
Thyme, Oregano, and Garlic Essential Oils and Their Main Active Compounds Influence Eimeria tenella Intracellular Development
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Parasites and Birds
2.2. Cell Culture
2.3. Chemicals and Reagents
2.4. Development Assay
2.5. Isolation of Nucleic Acids
2.6. Real Time Quantitative PCR (qPCR)
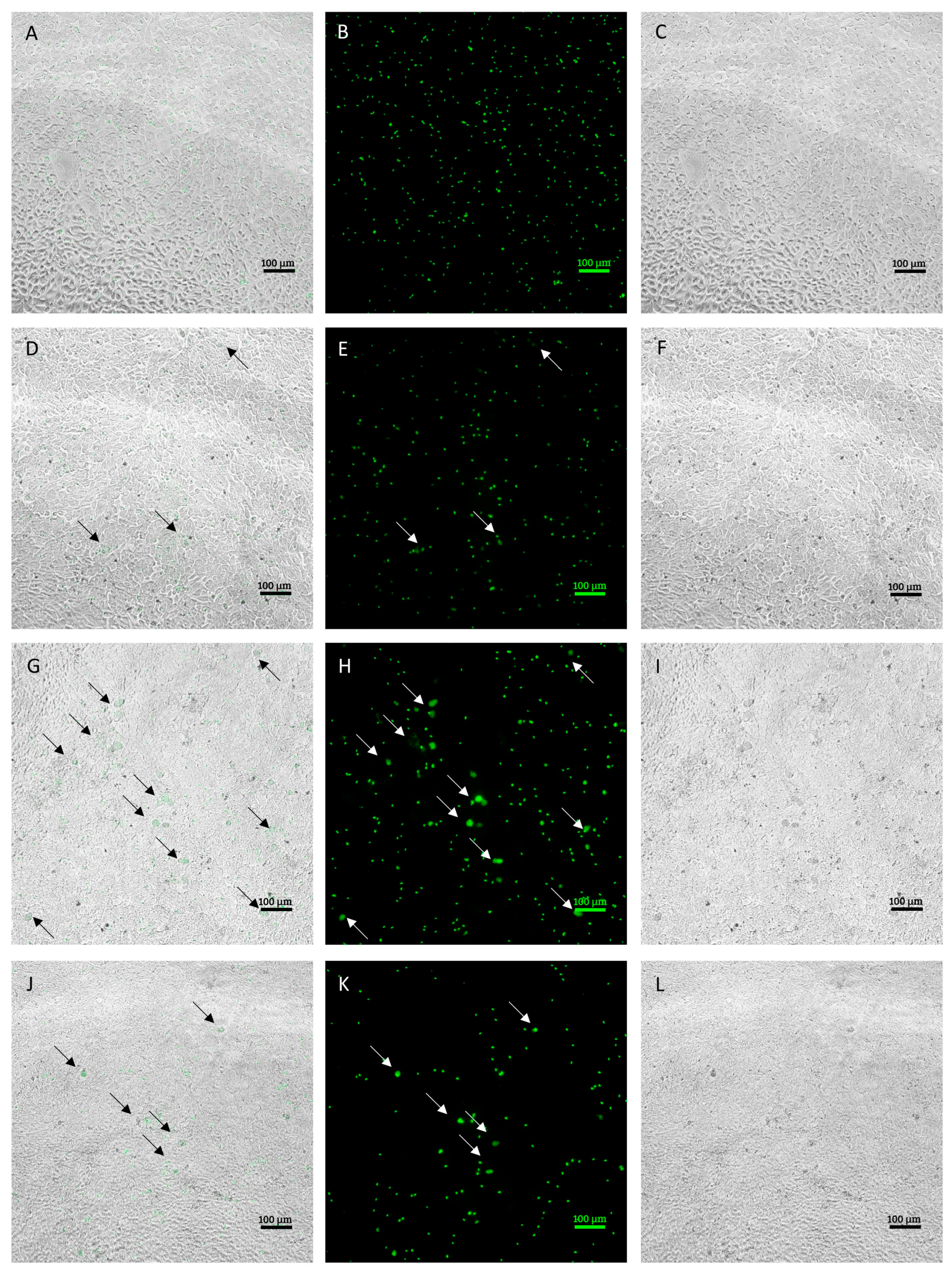
2.7. Image Analysis
2.8. Statistical Analysis
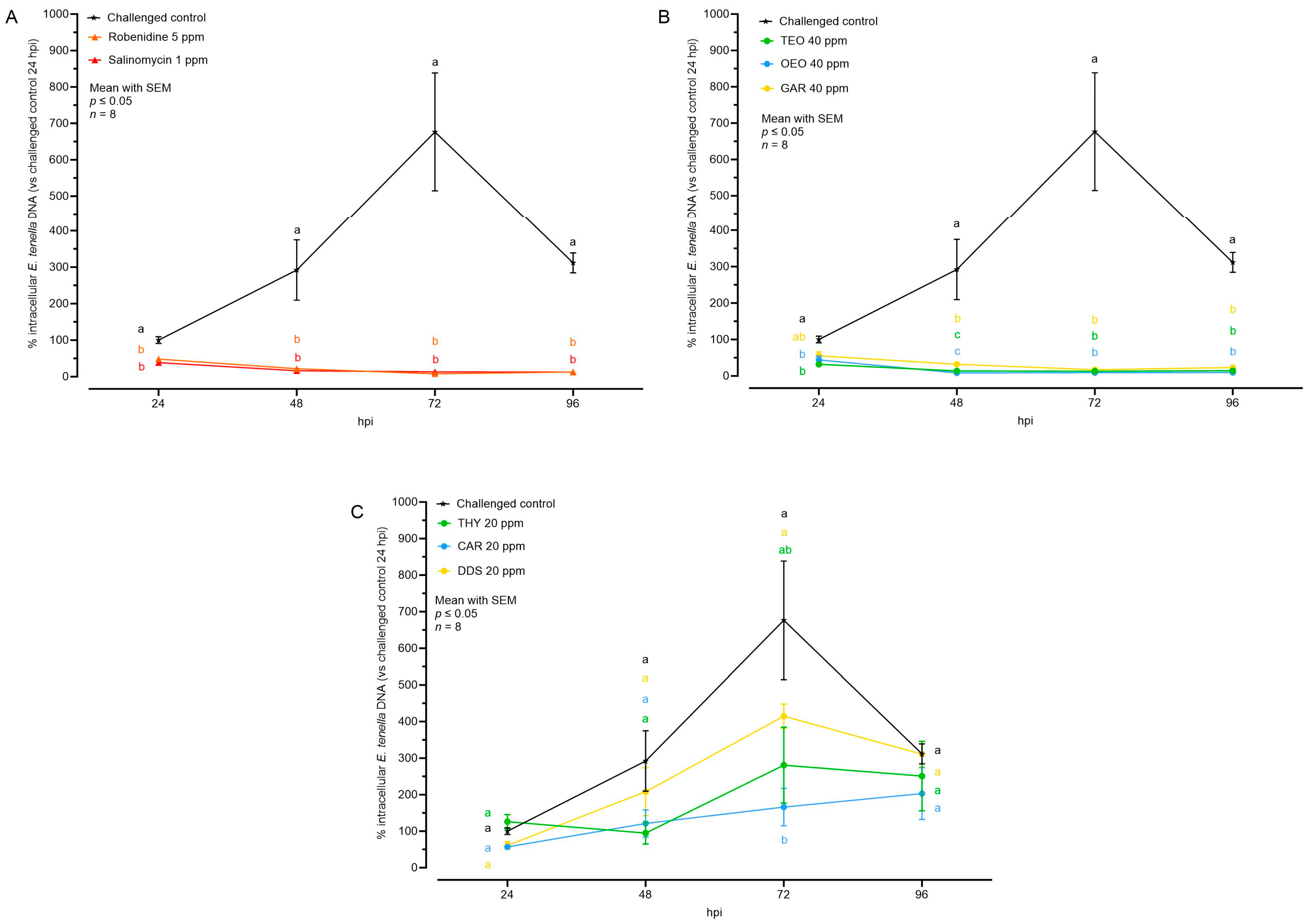
3. Results
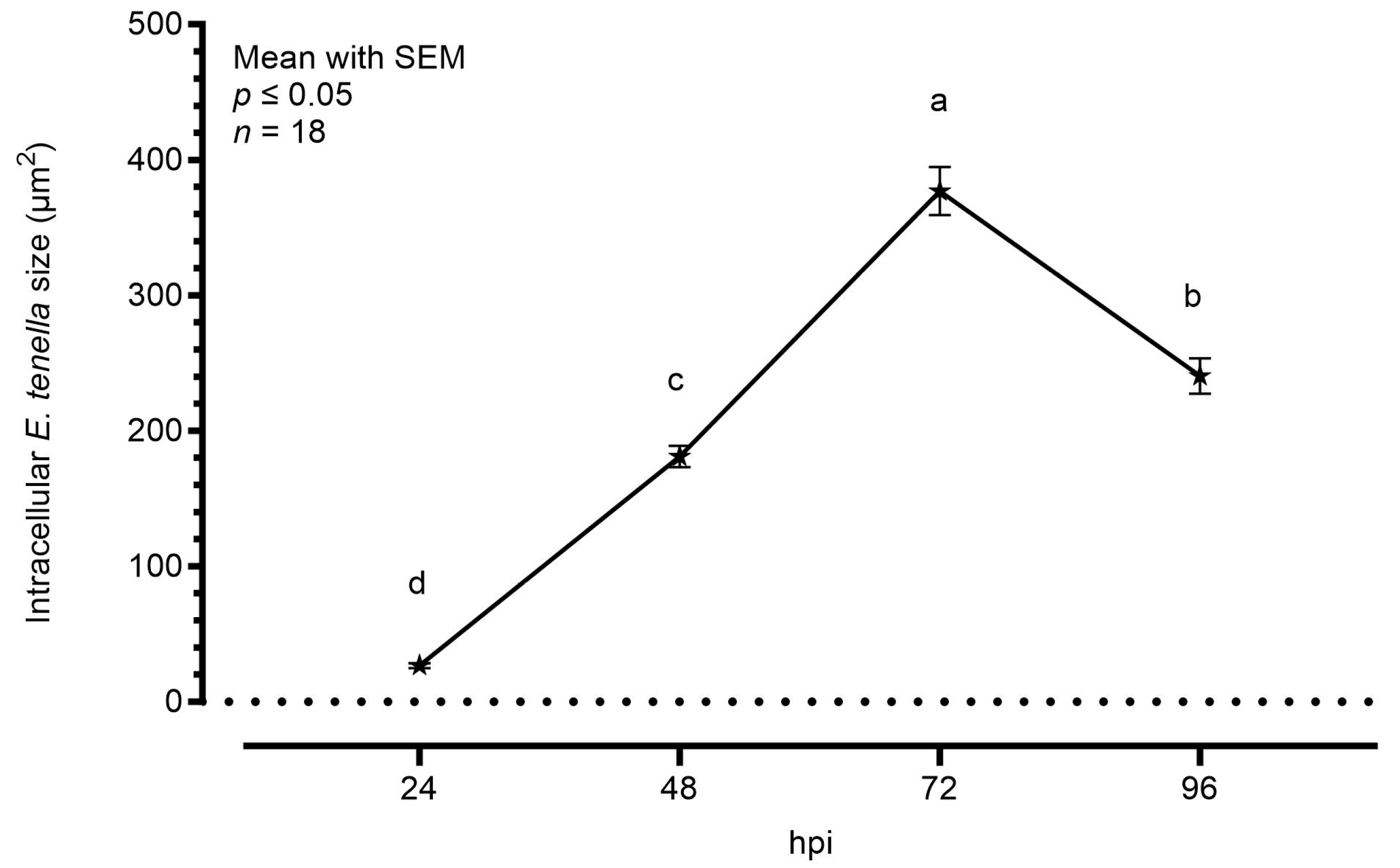
3.1. Characterization of E. tenella Intracellular Growth in MDBK Cells
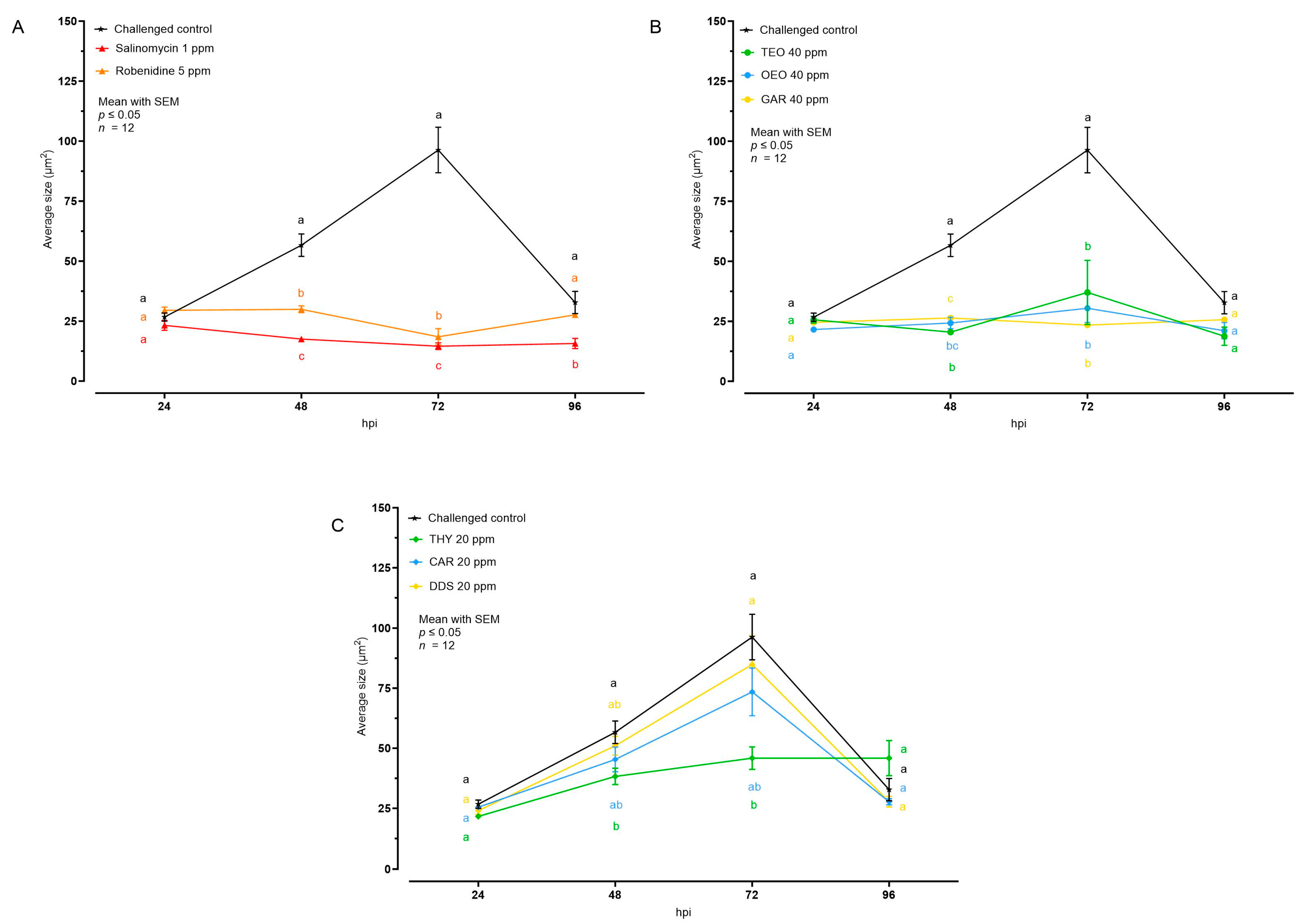
3.2. Effects of Natural Compounds on Intracellular Growth
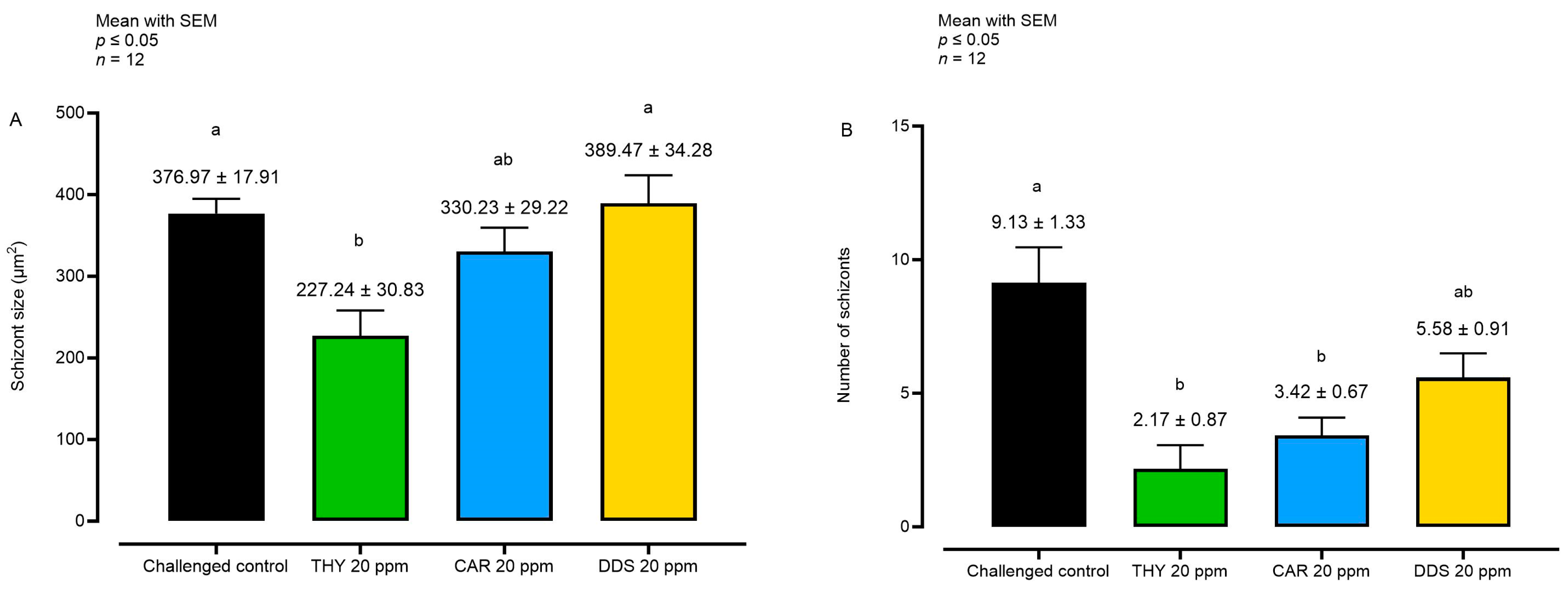
3.3. Effects of Natural Compounds on Parasite Invasion and Replication
4. Discussion
4.1. Anticoccidial Drugs
4.2. Essential Oils
4.3. Nature-Identical Compounds
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Jatau, I.D.; et al. Re-calculating the cost of coccidiosis in chickens. Vet. Res. 2020, 51, 115. [Google Scholar] [CrossRef] [PubMed]
- Collier, C.T.; Hofacre, C.L.; Payne, A.M.; Anderson, D.B.; Kaiser, P.; Mackie, R.I.; Gaskins, H.R. Coccidia-induced mucogenesis promotes the onset of necrotic enteritis by supporting Clostridium perfringens growth. Vet. Immunol. Immunopathol. 2008, 122, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, S.E.; Van Diemen, P.M.; Martineau, H.; Stevens, M.P.; Tomley, F.M.; Stabler, R.A.; Blake, P. Impact of Eimeria tenella Coinfection on Campylobacter jejuni Colonization of the Chicken. J. Hepatol. 2019, 87, e00772-18. [Google Scholar] [CrossRef] [PubMed]
- Rimet, C.S.; Maurer, J.J.; Berghaus, R.D.; Jordan, B.J.; Antoniassi Da Silva, L.H.; Stabler, L.J.; Johnson, K.K.; Tensa, L.R.; Segovia, K.M.; França, M.S. The Contribution of Eimeria Coinfection and Intestinal Inflammation to Cecal Colonization and Systemic Spread of Salmonella Typhimurium Deficient in Tetrathionate Reductase or Type III Secretion Systems Salmonella Pathogenicity Island 1 or 2. Avian Dis. 2019, 63, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Noack, S.; Chapman, H.D.; Selzer, P.M. Anticoccidial drugs of the livestock industry. Parasitol. Res. 2019, 118, 2009–2026. [Google Scholar] [CrossRef] [PubMed]
- Muthamilselvan, T.; Kuo, T.F.; Wu, Y.C.; Yang, W.C. Herbal remedies for coccidiosis control: A review of plants, compounds, and anticoccidial actions. Evid.-Based Complement. Altern. Med. 2016, 2016, 2657981. [Google Scholar] [CrossRef] [PubMed]
- Gordillo Jaramillo, F.X.; Kim, D.H.; Lee, S.H.; Kwon, S.K.; Jha, R.; Lee, K.W. Role of oregano and Citrus species-based essential oil preparation for the control of coccidiosis in broiler chickens. J. Anim. Sci. Biotechnol. 2021, 12, 47. [Google Scholar] [CrossRef]
- Chang, L.Y.; Di, K.Q.; Xu, J.; Chen, Y.F.; Xi, J.Z.; Wang, D.H.; Hao, E.Y.; Xu, L.J.; Chen, H.; Zhou, R.Y. Effect of natural garlic essential oil on chickens with artificially infected Eimeria tenella. Vet. Parasitol. 2021, 300, 109614. [Google Scholar] [CrossRef]
- Sidiropoulou, E.; Skoufos, I.; Marugan-Hernandez, V.; Giannenas, I.; Bonos, E.; Aguiar-Martins, K.; Lazari, D.; Blake, D.P.; Tzora, A. In vitro Anticoccidial Study of Oregano and Garlic Essential Oils and Effects on Growth Performance, Fecal Oocyst Output, and Intestinal Microbiota in vivo. Front. Vet. Sci. 2020, 7, 420. [Google Scholar] [CrossRef]
- Gholami-Ahangaran, M.; Ahmadi-Dastgerdi, A.; Azizi, S.; Basiratpour, A.; Zokaei, M.; Derakhshan, M. Thymol and carvacrol supplementation in poultry health and performance. Vet. Med. Sci. 2022, 8, 267–288. [Google Scholar] [CrossRef]
- Spisni, E.; Petrocelli, G.; Imbesi, V.; Spigarelli, R.; Azzinnari, D.; Sarti, M.D.; Campieri, M.; Valerii, M.C. Antioxidant, anti-inflammatory, and microbial-modulating activities of essential oils: Implications in colonic pathophysiology. Int. J. Mol. Sci. 2020, 21, 4152. [Google Scholar] [CrossRef] [PubMed]
- Reza Youssefi, M.; Moghaddas, E.; Abouhosseini Tabari, M.; Moghadamnia, A.A.; Hosseini, S.M.; Hosseini Farash, B.R.; Amin Ebrahimi, M.; Nabavi Mousavi, N.; Fata, A.; Maggi, F.; et al. In Vitro and In Vivo Effectiveness of Carvacrol, Thymol and Linalool against Leishmania infantum Mohammad. Molecules 2019, 24, 2072. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Uscanga, A.; Aycart, D.F.; Li, K.; Witola, W.H.; Andrade Laborde, J.E. Anti-protozoal activity of Thymol and a Thymol ester against Cryptosporidium parvum in cell culture. Int. J. Parasitol. Drugs Drug Resist. 2021, 15, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Bastaki, S.M.A.; Ojha, S.; Kalasz, H.; Adeghate, E. Chemical constituents and medicinal properties of Allium species. Mol. Cell. Biochem. 2021, 476, 4301–4321. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Chand, N.; Khan, R.U.; Naz, S.; Gul, S. Anticoccidial effect of garlic (Allium sativum) and ginger (Zingiber officinale) against experimentally induced coccidiosis in broiler chickens. J. Appl. Anim. Res. 2019, 47, 79–84. [Google Scholar] [CrossRef]
- Felici, M.; Tugnoli, B.; Piva, A.; Grilli, E. In vitro assessment of anticoccidials: Methods and molecules. Animals 2021, 11, 1962. [Google Scholar] [CrossRef] [PubMed]
- Patton, W.H. Eimeria tenella: Cultivation of the Asexual Stages in Cultured Animal Cells. Science 1965, 150, 767–769. [Google Scholar] [CrossRef] [PubMed]
- Marugan-Hernandez, V.; Jeremiah, G.; Aguiar-Martins, K.; Burrell, A.; Vaughan, S.; Xia, D.; Randle, N.; Tomley, F. The Growth of Eimeria tenella: Characterization and Application of Quantitative Methods to Assess Sporozoite Invasion and Endogenous Development in Cell Culture. Front. Cell. Infect. Microbiol. 2020, 10, 579833. [Google Scholar] [CrossRef]
- McDougald, L.R.; Galloway, R.B. Anticoccidial drugs: Effects on infectivity and survival intracellularly of Eimeria tenella sporozoites. Exp. Parasitol. 1976, 40, 314–319. [Google Scholar] [CrossRef]
- Clark, J.D.; Billington, K.; Bumstead, J.M.; Oakes, R.D.; Soon, P.E.; Sopp, P.; Tomley, F.M.; Blake, D.P. A toolbox facilitating stable transfection of Eimeria species. Mol. Biochem. Parasitol. 2008, 162, 77–86. [Google Scholar] [CrossRef]
- Shirley, M.W. Eimeria species and strains of chickens. In Guidelines on Techniques in Coccidiosis Research; Office for Official Publications of the European Communities: Luxembourg, 1995; pp. 1–25. ISBN 9282749703. [Google Scholar]
- Pastor-Fernández, I.; Pegg, E.; Macdonald, S.E.; Tomley, F.M.; Blake, D.P.; Marugán-Hernández, V. Laboratory Growth and Genetic Manipulation of Eimeria tenella. Curr. Protoc. Microbiol. 2019, 53, e81. [Google Scholar] [CrossRef] [PubMed]
- Felici, M.; Tugnoli, B.; Ghiselli, F.; Baldo, D.; Ratti, C.; Piva, A.; Grilli, E. Investigating the effects of essential oils and pure botanical compounds against Eimeria tenella in vitro. Poult. Sci. 2023, 102, 102898. [Google Scholar] [CrossRef] [PubMed]
- Marugan-Hernandez, V.; Cockle, C.; Macdonald, S.; Pegg, E.; Crouch, C.; Blake, D.P.; Tomley, F.M. Viral proteins expressed in the protozoan parasite Eimeria tenella are detected by the chicken immune system. Parasites Vectors 2016, 9, 463. [Google Scholar] [CrossRef] [PubMed]
- Arias-Maroto, S.; Aguiar-Martins, K.; Regidor-Cerrillo, J.; Ortega-Mora, L.; Marugan-Hernandez, V. Reduction of chickens use to perform in vitro pre-screening of novel anticoccidials by miniaturisation and increased throughput of the current Eimeria tenella compound-screening model. F1000Research 2022, 11, 1135. [Google Scholar] [CrossRef]
- Thabet, A.; Alnassan, A.A.; Daugschies, A.; Bangoura, B. Combination of cell culture and qPCR to assess the efficacy of different anticoccidials on Eimeria tenella sporozoites. Parasitol. Res. 2015, 114, 2155–2163. [Google Scholar] [CrossRef]
- Müller, J.; Hemphill, A. In vitro culture systems for the study of apicomplexan parasites in farm animals. Int. J. Parasitol. 2013, 43, 115–124. [Google Scholar] [CrossRef]
- Sidiropoulou, E.; Marugán-Hernández, V.; Skoufos, I.; Giannenas, I.; Bonos, E.; Aguiar-Martins, K.; Lazari, D.; Papagrigoriou, T.; Fotou, K.; Grigoriadou, K.; et al. In Vitro Antioxidant, Antimicrobial, Anticoccidial, and Anti-Inflammatory Study of Essential Oils of Oregano, Thyme, and Sage from Epirus, Greece. Life 2022, 12, 1783. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, D.K. Pharmacological effects of garlic (Allium sativum L.). Annu. Rev. Biomed. Sci. 2008, 10, 6–26. [Google Scholar] [CrossRef]
- Long, P.L. Endogenous stages of a ‘chick embryo-adapted’ strain of Eimeria tenella. Parasitology 1973, 66, 55–62. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average Size (μm2) | ||||
|---|---|---|---|---|
| 24 hpi | 48 hpi | 72 hpi | 96 hpi | |
| Challenged control | 26.70 ± 1.72 | 56.65 ± 4.70 | 96.30 ± 9.49 | 32.8 ± 4.65 |
| Robenidine 5 ppm | 29.49 ± 1.32 | 29.90 ± 1.44 | 18.47 ± 3.38 | 27.60 ± 1.14 |
| Salinomycin 1 ppm | 23.21 ± 2.07 | 17.50 ± 1.13 | 14.54 ± 1.40 | 15.66 ± 2.12 |
| TEO 40 ppm | 25.62 ± 1.40 | 20.46 ± 1.14 | 37.01 ± 13.38 | 18.76 ± 3.81 |
| OEO 40 ppm | 21.58 ± 1.22 | 24.30 ± 2.41 | 30.41 ± 5.89 | 21.08 ± 3.39 |
| GAR 40 ppm | 24.54 ± 1.16 | 26.35 ± 0.60 | 23.38 ± 1.21 | 25.67 ± 0.97 |
| THY 20 ppm | 21.59 ± 0.92 | 38.26 ± 3.39 | 45.88 ± 4.71 | 45.85 ± 7.33 |
| CAR 20 ppm | 25.45 ± 1.04 | 45.31 ± 5.16 | 73.44 ± 9.94 | 27.71 ± 1.26 |
| DDS 20 ppm | 24.05 ± 1.60 | 51.02 ± 3.91 | 84.870 ± 11.98 | 27.76 ± 2.23 |
| Rate (μm2/h) | |
|---|---|
| Challenged control | 1.21 ± 0.08 a |
| Robenidine 5 ppm | −0.23 ± 0.09 d |
| Salinomycin 1 ppm | −0.17± 0.04 d |
| TEO 40 ppm | −0.04 ± 0.03 cd |
| OEO 40 ppm | 0.07 ± 0.06 cd |
| GAR 40 ppm | −0.03 ± 0.03 cd |
| THY 20 ppm | 0.51 ± 0.10 bc |
| CAR 20 ppm | 0.87 ± 0.24 ab |
| DDS 20 ppm | 0.97 ± 0.27 ab |
| Level of Inhibition (%) | |
|---|---|
| Challenged control | 0.00 ± 9.29 bc |
| Robenidine 5 ppm | 51.65 ± 5.44 a |
| Salinomycin 1 ppm | 61.40 ± 3.04 a |
| TEO 40 ppm | 67.76 ± 5.88 a |
| OEO 40 ppm | 55.86 ± 10.63 a |
| GAR 40 ppm | 44.70± 10.81 a |
| THY 20 ppm | −26.08 ± 19.74 c |
| CAR 20 ppm | 43.05 ± 3.62 a |
| DDS 20 ppm | 38.96± 10.47 ab |
| Intracellular E. tenella DNA (%) | ||||
|---|---|---|---|---|
| 24 hpi | 48 hpi | 72 hpi | 96 hpi | |
| Challenged control | 100.00 ± 9.29 | 291.99 ± 82.71 | 676.43 ± 162.14 | 311.58 ± 27.35 |
| Robenidine 5 ppm | 48.35 ± 5.44 | 21.93 ± 4.14 | 8.06 ± 2.36 | 12.87 ± 4.75 |
| Salinomycin 1 ppm | 38.61 ± 3.04 | 16.28 ± 2.62 | 13.34 ± 3.97 | 12.80 ± 3.81 |
| TEO 40 ppm | 32.24 ± 5.88 | 13.73 ± 2.88 | 13.34 ± 2.14 | 14.25 ± 3.14 |
| OEO 40 ppm | 44.14 ± 10.63 | 8.54 ± 2.18 | 8.98 ± 1.59 | 9.43 ± 2.77 |
| GAR 40 ppm | 64.15 ± 12.88 | 32.12 ± 3.72 | 16.99 ± 3.78 | 23.31 ± 7.41 |
| THY 20 ppm | 126.08 ± 19.74 | 94.65 ± 30.03 | 280.62 ± 103.94 | 250.85 ± 95.30 |
| CAR 20 ppm | 71.43 ± 14.82 | 121.33 ± 37.52 | 166.03 ± 51.33 | 203.15 ± 71.00 |
| DDS 20 ppm | 61.04 ± 10.47 | 208.01 ± 65.58 | 414.91 ± 33.20 | 310.68 ± 34.73 |
| Rate (genomes/h) | |
|---|---|
| Challenged control | 6.44 ± 0.94 ab |
| Robenidine 5 ppm | −0.84 ± 0.14 c |
| Salinomycin 1 ppm | −0.53 ± 0.09 c |
| TEO 40 ppm | −0.39 ± 0.09 c |
| OEO 40 ppm | −0.73 ± 0.23 c |
| GAR 40 ppm | −0.78 ± 0.30 c |
| THY 20 ppm | 3.55 ± 2.45 abc |
| CAR 20 ppm | 2.63 ± 1.16 ac |
| DDS 20 ppm | 7.37 ± 0.82 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Felici, M.; Tugnoli, B.; De Hoest-Thompson, C.; Piva, A.; Grilli, E.; Marugan-Hernandez, V. Thyme, Oregano, and Garlic Essential Oils and Their Main Active Compounds Influence Eimeria tenella Intracellular Development. Animals 2024, 14, 77. https://doi.org/10.3390/ani14010077
Felici M, Tugnoli B, De Hoest-Thompson C, Piva A, Grilli E, Marugan-Hernandez V. Thyme, Oregano, and Garlic Essential Oils and Their Main Active Compounds Influence Eimeria tenella Intracellular Development. Animals. 2024; 14(1):77. https://doi.org/10.3390/ani14010077
Chicago/Turabian StyleFelici, Martina, Benedetta Tugnoli, Callum De Hoest-Thompson, Andrea Piva, Ester Grilli, and Virginia Marugan-Hernandez. 2024. "Thyme, Oregano, and Garlic Essential Oils and Their Main Active Compounds Influence Eimeria tenella Intracellular Development" Animals 14, no. 1: 77. https://doi.org/10.3390/ani14010077