
Resveratrol Reduces ROS by Increasing GSH in Vitrified Sheep Embryos
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Oocyte Collection
2.2. In Vitro Maturation (IVM) of Oocytes
2.3. In Vitro Fertilization (IVF) and In Vitro Development (IVD) of Embryos
2.4. Vitrification and Devitrification of Embryos
2.5. Quantification of ROS and GSH in Devitrified Embryos
2.6. DAPI Staining of Nuclei
2.7. Statistical Analysis
3. Results
3.1. Oocyte Collection
3.2. In Vitro Maturation (IVM) of Oocytes, In Vitro Fertilization (IVF), and In Vitro Development (IVD) of Embryos
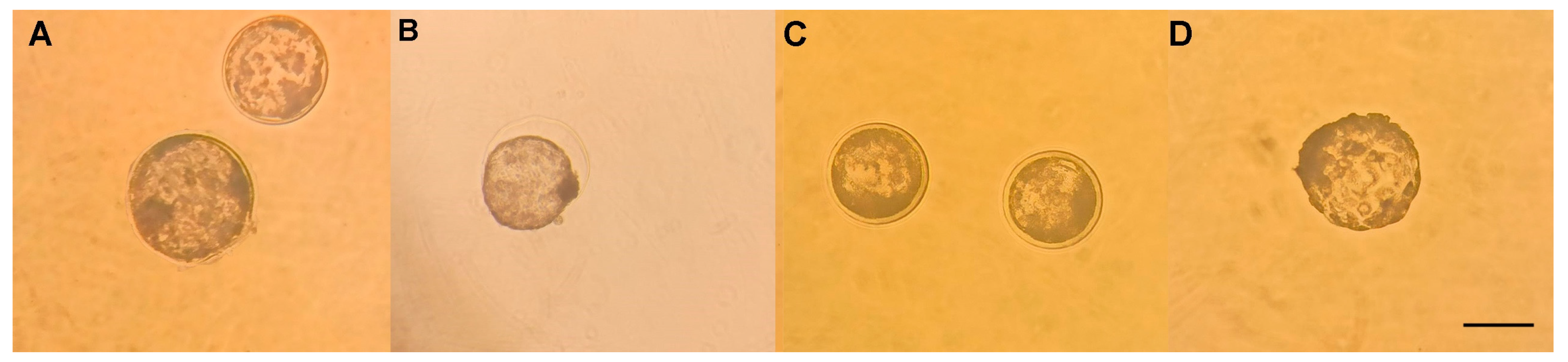
3.3. Vitrification and Devitrification of Embryos
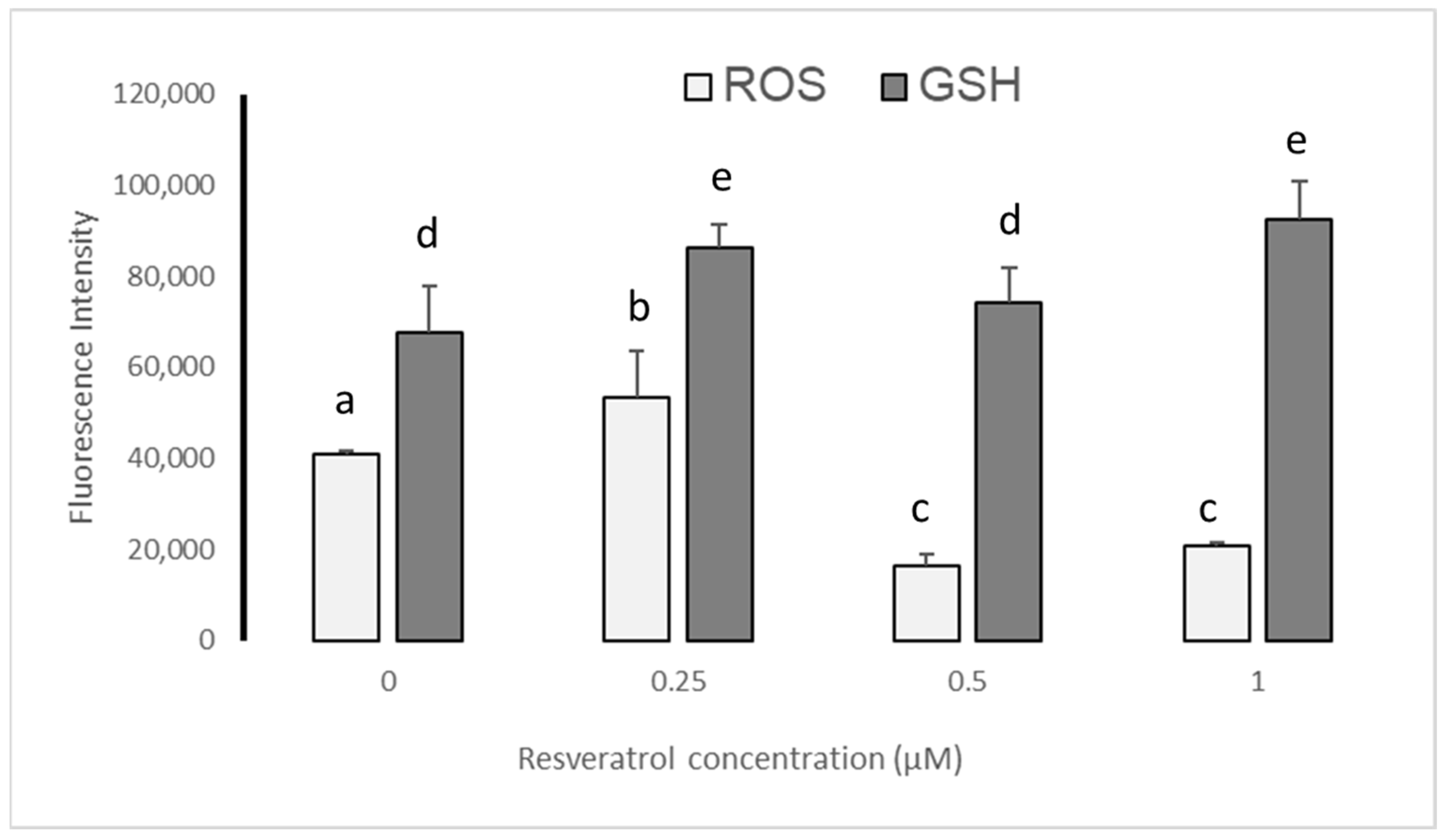
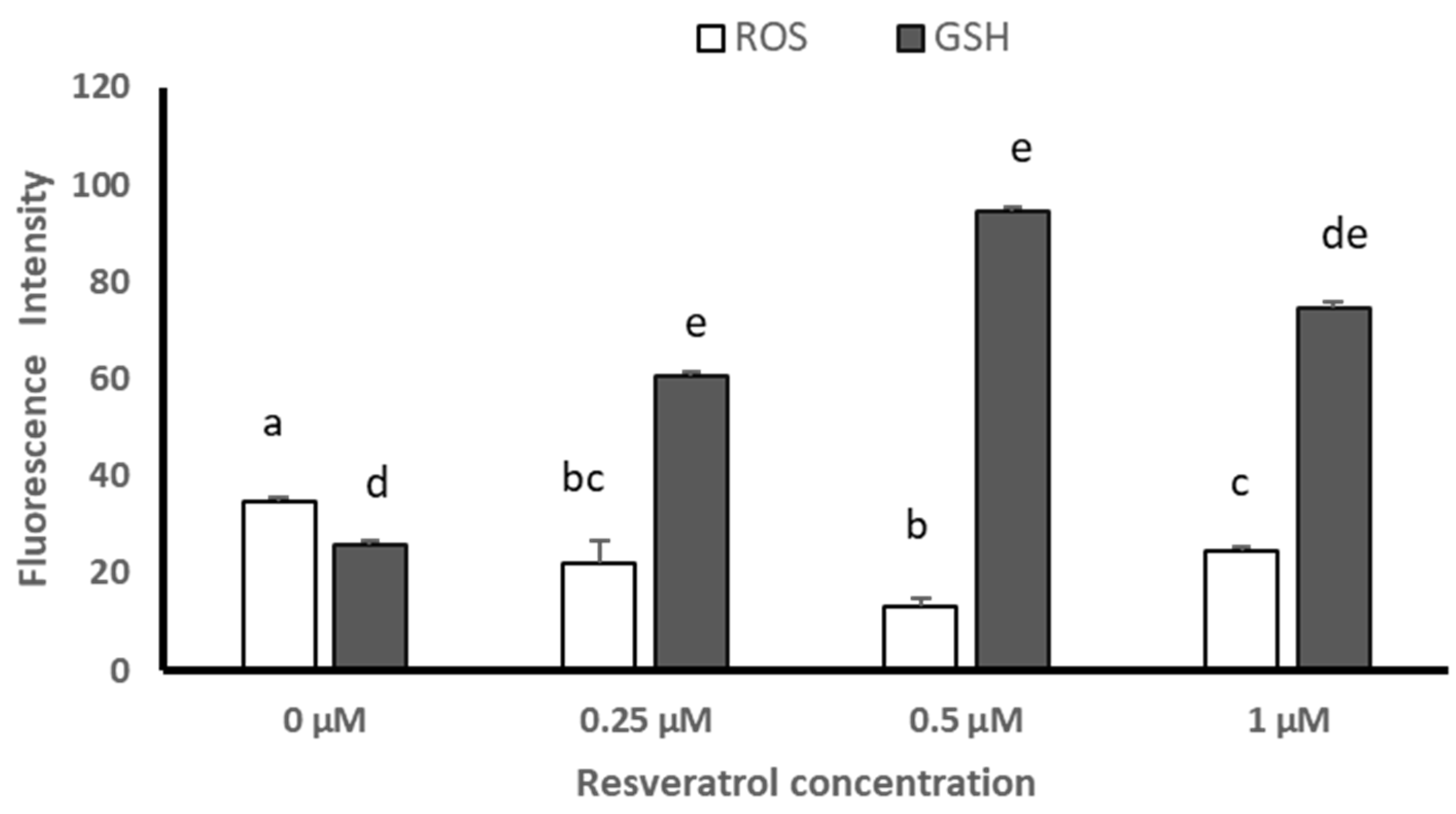
3.4. ROS and GSH Levels in Devitrified Embryos
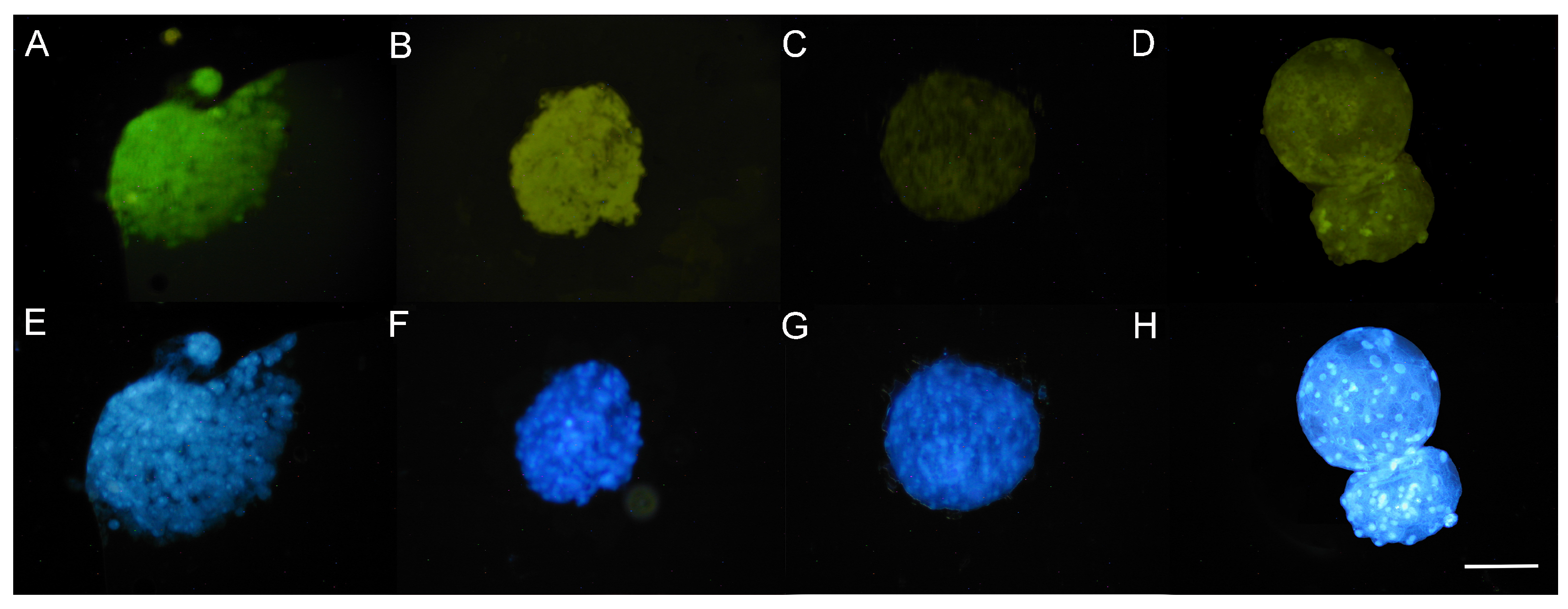
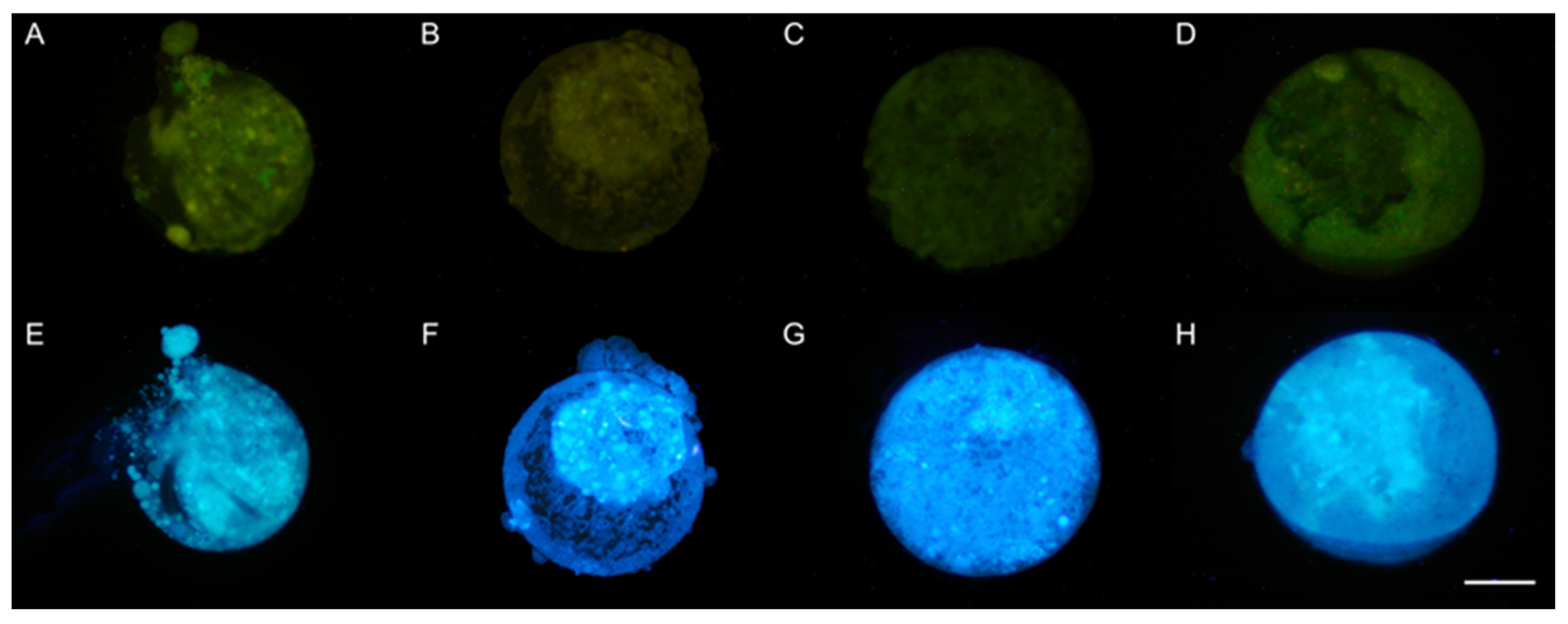
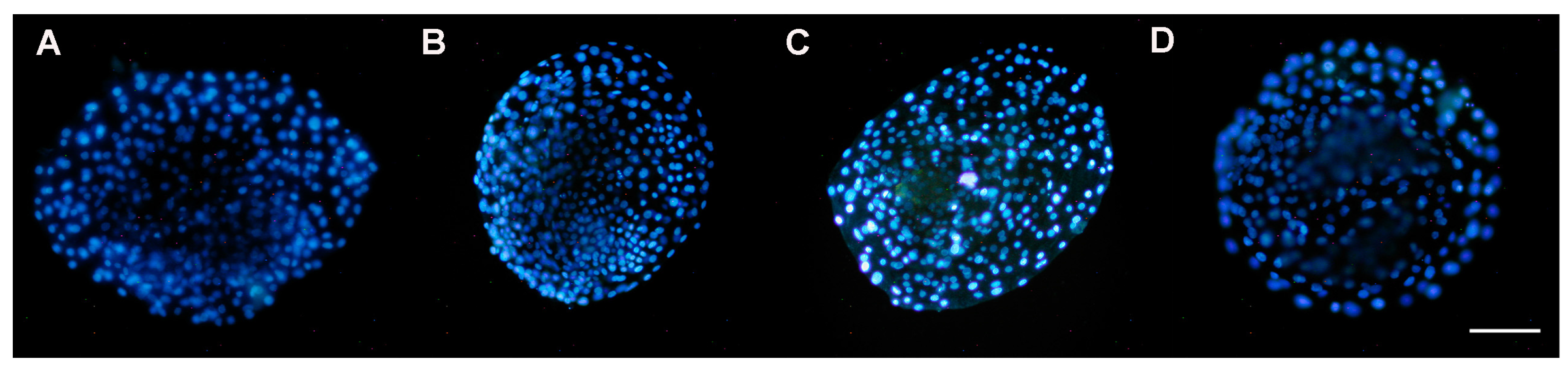
3.5. DAPI Staining of Nuclei
3.5.1. Presence of Nuclei in Early Blastocysts Prior to Vitrification
3.5.2. Presence of Nuclei in Late Blastocysts after Devitrification
4. Discussion
4.1. Effect of Resveratrol on IVM, DIV, and Blastocyst Production
4.2. Effect of Resveratrol on Embryo Vitrification and Devitrification
4.3. Effect of Resveratrol on ROS and GSH Levels in Devitrified Embryos
4.4. Effect of Resveratrol on Embryo Quality
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parra-Cortés, R.I.; Valderrama-Rodas, M.; Martínez-Correal, G. Situación actual y perspectivas de la ganadería de bovinos criollos en América Latina. Arch. Latinoam. Prod. Anim. 2021, 29, 79–90. [Google Scholar] [CrossRef]
- Rodríguez Suástegui, J.L.; Romo García, S.D.; Casas Hernández, E.; Hernández Pichardo, J.E. Desarrollo de mórulas de ovino en medio simple o secuencial: Relación entre evaluación morfológica y viabilidad embrionaria. Rev. Salud Anim. 2017, 39, 9–18. [Google Scholar]
- Martínez-Ibarra, J.L.; Espinoza-Mendoza, E.A.; Rangel-Santos, R.; Ambriz-García, D.A.; Navarro-Maldonado, M.D.C. Effect of resveratrol on the in vitro maturation of ovine (Ovis aries) oocytes and the subsequent development of handmade cloned embryos. Vet. Méx. 2019, 5, 1–14. [Google Scholar] [CrossRef]
- Mukherjee, A.; Malik, H.; Saha, A.P.; Dubey, A.; Singhal, D.K.; Boateng, S.; Saugandhika, S.; Kumar, S.; De, S.; Guha, S.K.; et al. Resveratrol treatment during goat oocytes maturation enhances developmental competence of parthenogenetic and hand-made cloned blastocysts by modulating intracellular glutathione level and embryonic gene expression. J. Assist. Reprod. Genet. 2014, 31, 229–239. [Google Scholar] [CrossRef]
- Zabihi, A.; Shabankareh, H.K.; Hajarian, H.; Foroutanifar, S. In vitro maturation medium supplementation with resveratrol improves cumulus cell expansion and developmental competence of Sanjabi sheep oocytes. Livest. Sci. 2021, 243, 104378. [Google Scholar] [CrossRef]
- Gaviria, S.M.; Herrera, A.L.; Urrego, R.; Betancur, G.R.; Zuluaga, J.J.E. Effect of resveratrol on vitrified in vitro produced bovine embryos: Recovering the initial quality. Cryobiology 2019, 89, 42–50. [Google Scholar] [CrossRef]
- Kwak, S.-S.; Cheong, S.-A.; Jeon, Y.; Lee, E.; Choi, K.-C.; Jeung, E.-B.; Hyun, S.-H. The effects of resveratrol on porcine oocyte in vitro maturation and subsequent embryonic development after parthenogenetic activation and in vitro fertilization. Theriogenology 2012, 78, 86–101. [Google Scholar] [CrossRef]
- Abdul Rahman, N.-S.; Mohamed Noor Khan, N.-A.; Eshak, Z.; Sarbandi, M.-S.; Mohammad Kamal, A.-A.; Abd Malek, M.; Abdullah, F.; Abdullah, M.A.; Othman, F. Exogenous L-Glutathione Improves Vitrification Outcomes in Murine Preimplantation Embryos. Antioxidants 2022, 11, 2100. [Google Scholar] [CrossRef]
- García-Martínez, T.; Vendrell-Flotats, M.; Martínez-Rodero, I.; Ordóñez-León, E.A.; Álvarez-Rodríguez, M.; López-Béjar, M.; Yeste, M.; Mogas, T. Glutathione Ethyl Ester Protects In Vitro-Maturing Bovine Oocytes against Oxidative Stress Induced by Subsequent Vitrification/Warming. Int. J. Mol. Sci. 2020, 21, 7547. [Google Scholar] [CrossRef]
- Xiang, D.; Jia, B.; Zhang, B.; Liang, J.; Hong, Q.; Wei, H.; Wu, G. Astaxanthin supplementation improves the subsequent developmental competence of vitrified porcine zygotes. Front. Vet. Sci. 2022, 9, 871289. [Google Scholar] [CrossRef]
- Giraldo Giraldo, J.J. Efecto del Crioprotector Dimetilformamida Sobre la Viabilidad de Embriones Bovinos Producidos In Vitro. Master’s Thesis, Universidad Nacional de Colombia, Facultad de Ciencias, Sede Medellín, Bogotá, Colombia, 2011; 96p. [Google Scholar]
- Asociación para el Estudio de la Biología de la Reproducción (ASEBIR). Criterios ASEBIR de Valoración Morfológica de Oocitos, Embriones Tempranos y Blastocistos Humanos, 3rd ed.; Cuadernos de Embriología Clínica: Madrid, Spain, 2015; pp. 9–75. [Google Scholar]
- Filipiak, Y.; Larocca, C.; Martínez, M. Comportamiento del Semen Bovino Sexado Congelado-Descongelado en Fertilización in vitro (FIV) Capacitado Mediante BO en dos Concentraciones versus Percoll. Int. J. Morphol. 2017, 35, 1337–1341. [Google Scholar] [CrossRef]
- Salgado-Cruz, E.; Vasquez, R.L. Aspectos esenciales sobre las técnicas de fertilización in vitro en bovinos. Rev. Investig. Vet. Perú 2020, 31, e17138. [Google Scholar] [CrossRef]
- Bhat, M.H.; Sharma, V.; Khan, F.A.; Naykoo, N.A.; Yaqoob, S.H.; Vajta, G.; Khan, H.M.; Fazili, M.R.; Ganai, N.A.; Shah, R.A. Open pulled straw vitrification and slow freezing of sheep IVF embryos using different cryoprotectants. Reprod. Fertil. Dev. 2015, 27, 1175–1180. [Google Scholar] [CrossRef]
- Vajta, G. Vitrification of the oocytes and embryos of domestic animals. Anim. Reprod. Sci. 2000, 60–61, 357–364. [Google Scholar] [CrossRef]
- Sosa, F.; Hansen, P.J. Protocol for embryo vitrification using open pulled straws. In Procedures for In Vitro Production of Bovine Embryos; University of Florida: Gainesville, FL, USA, 2019. [Google Scholar]
- González Mendoza, D.F. Efecto del Protocolo de Vitrificación y Sistemas de Empaque Sobre la Tasa de Supervivencia de Embriones Ovinos Obtenidos In Vivo. Master’s Thesis, Universidad Nacional de Colombia, Facultad de Medicina Veterinaria y de Zootecnia, Programas de Posgrados, Bogotá, Colombia, 2020. [Google Scholar]
- Vazquez-Avendaño, J.R.; Hernández-Martínez, S.; Hernández-Pichardo, J.E.; Rivera-Rebolledo, J.A.; Ambriz-García, D.A.; Navarro-Maldonado, M.C. Efecto del uso de medio secuencial humano en la producción de blastocistos de hembra ovis canadensis mexicana por clonación manual. Acta Zool. Mex. 2017, 33, 328–338. [Google Scholar] [CrossRef]
- Guerin, P.; El Mouatassim, S.; Menezo, Y. Oxidative stress y protección contra especies reactivas de oxígeno en el embrión preimplantacional y su entorno. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef]
- Méndez, M.S.; Argudo, D.E.; Soria, M.E.; Galarza, L.R.; Perea, F.P. Efecto de la adición de melatonina en el medio de maduración y/o vitrificación de ovocitos sobre la producción in vitro de embriones bovinos. Rev. Investig. Vet. Perú 2020, 31, e17557. [Google Scholar] [CrossRef]
- Soberano, M.A. Participación de Compuestos Tiol en el Medio de Maduración de Ovocitos de Cabra y su Efecto Sobre la Producción de Embriones In Vitro. Ph.D. Thesis, Universidad Michoacana de San Nicolás de Hidalgo, Doctorado en Biotecnología Molecular Agropecuaria, Morelia, Michoacán, Mexico, 2011. [Google Scholar]
- Sollecito, N.V.; Pereira, E.C.M.; Grázia, J.G.V.; Neves, B.P.; Couto, B.V.R.; Andrade, V.B.; Miranda, M.S.; Silva, J.K.R.; Borges, A.M. La actividad antioxidante del extracto oleoso obtenido de Lippia origanoides mejora la calidad de los embriones bovinos producidos in vitro. Arq. Bras. Med. Vet. Zootec. 2019, 71, 723–731. [Google Scholar] [CrossRef]
- Brykczynska, U.; Hisano, M.; Erkek, S.; Ramos, L.; Oakeley, E.J.; Roloff, T.C.; Beisel, C.; Schübeler, D.; Stadler, M.B.; Peters, A.H.F.M. Repressive and active histone methylation mark distinct promoters in human and mouse spermatozoa. Nat. Struct. Mol. Biol. 2010, 17, 679–687. [Google Scholar] [CrossRef]
- de Castro, L.S.; de Assis, P.M.; Siqueira, A.F.P.; Hamilton, T.R.S.; Mendes, C.M.; Losano, J.D.A.; Nichi, M.; Visintin, J.A.; Assumpção, M.E.O.A. Sperm oxidative stress is detrimental to embryo development: A dose-dependent study model and a new and more sensitive oxidative status evaluation. Oxidative Med. Cell. Longev. 2015, 2016, 8213071. [Google Scholar] [CrossRef]
- Selivanov, V.A.; Votyakova, T.V.; Pivtoraiko, V.N.; Zeak, J.; Sukhomlin, T.; Trucco, M.; Roca, J.; Cascante, M. Reactive oxygen species production by forward and reverse electron fluxes in the mitochondrial respiratory chain. PLoS Comput. Biol. 2011, 7, e1001115. [Google Scholar] [CrossRef]
- Yakes, F.M.; Van Houten, B. Mitochondrial DNA damage is more extensive and persists longer than nuclear DNA damage in human cells following oxidative stress. Proc. Natl. Acad. Sci. USA 1997, 94, 514–519. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Chinnery, P.F.; Elliott, H.R.; Hudson, G.; Samuels, D.C.; Relton, C.L. Epigenetics, epidemiology and mito-chondrial DNA diseases. Int. J. Epidemiol. 2012, 41, 177–187. [Google Scholar] [CrossRef]
- Han, Y.; Chen, J.Z. Oxidative stress induces mitochondrial dna damage and cytotoxicity through independent mechanisms in human cancer cells. BioMed Res. Int. 2013, 2013, 825065. [Google Scholar] [CrossRef]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef]
- Kirkinezos, I.G.; Moraes, C.T. Reactive oxygen species and mitochondrial diseases. Semin. Cell Dev. Biol. 2001, 12, 449–457. [Google Scholar] [CrossRef]
- Combelles, C.M.; Gupta, S.; Agarwal, A. Podría el estrés oxidativo influir en la maduración in vitro de los ovocitos? Reprod. Biomed. Online 2009, 18, 864–880. [Google Scholar] [CrossRef]
- Asiye, I.S.; Ozen, B.O.; Alper, B. Maduración in vitro de ovocitos bovinos: Efectos beneficiosos de la cisteamina. J. Dairy Vet. Anim. Res. 2018, 7, 64–65. [Google Scholar]
- Miclea, I.; Pacala, N.; Hettig, A.; Zahan, M.; Miclea, V. Alpha-tocopherol and Ascorbic Acid Combinations Influence the Maturation of Sheep Oocytes. Sci. Pap. Anim. Sci. Biotechnol. 2012, 45, 310–313. [Google Scholar]
- Sunderam, S.; Kissin, D.M.; Crawford, S.B.; Folger, S.G.; Boulet, S.L.; Warner, L.; Barfield, W.D. Assisted Reproductive Technology Surveillance—United States. Surveill. Summ. 2015, 67, 1–28. [Google Scholar]
- Lees, J.G.; Gardner, D.K.; Harvey, A.J. Pluripotent stem cell metabolism and mitochondria: Beyond ATP. Stem Cells Int. 2017, 2017, 2874283. [Google Scholar] [CrossRef]
- Vargas Reyes, J.N. Efecto de las Técnicas de Congelación Lenta y Vitrificación con Etilenglicol Sobre la Calidad Poscriopreservación de Embriones Bovinos Producidos In Vitro. Master’s Thesis, Maestría en Ciencias Veterinarias Universidad de La Salle, Facultad de Ciencias Agropecuarias, Bogotá, Colombia, 2013; pp. 1–69. Available online: https://ciencia.lasalle.edu.co/maest_ciencias_veterinarias/10 (accessed on 10 March 2022).
- Youngs, C.R. Cryopreservation of Preimplantation Embryos of Cattle, Sheep, and Goats. J. Vis. Exp. 2011, 54, e2764. [Google Scholar] [CrossRef]
- Giaretta, E.; Spinaci, M.; Bucci, D.; Tamanini, C.; Galeati, G. Effects of resveratrol on vitrified porcine oocytes. Oxidative Med. Cell. Longev. 2013, 2013, 920257. [Google Scholar] [CrossRef]
- Salzano, A.; Albero, G.; Zullo, G.; Neglia, G.; Abdel-Wahab, A.; Bifulco, G.; Zicarelli, L.; Gasparrini, B. Effect of resveratrol supplementation during culture on the quality and cryotolerance of bovine in vitro produced embryos. Anim. Reprod. Sci. 2014, 151, 91–96. [Google Scholar] [CrossRef]
- Battin, E.E.; Brumaghim, J.L. Antioxidant activity of sulfur and selenium: A review of reactive oxygen species scavenging, glutathione peroxidase, and metal-binding antioxidant mechanisms. Cell Biochem. Biophys. 2009, 55, 1–23. [Google Scholar] [CrossRef]
- de Matos, D.G.; Gasparrini, B.; Pasqualini, S.R.; Thompson, J.G. Effect of glutathione synthesis stimulation during in vitro maturation of ovine oocytes on embryo development and intracellular peroxide content. Theriogenology 2002, 57, 1443–1451. [Google Scholar] [CrossRef]
- Zuelke, K.A.; Jeffay, S.C.; Zucker, R.M.; Perreault, S.D. Glutathione (GSH) concentrations vary with the cell cycle in maturing hamster oocytes, zygotes, and pre-implantation stage embryos. Mol. Reprod. Dev. 2003, 64, 106–112. [Google Scholar] [CrossRef]
- De Matos, D.G.; Furnus, C.C.; Moses, D.F.; Martinez, A.G.; Matkovic, M. Stimulation of glutathione synthesis of in vitro matured bovine oocytes and its effect on embryo development and freezability. Mol. Reprod. Dev. 1996, 45, 451–457. [Google Scholar] [CrossRef]
- de Matos, D.G.; Furnus, C.C.; Moses, D.F. Glutathione Synthesis during in vitro maturation of bovine oocytes: Role of cumulus cells. Biol. Reprod. 1997, 57, 1420–1425. [Google Scholar] [CrossRef]
- Hernández Martínez, S.; Hernández Pichardo, J.E.; Vazquez Avendaño, J.R.; Ambríz García, D.A.; Navarro Maldonado, M.C. Developmental dynamics of cloned Mexican bighorn sheep embryos using morphological quality standards. Vet. Med. Sci. 2020, 6, 382–392. [Google Scholar] [CrossRef]
- Lorenzo-Torres, A.; Rangel-Santos, R.; Ruíz-Flores, A.; Ambríz-García, D.A. In vitro embryo production from ewes at different physiological stages. J. Vet. Sci. 2022, 23, e87. [Google Scholar] [CrossRef]
- Anzalone, D.A.; Palazzese, L.; Czernik, M.; Sabatucci, A.; Valbonetti, L.; Capra, E.; Loi, P. Controlled spermatozoa–oocyte interaction improves embryo quality in sheep. Sci. Rep. 2021, 11, 22629. [Google Scholar] [CrossRef]
- Mastrorocco, A.; Cacopardo, L.; Lamanna, D.; Temerario, L.; Brunetti, G.; Carluccio, A.; Robbe, D.; Dell’Aquila, M.E. Bioengineering approaches to improve in vitro performance of prepubertal lamb oocytes. Cells 2021, 10, 1458. [Google Scholar] [CrossRef]
- Nadri, T.; Towhidi, A.; Zeinoaldini, S.; Riazi, G.; Sharafi, M.; Zhandi, M.; Kastelic, J.; Gholami, D. Supplementation of freezing medium with encapsulated or free glutathione during cryopreservation of bull sperm. Reprod. Domest. Anim. Zuchthyg. 2022, 57, 515–523. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resveratrol | 0 µM | 0.25 µM | 0.5 µM | 1 µM |
|---|---|---|---|---|
| IVM | 75 ± 6.5 a | 74 ± 8.5 a | 81 ± 6.1 b | 81 ± 6.5 b |
| Cleavage (IVD) | 63.1 ± 5.4 a | 60 ± 4.6 a | 68.6 ± 4.7 b | 69.2 ± 4.7 b |
| Early Blastocysts | 8.4 ± 3.4 a | 13.5 ± 4.1 a | 16.2 ± 6 a | 16.8 ± 6.5 a |
| Late Blastocysts | 30.2 ± 3.9 a | 25.6 ± 3.1 a | 30.2 ± 3.5 a | 31.4 ± 4.5 a |
| Resveratrol | ||||
|---|---|---|---|---|
| Late Blastocysts | 0 µM | 0.25 µM | 0.5 µM | 1 µM |
| Mean ± SE | 58 ± 20.9 a | 41 ± 23.6 a | 59 ± 7.3 a | 64 ± 13.8 a |
| Variable | Resveratrol Concentration | |||
|---|---|---|---|---|
| Number of nuclei | 0 µM | 0.25 µM | 0.5 µM | 1 µM |
| Mean ± SE | 267 ± 28.8 a | 260 ± 33.2 ac | 185 ± 11.7 bc | 354 ± 21.5 d |
| Variable | Resveratrol Concentration | |||
|---|---|---|---|---|
| Number of nuclei | 0 µM | 0.25 µM | 0.5 µM | 1 µM |
| Mean ± SD | 109 ± 4.3 a | 90 ± 12.3 a | 147 ± 21 b | 150 ± 19.4 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Garzón, A.C.; Ramón-Ugalde, J.P.; Ambríz-García, D.A.; Vazquez-Avendaño, J.R.; Hernández-Pichardo, J.E.; Rodríguez-Suastegui, J.L.; Cortez-Romero, C.; Navarro-Maldonado, M.d.C. Resveratrol Reduces ROS by Increasing GSH in Vitrified Sheep Embryos. Animals 2023, 13, 3602. https://doi.org/10.3390/ani13233602
González-Garzón AC, Ramón-Ugalde JP, Ambríz-García DA, Vazquez-Avendaño JR, Hernández-Pichardo JE, Rodríguez-Suastegui JL, Cortez-Romero C, Navarro-Maldonado MdC. Resveratrol Reduces ROS by Increasing GSH in Vitrified Sheep Embryos. Animals. 2023; 13(23):3602. https://doi.org/10.3390/ani13233602
Chicago/Turabian StyleGonzález-Garzón, Andrea Carolina, Julio Porfirio Ramón-Ugalde, Demetrio Alonso Ambríz-García, José Roberto Vazquez-Avendaño, José Ernesto Hernández-Pichardo, José Luis Rodríguez-Suastegui, César Cortez-Romero, and María del Carmen Navarro-Maldonado. 2023. "Resveratrol Reduces ROS by Increasing GSH in Vitrified Sheep Embryos" Animals 13, no. 23: 3602. https://doi.org/10.3390/ani13233602