


Genetic Variance Estimation over Time in Broiler Breeding Programmes for Growth and Reproductive Traits

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Dataset
2.2. Multivariate Model and Computation of Variances
2.3. Sliding Overlapping Windows
2.4. Expected Variance Components
2.5. Doubling Traits to Assess Covariances and Correlations
3. Results
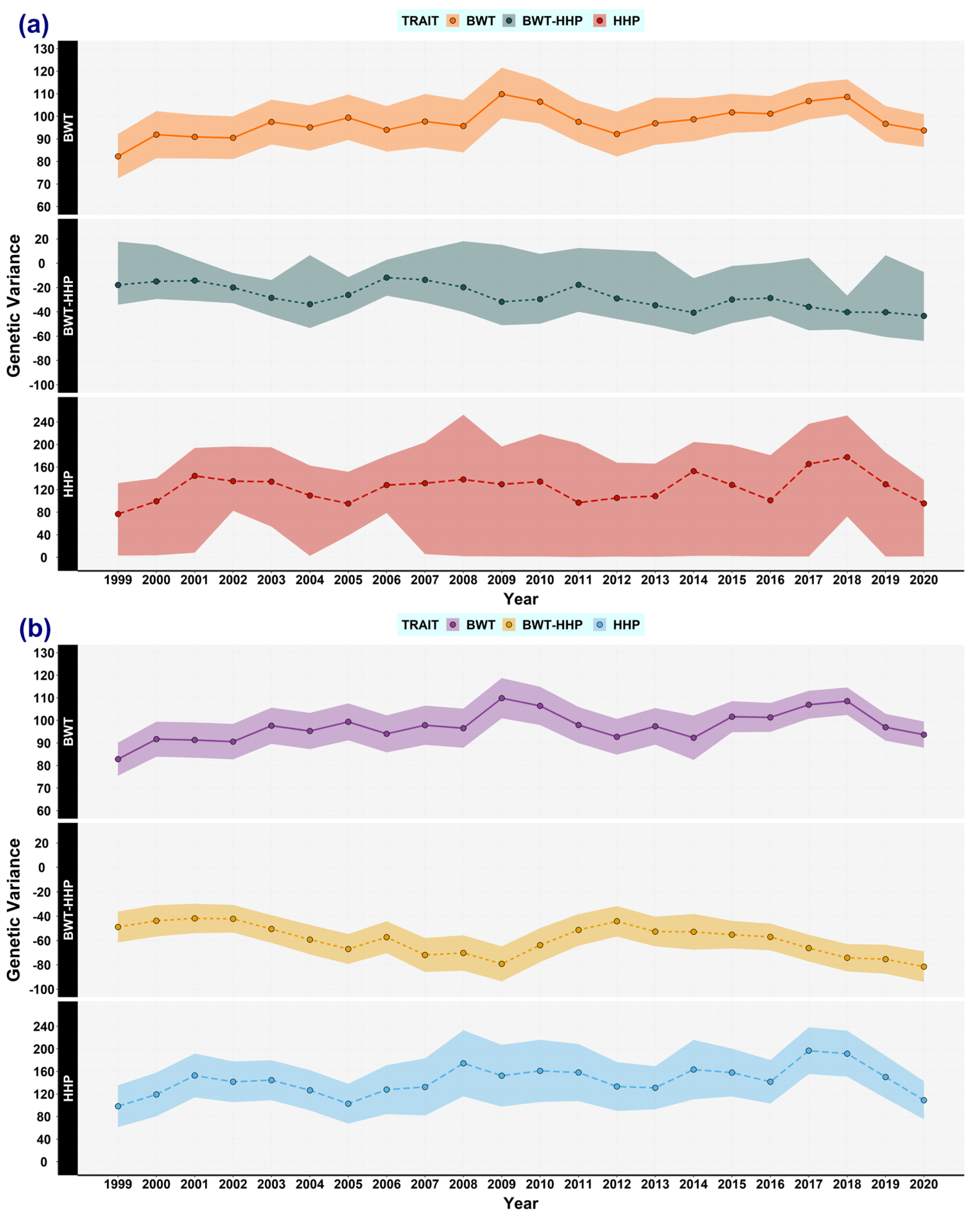
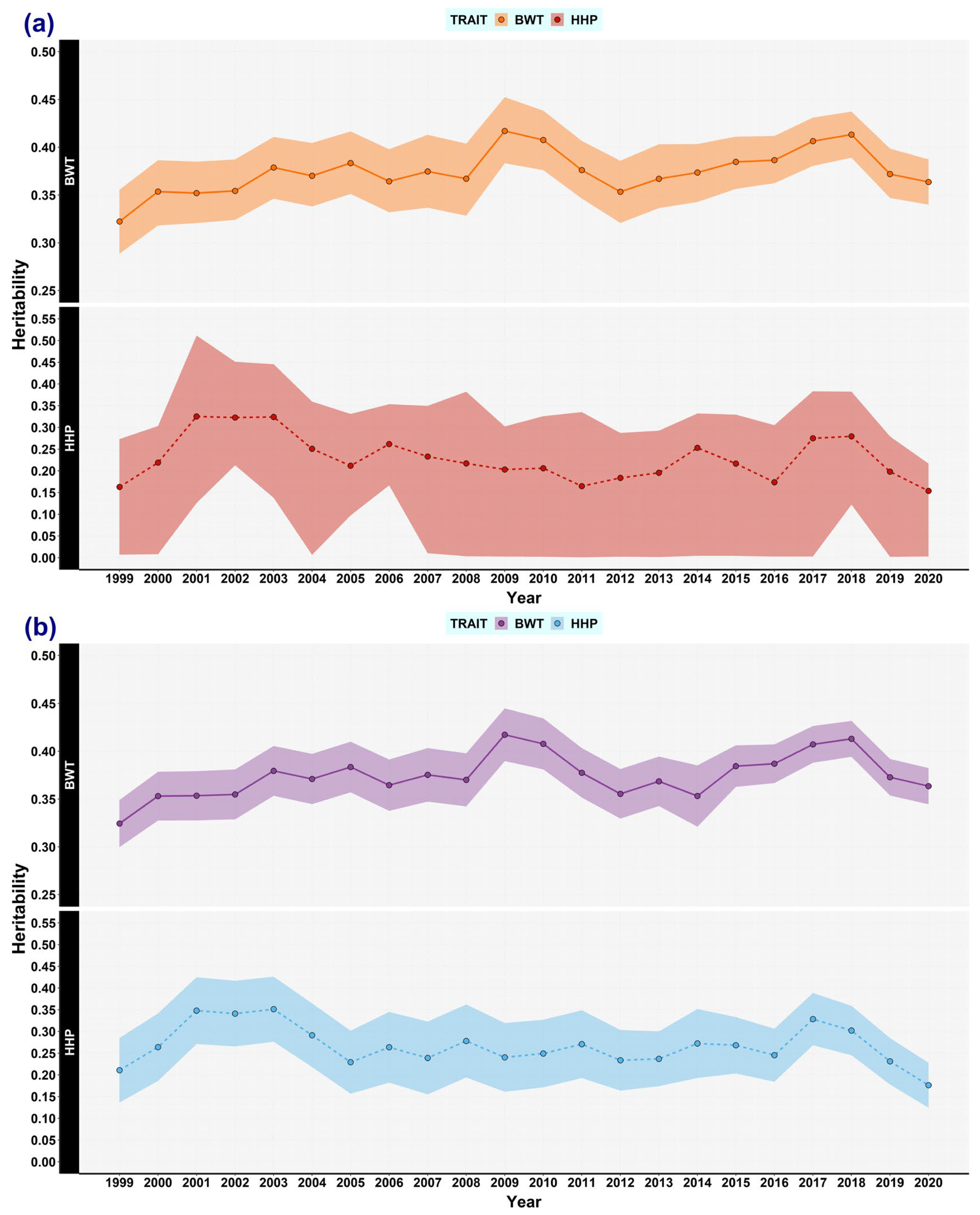
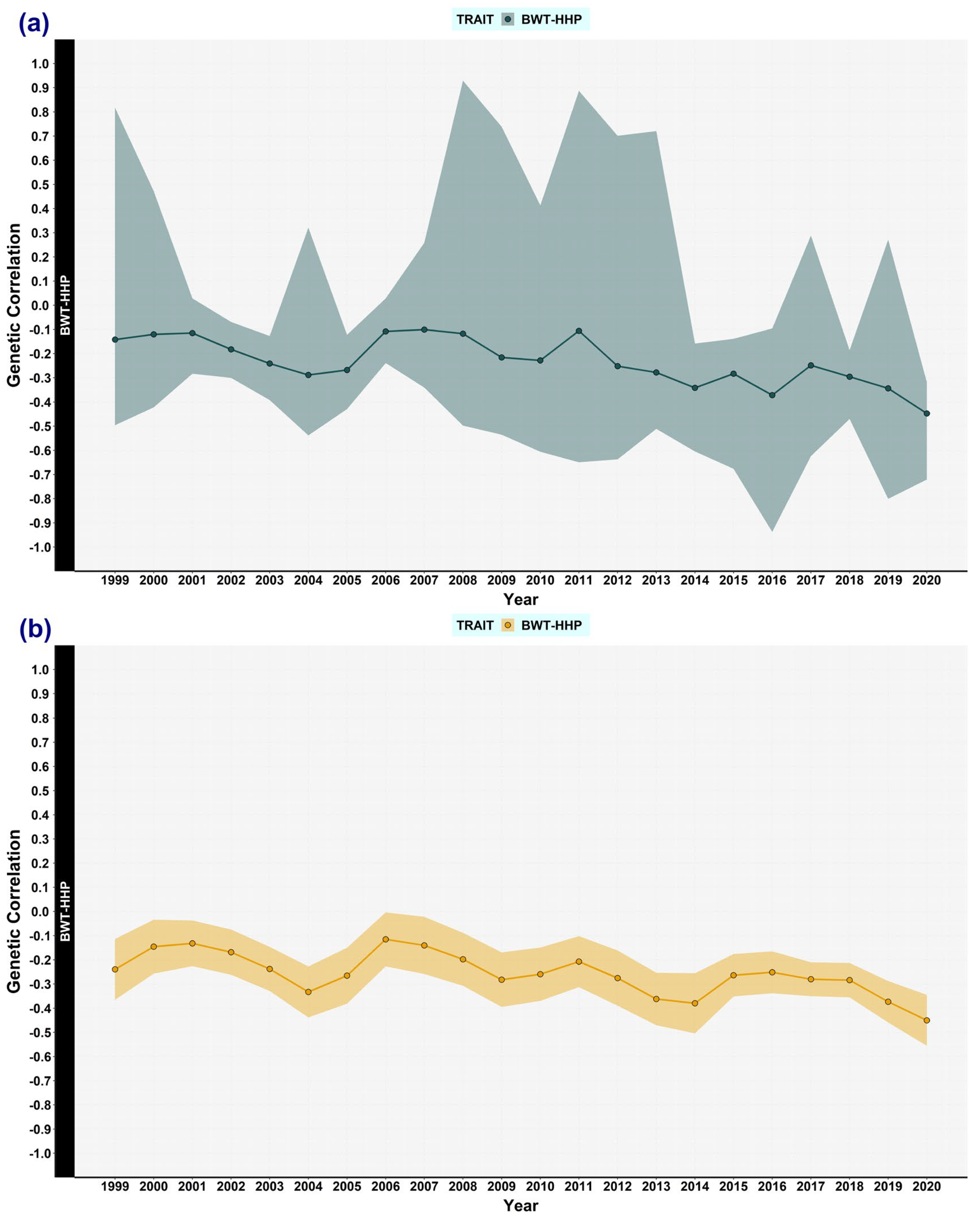
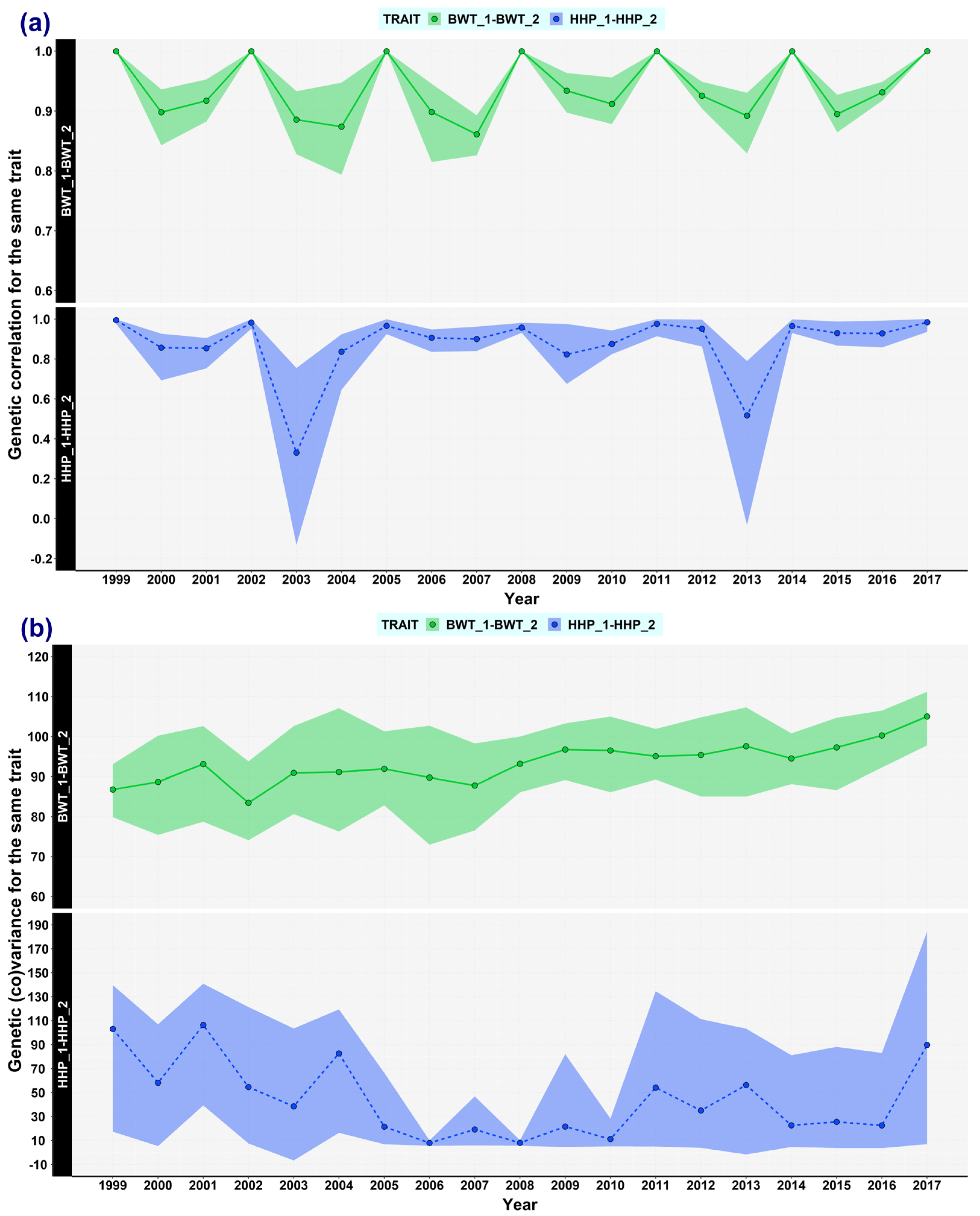
3.1. Genetic (Co)variances, Heritability, and Correlation Genetics



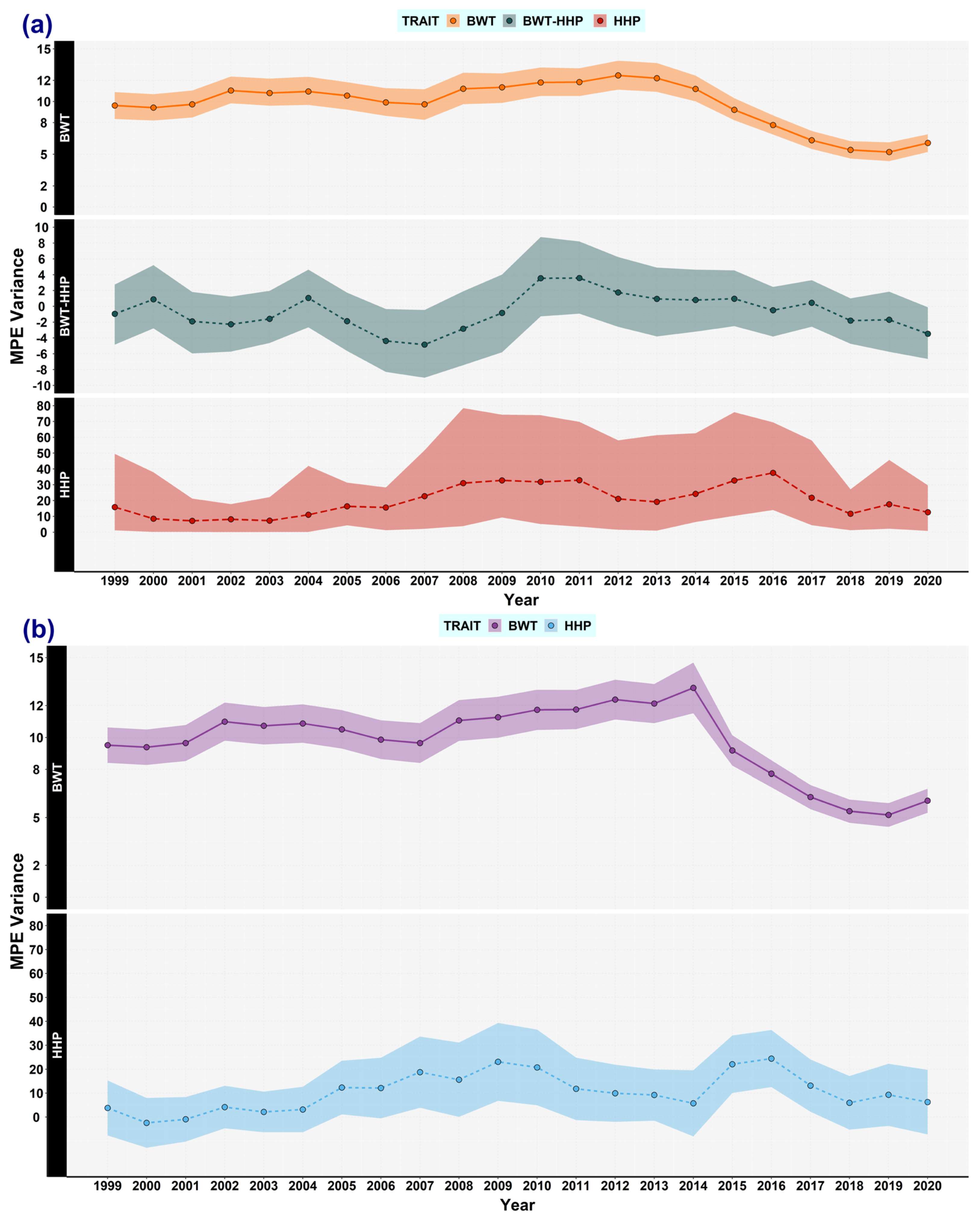
3.2. Maternal Permanent Environmental Effects

3.3. Residual and Phenotypic Variances
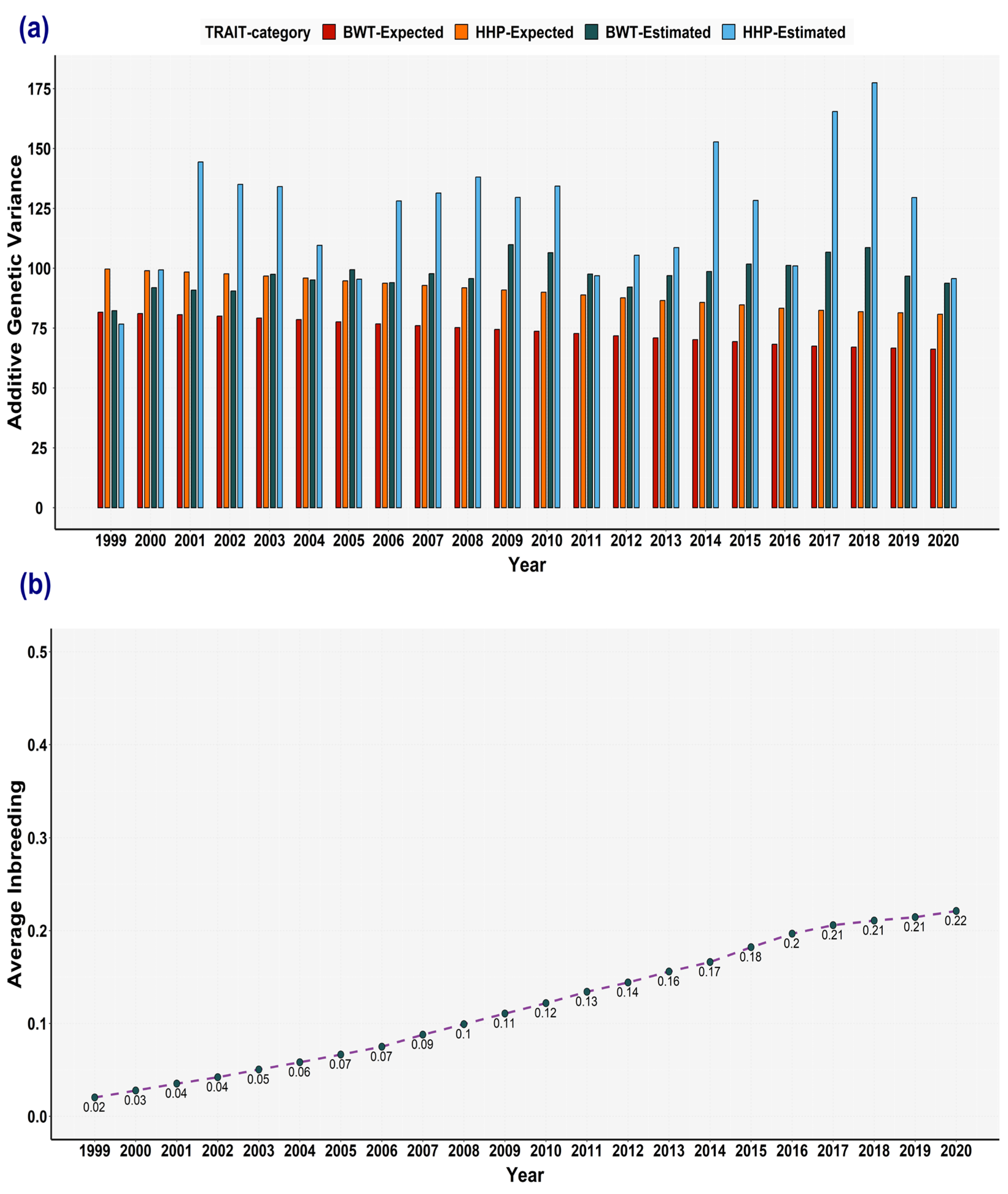
3.4. Inbreeding Effect on the Computation of Variance Components
3.5. Expected Variances and the Broiler Selection Effect on Genetic Variances

3.6. Changes Examined by Using Double Trait Covariance Analysis

4. Discussion
4.1. Genetic, Inbreeding, Coancestry, and Drift Parameters
4.2. Monitoring the Dynamics of Genetic Variance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Walsh, B.; Lynch, M. Evolution and Selection of Quantitative Traits; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Charlesworth, B.; Charlesworth, D. Elements of Evolutionary Genetics; Roberts and Company Publishers: Greenwood, CO, USA, 2010. [Google Scholar]
- Bulmer, M.G. The Effect of Selection on Genetic Variability. Am. Nat. 1971, 105, 201–211. [Google Scholar] [CrossRef]
- Falconer, D.S.; Mackay, T.F. Introduction to Quantitative Genetics, 4th ed.; Longman: Essex, UK, 1996. [Google Scholar]
- Mulder, H.A.; Lee, S.H.; Clark, S.; Hayes, B.J.; Van Der Werf, J.H.J. The Impact of Genomic and Traditional Selection on the Contribution of Mutational Variance to Long-Term Selection Response and Genetic Variance. Genetics 2019, 213, 361–378. [Google Scholar] [CrossRef]
- Chapuis, H.; Ducrocq, V.; Phocas, F.; Delabrosse, Y. Modelling and Optimizing of Sequential Selection Schemes: A Poultry Breeding Application. Genet. Sel. Evol. 1997, 29, 327. [Google Scholar] [CrossRef]
- Gorjanc, G.; Bijma, P.; Hickey, J.M. Reliability of Pedigree-Based and Genomic Evaluations in Selected Populations. Genet. Sel. Evol. 2015, 47, 65. [Google Scholar] [CrossRef]
- Sorensen, D.; Fernando, R.; Gianola, D. Inferring the Trajectory of Genetic Variance in the Course of Artificial Selection. Genet. Res. 2001, 77, 83–94. [Google Scholar] [CrossRef]
- Legarra, A. Comparing Estimates of Genetic Variance across Different Relationship Models. Theor. Popul. Biol. 2016, 107, 26–30. [Google Scholar] [CrossRef]
- Fernández, E.N.; Legarra, A.; Martínez, R.; Sánchez, J.P.; Baselga, M. Pedigree-Based Estimation of Covariance between Dominance Deviations and Additive Genetic Effects in Closed Rabbit Lines Considering Inbreeding and Using a Computationally Simpler Equivalent Model. J. Anim. Breed. Genet. 2017, 134, 184–195. [Google Scholar] [CrossRef]
- Hidalgo, J.; Tsuruta, S.; Lourenco, D.; Masuda, Y.; Huang, Y.; Gray, K.A.; Misztal, I. Changes in Genetic Parameters for Fitness and Growth Traits in Pigs under Genomic Selection. J. Anim. Sci. 2020, 98, skaa032. [Google Scholar] [CrossRef]
- Macedo, F.L.; Christensen, O.F.; Legarra, A. Selection and Drift Reduce Genetic Variation for Milk Yield in Manech Tête Rousse Dairy Sheep. JDS Commun. 2021, 2, 31–34. [Google Scholar] [CrossRef]
- Tarsani, E.; Kranis, A.; Maniatis, G.; Theodorides, A.L.H. Detection of Loci Exhibiting Pleiotropic Effects on Body Weight and Egg Number in Female Broilers. Sci. Rep. 2021, 11, 7441. [Google Scholar] [CrossRef]
- Butler, D.G.; Cullis, B.R.; Gilmour, A.R.; Gogel, B.J.; Thompson, R. ASReml-R Reference Manual Version 4; VSN International Ltd.: Hemel Hempstead, UK, 2017. [Google Scholar]
- Misztal, I.; Tsuruta, S.; Lourenco, D.A.L.; Masuda, Y.; Aguilar, I.; Legarra, A.; Vitezica, Z. Manual for BLUPF90 Family of Programs; University of Georgia: Athens, GA, USA, 2018. [Google Scholar]
- Cheng, H.; Fernando, R.; Garrick, D. JWAS: Julia Implementation of Whole-Genome Analysis Software. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Auckland, New Zealand, 7–11 February 2018; Volume 11, p. 859. [Google Scholar]
- Colleau, J.J. An Indirect Approach to the Extensive Calculation of Relationship Coefficients. Genet. Sel. Evol. 2002, 34, 409–421. [Google Scholar] [CrossRef]
- Colleau, J.J.; Palhière, I.; Rodríguez-Ramilo, S.T.; Legarra, A. A Fast Indirect Method to Compute Functions of Genomic Relationships Concerning Genotyped and Ungenotyped Individuals, for Diversity Management. Genet. Sel. Evol. 2017, 49, 87. [Google Scholar] [CrossRef]
- Haile-Mariam, M.; Pryce, J.E. Variances and Correlations of Milk Production, Fertility, Longevity, and Type Traits over Time in Australian Holstein Cattle. J. Dairy Sci. 2015, 98, 7364–7379. [Google Scholar] [CrossRef]
- Hill, W.G. Is Continued Genetic Improvement of Livestock Sustainable? Genetics 2016, 202, 877–881. [Google Scholar] [CrossRef]
- Chu, T.T.; Madsen, P.; Norberg, E.; Wang, L.; Marois, D.; Henshall, J.; Jensen, J. Genetic Analysis on Body Weight at Different Ages in Broiler Chicken Raised in Commercial Environment. J. Anim. Breed. Genet. 2020, 137, 245–259. [Google Scholar] [CrossRef]
- Kapell, D.N.R.G.; Hill, W.G.; Neeteson, A.M.; McAdam, J.; Koerhuis, A.N.M.; Avendaño, S. Genetic Parameters of Foot-Pad Dermatitis and Body Weight in Purebred Broiler Lines in 2 Contrasting Environments. Poult. Sci. 2012, 91, 565–574. [Google Scholar] [CrossRef]
- Kapell, D.N.R.G.; Hill, W.G.; Neeteson, A.M.; McAdam, J.; Koerhuis, A.N.M.; Avendaño, S. Twenty-Five Years of Selection for Improved Leg Health in Purebred Broiler Lines and Underlying Genetic Parameters. Poult. Sci. 2012, 91, 3032–3043. [Google Scholar] [CrossRef]
- Momen, M.; Mehrgardi, A.A.; Sheikhy, A.; Esmailizadeh, A.; Fozi, M.A.; Kranis, A.; Valente, B.D.; Rosa, G.J.M.; Gianola, D. A Predictive Assessment of Genetic Correlations between Traits in Chickens Using Markers. Genet. Sel. Evol. 2017, 49, 16. [Google Scholar] [CrossRef]
- Maniatis, G.; Demiris, N.; Kranis, A.; Banos, G.; Kominakis, A. Model Comparison and Estimation of Genetic Parameters for Body Weight in Commercial Broilers. Can. J. Anim. Sci. 2013, 93, 67–77. [Google Scholar] [CrossRef]
- Mebratie, W.; Madsen, P.; Hawken, R.; Romé, H.; Marois, D.; Henshall, J.; Bovenhuis, H.; Jensen, J. Genetic Parameters for Body Weight and Different Definitions of Residual Feed Intake in Broiler Chickens. Genet. Sel. Evol. 2019, 51, 53. [Google Scholar] [CrossRef]
- Mebratie, W.; Madsen, P.; Hawken, R.; Jensen, J. Multi-Trait Estimation of Genetic Parameters for Body Weight in a Commercial Broiler Chicken Population. Livest. Sci. 2018, 217, 15–18. [Google Scholar] [CrossRef]
- Rajkumar, U.; Leslie Leo Prince, L.; Rajaravindra, K.S.; Haunshi, S.; Niranjan, M.; Chatterjee, R.N. Analysis of (Co) Variance Components and Estimation of Breeding Value of Growth and Production Traits in Dahlem Red Chicken Using Pedigree Relationship in an Animal Model. PLoS ONE 2021, 16, e0247779. [Google Scholar] [CrossRef]
- Koerhuis, A.N.M.; McKay, J.C. Restricted Maximum Likelihood Estimation of Genetic Parameters for Egg Production Traits in Relation to Juvenile Body Weight in Broiler Chickens. Livest. Prod. Sci. 1996, 46, 117–127. [Google Scholar] [CrossRef]
- Begli, H.E.; Torshizi, R.V.; Masoudi, A.A.; Ehsani, A.; Jensen, J. Longitudinal Analysis of Body Weight, Feed Intake and Residual Feed Intake in F2 Chickens. Livest. Sci. 2016, 184, 28–34. [Google Scholar] [CrossRef]
- Sheng, Z.; Pettersson, M.E.; Honaker, C.F.; Siegel, P.B.; Carlborg, Ö. Standing Genetic Variation as a Major Contributor to Adaptation in the Virginia Chicken Lines Selection Experiment. Genome Biol. 2015, 16, 219. [Google Scholar] [CrossRef]
- Mebratie, W.; Shirali, M.; Madsen, P.; Sapp, R.L.; Hawken, R.; Jensen, J. The Effect of Selection and Sex on Genetic Parameters of Body Weight at Different Ages in a Commercial Broiler Chicken Population. Livest. Sci. 2017, 204, 78–87. [Google Scholar] [CrossRef]
- Tongsiri, S.; Jeyaruban, G.M.; Hermesch, S.; van der Werf, J.H.J.; Li, L.; Chormai, T. Genetic Parameters and Inbreeding Effects for Production Traits of Thai Native Chickens. Asian Australasian J. Anim. Sci. 2019, 32, 930–938. [Google Scholar] [CrossRef]
- Luo, P.T.; Yang, R.Q.; Yang, N. Estimation of Genetic Parameters for Cumulative Egg Numbers in a Broiler Dam Line by Using a Random Regression Model. Poult. Sci. 2007, 86, 30–36. [Google Scholar] [CrossRef]
- Farzin, N.; Vaez Torshizi, R.; Gerami, A.; Seraj, A. Estimates of Genetic Parameters for Monthly Egg Production in a Commercial Female Broiler Line Using Random Regression Models. Livest. Sci. 2013, 153, 33–38. [Google Scholar] [CrossRef]
- Tsuruta, S.; Misztal, I.; Lawlon, T.J.; Klei, L. Estimation of Changes of Genetic Parameters over Time for Type Traits in Holstein Using Random Regression Models. In Proceedings of the 7th World Congress on Genetics Aplied to Livestock Production, Montpellier, France, 19–23 August 2002; pp. 17–20. [Google Scholar]
- Dana, N.; vander Waaij, E.H.; van Arendonk, J.A.M. Genetic and Phenotypic Parameter Estimates for Body Weights and Egg Production in Horro Chicken of Ethiopia. Trop. Anim. Health Prod. 2011, 43, 21–28. [Google Scholar] [CrossRef]
- Prince, L.L.L.; Rajaravindra, K.S.; Rajkumar, U.; Reddy, B.L.N.; Paswan, C.; Haunshi, S.; Chatterjee, R.N. Genetic Analysis of Growth and Egg Production Traits in Synthetic Colored Broiler Female Line Using Animal Model. Trop. Anim. Health Prod. 2020, 52, 3153–3163. [Google Scholar] [CrossRef]
- Abdollahi-Arpanahi, R.; Morota, G.; Valente, B.D.; Kranis, A.; Rosa, G.J.M.; Gianola, D. Differential Contribution of Genomic Regions to Marked Genetic Variation and Prediction of Quantitative Traits in Broiler Chickens. Genet. Sel. Evol. 2016, 48, 10. [Google Scholar] [CrossRef]
- Misztal, I.; Aguilar, I.; Lourenco, D.; Ma, L.; Steibel, J.P.; Toro, M. Emerging Issues in Genomic Selection. J. Anim. Sci. 2021, 99, skab092. [Google Scholar] [CrossRef]
- Neeteson-van Nieuwenhoven, A.M.; Knap, P.; Avendaño, S. The Role of Sustainable Commercial Pig and Poultry Breeding for Food Security. Anim. Front. 2013, 3, 52–57. [Google Scholar] [CrossRef]
- Fernández, E.N.; Sánchez, J.P.; Martínez, R.; Legarra, A.; Baselga, M. Role of Inbreeding Depression, Non-Inbred Dominance Deviations and Random Year-Season Effect in Genetic Trends for Prolificacy in Closed Rabbit Lines. J. Anim. Breed. Genet. 2017, 134, 441–452. [Google Scholar] [CrossRef]
- FAO. Secondary Guidelines for Development of National Farm Animal Genetic Resources Management Plans. In Management of Small Populations at Risk; FAO: Québec City, Canada, 1998; p. 63. [Google Scholar]
- Villanueva, B.; Kennedy, B.W. Effect of Selection on Genetic Parameters of Correlated Traits. Theor. Appl. Genet. 1990, 80, 746–752. [Google Scholar] [CrossRef]
- Villanueva, B.; Wray, N.R.; Thompson, R. Prediction of Asymptotic Rates of Response from Selection on Multiple Traits Using Univariate and Multivariate Best Linear Unbiased Predictors. Anim. Prod. 1993, 57, 1–13. [Google Scholar] [CrossRef]
- Svitáková, A.; Bauer, J.; Přibyl, J.; Veselá, Z.; Vostrý, L. Changes over Time in Genetic Parameters for Growth in Bulls and Assessment of Suitability of Test Methods. Czech J. Anim. Sci. 2014, 59, 19–25. [Google Scholar] [CrossRef]
- Tsuruta, S.; Misztal, I.; Lawlor, T.J. Genetic Correlations among Production, Body Size, Udder, and Productive Life Traits over Time in Holsteins. J. Dairy Sci. 2004, 87, 1457–1468. [Google Scholar] [CrossRef]
- Renema, R.A.; Rustad, M.E.; Robinson, F.E. Implications of Changes to Commercial Broiler and Broiler Breeder Body Weight Targets over the Past 30 Years. Worlds. Poult. Sci. J. 2007, 63, 457–472. [Google Scholar] [CrossRef]
- Lillie, M.; Sheng, Z.Y.; Honaker, C.F.; Andersson, L.; Siegel, P.B.; Carlborg, Ö. Genomic Signatures of 60 Years of Bidirectional Selection for 8-Week Body Weight in Chickens. Poult. Sci. 2018, 97, 781–790. [Google Scholar] [CrossRef]
- Lillie, M.; Honaker, C.F.; Siegel, P.B.; Carlborg, Ö. Bidirectional Selection for Body Weight on Standing Genetic Variation in a Chicken Model. G3 Genes Genomes Genet. 2019, 9, 1165–1173. [Google Scholar] [CrossRef]
- Houle, D. How Should We Explain Variation in the Genetic Variance of Traits? Genetica 1998, 102–103, 241–253. [Google Scholar] [CrossRef]
- Casellas, J.; Caja, G.; Piedrafita, J. Accounting for Additive Genetic Mutations on Litter Size in Ripollesa Sheep1. J. Anim. Sci. 2010, 88, 1248–1255. [Google Scholar] [CrossRef]
- Ezzeroug, R.; Belabbas, R.; Argente, M.J.; Berbar, A.; Diss, S.; Boudjella, Z.; Talaziza, D.; Boudahdir, N.; De La Luz García, M. Genetic Correlations for Reproductive and Growth Traits in Rabbits. Can. J. Anim. Sci. 2020, 100, 317–322. [Google Scholar] [CrossRef]
- Neyhart, J.L.; Lorenz, A.J.; Smith, K.P. Multi-Trait Improvement by Predicting Genetic Correlations in Breeding Crosses. G3 Genes Genomes Genet. 2019, 9, 3153–3165. [Google Scholar] [CrossRef]
- Fikse, W.F.; Liu, Z.; Sullivan, P.G. Tolerance Values for Validation of Trends in Genetic Variances Over Time. Interbull Bull. 2005, 33, 200. [Google Scholar]
- Tyrisevä, A.M.; Fikse, W.F.; Mäntysaari, E.A.; Jakobsen, J.; Aamand, G.P.; Dürr, J.; Lidauer, M.H. Validation of Consistency of Mendelian Sampling Variance. J. Dairy Sci. 2018, 101, 2187–2198. [Google Scholar] [CrossRef]
- Lidauer, M.; Vuori, K.; Stranden, I.; Mantysaari, E. Experiences with Interbull Test IV: Estimation of Genetic Variance. Interbull Bull. 2007, 37, 69–72. [Google Scholar]
- Gao, H.; Madsen, P.; Aamand, G.P.; Thomasen, J.R.; Sørensen, A.C.; Jensen, J. Bias in Estimates of Variance Components in Populations Undergoing Genomic Selection: A Simulation Study. BMC Genomics 2019, 20, 956. [Google Scholar] [CrossRef]
- Cesarani, A.; Pocrnic, I.; Macciotta, N.P.P.; Fragomeni, B.O.; Misztal, I.; Lourenco, D.A.L. Bias in Heritability Estimates from Genomic Restricted Maximum Likelihood Methods under Different Genotyping Strategies. J. Anim. Breed. Genet. 2019, 136, 40–50. [Google Scholar] [CrossRef]
- Bussiman, F.; Chen, C.Y.; Holl, J.; Bermann, M.; Legarra, A.; Misztal, I.; Lourenco, D. Boundaries for Genotype, Phenotype, and Pedigree Truncation in Genomic Evaluations in Pigs. J. Anim. Sci. 2023, 101, skad273. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overlapping Window | Pedigree 1 | Number of Generations 5 | BWT 2 | HHP 4 | ||||
|---|---|---|---|---|---|---|---|---|
| Number 3 | Mean | SD | Number 3 | Mean | SD | |||
| 1999–2001 | 209,977 | 9 | 207,734 | 178.11 | 22.62 | 4711 | 108 | 22.46 |
| 2000–2002 | 221,795 | 9 | 219,618 | 179.10 | 22.34 | 4544 | 111 | 21.43 |
| 2001–2003 | 231,672 | 10 | 224,297 | 178.80 | 22.26 | 4908 | 113 | 21.17 |
| 2002–2004 | 229,175 | 11 | 226,534 | 179.90 | 23.06 | 5126 | 115 | 21.42 |
| 2003–2005 | 232,122 | 10 | 230,104 | 185.58 | 24.62 | 5166 | 117 | 21.04 |
| 2004–2006 | 233,058 | 11 | 230,458 | 192.90 | 24.93 | 4841 | 116 | 21.48 |
| 2005–2007 | 218,953 | 10 | 211,600 | 199.45 | 24.17 | 4983 | 115 | 21.01 |
| 2006–2008 | 203,089 | 11 | 200,852 | 200.77 | 21.13 | 4694 | 118 | 22.66 |
| 2007–2009 | 203,461 | 11 | 201,163 | 202.50 | 24.06 | 4732 | 121 | 24.05 |
| 2008–2010 | 213,624 | 11 | 211,513 | 203.85 | 24.13 | 4964 | 125 | 25.17 |
| 2009–2011 | 207,383 | 10 | 205,448 | 205.21 | 23.93 | 5199 | 125 | 25.30 |
| 2010–2012 | 214,657 | 11 | 212,462 | 207.21 | 23.77 | 5445 | 125 | 27.42 |
| 2011–2013 | 224,160 | 11 | 221,943 | 205.99 | 23.88 | 5652 | 116 | 31.09 |
| 2012–2014 | 234,220 | 10 | 231,895 | 205.87 | 24.17 | 6027 | 118 | 31.28 |
| 2013–2015 | 243,054 | 9 | 240,617 | 205.03 | 24.41 | 6412 | 119 | 29.55 |
| 2014–2016 | 288,859 | 10 | 286,084 | 206.12 | 24.37 | 7490 | 122 | 25.54 |
| 2015–2017 | 329,530 | 10 | 326,546 | 205.71 | 24.34 | 7465 | 122 | 24.99 |
| 2016–2018 | 362,340 | 10 | 359,044 | 205.26 | 23.98 | 7624 | 121 | 24.79 |
| 2017–2019 | 380,101 | 10 | 376,786 | 204.79 | 23.51 | 7971 | 123 | 25.03 |
| 2018–2020 | 361,019 | 9 | 357,614 | 205.41 | 22.86 | 8115 | 125 | 27.26 |
| 2019–2021 | 340,604 | 9 | 336,671 | 205.48 | 23.74 | 7159 | 127 | 28.18 |
| 2020–2022 | 355,976 | 10 | 351,908 | 210.48 | 25.17 | 5876 | 126 | 28.65 |
| Overlapping Window | Pedigree within Window 1 | Accumulated Pedigree 4 | ||||
|---|---|---|---|---|---|---|
| Mean | HPD95% Interval 2 | IChS 3 | Mean | HPD95% Interval 2 | IChS 3 | |
| 2000–2002 | 91.91 | [81.42, 102.30] | 182 | 90.43 | [80.81, 99.30] | 92 |
| 2001–2003 | 90.89 | [81.33, 100.70] | 142 | 90.53 | [81.98, 100.30] | 20 |
| 2002–2004 | 90.51 | [81.07, 100.00] | 64 | 91.89 | [82.66, 101.10] | 32 |
| 2003–2005 | 97.52 | [87.55, 107.40] | 228 | 97.45 | [88.28, 106.70] | 156 |
| 2004–2006 | 95.12 | [84.76, 104.90] | 114 | 96.50 | [84.36, 107.20] | 286 |
| 2005–2007 | 99.39 | [89.51, 109.70] | 114 | 95.81 | [86.53, 106.10] | 68 |
| 2006–2008 | 94.02 | [84.37, 104.60] | 50 | 92.27 | [79.32, 106.10] | 214 |
| 2007–2009 | 97.73 | [86.29, 109.90] | 224 | 92.56 | [83.16, 103.30] | 60 |
| 2008–2010 | 95.70 | [84.03, 107.30] | 544 | 94.26 | [83.93, 104.10] | 120 |
| 2009–2011 | 109.87 | [99.20, 121.60] | 332 | 103.76 | [90.31, 115.60] | 60 |
| 2010–2012 | 106.49 | [96.84, 116.60] | 110 | 110.25 | [99.45, 121.00] | 46 |
| 2011–2013 | 97.58 | [88.42, 107.00] | 174 | 103.63 | [89.92, 113.40] | 256 |
| 2012–2014 | 92.15 | [82.18, 102.10] | 134 | 97.67 | [86.74, 108.40] | 256 |
| 2013–2015 | 96.93 | [87.42, 108.30] | 272 | 101.40 | [89.96, 114.20] | 258 |
| 2014–2016 | 98.68 | [88.99, 108.10] | 36 | 98.44 | [84.85, 114.40] | 334 |
| 2015–2017 | 101.74 | [92.68, 110.00] | 32 | 109.19 | [98.05, 123.50] | 342 |
| 2016–2018 | 101.18 | [93.42, 109.00] | 40 | 112.57 | [98.33, 122.20] | 254 |
| 2017–2019 | 106.75 | [98.62, 114.80] | 482 | 119.45 | [110.40, 129.10] | 334 |
| 2018–2020 | 108.63 | [100.90, 116.40] | 68 | 123.41 | [107.00, 137.50] | 322 |
| 2019–2021 | 96.71 | [88.71, 104.60] | 482 | 108.18 | [97.88, 119.10] | 370 |
| 2020–2022 | 93.75 | [86.42, 100.90] | 206 | 105.07 | [94.59, 113.70] | 288 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sosa-Madrid, B.S.; Maniatis, G.; Ibáñez-Escriche, N.; Avendaño, S.; Kranis, A. Genetic Variance Estimation over Time in Broiler Breeding Programmes for Growth and Reproductive Traits. Animals 2023, 13, 3306. https://doi.org/10.3390/ani13213306
Sosa-Madrid BS, Maniatis G, Ibáñez-Escriche N, Avendaño S, Kranis A. Genetic Variance Estimation over Time in Broiler Breeding Programmes for Growth and Reproductive Traits. Animals. 2023; 13(21):3306. https://doi.org/10.3390/ani13213306
Chicago/Turabian StyleSosa-Madrid, Bolívar Samuel, Gerasimos Maniatis, Noelia Ibáñez-Escriche, Santiago Avendaño, and Andreas Kranis. 2023. "Genetic Variance Estimation over Time in Broiler Breeding Programmes for Growth and Reproductive Traits" Animals 13, no. 21: 3306. https://doi.org/10.3390/ani13213306