
Assessment Heartworm Disease in the Canary Islands (Spain): Risk of Transmission in a Hyperendemic Area by Ecological Niche Modeling and Its Future Projection
 , , ,
, , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
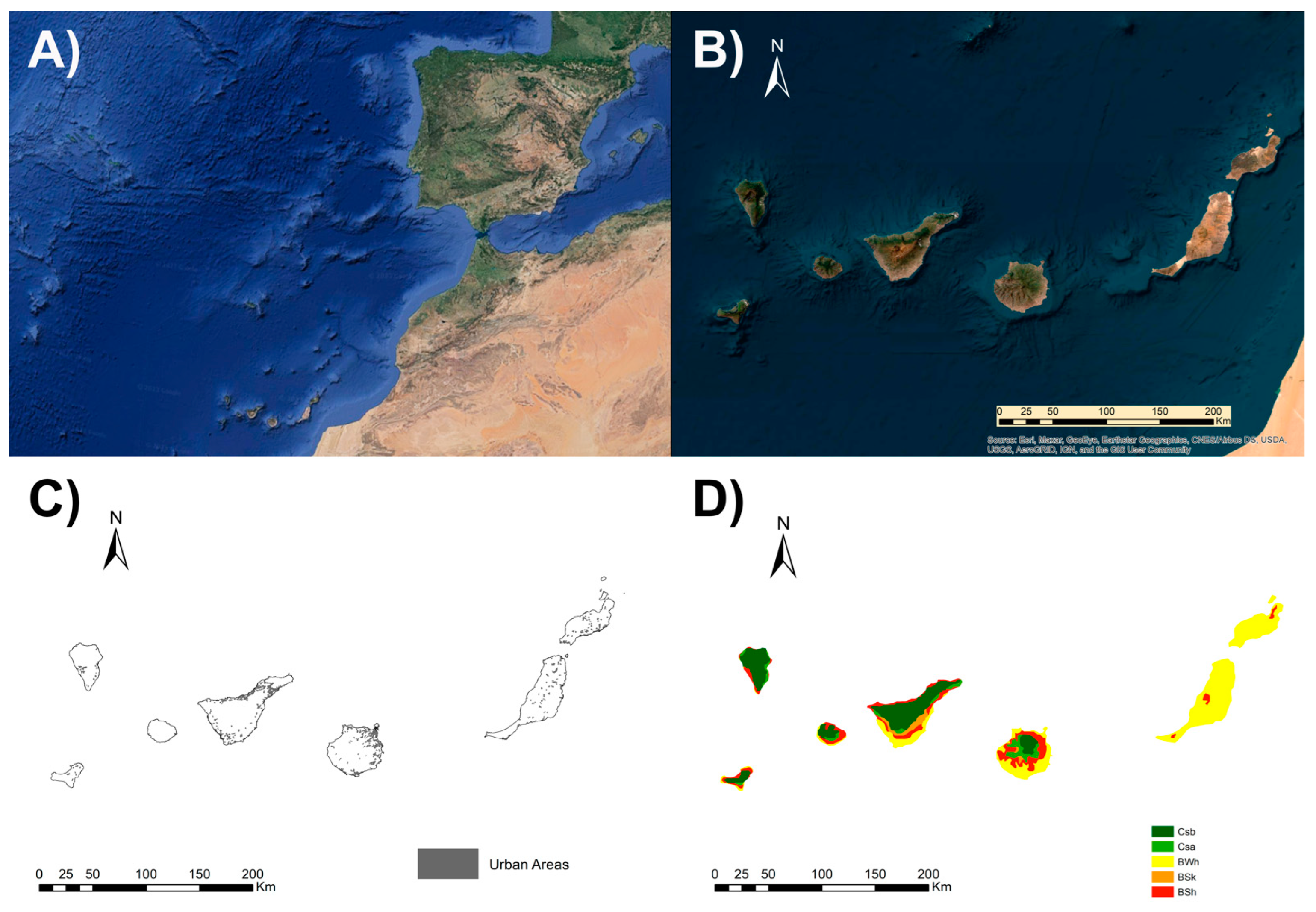
2.1. Study Area Description
2.2. Data
2.2.1. Culex spp. Collection
2.2.2. Environmental and Bioclimatic Data
2.3. Culex spp. Ecological Niche Model
2.4. Dirofilaria Immitis Generations
2.5. Dirofilaria Immitis Risk Map
2.6. Forward Projection and Rank Change Analysis
3. Results
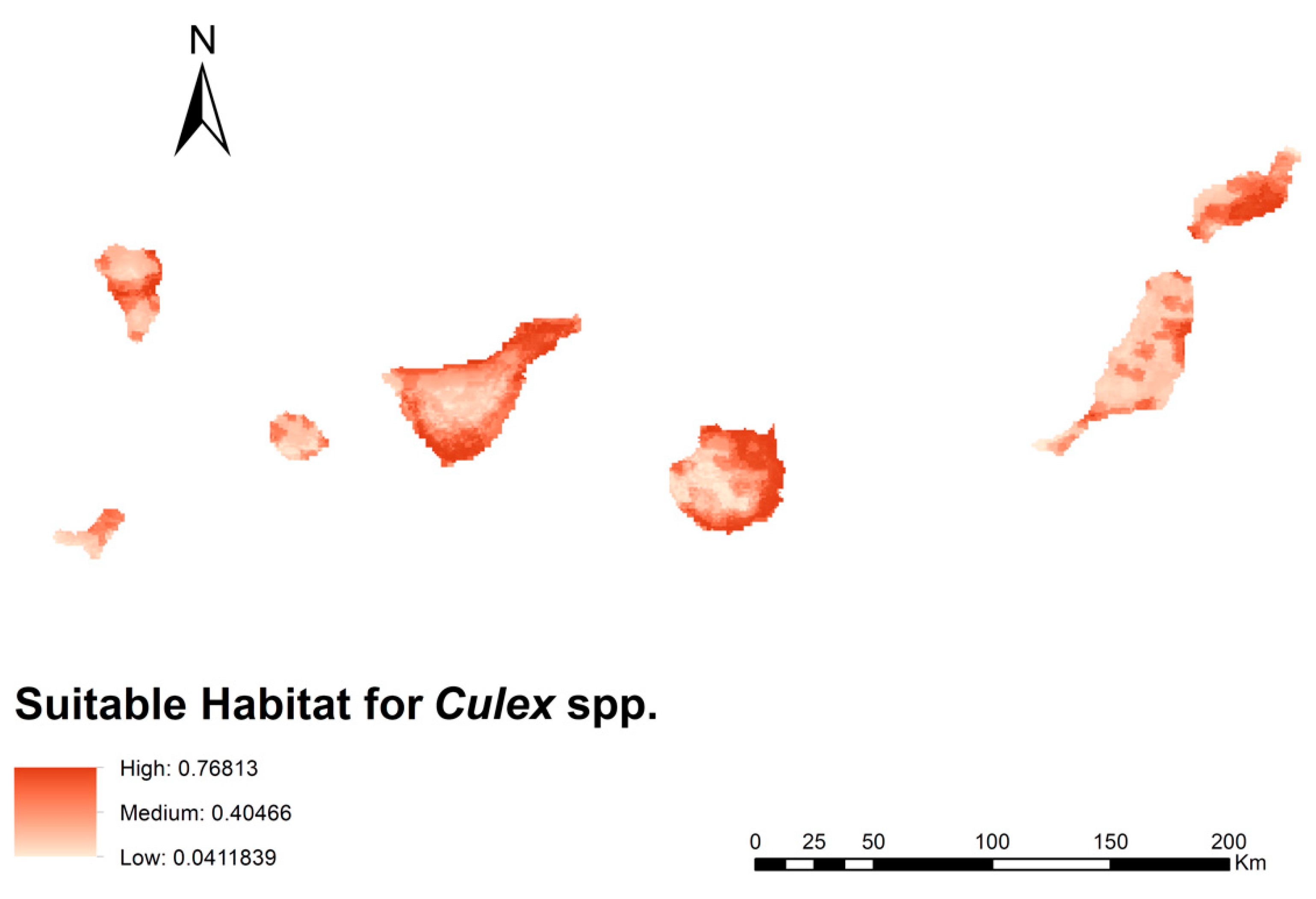
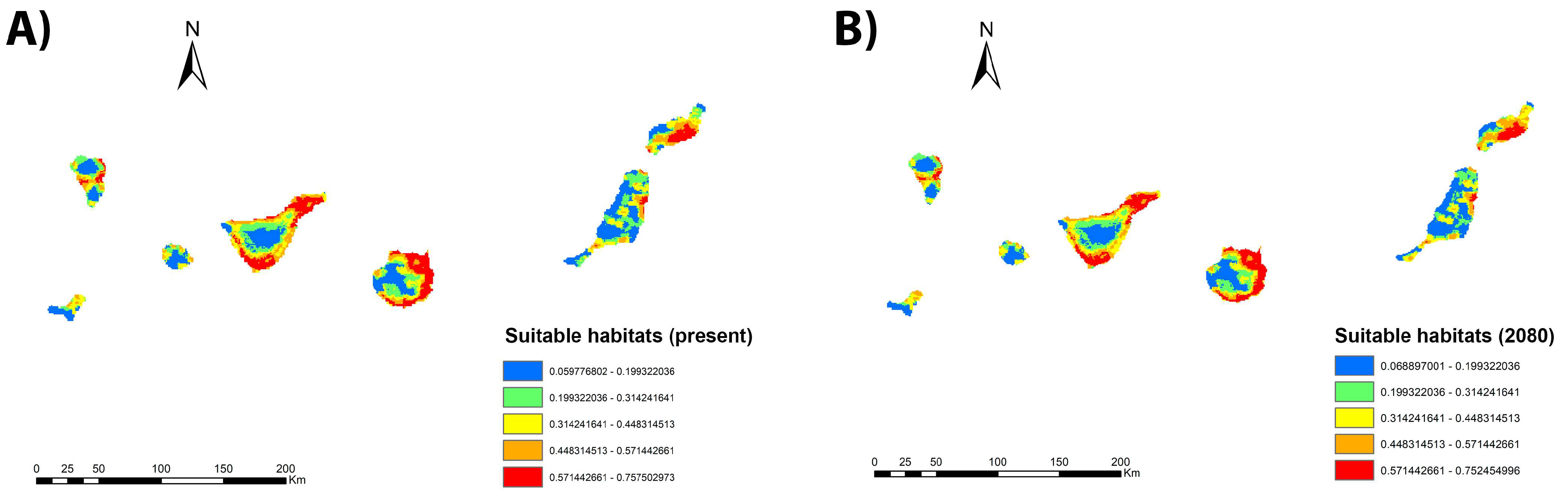
3.1. Culex spp. Habitat Suitability
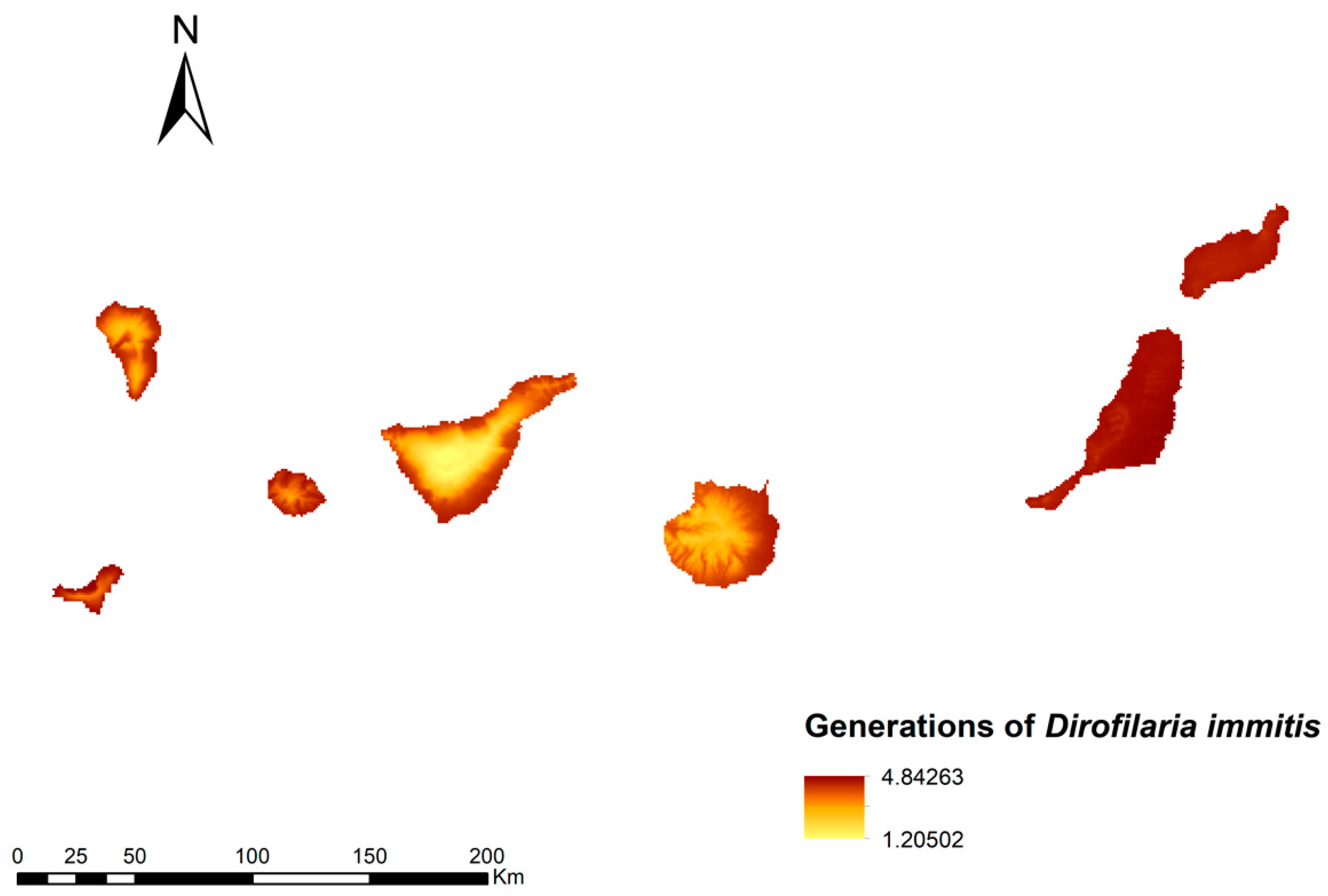
3.2. Number Dirofilaria Immitis Generations
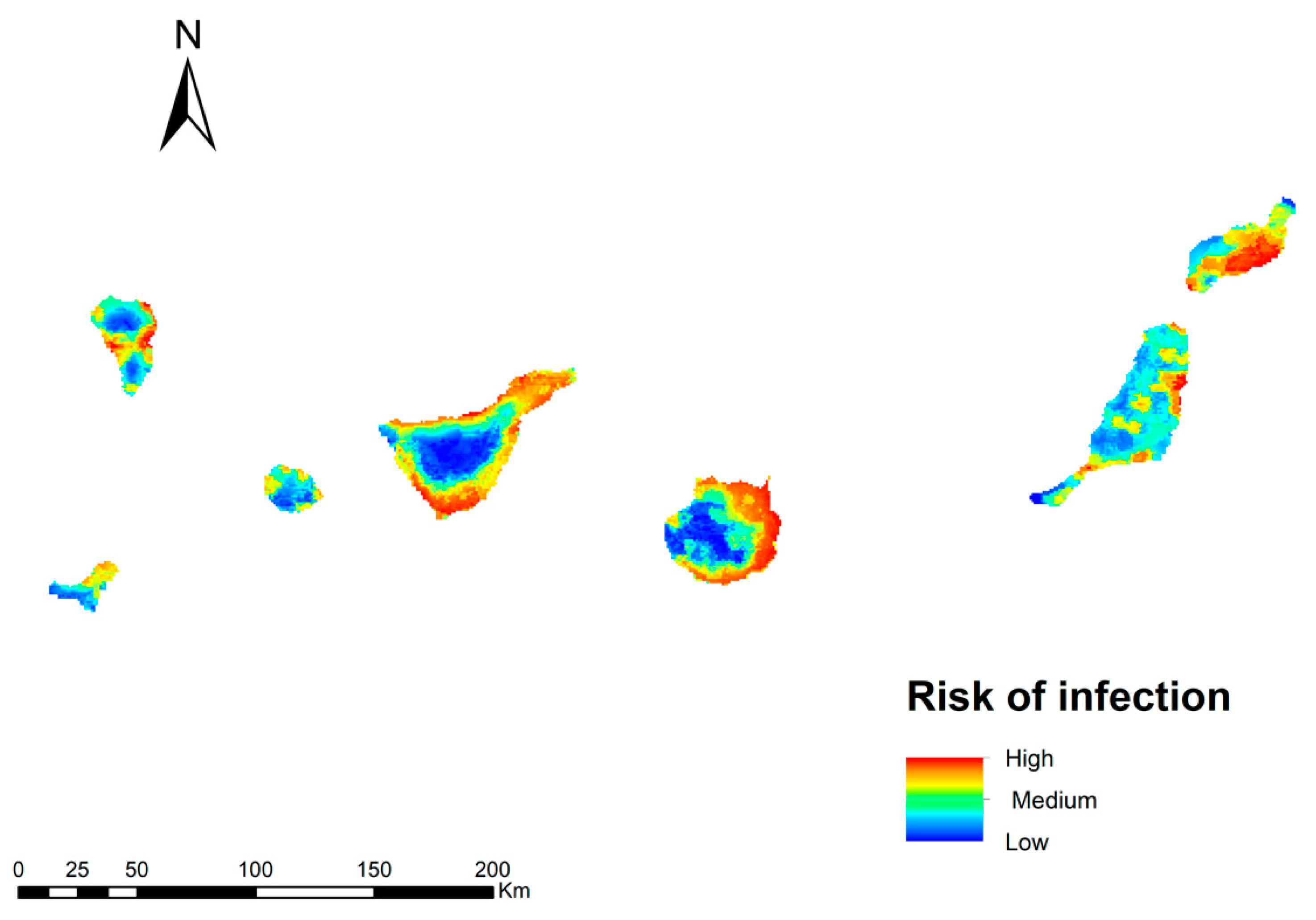
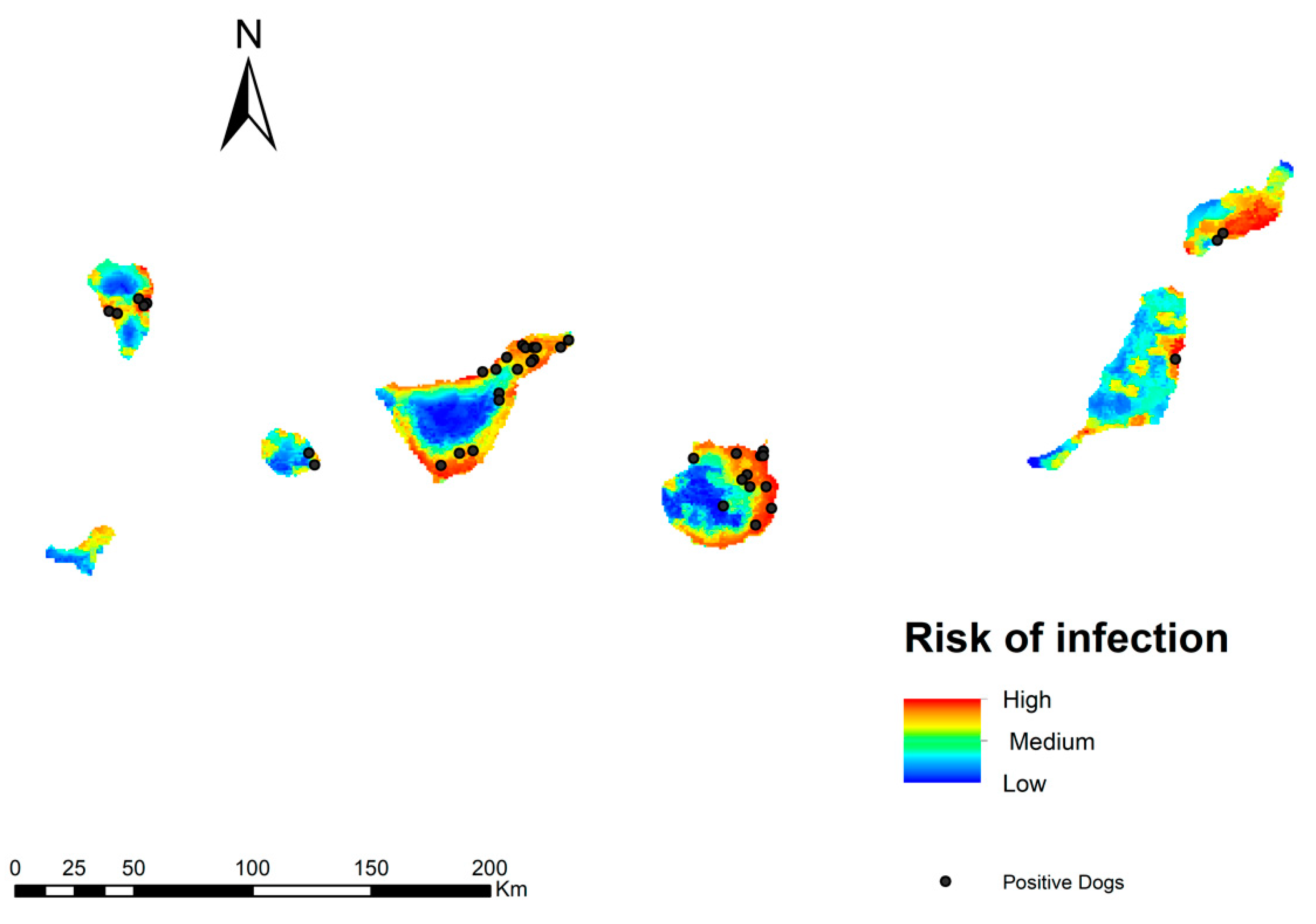
3.3. Potential Risk of Transmission of Dirofilaria Immitis
3.4. Future Projection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simón, F.; Siles-Lucas, M.; Morchón, R.; González-Miguel, J.; Mellado, I.; Carretón, E.; Montoya-Alonso, J.A. Human and animal dirofilariasis: The emergence of a zoonotic mosaic. Clin. Microbiol. Rev. 2012, 25, 507–544. [Google Scholar] [CrossRef] [PubMed]
- Capelli, G.; Genchi, C.; Baneth, G.; Bourdeau, P.; Brianti, E.; Cardoso, L.; Danesi, P.; Fuehrer, H.P.; Giannelli, A.; Ionică, A.M.; et al. Recent advances on Dirofilaria repens in dogs and humans in Europe. Parasites Vectors 2018, 11, 663. [Google Scholar] [CrossRef] [PubMed]
- Morchón, R.; Montoya-Alonso, J.A.; Rodríguez-Escolar, I.; Carretón, E. What Has Happened to Heartworm Disease in Europe in the Last 10 Years? Pathogens 2022, 11, 1042. [Google Scholar] [CrossRef]
- Cancrini, G.; Scaramozzino, P.; Gabrielli, S.; Di Paolo, M.; Toma, L.; Romi, R. Aedes albopictus and Culex pipiens implicated as natural vectors of Dirofilaria repens in central Italy. J. Med. Entomol. 2007, 44, 1064–1066. [Google Scholar] [CrossRef]
- Genchi, C.; Rinaldi, L.; Mortarino, M.; Genchi, M.; Cringoli, G. Climate and Dirofilaria infection in Europe. Vet. Parasitol. 2009, 163, 286–292. [Google Scholar] [CrossRef]
- Otranto, D.; Dantas-Torres, F.; Brianti, E.; Traversa, D.; Petrić, D.; Genchi, C.; Capelli, G. Vector-borne helminths of dogs and humans in Europe. Parasites Vectors 2013, 6, 16. [Google Scholar] [CrossRef]
- Gangoso, L.; Aragonés, D.; Martínez-de la Puente, J.; Lucientes, J.; Delacour-Estrella, S.; Estrada Peña, R.; Montalvo, T.; Bueno-Marí, R.; Bravo-Barriga, D.; Frontera, E.; et al. Determinants of the current and future distribution of the West Nile virus mosquito vector Culex pipiens in Spain. Environ. Res. 2020, 188, 109837. [Google Scholar] [CrossRef] [PubMed]
- Simón, F.; González-Miguel, J.; Diosdado, A.; Gómez, P.J.; Morchón, R.; Kartashev, V. The Complexity of Zoonotic Filariasis Episystem and Its Consequences: A Multidisciplinary View. Biomed. Res. Int. 2017, 2017, 6436130. [Google Scholar] [CrossRef]
- Montoya-Alonso, J.A.; Morchón, R.; García-Rodríguez, S.N.; Falcón-Cordón, Y.; Costa-Rodríguez, N.; Matos, J.I.; Rodríguez-Escolar, I.; Carretón, E. Expansion of Canine Heartworm in Spain. Animals 2022, 12, 1268. [Google Scholar] [CrossRef]
- Montoya-Alonso, J.A.; García Rodríguez, S.N.; Carretón, E.; Rodríguez Escolar, I.; Costa-Rodríguez, N.; Matos, J.I.; Morchón, R. Seroprevalence of Feline Heartworm in Spain: Completing the Epidemiological Puzzle of a Neglected Disease in the Cat. Front. Vet. Sci. 2022, 9, 900371. [Google Scholar] [CrossRef]
- Simón, F.; Diosdado, A.; Siles-Lucas, M.; Kartashev, V.; González-Miguel, J. Human dirofilariosis in the 21st century: A scoping review of clinical cases reported in the literature. Transbound Emerg. Dis. 2022, 69, 2424–2439. [Google Scholar] [CrossRef] [PubMed]
- Morchón, R.; Bargues, M.D.; Latorre-Estivalis, J.M.; Pou-Barreto, C.; Melero-Alcibar, R.; Moreno, M.; Valladares, B.; Molina, R.; Montoya Alonso, J.A.; Mas-Coma, S.; et al. Molecular Characterization of Culex theileri from Canary Islands.; Spain.; a potential vector of Dirofilaria immitis. J. Clin. Experiment. Pathol. 2011, S3, 001. [Google Scholar]
- Morchón, R.; Bargues, M.D.; Latorre, J.M.; Melero-Alcíbar, R.; Pou-Barreto, C.; Mas-Coma, S.; Simón, F. Haplotype H1 of Culex pipiens implicated as natural vector of Dirofilaria immitis in an endemic area of Western Spain. Vector-Borne Zoonotic. Dis. 2007, 7, 653–658. [Google Scholar] [CrossRef]
- Bravo-Barriga, D.; Parreira, R.; Almeida, A.P.; Calado, M.; Blanco-Ciudad, J.; Serrano-Aguilera, F.J.; Pérez-Martín, J.E.; Sánchez-Peinado, J.; Pinto, J.; Reina, D.; et al. Culex pipiens as a potential vector for transmission of Dirofilaria immitis and other unclassified Filarioidea in Southwest Spain. Vet. Parasitol. 2016, 223, 173–180. [Google Scholar] [CrossRef]
- Genchi, C.; Rinaldi, L.; Cascone, C.; Mortarino, M.; Cringoli, G. Is heartworm disease really spreading in Europe? Vet. Parasitol. 2005, 133, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Genchi, C.; Mortarino, M.; Rinaldi, L.; Cringoli, G.; Traldi, G.; Genchi, M. Changing climate and changing vector-borne disease distribution: The example of Dirofilaria in Europe. Vet. Parasitol. 2011, 176, 295–299. [Google Scholar] [CrossRef]
- Rinaldi, L.; Musella, V.; Biggeri, A.; Cringoli, G. New insights into the application of geographical information systems and remote sensing in veterinary parasitology. Geospat. Health. 2006, 1, 33–47. [Google Scholar] [CrossRef]
- Medlock, J.M.; Barrass, I.; Kerrod, E.; Taylor, M.A.; Leach, S. Analysis of climatic predictions for extrinsic incubation of Dirofilaria in the United Kingdom. Vector-Borne Zoonotic. Dis. 2007, 7, 4–14. [Google Scholar] [CrossRef]
- Mortarino, M.; Musella, V.; Costa, V.; Genchi, C.; Cringoli, G.; Rinaldi, L. GIS modeling for canine dirofilariosis risk assessment in central Italy. Geospat. Health. 2008, 2, 253–261. [Google Scholar] [CrossRef]
- Sassnau, R.; Czajka, C.; Kronefeld, M.; Werner, D.; Genchi, C.; Tannich, E.; Kampen, H. Dirofilaria repens and Dirofilaria immitis DNA findings in mosquitoes in Germany: Temperature data allow autochthonous extrinsic development. Parasitol. Res. 2014, 113, 3057–3061. [Google Scholar] [CrossRef]
- Kartashev, V.; Afonin, A.; González-Miguel, J.; Sepúlveda, R.; Simón, L.; Morchón, R.; Simón, F. Regional warming and emerging vector-borne zoonotic dirofilariosis in the Russian Federation, Ukraine, and other post-Soviet states from 1981 to 2011 and projection by 2030. BioMed Res. Intern. 2014, 2014, 858936. [Google Scholar] [CrossRef] [PubMed]
- Ciucă, L.; Musella, V.; Miron, L.D.; Maurelli, M.P.; Cringoli, G.; Bosco, A.; Rinaldi, L. Geographic distribution of canine heartworm (Dirofilaria immitis) infection in stray dogs of eastern Romania. Geospat. Health 2016, 11, 499. [Google Scholar] [CrossRef] [PubMed]
- Farkas, R.; Mag, V.; Gyurkovszky, M.; Takács, N.; Vörös, K.; Solymosi, N. The current situation of canine dirofilariosis in Hungary. Parasitol. Res. 2020, 119, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Simón, L.; Afonin, A.; López-Díez, L.I.; González-Miguel, J.; Morchón, R.; Carretón, E.; Montoya-Alonso, J.A.; Kartashev, V.; Simón, F. Geo-environmental model for the prediction of potential transmission risk of Dirofilaria in an area with dry climate and extensive irrigated crops. The case of Spain. Vet. Parasitol. 2014, 200, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Alonso, J.A.; Carretón, E.; Simón, L.; González-Miguel, J.; García-Guasch, L.; Morchón, R.; Simón, F. Prevalence of Dirofilaria immitis in dogs from Barcelona: Validation of a geospatial prediction model. Vet. Parasitol. 2015, 212, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Escolar, I.; Hernández-Lambraño, R.E.; Sánchez-Agudo, J.A.; Collado, M.; Pérez-Pérez, P.; Morchón, R. Current Risk of Dirofilariosis Transmission in the Iberian Peninsula (Spain and Portugal) and the Balearic Islands (Spain) and Its Future Projection under Climate Change Scenarios. Animals 2023, 13, 1764. [Google Scholar] [CrossRef]
- Gobierno de Canarias. Available online: https://www3.gobiernodecanarias.org/sanidad/scs/scs/1/plansalud/psc02/psc02_11.htm (accessed on 23 May 2023).
- Climate shifts. Worldmaps of Köppen-Geiger Climate Classification. Available online: http://koeppen-geiger.vuwien.ac.at/shifts.html (accessed on 12 April 2023).
- Melero-Alcíbar, R.; Molina, R.; Morchón, R.; Simón, F.; Latorre-Estivalis, J.; Bargues, M.D.; Mas-Coma, S.; Báez, M. Nota sobre la fauna de Culícidos de las Islas Canarias: Isla de Gran Canaria. Boletín de la SEA. 2006, 39, 427–428. [Google Scholar]
- Melero-Alcíbar, R.; Molina, R.; Morchón, R.; Simón, F.; Latorre-Estívalis, J.M.; Bargues, M.D.; Mas-Coma, S.; Pou-Barreto, C.; Valladares, B. Nota sobre la fauna de Culícidos de las Islas Canarias II: Isla de Tenerife*. Boletín de la SEA. 2008, 42, 463–464. [Google Scholar]
- Bravo-Barriga, D.; Gomes, B.; Almeida, A.P.; Serrano-Aguilera, F.J.; Pérez-Martín, J.E.; Calero-Bernal, R.; Reina, D.; Frontera, E.; Pinto, J. The mosquito fauna of the western region of Spain with emphasis on ecological factors and the characterization of Culex pipiens forms. J. Vector Ecol. 2017, 42, 136–147. [Google Scholar] [CrossRef]
- Spanish Government. Entomological Surveillance in Airports and Ports Against Imported Vectors of Exotic Infectious Diseases and Surveillance of Potential Autonomous Vectors of These Diseases. Available online: https://www.sanidad.gob.es/profesionales/saludPublica/ccayes/activPreparacionRespuesta/doc/Resumen_Proyecto_vigentomologica.2018.pdf (accessed on 26 May 2023).
- Serafín-Pérez, I.; Fernández-Serafín, C.; Rodríguez-Camacho, S.; Molina, R.; Lucientes, J.; del Castillo-Remiro, A.; Valladares, B. Implementación de la Vigilancia Entomológica en las islas de La Gomera y El Hierro (Islas Canarias). Proceedings of the XXII Congreso de la Sociedad Española de Parasitología, Madrid, Spain. Para J. 2022, 1, 204. [Google Scholar]
- Global Biodiversity Information Facility. Available online: http://www.gbif.org (accessed on 23 March 2023).
- World Clim. Available online: http://www.worldclim.org (accessed on 11 February 2023).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Intern. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Ripley, B.D. The R project in statistical computing.” MSOR Connections. The newsletter of the LTSN Maths, Stats OR Network 1.1. MSOR Connect. 2001, 1, 23–25. [Google Scholar] [CrossRef]
- Socioeconomic Data and Applications Center. 2023. Available online: https://sedac.ciesin.columbia.edu (accessed on 22 May 2023).
- Corine Land Cover. Copernicus Global Land Service. Available online: https://land.copernicus.eu/pan-european/corine-land-cover/clc2018 (accessed on 12 April 2023).
- EarthEnv. Available online: http://www.earthenv.org/landcover (accessed on 12 April 2023).
- Hernández-Lambraño, R.E.; González-Moreno, P.; Sánchez-Agudo, J.A. Towards the top: Niche expansion of Taraxacum officinale and Ulex europaeus in mountain regions of South America. Austral. Ecol. 2017, 42, 577–589. [Google Scholar] [CrossRef]
- Fath, B. Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Battini, N.; Farías, N.; Giachetti, C.B.; Schwindt, E.; Bortolus, A. Staying ahead of invaders: Using species distribution modeling to predict alien species’ potential niche shifts. Mar. Ecol. Prog. Ser. 2019, 612, 127–140. [Google Scholar] [CrossRef]
- Omar, K.; Thabet, H.S.; TagEldin, R.A.; Asadu, C.C.; Chukwuekezie, O.C.; Ochu, J.C.; Dogunro, F.; Nwangwu, U.; Onwude, O.; Ezihe, E.; et al. Ecological niche modeling for predicting the potential geographical distribution of Aedes species (Diptera: Culicidae): A case study of Enugu State, Nigeria. Parasite Epidemiol. Control. 2021, 15, e00225. [Google Scholar] [CrossRef]
- Fleitas, P.E.; Kehl, S.D.; Lopez, W.; Travacio, M.; Nieves, E.; Gil, J.F.; Cimino, R.O.; Krolewiecki, A.J. Mapping the global distribution of Strongyloides stercoralis and hookworms by ecological niche modeling. Parasites Vectors 2022, 15, 19. [Google Scholar] [CrossRef]
- Boorgula, G.D.Y.; Peterson, A.T.; Foley, D.H.; Ganta, R.R.; Raghavan, R.K. Assessing the current and future potential geographic distribution of the American dog tick, Dermacentor variabilis (Say) (Acari: Ixodidae) in North America. PLoS ONE 2020, 15, e0237191. [Google Scholar] [CrossRef]
- Cunze, S.; Kochmann, J.; Klimpel, S. Global occurrence data improve potential distribution models for Aedes japonicus japonicus in non-native regions. Pest. Manag. Sci. 2020, 76, 1814–1822. [Google Scholar] [CrossRef]
- Hanafi-Bojd, A.A.; Vatandoost, H.; Yaghoobi-Ershadi, M.R. Climate Change and the Risk of Malaria Transmission in Iran. J. Med. Entomol. 2020, 57, 50–64. [Google Scholar] [CrossRef]
- Rochat, E.; Vuilleumier, S.; Aeby, S.; Greub, G.; Joost, S. Nested Species Distribution Models of Chlamydiales in Ixodes ricinus (Tick) Hosts in Switzerland. App. Environ. Microbiol. 2020, 87, e01237-20. [Google Scholar] [CrossRef]
- Charrahy, Z.; Yaghoobi-Ershadi, M.R.; Shirzadi, M.R.; Akhavan, A.A.; Rassi, Y.; Hosseini, S.Z.; Webb, N.J.; Haque, U.; Bozorg Omid, F.; Hanafi-Bojd, A.A. Climate change and its effect on the vulnerability to zoonotic cutaneous leishmaniasis in Iran. Transbound. Emerg. Dis. 2022, 69, 1506–1520. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Edwards, T.C., Jr.; Guisan, A.; McMahon, S.M.; Normand, S.; Thuiller, W.; Wüest, R.O.; Zimmermann, N.E.; Elith, J. What do we gain from simplicity versus complexity in species distribution models? Ecography 2014, 37, 1267–1281. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. Kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Climatologies at High Resolution for the Earth’s Land Surface Areas. Available online: https://chelsa-climate.org/ (accessed on 23 March 2023).
- Karger, D.N.; Lange, S.; Hari, C.; Reyer, C.P.; Zimmermann, N.E. CHELSA-W5E5 v1. 0: W5E5 v1. 0 downscaled with CHELSA v2. 0. 2021; ISIMIP Repository. 2022. [Google Scholar]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD: Una plataforma para la predicción por conjuntos de la distribución de especies. Ecografía 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Morchón, R.; Moya, I.; González-Miguel, J.; Montoya, M.N.; Simón, F. Zoonotic Dirofilaria immitis infections in a province of Northern Spain. Epidemiol. Infect. 2010, 138, 380–383. [Google Scholar] [CrossRef]
- Montoya-Alonso, J.A.; Morchón, R.; Falcón-Cordón, Y.; Falcón-Cordón, S.; Simón, F.; Carretón, E. Prevalence of heartworm in dogs and cats of Madrid, Spain. Parasit Vectors 2017, 10, 354. [Google Scholar] [CrossRef]
- Pérez Pérez, P.; Rodríguez-Escolar, I.; Carretón, E.; Sánchez Agudo, J.A.; Lorenzo-Morales, J.; Montoya-Alonso, J.A.; Morchón, R. Serological Survey of Canine Vector-Borne Infections in North-Center Spain. Front. Vet. Sci. 2021, 8, 784331. [Google Scholar] [CrossRef] [PubMed]
- Montoya, J.A.; Morales, M.; Ferrer, O.; Molina, J.M.; Corbera, J.A. The prevalence of Dirofilaria immitis in Gran Canaria, Canary Islands, Spain (1994–1996). Vet. Parasitol. 1998, 75, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Valladares, B.; Gijon, H.; López-Roman, R. Dirofilaria immitis en la isla de Tenerife. Algunos datos de su fisiopatología. Rev. Iber. Parasitol. 1987, 47, 377–380. [Google Scholar]
- Guerrero, J.; Rojo, F.; Ródenas, A. Estudio de la incidencia del gusano del corazón en la población canina española. Med. Vet. 1989, 6, 217–220. [Google Scholar]
- Stenzenberger, R.; Gothe, R. Arthropoden Üebertragene parasitaere Infektionen und Zecken bei Hunden auf Teneriffa. Tierarztl. Prax. 1999, 1, 47–52. [Google Scholar]
- Montoya-Alonso, J.A.; Mellado, I.; Carretón, E.; Cabrera-Pedrero, E.D.; Morchón, R.; Simón, F. Canine dirofilariosis caused by Dirofilaria immitis is a risk factor for the human population on the island of Gran Canaria, Canary Islands, Spain. Parasitol. Res. 2010, 107, 1265–1269. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.T.; Cardoso, P.; Borges, P.A.; Gabriel, R.; Brito de Azevedo, E.; Francisco Reis, F.; Araújo, M.B.; Bento Elías, R. Effects of climate change on the distribution of indigenous species in oceanic islands (Azores). Clim. Change 2016, 138, 603–615. [Google Scholar] [CrossRef]
- Misslin, R.; Telle, O.; Daudé, E.; Vaguet, A.; Paul, R.E. Urban climate versus global climate change-what makes the difference for dengue? Ann. N. Y. Acad. Sci. 2016, 1382, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, J.; Hernández-Barrera, S.; Expósito, F.J.; Díaz, J.P.; González, A.; Pérez, J.C. The uneven impact of climate change on drought with elevation in the Canary Islands. NPJ Clim. Atmos. Sci. 2023, 6, 31. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Percent Contribution |
|---|---|
| Human footprint | 83.32% |
| Shrub density | 6.78% |
| Artificial water bodies | 4.02% |
| BIO4 (temperature seasonality) | 2.98% |
| BIO8 (mean temperature of wettest quarter) | 1.36% |
| BIO12 (annual precipitation) | 0.74% |
| Natural water bodies | 0.61% |
| Irrigated crops | 0.11% |
| BIO3 (isothermality) | 0.05% |
| Herbaceous density | 0.04% |
| BIO9 (mean temperature of driest quarter) | 0% |
| BIO15 (precipitation seasonality) | 0% |
| BIO1 (annual mean temperature) | 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morchón, R.; Rodríguez-Escolar, I.; Lambraño, R.E.H.; Agudo, J.Á.S.; Montoya-Alonso, J.A.; Serafín-Pérez, I.; Fernández-Serafín, C.; Carretón, E. Assessment Heartworm Disease in the Canary Islands (Spain): Risk of Transmission in a Hyperendemic Area by Ecological Niche Modeling and Its Future Projection. Animals 2023, 13, 3251. https://doi.org/10.3390/ani13203251
Morchón R, Rodríguez-Escolar I, Lambraño REH, Agudo JÁS, Montoya-Alonso JA, Serafín-Pérez I, Fernández-Serafín C, Carretón E. Assessment Heartworm Disease in the Canary Islands (Spain): Risk of Transmission in a Hyperendemic Area by Ecological Niche Modeling and Its Future Projection. Animals. 2023; 13(20):3251. https://doi.org/10.3390/ani13203251
Chicago/Turabian StyleMorchón, Rodrigo, Iván Rodríguez-Escolar, Ricardo E. Hernández Lambraño, José Ángel Sánchez Agudo, José Alberto Montoya-Alonso, Irene Serafín-Pérez, Carolina Fernández-Serafín, and Elena Carretón. 2023. "Assessment Heartworm Disease in the Canary Islands (Spain): Risk of Transmission in a Hyperendemic Area by Ecological Niche Modeling and Its Future Projection" Animals 13, no. 20: 3251. https://doi.org/10.3390/ani13203251