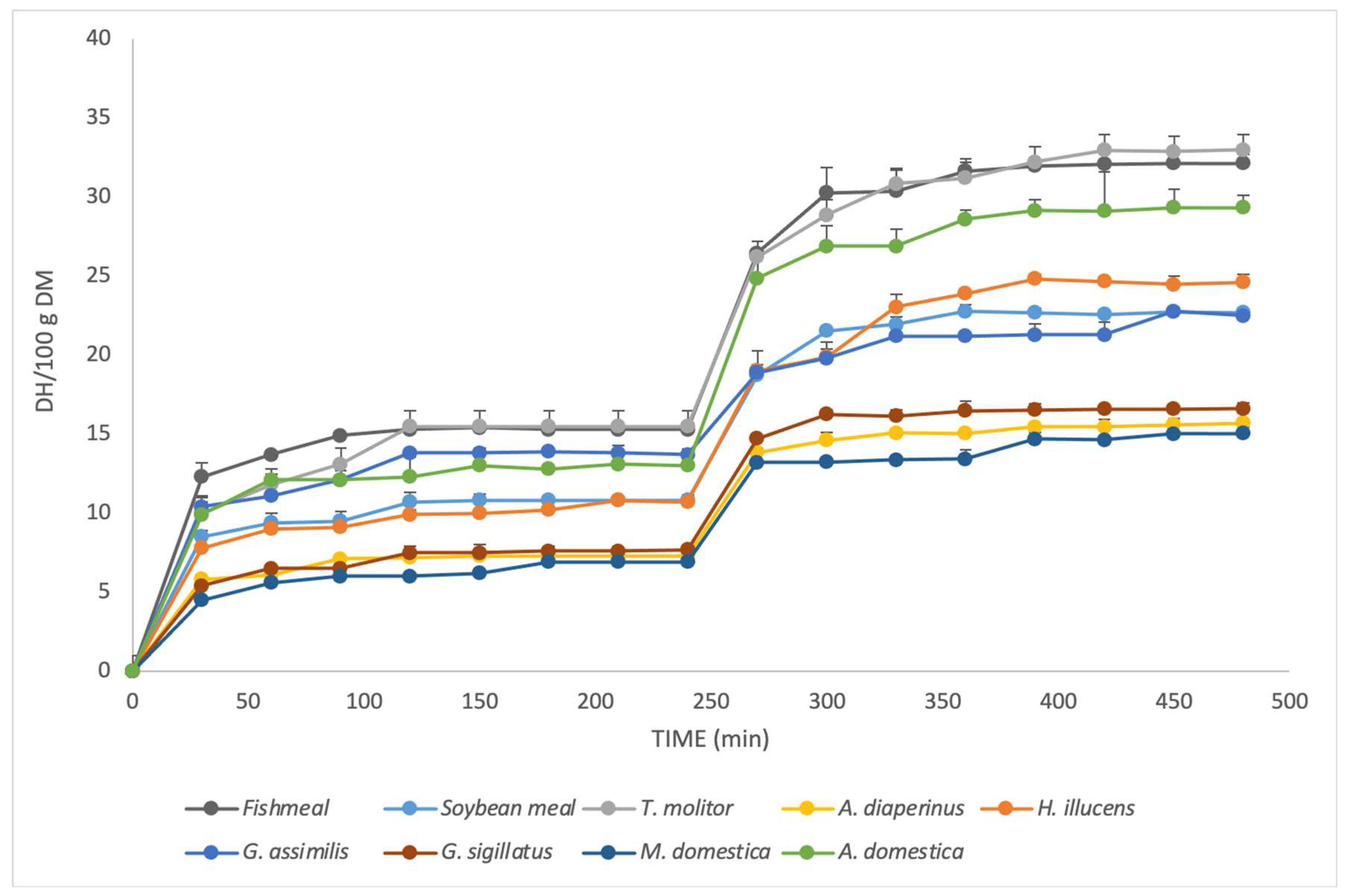
3.4. Degree of Hydrolysis (DH/NH2) and Total Hydrolysis (TH)
3.5. Gastric and Intestinal Digestion
3.6. Correlations
4. Discussion
4.1. Organic Matter Digestibility (OMd) and Crude Protein Digestibility (CPd)
4.2. Degree of Hydrolysis (DH) and Total Hydrolysis (TH)
4.3. Gastric and Intestinal Digestion
4.4. Correlations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Commission Regulation (EU) 2017/893 of 24 May 2017 Amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as Regards the Provisions on Processed Animal Protein. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32017R0893 (accessed on 22 July 2022).
- Commission Regulation (EU) 2021/1925 of 5 November 2021 Amending Certain Annexes to Regulation (EU) No 142/2011 as Regards the Requirements for Placing on the Market of Certain Insect Products and the Adaptation of a Containment Method. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32021R1925 (accessed on 22 July 2022).
- Dumas, A.; Raggi, T.; Barkhouse, J.; Lewis, E.; Weltzien, E. The oil fraction and partially defatted meal of black soldier fly larvae (Hermetia illucens) affect differently growth performance, feed efficiency, nutrient deposition, blood glucose and lipid digestibility of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 492, 24–34. [Google Scholar] [CrossRef]
- Fabrikov, D.; Sánchez-Muros, M.J.; Barroso, F.G.; Tomás-Almenar, C.; Melenchón, F.; Hidalgo, M.C.; Morales, A.E.; Rodriguez-Rodriguez, M.; Montes-Lopez, J. Comparative study of growth performance and amino acid catabolism in Oncorhynchus mykiss, Tinca tinca and Sparus aurata and the catabolic changes in response to insect meal inclusion in the diet. Aquaculture 2020, 529, 735731. [Google Scholar] [CrossRef]
- Kozłowski, K.; Ognik, K.; Stępniowska, A.; Juśkiewicz, J.; Zduńczyk, Z.; Kierończyk, B.; Benzertiha, A.; Józefiak, D.; Jankowski, J. Growth performance, immune status and intestinal fermentative processes of young turkeys fed diet with additive of full fat meals from Tenebrio molitor and Hermetia illucens. Anim. Feed Sci. Technol. 2021, 278, 114994. [Google Scholar] [CrossRef]
- Melenchón, F.; Larrán, A.M.; de Mercado, E.; Hidalgo, M.C.; Cardenete, G.; Barroso, F.G.; Fabrikov, D.; Lourenço, H.M.; Pessoa, M.F.; Tomás-Almenar, C. Potential use of black soldier fly (Hermetia illucens) and mealworm (Tenebrio molitor) insectmeals in diets for rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2021, 27, 491–505. [Google Scholar] [CrossRef]
- Gravel, A.; Doyen, A. The use of edible insect proteins in food: Challenges and issues related to their functional properties. Innov. Food Sci. Emerg. Technol. 2020, 59, 102272. [Google Scholar] [CrossRef]
- Raheem, D.; Raposo, A.; Oluwole, O.B.; Nieuwland, M.; Saraiva, A.; Carrascosa, C. Entomophagy: Nutritional, ecological, safety and legislation aspects. Food Res. Int. 2019, 126, 108672. [Google Scholar] [CrossRef] [PubMed]
- Sun-Waterhouse, D.; Waterhouse, G.I.N.; You, L.; Zhang, J.; Liu, Y.; Ma, L.; Gao, J.; Dong, Y. Transforming insect biomass into consumer wellness foods: A review. Food Res. Int. 2016, 89, 129–151. [Google Scholar] [CrossRef] [PubMed]
- Kouřimská, L.; Adámková, A. Nutritional and sensory quality of edible insects. NFS J. 2016, 4, 22–26. [Google Scholar] [CrossRef]
- de Castro, R.J.S.; Ohara, A.; dos Santos Aguilar, J.G.; Domingues, M.A.F. Nutritional, functional and biological properties of insect proteins: Processes for obtaining, consumption and future challenges. Trends Food Sci. Technol. 2018, 76, 82–89. [Google Scholar] [CrossRef]
- Gómez, B.; Munekata, P.E.S.; Zhu, Z.; Barba, F.J.; Toldrá, F.; Putnik, P.; Bursać Kovačević, D.; Lorenzo, J.M. Challenges and opportunities regarding the use of alternative protein sources: Aquaculture and insects. Adv. Food Nutr. Res. 2019, 89, 259–295. [Google Scholar] [CrossRef]
- Huang, C.; Feng, W.; Xiong, J.; Wang, T.; Wang, W.; Wang, C.; Yang, F. Impact of drying method on the nutritional value of the edible insect protein from black soldier fly (Hermetia illucens L.) larvae: Amino acid composition, nutritional value evaluation, in vitro digestibility, and thermal properties. Eur. Food Res. Technol. 2019, 245, 11–21. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.-J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Manditsera, F.A.; Luning, P.A.; Fogliano, V.; Lakemond, C.M.M. Effect of domestic cooking methods on protein digestibility and mineral bioaccessibility of wild harvested adult edible insects. Food Res. Int. 2019, 121, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Jo, K.; Yong, H.I.; Choi, Y.-S.; Jung, S. Comparison of the in vitro protein digestibility of Protaetia brevitarsis larvae and beef loin before and after defatting. Food Chem. 2021, 338, 128073. [Google Scholar] [CrossRef] [PubMed]
- Elia, A.C.; Capucchio, M.T.; Caldaroni, B.; Magara, G.; Dörr, A.J.M.; Biasato, I.; Biasibetti, E.; Righetti, M.; Pastorino, P.; Prearo, M.; et al. Influence of Hermetia illucens meal dietary inclusion on the histological traits, gut mucin composition and the oxidative stress biomarkers in rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 496, 50–57. [Google Scholar] [CrossRef]
- Azzollini, D.; Derossi, A.; Fogliano, V.; Lakemond, C.M.M.; Severini, C. Effects of formulation and process conditions on microstructure, texture and digestibility of extruded insect-riched snacks. Innov. Food Sci. Emerg. Technol. 2018, 45, 344–353. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists International, 18th ed.; AOAC International: Gaitherburg, MD, USA, 2005. [Google Scholar]
- ANKOM Technology (2006) Acid Detergent Fiber in Feeds Filter Bag Technique, Method 5, ANKOM Technology. Available online: http://www.ankom.com/media/documents/ADF_81606_A200.pdf (accessed on 21 November 2023).
- Ottoboni, M.; Spranghers, T.; Pinotti, L.; Baldi, A.; De Jaeghere, W.; Eeckhout, M. Inclusion of Hermetia illucens larvae or prepupae in an experimental extruded feed: Process optimisation and impact on in vitro digestibility. Ital. J. Anim. Sci. 2018, 17, 418–427. [Google Scholar] [CrossRef]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G. Spectrophotometric assay using o-phthaldehyde for determination of proteolysis in milk proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.-J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422–423, 193–201. [Google Scholar] [CrossRef]
- Mastoraki, M.; Panteli, N.; Kotzamanis, Y.P.; Gasco, L.; Antonopoulou, E.; Chatzifotis, S. Nutrient digestibility of diets containing five different insect meals in gilthead sea bream (Sparus aurata) and European sea bass (Dicentrarchus labrax). Anim. Feed Sci. Technol. 2022, 292, 115425. [Google Scholar] [CrossRef]
- Kovitvadhi, A.; Chundang, P.; Thongprajukaew, K.; Tirawattanawanich, C.; Srikachar, S.; Chotimanothum, B. Potential of insect meals as protein sources for meat-type ducks based on in vitro digestibility. Animals 2019, 9, 155. [Google Scholar] [CrossRef] [PubMed]
- Kovitvadhi, A.; Chundang, P.; Pliantiangtam, N.; Thongprajukaew, K.; Tirawattanawanich, C.; Suwanasopee, T.; Koonawootrittriron, S. Screening of in vitro nutrient digestibility coefficients of selected insect meals in broiler chickens, black-meat chickens and quails. J. Anim. Physiol. Anim. Nutr. 2021, 105, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Poelaert, C.; Beckers, Y.; Despret, X.; Portetelle, D.; Francis, F.; Bindelle, J. In vitro evaluation of fermentation characteristics of two types of insects as potential novel protein feeds for pigs1. J. Anim. Sci. 2016, 94 (Suppl. S3), 198–201. [Google Scholar] [CrossRef]
- Bosch, G.; Zhang, S.; Oonincx, D.G.A.B.; Hendriks, W.H. Protein quality of insects as potential ingredients for dog and cat foods. J. Nutr. Sci. 2014, 3, e29. [Google Scholar] [CrossRef]
- Bosch, G.; Vervoort, J.J.M.; Hendriks, W.H. In vitro digestibility and fermentability of selected insects for dog foods. Anim. Feed Sci. Technol. 2016, 221, 174–184. [Google Scholar] [CrossRef]
- Ndiritu, A.K.; Kinyuru, J.N.; Kenji, G.M.; Gichuhi, P.N. Extraction technique influences the physico-chemical characteristics and functional properties of edible crickets (Acheta domesticus) protein concentrate. J. Food Meas. Charact. 2017, 11, 2013–2021. [Google Scholar] [CrossRef]
- Marono, S.; Piccolo, G.; Loponte, R.; Di Meo, C.; Attia, Y.A.; Nizza, A.; Bovera, F. In vitro crude protein digestibility of Tenebrio Molitor and Hermetia illucens insect meals and its correlation with chemical composition traits. Ital. J. Anim. Sci. 2015, 14, 3889. [Google Scholar] [CrossRef]
- Caparros, M.R.; Poelaert, C.; Ernens, M.; Liotta, M.; Blecker, C.; Danthine, S.; Tyteca, E.; Haubruge, É.; Alabi, T.; Bindelle, J.; et al. Effect of household cooking techniques on the microbiological load and the nutritional quality of mealworms (Tenebrio molitor L. 1758). Food Res. Int. 2018, 106, 503–508. [Google Scholar] [CrossRef]
- Mancini, S.; Mattioli, S.; Paolucci, S.; Fratini, F.; Dal Bosco, A.; Tuccinardi, T.; Paci, G. Effect of cooking techniques on the in vitro protein digestibility, fatty acid profile, and oxidative status of mealworms (Tenebrio molitor). Front. Vet. Sci. 2021, 8, 675572. [Google Scholar] [CrossRef]
- Yang, Q.; Liu, S.; Sun, J.; Yu, L.; Zhang, C.; Bi, J.; Yang, Z. Nutritional composition and protein quality of the edible beetle Holotrichia parallela. J. Insect Sci. 2014, 14, 139. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Choi, Y.-S.; Jo, K.; Kim, T.-K.; Yong, H.I.; Jung, S. Quality characteristics and protein digestibility of Protaetia brevitarsis larvae. J. Anim. Sci. Technol. 2020, 62, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Janssen, R.H.; Vincken, J.-P.; Arts, N.J.G.; Fogliano, V.; Lakemond, C.M.M. Effect of endogenous phenoloxidase on protein solubility and digestibility after processing of Tenebrio molitor, Alphitobius diaperinus and Hermetia illucens. Food Res. Int. 2019, 121, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, E.; Baraniak, B.; Karaś, M.; Rybczyńska, K.; Jakubczyk, A. Selected species of edible insects as a source of nutrient composition. Food Res. Int. 2015, 77, 460–466. [Google Scholar] [CrossRef]
- Fabrikov, D.; Vargas-García, C.; Barroso, F.G.; Sánchez-Muros, M.J.; Cacua-Ortíz, S.M.; Morales, A.E.; Cardenete, G.; Tomás-Almenar, C.; Melenchón, F. Effect on Intermediary Metabolism and Digestive Parameters of the High Substitution of Fishmeal with Insect Meal in Sparus aurata Feed. Insects 2021, 12, 965. [Google Scholar] [CrossRef] [PubMed]
- Melenchón, F.; Mercado, E.; De Pula, H.J.; Cardenete, G.; Barroso, F.G.; Fabrikov, D.; Lourenço, H.M.; Pessoa, M.-F.; Lagos, L.; Weththasinghe, P.; et al. Fishmeal Dietary Replacement up to 50%: A Comparative Study of Two Insect Meals for Rainbow Trout (Oncorhynchus mykiss). Animals 2022, 12, 179. [Google Scholar] [CrossRef] [PubMed]
- Purschke, B.; Meinlschmidt, P.; Horn, C.; Rieder, O.; Jäger, H. Improvement of techno-functional properties of edible insect protein from migratory locust by enzymatic hydrolysis. Eur. Food Res. Technol. 2018, 244, 999–1013. [Google Scholar] [CrossRef]
- Quinto, B.P.T.; Albuquerque, J.V.; Bezerra, R.S.; Peixoto, S.; Soares, R. Replacement of fishmeal by two types of fish protein hydrolysate in feed for postlarval shrimp Litopenaeus vannamei. Aquac. Nutr. 2018, 24, 768–776. [Google Scholar] [CrossRef]
- Zheng, K.; Liang, M.; Yao, H.; Wang, J.; Chang, Q. Effect of size-fractionated fish protein hydrolysate on growth and feed utilization of turbot (Scophthalmus maximus L.). Aquac. Res. 2013, 44, 895–902. [Google Scholar] [CrossRef]
- Cardoza, A.L.; Guerra, M.G.; Palomino, A.R. Use of fish hydrolysate in aquaculture: A review of some beneficial results in aquafeeds. Manglar 2021, 18, 215–222. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Caligiani, A.; Marseglia, A.; Leni, G.; Baldassarre, S.; Maistrello, L.; Dossena, A.; Sforza, S. Composition of black soldier fly prepupae and systematic approaches for extraction and fractionation of proteins, lipids and chitin. Food Res. Int. 2018, 105, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Van Boekel, M.A.J.S.; Boeren, S.; Lakemond, C.M.M. Protein identification and in vitro digestion of fractions from Tenebrio molitor. Eur. Food Res. Technol. 2016, 242, 1285–1297. [Google Scholar] [CrossRef]
- Bishop, G.H. Body fluid of the honey bee larva. J. Biol. Chem. 1923, 58, 543–565. [Google Scholar] [CrossRef]
- Mukundan, M.K.; Radhakrishnan, A.G.; Stephen, J.; Antony, P.D. Nutritional evaluation of some fresh water fishes. Fish. Technol. 1986, 23, 189–195. [Google Scholar]
- Jonas-Levi, A.; Martinez, J.-J.I. The high level of protein content reported in insects for food and feed is overestimated. J. Food Compos. Anal. 2017, 62, 184–188. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CP 10 | EE 11 | ADF 12 | Ash | |
|---|---|---|---|---|
| FM 1 | 60.9 ± 0.6 bc | 14.4 ± 0.3 g | 7.84 ± 0.4 c | 19.5 ± 0.0 a |
| SBM 2 | 50.0 ± 0.5 e | 1.5 ± 0.1 h | 8.35 ± 0.8 c | 6.91 ± 0.0 c |
| HI 3 | 55.4 ± 1.3 d | 31.8 ± 0.4 a | 8.52 ± 0.5 bc | 8.75 ± 0.1 b |
| TM 4 | 63.4 ± 3.2 b | 29.4 ± 0.3 b | 7.82 ± 0.3 c | 4.97 ± 0.0 f |
| GA 5 | 60.4 ± 0.8 bc | 24.8 ± 0.4 c | 7.93 ± 0.3 c | 4.43 ± 0.0 g |
| GS 6 | 68.9 ± 0.5 a | 21.7 ± 0.4 d | 9.20 ± 0.4 bc | 5.43 ± 0.0 d |
| AchD 7 | 72.0 ± 0.3 a | 20.5 ± 0.4 e | 8.23 ± 0.3 c | 5.28 ± 0.1 e |
| AlpD 8 | 72.0 ± 0.2 a | 18.7 ± 0.2 f | 9.99 ± 0.9 b | 4.42 ± 0.0 g |
| MD 9 | 57.3 ± 1.2 cd | 29.5 ± 1.5 b | 16.4 ± 0.7 a | 5.38 ± 0.1 d |
| Sample | Prehydrolysis (0 min) | Gastric Phase (240 min) | Intestinal Phase (480 min) |
|---|---|---|---|
| Fishmeal | 5.10 ± 0.13 d | 15.31 ± 0.19 a | 32.14 ± 0.57 a |
| Soybean meal | 3.72 ± 0.09 e | 10.75 ± 0.36 c | 22.67 ± 0.20 d |
| T. molitor | 13.23 ± 0.20 a | 15.42 ± 0.24 b | 32.63 ± 0.33 a |
| H. illucens | 10.63 ± 0.36 b | 10.66 ± 0.24 c | 24.47 ± 0.48 c |
| G. assimilis | 10.89 ± 0.33 b | 13.71 ± 0.37 b | 22.75 ± 0.18 d |
| G. sigillatus | 3.62 ± 0.17 e | 7.66 ± 0.17 d | 16.60 ± 0.38 e |
| A. domestica | 9.19 ± 0.76 b | 12.98 ± 0.89 b | 29.32 ± 0.77 b |
| A. diaperinus | 5.36 ± 0.07 d | 7.34 ± 0.25 d | 15.69 ± 0.25 ef |
| M. domestica | 3.31 ± 0.07 e | 6.87 ± 0.38 d | 15.03 ± 0.39 f |
| TH | DH | OMd | CPd | ADF | CP | |
|---|---|---|---|---|---|---|
| EE | nsd | nsd | nsd | −0.5597 | nsd | nsd |
| CP | nsd | −0.4416 * | nsd | nsd | nsd | - |
| ADF | nsd | −0.5981 | −0.6456 | −0.5970 | - | - |
| CPd | nsd | nsd | 0.4509 * | - | - | - |
| OMd | nsd | nsd | - | - | - | - |
| DH | 0.7016 | - | - | - | - | - |
| TH | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Rodríguez, M.; Sánchez-Muros, M.J.; Vargas-García, M.d.C.; Varga, A.T.; Fabrikov, D.; Barroso, F.G. Evaluation of In Vitro Protein Hydrolysis in Seven Insects Approved by the EU for Use as a Protein Alternative in Aquaculture. Animals 2024, 14, 96. https://doi.org/10.3390/ani14010096
Rodríguez-Rodríguez M, Sánchez-Muros MJ, Vargas-García MdC, Varga AT, Fabrikov D, Barroso FG. Evaluation of In Vitro Protein Hydrolysis in Seven Insects Approved by the EU for Use as a Protein Alternative in Aquaculture. Animals. 2024; 14(1):96. https://doi.org/10.3390/ani14010096
Chicago/Turabian StyleRodríguez-Rodríguez, María, María José Sánchez-Muros, María del Carmen Vargas-García, Agnes Timea Varga, Dmitri Fabrikov, and Fernando G. Barroso. 2024. "Evaluation of In Vitro Protein Hydrolysis in Seven Insects Approved by the EU for Use as a Protein Alternative in Aquaculture" Animals 14, no. 1: 96. https://doi.org/10.3390/ani14010096