

Phenotype Alterations in the Cecal Ecosystem Involved in the Asymptomatic Intestinal Persistence of Paratyphoid Salmonella in Chickens

1
Southern Plains Agricultural Research Center, USDA-ARS, College Station, TX 77845, USA
2
Instituto de Patobiología, Instituto Nacional de Tecnología, Nicolas Repetto y Los Reseros S/N, Hurlingham 1686, Buenos Aires, Argentina
*
Author to whom correspondence should be addressed.
Animals 2023, 13(18), 2824; https://doi.org/10.3390/ani13182824
Submission received: 25 July 2023
/
Revised: 28 August 2023
/
Accepted: 4 September 2023
/
Published: 6 September 2023
(This article belongs to the Special Issue EBM Collection: Composition and Function of Poultry Gut Microbiota)
{kind=link}
Abstract
:Simple Summary
To combat infections, hosts employ a combination of antagonistic and cooperative host defense strategies. The former refers to pathogen killing mediated by host immunity (disease resistance), while the latter refers to physiological defense mechanisms that promote host health during infection independent of pathogen killing, leading to a cooperation between the host and the pathogen (diseases tolerance). In chickens, the paratyphoid Salmonella evolved the capacity to survive the initial robust immune response and persist in the avian ceca for months without triggering clinical signs. The persistent or carrier phase of a Salmonella infection in the avian host involves a complex balance of antagonistic and cooperative host defense strategies. Initially, the host reacts with a normal inflammatory response that controls bacterial invasion. After 3–4 days of inflammation, the host response changes to a more anti-inflammatory response characterized by changes in the local intestinal physiology that is no longer antagonistic to the bacterial pathogen, but instead ‘tolerates’ its presence. Thus, the chicken becomes a carrier of the pathogen allowing it to survive in the intestine without causing disease. It is hoped that understanding these mechanisms of pathogen survival in the chicken will allow future research to exploit these novel disease tolerance mechanisms to impact poultry health and reduced foodborne infections of Salmonella.
Abstract
The gastrointestinal ecosystem involves interactions between the host, gut microbiota, and external environment. To colonize the gut of poultry, Salmonella must surmount barriers levied by the intestine including mucosal innate immune responses and microbiota-mediated niche restrictions. Accordingly, comprehending Salmonella intestinal colonization in poultry requires an understanding of how the pathogen interacts with the intestinal ecosystem. In chickens, the paratyphoid Salmonella have evolved the capacity to survive the initial immune response and persist in the avian ceca for months without triggering clinical signs. The persistence of a Salmonella infection in the avian host involves both host defenses and tolerogenic defense strategies. The initial phase of the Salmonella–gut ecosystem interaction is characteristically an innate pro-inflammatory response that controls bacterial invasion. The second phase is initiated by an expansion of the T regulatory cell population in the cecum of Salmonella-infected chickens accompanied by well-defined shifts in the enteric neuro-immunometabolic pathways that changes the local phenotype from pro-inflammatory to an anti-inflammatory environment. Thus, paratyphoid Salmonella in chickens have evolved a unique survival strategy that minimizes the inflammatory response (disease resistance) during the initial infection and then induces an immunometabolic reprogramming in the cecum that alters the host defense to disease tolerance that provides an environment conducive to drive asymptomatic carriage of the bacterial pathogen.
1. Introduction
A feature of paratyphoid Salmonella serovars (broad-host range) in poultry is a persistent intestinal infection, or asymptomatic carrier state [1,2,3,4]. Further, these Salmonella asymptomatic carriers excrete high numbers of Salmonella into the environment [1,2,3,4], thereby increasing their propagation and facilitating contamination of other birds by horizontal transmission or affecting newly hatched chicks [5,6].
The establishment of persistence in the chicken gut by Salmonella requires overcoming both host-mediated elements such as a substantial immunometabolic mucosal response [7], but also microbiota-linked features such as production of antimicrobial fermentation products and the occupation of nutrient and adhesion niches [8,9]. Surmounting both features of the avian intestinal ecosystem by Salmonella is quite different than that observed in the human and swine gut [9,10,11]. The mechanisms that underlie pathogen persistence in the gut form the basis for this review.
2. The Gut Ecosystem
Optimal gut health is of vital importance to the performance of production animals, including poultry, to be able to perform to their genetic potential. Understanding a ‘healthy gut’ requires knowledge of the functional interactions of all components of the enteric ecosystem: the host, the microbiota, and the environment. The connections between these diverse physiological features of the enteric ecosystem underscore the extent of areas encompassed by the gut and the difficulty in correlating specific components of gut health with the ability to regulate poultry performance.
The gut can be considered as a complex and dynamic ecosystem molded during the interactions between the host, the gut microbiota, and environmental factors, including diet, temperature and humidity, animal density, infections, and mycotoxins. A diverse number of bacteria, commensal, potential beneficial, and pathogenic, must overcome several physical, chemical, and biological barriers imposed by this ecosystem, including host immune responses, the epithelial cell barrier, and microbiota-mediated events [12]. Consequently, bacterial intestinal colonization requires understanding the mechanisms by which the gut ecosystem interacts with microbes attempting to associate with the established community.
2.1. Components of the Intestinal Ecosystem
In general, there are three foundational components of the intestinal ecosystem: (a) host factors, (b) microbiota-linked factors, and (c) environmental factors, as recently detailed by Barron and Young [13].
2.1.1. Host Factors
The host provides the physical and biochemical foundations of the intestinal ecosystem with the intestinal epithelium and mucosal immune system forming the basis of the gut milieu.
- (A)
- Intestinal epithelium
The epithelium physical firewall is a single layer of epithelial cells that separate the densely colonized, and environmentally exposed, intestinal lumen from the largely sterile subepithelial tissue. The intestinal epithelial cell layer displays a number of distinctive functions including production of antimicrobial peptides and the secretion of mucus secrete antimicrobial peptides (defensins, cathelicidins, C-type lectins), which are a key defense against luminal microbes. Linking the epithelial cells are tight junctions, which help form a continuous luminal surface and help seal the intercellular space, near the apical surface, from the external environment [14]. Besides being the primary barrier preventing a microbial breach of the intestine, the epithelial cells should also be considered part of the cellular component of the innate immune response possessing pattern recognition receptors (PRRs) for sensing microbial-associated molecular patterns (MAMPs), but also capable of producing cytokines and chemokines to drive an inflammatory response against pathogen infection.
- (B)
- Immune system
Below the epithelial layer is the final component of the intestinal barrier: the immunological barrier where the professional immune cells (macrophages, DC, and lymphocytes) reside in the lamina propria [14]. This intestinal immune barrier has two distinct functions: the ability to respond to pathobionts (potential pathogenic microbes), invasive pathogens, and microbial products while also maintaining a state of tolerance to the diverse and beneficial commensal intestinal microbes [15]. Both systems working together through innate immune sensing using PRRs on epithelial cells and professional immune cells in the lamina propria (dendritic cells (DC) and macrophages) trigger immune pathways resulting in microbial killing and the activation of various acquired immune effector T cells (Th1, Th2, Th17, and Treg) all while keeping the resident microbiota in check without generating an overt inflammatory response. IgA-producing plasma cells, intraepithelial lymphocytes, and γδT cell receptor-expressing T cells are lymphocytes that are uniquely present in the mucosa. In addition, of the γδT cells in the intestinal lamina propria, there are significant numbers of IL-17-producing T cells and regulatory T cells (reviewed in [14]).
- (C)
- Enteric neuroendocrine system
The gut is the largest neuroendocrine organ in the body owing to the large numbers of neurons, gut hormones, and secondary messengers involved in regulating physiological functions in the host [16,17]. The neuroendocrine system (NES) of the gut involves two components including the gut endocrine cells, which are in the gut mucosa, and the enteric nervous system (ENS) in the gut submucosa. This system regulates several functions of the GI tract, such as motility, secretion, absorption, microcirculation in the gut, local immune defense, and cell proliferation [16,17]. The ENS comprises a large variety of neurotransmitters and associated receptors.
The gut contains a large number of enterochromaffin cells (endocrine cells that produce serotonin) dispersed among the epithelial cells of the gut mucosa in the intestine of the chicken [18,19]. The gut endocrine cells secrete signaling peptides into the lamina propria of the gut lining, where they have regulatory activity on the enteric nervous system (ENS), afferent and efferent nerve fibers of the central nervous system (CNS), and the autonomic nervous system (reviewed in [19]). Further, neurochemicals play a recognized role in determining bacterial colonization and interaction with the gut epithelium [20].
The gut–brain axis is a bidirectional information exchange network that connects the gut, the enteric endocrine system, and the CNS to the brain [21]. Villageliu and Lyte [22] corroborated the presence of the gut–brain axis in chickens, albeit not functional characterized. However, both Wickramasuriya and colleagues [23] and Cao et al. [24] have described the effects of environmental stresses on the gut–brain axis functional regulation of the physiology of the chicken.
2.1.2. Microbiota Factors
The chicken gastrointestinal (GIT) tract is home to a complex microbial community that links the environment to the health status of the host. The avian commensal microbiota are strategic managers of host physiology involved in regulating bird health [25,26], directing host intestinal metabolism and immunity, and directing a metabolome that affects energy balance and body weight [27]. Lastly, the residential microbes in the gut play a significant role in inhibiting pathogens from colonizing by a process called colonization resistance [8,9].
- (A)
- Microbial composition: nutrient competition
Effective nutrient acquisition in the competitive environment of the gut is essential for persistence of indigenous microbes. Nutrient resources for the microbiota are provided from dietary components or metabolites produced by the host either from the diet or mucus secretions [28,29]. Indigenous microbiota utilize dietary amino acids, carbohydrates, essential trace metals (iron, zinc, copper, manganese), and respiratory electron acceptors (O2 and NO3−), thereby starving the pathogens of essential nutrients and molecules [29,30,31].
- (B)
- Microbial composition: site competition
Commensal bacteria are able to control host membrane glycosylation and/or use it as a nutrient, thus creating a novel niche that reduces pathogen access to the epithelial barrier [32]. Further, commensal microbes occupy potential binding sites on the intestinal epithelium by deploying numerous molecular structures including outer membrane proteins, capsules, lectins, adhesins, and fimbriae [33]. Interestingly, Donaldson and colleagues [34] have shown that some symbiotic bacteria can co-opt the secretory IgA response to mediate stable colonization of the intestine which excludes colonization of indigenous pathobionts and pathogens access to the intestine.
- (C)
- Microbial composition and antimicrobial peptides
Commensal microbes can limit enteric bacterial colonization through direct microbe-microbe interactions that include contact-dependent killing competitor bacteria via the type 6 secretion system (T6SS), suppression of competitor bacteria growth by contact-dependent secretion of effector proteins which bind to specific receptors on the competitor bacteria that activate a toxic effector domain, or the production of bacteriocins which form pores in the competitor symbionts, pathobionts, or pathogens which induces leakage of cellular contents [32,35].
- (D)
- Microbial metabolite production
The microbiota, using a number of biochemical pathways, metabolize diet- and host-derived metabolites that can have a direct impact on the intestinal immune system and inhibit colonization of the intestine by competitor bacteria. For example, bacterial metabolites such as short chain fatty acids (SCFA) serve as an energy source to the epithelial cells but also have antimicrobial activity and limit virulence factor expression on pathogenic bacteria [36,37,38]. Further, microbiota can degrade dietary tryptophan to promote epithelial cell barrier function and breakdown dietary arginine which inhibits pro-inflammatory cytokine production [39].
3. Salmonella Interactions with the Intestinal Ecosystem in Chickens
One of the major causes of human gastroenteritis is Salmonella enterica (S. enterica) due to infected poultry products. Serovars, such as S. Typhimurium and S. Enteritidis, cause acute gastroenteritis in humans, but these bacteria colonize the intestines of chickens without causing disease [40]. This subclinical colonization poses a challenge for the prevention of foodborne transmission of Salmonella since colonized poultry are difficult to identify. The basis of the difference in the clinical consequences of S. Typhimurium and S. Enteritidis infection between mammals and birds is still vague, but recent studies have begun to unravel the fundamental differences which include the differential expression of virulence genes [41,42,43], host inflammation [42,44,45], and bacterial dietary and metabolic requirements [10,46,47].
3.1. Salmonella and Host Factors
3.1.1. Immune System
Paratyphoid Salmonella have evolved a unique survival strategy in poultry by surviving the initial robust immune response and persistently infecting the intestine. This chronic colonization of the intestinal tract is an important aspect of persistent Salmonella infection because it results in a silent propagation of bacteria in poultry stocks due to the impossibility to isolate contaminated animals. Data from our lab promote the hypothesis that Salmonella have evolved a unique survival strategy in poultry that minimizes host defenses (disease resistance) during the initial infection and then exploits and/or induces a dramatic immunometabolic reprogramming in the cecum that alters the host defense to disease tolerance.
- (A)
- Disease Resistance
Disease resistance is the host defense strategy grounded on the immune response’s capacity to detect and eliminate pathogens, i.e., host immunity [48,49,50]. The initial phase of the chicken cecum to Salmonella infection is manifested by the absence of clinical disease and functions to control pathogen invasion and reduce or eliminate the invading pathogen [48]. Specifically, Salmonella infection: (1) activated both Toll-like receptor (TLR) and Nod-like receptor (NLR) signaling pathways to initiate an innate immune response; (2) induced the production of chemokines CXCLi2 (IL-8) and cytokines IL-2, IL-6, IFN-α, and IFN-γ; (3) induced the phosphorylation of Janus kinase (JAK)/signal transducer and activator of transcription (STAT) signaling pathway that initiates innate immunity and coordinates adaptive immune mechanisms; (4) triggered both the intrinsic and extrinsic apoptotic pathways; and (5) activated the T cell receptor signaling pathway through the AP-1 and NF-κB transcription factor cascades [48].
- (B)
- Disease Tolerance
Disease tolerance is a second host defense strategy that limits the damage caused by a pathogen’s growth without affecting or reducing pathogen numbers [51,52]. In chickens, a second phase (starting at 4 days post-infection) of Salmonella persistence is characterized by an increase in the CD4+ CD25+ T cell (T regulatory [Tregs]) population in the cecum of Salmonella-infected chickens. Functionally, the cecal Tregs had increased suppressive activity for T effector cells and had a profound increase in IL-10 mRNA transcription [49,50].
Using chicken-specific kinomic immune-metabolism peptide arrays and quantitative real-time PCR of Salmonella-infected cecal tissue 4 to 17 days post challenge, distinct immune and metabolic pathways are altered and changed the local immunometabolic environment. In general, two energy sensory kinases, AMPK and mTOR, are key players linking specific extracellular milieu and intracellular metabolism. Phenotypically, the early response (4 to 72 h) to Salmonella infection is pro-inflammatory, fueled by glycolysis and mTOR-mediated protein synthesis, whereas after 4 days post-infection, the local environment has undergone an immune-metabolic reprogramming to an anti-inflammatory state driven by AMPK-directed oxidative phosphorylation [51,52].
3.1.2. Paratyphoid Salmonella Manipulation of the Enteric Nervous System
- (A)
- Salmonella and neurochemical release
The Mellata lab at Iowa State University conducted a series of experiments to understand interactions between the nervous and immune systems during paratyphoid Salmonella infections in chickens [53]. Using the drug reserpine, which induces the release of intracellular storage of catecholamines like norepinephrine (NE) to treat cecal explants and isolated Tregs from chickens, NE was increased [53]. Further, reserpine treatment in vivo reduced the level of intestinal Salmonella Typhimurium and other Enterobacteriaceae. These results provided the first direct data that Salmonella colonization inhibits the release of neurochemicals that participate in the regulation of the enteric neuro-immunological responses to infection [53].
- (B)
- Salmonella and neurotrophin signaling
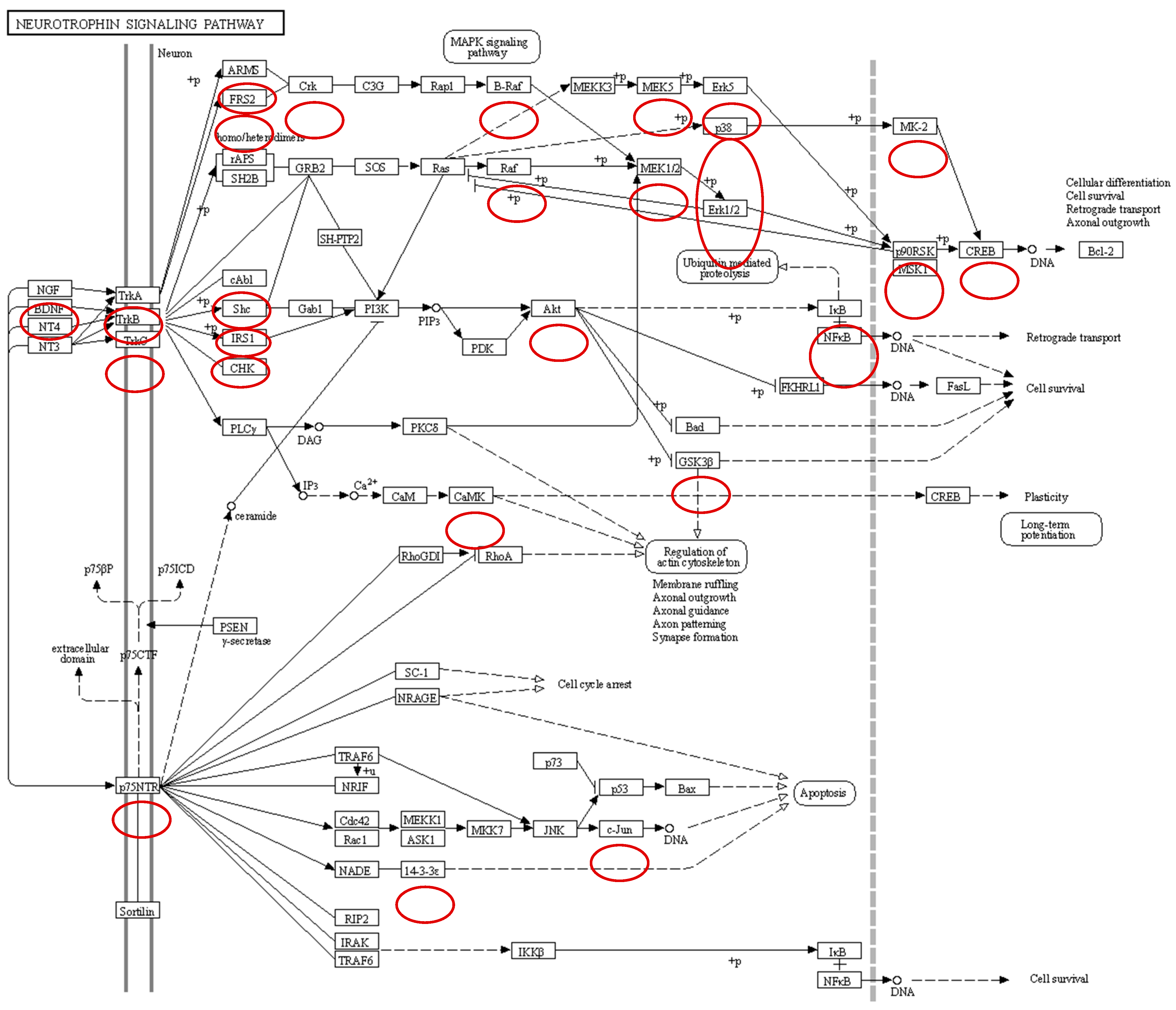
Neurotrophins are a family of growth factors critical to the functioning of the nervous system including neuron formation and survival. Neurotrophies are ligands of Trk receptors. Trk receptors are a family of tyrosine kinases that regulate gut sensation, motility, and epithelial barrier function, and protect enteric neurons and glial cells from damaging insults in the microenvironment of the gut via several signaling cascades including the extracellular signal-regulated kinases (ERK) pathway, the phosphoinositide 3-kinase (PI3K)/Akt pathway, and the phospholipase C (PLC)-γ pathway. Salmonella infection of the chicken cecum dephosphorylates the Trk A and C receptors resulting in the dephosphorylation of the ERK pathway, PI3K/Akt pathway, and the PLC-γ pathways (Figure 1). These results provide evidence that Salmonella infection manipulates enteric neuron functionality during colonization of the cecum, thus blocking the gut–brain axis that controls the enteric host response to the pathogen.
- (C)
- Infection-induced feeding behavior
A common feature of enteric infections is a reduction in feeding behavior or anorexia. However, paratyphoid Salmonella colonize the intestines of chickens without causing disease [40] including a lack of effect on growth feed intake, growth, and production [54]. As a function of disease tolerance host response, it has been shown that Salmonella enterica manipulates the gut–brain axis to inhibit anorexia, which reduces its virulence but promotes its transmission [52,54,55].
3.1.3. Salmonella and Microbiota-Linked Factors
- (A)
- Gastrointestinal microbiota
The crop, proventriculus and gizzard, duodenum, jejunum, and ileum (small intestine), ceca, large intestine, and cloaca is the basic structure of the poultry gastrointestinal tract [31] and each part plays a different role that is influenced by the dynamics of the microbiota. The gut chicken microbiota at various stages of the animal’s life supports functions that range from protection against pathogens and nutrient production to maturation of the immune system. Salmonella face an important competition for space and nutrients with this large and diverse community of gastrointestinal microorganisms.
Ileal and cecal microbiomes have been characterized in commercial chickens as a means of evaluating the influence of the microbiota on performance, health, and disease [25,27,56,57]. Microbiota composition is affected by numerous factors such as sex [58], genetics [59], diet [60], environment [25], and host stress [61,62]; yet, age is consistently the primary driving factor with the microbiota undergoing successional changes during the life cycle of a bird [63,64]. The richness and the composition of microbiota strongly influence the gut ecosystem functioning. As the individual grows, the gut microbiota undergo dynamic changes with greater diversity and complexity as the host ages, and variably by intestinal tract segments [56]. To describe and investigate the dynamics of the microbiota or to detect changes in composition, usually mathematical tools such as α-diversity indices (diversity within a sample) and β-diversity indices (diversity among samples) as well as the description of taxonomic composition are used. Salmonella colonization impact both diversity and composition of chicken gut microbiota [65,66]. However, it must be considered that the colonization and the interaction of Salmonella with the microbiota are complex processes that depend not only on the characteristics of the pathogen but also on other variables such as the time of infection, host characteristics, nutrition, and the gut microbiota composition itself.
- (B)
- Microbiota and Salmonella intestinal colonization
Considering microbiota composition and timing, the intestine of newborn chicks is a relatively sterile environment and an excellent opportunity for certain pathogens (such as Salmonella) to rapidly colonize and spread freely in the intestine [67]. Proteobacteria, primarily Enterobacteriaceae, including members such as Escherichia, Shigella, Enterococcus, and Salmonella, are the most common genera detected in day-of-hatch birds, but also high abundance of Enterobacteriaceae has been observed in and at 3-day-old layer chickens [65,68]. Proteobacteria abundance decreased significantly with age as Firmicutes members, particularly Clostridia, and overall diversity increase [69]. Firmicutes increase in abundance and taxonomic diversity starting around day 7 [63]. The reduction of certain bacteria taxa from Enterobacteriaceae family during the early post-hatch period might enhance the host resistance to enteric pathogen infection as well as transient perturbation of the gut microbiota produced by different stressors including antibiotics [70]. Although Salmonella is a common gut colonizer in poultry [71], it had become evident that the taxonomic features of the microbiota are an important factor determining susceptibility and resistance to Salmonella colonization at the individual level [72].
Salmonella enters chickens through vertical transmission from infected hens or via the oral route through infected feed, water, or litter, and colonize the distal part of the ileum and cecum [73]. Oral inoculation of Salmonella after a couple of days after hatch resulted in an important shift in chicken cecal microbiota composition at 7 and 14 dpi [74]. Experimentally, Salmonella is able to induce an asymptomatic carrier state when 5 × 104 colony-forming units (CFU) are orally inoculated in chicks [75], but the same bacteria load in older chicks has no effect and it is required up to 1 × 108 CFU to reproduce a successful infection in 30-week-old hens [76], supporting the observation that hens are usually more resistance to Salmonella infection than chicks. These observations suggest that Salmonella colonization in the gastrointestinal tract of the chickens has a direct effect on altering the natural development of the gut microbiota.
The role of the microbiota is also important to explain the heterogeneity of infection associated with the presence of super-shedders which constantly disseminated Salmonella to the low-shedder chicks [77]. The shedding levels are highly influenced by gut microbiota composition at the moment of infection, with α-diversity indices correlating with the shedding level as low-shedder chicks showed the lowest α-diversities [72]. The homogeneity of microbial compositions within the shedding level was corroborated by the analysis of β-diversity indices. Differences in the composition of the intestinal microbiota show an important influence on the susceptibility or resistance to colonization by Salmonella as adult hen microbiota samples orally delivered may have a protective effect on one-day-old chicks but the absence of gut microbiota results in super-shedders animals after Salmonella colonization [72,78]. The outcome of the first exposure to Salmonella seems to be determinant for shedding as, contrary to super-shedder chicks, low shedders can block early colonization [77].
3.1.4. Salmonella Enterica Serovar Enteritidis (SE) and Microbiota
In the immediate post-hatch period in chickens, SE infection is disadvantageous to the expansion of the gut microbiota resulting in the reduction in microbial diversity and an increase in potential pathogens in the microbial community [65,79]. In contrast, SE infection in older animals did not show dominance of a specific species in the community when compared to non-infected animals as determined by Shannon index (α-diversity) [79]. However, 3 weeks after infection of SE in these older hens, an increased colonization by minority members of the community following the infection as observed in the changes of Chao1 index (α-diversity) was observed [79]. The impact of SE in microbial communities in challenged one-day-old layer chicks seems to be more substantial in later stages of the infection [65]. However, besides that SE can be considered a good gut competitor and bacterial colonization was exclusively localized in the cecum of the infected chicks, systemic infection was not observed in an oral challenge model, also when high doses of inoculum were administered [79].
SE infection in young chicks significantly reduces the overall diversity of the microbiota population due to the expansion of the Enterobacteriaceae family; however, infection had a more significant impact on microbial communities during the later stages of infection where a negative correlation between Enterobacteriaceae and Lachnospiraceae [70], Ruminococcaceae, Erysipelotrichaceae, and Peptostreptococcaceae was observed in SE challenged chicks [65]. Although SE infection in newly hatched chickens did not influence the predominant cecal microbiota, an increase of Lactobacillaceae was observed [70]. The Ruminococcaceae family, which is more abundant in non-infected animals, has been suggested as a signature of Salmonella infection [5,80]. Considering only the genera presenting a high relative abundance (>5%), Enterococcus genus was considered the main taxonomic feature allowing to predict the low- or super-shedder phenotypes, although SE colonization was not modified by oral inoculation of chicken strains of E. faecium [72]. Although Enterococcus spp. might not prevent Salmonella colonization by itself, their proportional increase can be associated with another change at microbial or host physiologic level and could be used as biomarker of Salmonella infection. However, although not opposed to the previous observation, the association of Enterococcus with well-defined commensal bacteria (E. coli, Clostridium, Lactobacillus) orally inoculated the day of hatch reduced SE excretion and increased the proportion of low-shedders animals, which was not observed when inoculated separately [72].
3.1.5. Salmonella Enterica Serovar Typhimurium (ST) and Microbiota
ST colonization of the chicken intestinal tract, either after experimental challenge or natural infection, alter microbiota composition with resultant decreases in cecal Enterococcus, Lactobacillus, Escherichia, and Bacillaceae [81]. Further, ST infection significantly reduced α-diversity indexes of ileal microbiota of broiler chickens [54] with a notable increase in Escherichia-Shigella genus levels [82].
A few days after ST colonization (3 days post-infection), an increased α-diversity was observed in cecal microbiota, but the change was rapidly inverted at 5 and 7 dpi, suggesting a differential effect depending on the dynamics of the innate immune response [83]. Additionally, the challenge decreased the cecal abundance of Lactobacillus, Bifidobacterium, Trabulsiella, Oscillospira, Holdemania, and Coprococcus, and increased Klebsiella and Escherichia [66,83]. In the feces of ST-challenged laying chickens, a significantly lower microbial α-diversity was observed when compared to control during several weeks after the challenge with a decrease in the abundance of Blautia, Enorma, Faecalibacterium, Shuttleworthia, Sellimonas, Intestinimonas, and Subdoligranulum and increase in the abundance of Butyricicoccus, Erysipelatoclostridium, Oscillibacter, and Flavonifractor [83].
- (C)
- Intestinal microbial functions and host metabolite production
Alteration in the overall microbial community following Salmonella infection in chickens has a consequent effect on the host regulation of cecum-associated metabolic networks [65,84,85,86]. Metagenome functional prediction of the chicken gut microbiome shown various altered pathways in 2-week-old SE-infected animals, including functional genes associated with ribosomal activity and nucleotide metabolism (purine and pyrimidine) which could suggest an interference of Salmonella with the metabolism of intestinal microorganisms and intestinal activity [79]. Metabolic pathway analysis of the cecal content from birds infected with SE revealed a disruption in microbiota metabolic pathways related to arginine and proline metabolism as well as reduced tricarboxylic acid cycle (TCA) activity. Similarly, ST infection of chick early post-hatch also found revealed differences with non-infected animals in metabolic composition of ceca content including lactate, the main product of glucose fermentation of Enterococcus [87], supporting the observation that enterococci are significant members of the cecal microbiota during Salmonella infection. Furthermore, the microbiota composition was unchanged in neonatal chicks infected with ST, but the functional activity of the microbiota was dramatically altered [86]. For example, ST infection induced the increase expression of genes involved in branched-chain amino acid (BCAA) production, such as leucine, isoleucine, and valine [86] which play a key role in the growth, production performance, immunity, and intestinal health of chickens [87,88].
Global gene expression pattern and metabolites profile in the host is altered during Salmonella infection. A significant accumulation of metabolites was identified in the ileum and cecum of Salmonella-infected birds [79,86]. Immediately after infection, a comparatively moderate number [6] of metabolites were affected in cecum of infected chickens. However, a week after infection, a considerable number of metabolites [78] are altered but the difference was dramatically reduced (three metabolites) a week later [79]. Up-regulation of arginine and proline metabolism was detected in association with Salmonella infection, suggesting the activation of a host metabolic adjustment strategy to reduce the intestinal inflammation during Salmonella infection to improve their intestinal colonization [79]. Arginine is a common amino acid substrate used by the inducible nitric oxide synthase (iNOS) for nitric oxide production, one of the key innate immune responses to induce inflammation as part of the host defense mechanism [89]. The analysis of the metabolites revealed reduced tricarboxylic acid cycle (TCA) in the Salmonella-infected group compared to the non-infected group, suggesting that a change in host cellular energy metabolism during Salmonella infection occurred [79]. The alteration in host metabolic response could be associated with the innate immune activation triggered by LPS stimuli from Salmonella infection [90].
4. Concluding Remarks
Because of the increase in antimicrobial-resistant microbes, the use of antibiotics as growth promoters has either been banned by government intervention or removed by producers due to the consumer demand for ‘no antibiotics ever’ or ‘raised without antibiotics’ poultry products. Thus, there is an ongoing demand for the development and use of alternatives to antibiotics for growth promotion and food safety.
Herein, we have provided a review of the literature on Salmonella interactions with the different components of the avian GIT ecosystem. Further, we have shared insights into how these interactions appear to be involved in the establishment of a persistent Salmonella infection in the cecal lumen. The dynamics of these host–pathogen interactions involve host neuroimmune and immunometabolic pathways, pathogen virulence, gut microbiota, and the crosstalk between all components which has been described by Troha and Ayres [91] as ‘a household of three’. In this context, Troha and Ayres [91] described that the metabolic needs of all three members of the household must be studied to develop novel strategies that target metabolic processes that underpin the intestinal ecosystem.
Traditionally, studies into the host response of poultry to infections with paratyphoid Salmonella serovars have concentrated on host resistance mechanisms which target the elimination of the pathogen by the immune system [46,48,51,52]. However, in poultry, paratyphoid Salmonella have evolved the capacity to survive this initial immune response and persist in the gut lumen for weeks without causing clinical disease in birds [40,48]. This persistent colonization of the intestinal tract is an important aspect of a Salmonella infection because it results in the silent propagation of bacteria in poultry stocks due to the impossibility to isolate contaminated animals. This persistence suggests that a second defense mechanism has evolved in chicken-Salmonella infection biology that functions to foster host health instead of removal of the pathogen [51,52]. This alternate defense strategy, known as disease tolerance, involves protecting the overall physiological homeostasis in the bird [51,52,53]. Collectively, the information from the literature implies that these neuro-immunometabolic connections between the host and its microbiota could be manipulated and that targeting the regulators of these neuro-immunometabolic pathways signify a promising translational approach to novel therapeutics in the future.
The use of omics technologies has opened the doors for better understanding of the Salmonella–intestinal ecosystem interactome, but more holistic approaches are required. All-inclusive studies using systems biology approaches are needed. For example, we have cited a number of studies describing the neuroimmune, immunometabolic, and microbiological alterations induced in the cecum of the host by Salmonella infections but provided no definitive information on the role that the microbiota play in this environment. Likewise, dozens of studies have provided ‘lists’ of microbiota compositional changes that have been found during Salmonella infections, but few, if any, of these studies showed a causal relationship to the host functional responsiveness to the infection. Our hope is that providing this overview in the literature on the Salmonella–intestinal ecosystem interactome will encourage more collaborative studies between laboratories concentrating on these interactions between the avian gut, the gut microbiota, and Salmonella.
Author Contributions
Conceptualization, M.H.K. and M.E.F.M.; writing—original draft preparation, M.H.K. and M.E.F.M.; writing—review and editing, M.H.K. and M.E.F.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barrow, P.A.; Simpson, J.M.; Lovell, M.A. Intestinal colonization in the chicken by food-poisoning Salmonella serotypes; microbial characteristics associated with fecal excretion. Avian Pathol. 1988, 17, 571–588. [Google Scholar] [CrossRef] [PubMed]
- Withanage, G.S.K.; Wigley, P.L.; Kaiser, P.; Mastroeni, P.; Brooks, H.; Powers, C.; Beal, R.; Barrow, P.; Maskell, D.; McConnell, I. Cytokine and chemokine responses associated with clearance of a primary Salmonella enterica Typhimurium infection in the chicken and in protective immunity to rechallenge. Infect. Immun. 2005, 73, 5173–5182. [Google Scholar] [CrossRef] [PubMed]
- Perron, G.G.; Quessy, S.; Bell, G. A reservoir of drug-resistant pathogenic bacteria in asymptomatic hosts. PLoS ONE 2008, 3, e3749. [Google Scholar] [CrossRef]
- Aksakal, A.; Boynukara, B.; Somaz, H.; Kutlu, I.; Gulhan, T. Occurrence and antibiotic susceptibility of Salmonella serotypes in apparently healthy slaughtered sheep in Van, Turkey. J. Anim. Vet. Adv. 2009, 8, 1455–1460. [Google Scholar]
- Cazals, A.; Estellé, J.; Bruneau, N.; Coville, J.L.; Menanteau, P.; Rossignol, M.N.; Jardet, D.; Bevilacqua, C.; Rau, A.; Bed’Hom, B.; et al. Differences in caecal microbiota composition and Salmonella carriage between experimentally infected inbred lines of chickens. Genet. Sel. Evol. 2022, 54, 7. [Google Scholar] [CrossRef]
- Cazals, A.; Rau, A.; Estelle, J.; Bruneau, N.; Coville, J.-L.; Menanteau, P.; Rossignol, M.-N.; Jardet, D.; Beveliacqua, C.; Bed’Hom, B.; et al. Comparative analysis of the cecal tonsil transcriptome in two chicken lines experimental infected with Salmonella Enteritidis. PLoS ONE 2022, 17, e0270012. [Google Scholar] [CrossRef] [PubMed]
- Wigley, P. Salmonella enterica in the chicken: How it has helped our understanding of immunology in a non-biomedical model species. Front. Immunol. 2014, 5, 482. [Google Scholar] [CrossRef]
- Lawley, T.D.; Walker, A.W. Intestinal colonization resistance. Immunology 2013, 138, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shealy, N.G.; Yoo, W.; Byndloss, M.X. Colonization resistance: Metabolic warfare as a strategy against pathogenic Enterobacteriaceae. Curr. Opin. Microbiol. 2021, 64, 82–90. [Google Scholar] [CrossRef]
- Taylor, S.J.; Winter, S.E. Salmonella finds a way: Metabolic versatility of Salmonella enterica serovar Typhimurium in diverse host environments. PLoS Pathog. 2020, 16, e100854. [Google Scholar] [CrossRef]
- Rogers, A.W.L.; Tsolis, R.M.; Baumler, A.J. Salmonella versus the microbiome. Microbiol. Mol. Biol. Rev. 2021, 85, e00027-19. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Barron, M.R.; Young, V.B. Viewing bacterial colonization through the lens of systems biology. mSystems 2022, 7, e01383-21. [Google Scholar] [CrossRef]
- Smith, A.L.; Powers, C.; Beal, R. Chapter 11.1. The avian enteric immune system in health and disease. In Avian Immunology, 3rd ed.; Kaspers, V., Schat, K., Gobel, R., Vervelde, L., Eds.; Academic Press: London, UK, 2021; pp. 303–326. [Google Scholar]
- Broom, L.J.; Kogut, M.H. The role of the gut microbiota in shaping the immune system of chickens. Vet. Immunol. Immunopathol. 2018, 204, 44–51. [Google Scholar] [CrossRef]
- Neuman, H.; Debelius, J.W.; Knight, R.; Koren, O. Microbial endocrinology: The interplay between the microbiota and the endocrine system. FEMS Microbiol. Rev. 2015, 39, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Knauf, C. How gut microbes talk to organs: The role of endocrine and nervous routes. Mol. Metab. 2016, 5, 743–752. [Google Scholar] [CrossRef]
- Rawdon, B.B. Gastrointestinal hormones in birds: Morphological, chemical, and developmental aspects. J. Exp. Zool. 1984, 232, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, K. Chicken intestinal L cells and glucagon-like peptide-1 secretion. J. Poult. Sci. 2020, 57, 1–6. [Google Scholar] [CrossRef]
- Lyte, J.M.; Martinez, D.A.; Robinson, K.; Donoghue, A.M.; Daniels, K.M.; Lyte, M. A neurochemical biogeography of the broiler chicken intestinal tract. Poult. Sci. 2021, 101, 101671. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Burford, W.B.; Pham, G.; Zhang, L.; Alto, L.T.; Ertelt, J.M.; Winter, M.G.; Winter, S.E.; Way, S.S.; Alto, N.M. Systemic reconstruction of an effector-gene network reveals determinants of Salmonella cellular and tissue tropism. Cell Host Microbe 2021, 29, 1531–1547. [Google Scholar] [CrossRef] [PubMed]
- Villageliu, D.N.; Lyte, M. Microbial endocrinology: Why the intersection of microbiology and neurobiology matters to poultry health. Poult. Sci. 2017, 96, 2501–2508. [Google Scholar] [CrossRef] [PubMed]
- Wickramasuriya, S.S.; Park, I.; Lee, K.; Lee, Y.; Kim, W.H.; Nam, H.; Lillehoj, H.S. Role of physiology, immunity, microbiota, and infectious diseases in the gut health of poultry. Vaccines 2022, 10, 172. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Chowdhury, V.S.; Cline, M.A.; Gilbert, E.R. The microbiota-gut-brain axis during heat stress in chickens: A review. Front. Physiol. 2022, 12, 752265. [Google Scholar] [CrossRef]
- Oakley, B.B.; Buhr, R.J.; Ritz, C.W.; Kiepper, B.H.; Berrang, M.E.; Seal, B.S.; Cox, N.A. Successional changes in the chicken cecal microbiome during 42 days of growth are independent of organic acid feed additives. BMC Vet. Res. 2014, 10, 282. [Google Scholar] [CrossRef]
- Sargeant, M.J.; Constantinidou, C.; Cogan, T.A.; Bedford, M.R.; Penn, C.W.; Pallen, M.J. Extensive microbial and functional diversity within the chicken cecal microbiome. PLoS ONE 2014, 9, e91941. [Google Scholar] [CrossRef]
- Carrasco, J.M.D.; Casanova, N.A.; Fernandez-Miyakawa, M.E.F. Microbiota, gut health, and chicken productivity: What is the connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef] [PubMed]
- Feye, K.M.; Baxter, M.F.A.; Tellez-Isaias, G.; Kogut, M.H.; Ricke, S.C. Influential factors on the composition of the conventionally raised broiler gastrointestinal microbiomes. Poult. Sci. 2020, 99, 653–659. [Google Scholar] [CrossRef]
- Aruwa, C.E.; Pillay, C.; Nyaga, M.M.; Sabiu, S. Poultry gut health—Microbiome functions, environmental impacts, microbiome engineering and advancements in characterization technologies. J. Anim. Sci. Biotechnol. 2021, 12, 119. [Google Scholar] [CrossRef] [PubMed]
- Apajalaht, J.; Vienola, K. Interaction between chicken intestinal microbiota and protein digestion. Ann. Feed Sci. Technol. 2016, 221, 323–330. [Google Scholar] [CrossRef]
- Rychlik, I. Composition, and function of chicken gut microbiota. Animals 2020, 10, 103. [Google Scholar] [CrossRef]
- Caballero-Flores, G.; Pickard, J.M.; Nunez, G. Microbiota-mediated colonization resistance: Mechanisms and regulation. Nat. Rev. Microbiol. 2023, 21, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, G.P.; Ladinsky, M.S.; Yu, K.B.; Sanders, J.G.; Yoo, B.B.; Chou, W.-C.; Conner, M.E.; Earl, A.M.; Knight, R.; Bjorkman, P.J.; et al. Gut microbiota utilize immunoglobulin A for mucosal colonization. Science 2017, 360, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Pickard, J.M.; Nunez, G. Pathogen colonization resistance in the gut and its manipulation for improved health. Am. J. Pathol. 2019, 189, 1300–1310. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.Y.; Packialakshmi, B.; Makkar, S.K.; Dridi, S.; Rath, N.C. Effect of butyrate on immune response of a chicken macrophage cell line. Vet. Immunol. Immunopathol. 2014, 162, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Ji, J.; Qu, H.; Wang, J.; Shu, D.M.; Liu, T.F.; Li, Y.; Luo, C.L. Effects of sodium butyrate on intestinal health and gut microbiota composition during intestinal inflammation progression in broilers. Poult. Sci. 2019, 98, 4449–4456. [Google Scholar] [CrossRef]
- Gupta, A.; Bansai, M.; Wagle, B.; Su, X.; Rath, N.; Donoghue, A.; Upadhyay, A. Sodium butyrate reduces Salmonella Enteritidis infection of chicken enterocytes and expression of inflammatory host genes in vitro. Front. Microbiol. 2020, 11, 553670. [Google Scholar] [CrossRef] [PubMed]
- Fouad, A.M.; El-Senousey, H.K.; Ruan, D.; Wang, S.; Xia, W.; Zheng, C. Tryptophan in poultry nutrition: Impacts and mechanisms of action. J. Anim. Physiol. Anim. Nutr. 2021, 105, 1146–1153. [Google Scholar] [CrossRef]
- Van Immerseel, F.; De Buck, J.; Pasmans, F.; Bohez, L.; Boyen, F.; Haesebrouck, F.; Ducatelle, R. Intermittent long-term shedding and induction of carrier birds after infection of chickens early post-hatch with a low or high dose of Salmonella Enteritidis. Poult. Sci. 2004, 83, 1911–1916. [Google Scholar] [CrossRef]
- Addwebi, T.T.; Call, D.R.; Shah, D.H. Contributions of Salmonella Enteritidis virulence factors to intestinal colonization and systemic dissemination in 1-day-old chicks. Poult. Sci. 2014, 93, 871–881. [Google Scholar] [CrossRef]
- Eade, C.R.; Bogomolnaya, L.; Hung, C.C.; Betteken, M.I.; Adams, L.G.; Rews-Polymenis, H.; Altier, C. Salmonella pathogenicity island 1 is expressed in the chicken intestine and promotes bacterial proliferation. Infect. Immun. 2018, 87, e00503-18. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, J.; Chen, Y. Regulation of neurotransmitters by the gut microbiotaand effects on cognition in neurological disorders. Nutrients 2012, 13, 2099. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.J.; Kendall, M.M. Salmonella enterica Serovar Typhimurium Strategies for Host Adaptation. Front. Microbiol. 2017, 8, 1983. [Google Scholar] [CrossRef] [PubMed]
- Galan, J.E. Salmonella Typhimurium and inflammation: A pathogen-centric affair. Nat. Rev. Microbiol. 2021, 19, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Barrow, P.A.; Berchieri, A.; Neto, O.C.F.; Lovett, M. The contribution of aerobic and anaerobic respiration to intestinal colonization and virulence for Salmonella typhimurium in the chicken. Avian Pathol. 2015, 44, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-J.; Bogomolnaya, L.; McClelland, M.; Andrews-Polymenis, H. De novo pyrimidine synthesis is necessary for intestinal colonization of Salmonella Typhimurium in chicks. PLoS ONE 2017, 12, e0183751. [Google Scholar] [CrossRef] [PubMed]
- Kogut, M.H.; Genovese, K.J.; Byrd, J.A.; Swaggerty, C.; He, H.; Farnell, Y.; Arsenault, R.J. Chicken-specific kinome analysis of early host immune signaling pathways in the cecum of newly hatched chickens infected with Salmonella enterica serovar Enteritidis. Front. Cell. Infect. Microbiol. 2022, 12, 899395. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, R.; Acevedo, K.; Mortada, M.; Akerele, G.; Applegate, T.; Kogut, M.; Selvaraj, R. Effects of Salmonella enterica ser. Enteritidis and Heidelberg on host CD4+CD25+ regulatory T cell suppressive immune responses in chickens. PLoS ONE 2021, 16, e0260280. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, R.; Kogut, M.H.; Arsenault, R.J.; Swaggerty, C.L.; Cole, K.Y.; Reddish, M.J.; Selvaraj, R.K. Effect of Salmonella infection on cecal tonsil regulatory T cell properties in chickens. Poult. Sci. 2015, 94, 1828–1835. [Google Scholar] [CrossRef]
- Kogut, M.H.; Genovese, K.J.; He, H.; Arsenault, R.J. AMPK and mTOR: Sensors and regulators of immunometabolic changes during Salmonella infection in the chicken. Poult. Sci. 2016, 95, 345–353. [Google Scholar] [CrossRef]
- Kogut, M.; Arsenault, R. Immunometabolic phenotype alterations associated with the induction of disease tolerance and persistent asymptomatic infection of Salmonella in the chicken intestine. Front. Immunol. 2017, 8, 372. [Google Scholar] [CrossRef] [PubMed]
- Redweik, G.A.J.; Kogut, M.H.; Arsenault, M.J.; Lyte, M.; Mellata, M. Reserpine induces antimicrobial responses in chicken intestine via neuro-immunometabolic signaling and MEK1/2 activation. Commun. Biol. 2021, 4, 1358. [Google Scholar] [CrossRef]
- He, Y.; Yang, Y.; Dong, Y.; Ito, K.; Zhang, B. Highly nutritious diet resists Salmonella Typhimurium infections by improving intestinal microbiota and morphology in broiler chickens. Poult Sci. 2020, 99, 7055–7065. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, R.J.; Napper, S.; Kogut, M.H. Salmonella enterica Typhimurium infection causes metabolic changes in chicken muscle involving AMPK, fatty acid and insulin/mTOR signaling. Vet. Res. 2013, 4, 35. [Google Scholar] [CrossRef]
- Stanley, D.; Hughes, R.; Moore, R.J. Microbiota of the chicken gastrointestinal tract: Influence on health, productivity, and disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef]
- Clavijo, V.; Florez, M.J.V. The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: A review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef]
- Lee, K.-C.; Kil, D.Y.; Sul, W.J. Cecal microbiome divergence of broiler chickens by sex, and body weight. J. Microbiol. 2017, 55, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, G.; Siegel, P.; He, C.; Wang, H.; Zhao, W.; Zhai, Z.; Tian, F.; Zhao, J.; Zhang, H.; et al. Quantitative genetic background of the host influences gut microbiomes in chickens. Sci. Rep. 2013, 3, 1163. [Google Scholar] [CrossRef]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef]
- Zhu, L.; Liao, R.; Wu, N.; Zhu, G.; Yang, C. Heat stress mediates changes in fecal microbiome and functional pathways of laying hens. Appl. Microbiol. Biotechnol. 2019, 103, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Bai, L.; Qu, Q.; Zhou, S.; Yang, M.; Guo, S.; Li, Q.; Liu, C. Impact of gut microbiota structure in heat-stressed broilers. Poult. Sci. 2019, 98, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Ballou, A.L.; Ali, R.A.; Mendoza, M.A.; Ellis, J.C.; Hassan, H.M.; Croom, W.J.; Koci, M.D. Development of the chick microbiome: How early exposure influences future microbial diversity. Front. Vet. Sci. 2016, 3, 163181. [Google Scholar] [CrossRef]
- Oakley, B.B.; Kogut, M.H. Spatial and temporal changes in the broiler chicken cecal and fecal microbiomes and correlations of bacterial taxa with cytokine gene expression. Front. Vet. Sci. 2016, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Mon, K.K.Z.; Saelao, P.; Halstead, M.M.; Chanthavixay, G.; Chang, H.C.; Garas, L.; Maga, E.A.; Zhou, H. Salmonella enterica serovars enteritidis infection alters the indigenous microbiota diversity in young layer chicks. Front. Vet. Sci. 2015, 2, 170740. [Google Scholar] [CrossRef] [PubMed]
- Joat, N.N.; Khan, S.; Chousalkar, K. Understanding the effects of intramuscular injection and feed withdrawal on Salmonella Typhimurium shedding and gut microbiota in pullets. J. Anim. Sci. Biotechnol. 2021, 12, 78. [Google Scholar] [CrossRef]
- Juricova, H.; Videnska, P.; Lukac, M.; Faldynova, M.; Babk, V.; Havickova, H.; Sisak, F.; Rychlik, I. Influence of Salmonella enterica serovar Enteritidis infection on the development of the cecum microbiota in newly hatched chicks. Appl. Environ. Microbiol. 2013, 79, 745–747. [Google Scholar] [CrossRef]
- Videnska, P.; Sisak, F.; Havlickova, H.; Faldynova, M.; Rychlik, I. Influence of Salmonella enterica serovar Enteritidis infection on the composition of chicken cecal microbiota. BMC Vet. Res. 2013, 9, 140. [Google Scholar] [CrossRef]
- Jurburg, S.D.; Brouwer, M.S.M.; Ceccarelli, D.; van der Goot, J.; Jansman, A.J.M.; Bossers, A. Patterns of community assembly in the developing chicken microbiome reveal rapid primary succession. Microbiol. Open 2019, 8, e00821. [Google Scholar] [CrossRef] [PubMed]
- Videnska, P.; Faldynova, M.; Juricova, H.; Babak, V.; Sisak, F.; Havlickova, H.; Rychlik, I. Chicken faecal microbiota and disturbances induced by single or repeated therapy with tetracycline and streptomycin. BMC Vet. Res. 2013, 9, 30. [Google Scholar] [CrossRef]
- Antunes, P.; Réu, C.; Sousa, J.C.; Peixe, L.; Pestana, N. Incidence of Salmonella from poultry products and their susceptibility to antimicrobial agents. Int. J. Food Microbiol. 2003, 82, 97–103. [Google Scholar] [CrossRef]
- Kempf, F.; Menanteau, P.; Rychlik, I.; Kubasová, T.; Trotereau, J.; Virlogeux-Payant, I.; Schaeffer, S.; Schouler, C.; Drumo, R.; Guitton, E.; et al. Gut microbiota composition before infection determines the Salmonella super- and low-shedder phenotypes in chicken. Microb. Biotechnol. 2020, 13, 1611–1630. [Google Scholar] [CrossRef]
- Ijaz, A.; Veldhuizen, E.J.A.; Broere, F.; Rutten, V.P.M.G.; Jansen, C. The interplay between Salmonella and intestinal innate immune cells in chickens. Pathogens 2021, 10, 1512. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Lin, L.; Zheng, L.; Tang, H.; Fan, X.; Xue, N.; Liu, M.; Li, X. Cecal microbiome profile altered by Salmonella enterica, serovar Enteritidis inoculation in chicken. Gut Pathog. 2018, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Sadeyen, J.R.; Trotereau, J.; Velge, P.; Marly, J.; Beaumont, C.; Barrow, P.A.; Bumstead, N.; Lalmanach, A.C. Salmonella carrier state in chicken: Comparison of expression of immune response genes between susceptible and resistant animals. Microbes Infect. 2004, 6, 1778–1786. [Google Scholar] [CrossRef] [PubMed]
- Sadeyen, J.R.; Trotereau, J.; Protais, J.; Beaumont, C.; Sellier, N.; Salvat, G.; Velge, P.; Lalmanach, A.C. Salmonella carrier-state in hens: Study of host resistance by a gene expression approach. Microbes Infect. 2016, 8, 1308–1314. [Google Scholar] [CrossRef]
- Menanteau, P.; Kempf, F.; Trotereau, J.; Virlogeux-Payant, I.; Gitton, E.; Dalifard, J.; Gabriel, I.; Rychlik, I.; Velge, P. Role of systemic infection, cross contaminations, and super-shedders in Salmonella carrier state in chicken. Environ. Microbiol. 2018, 20, 3246–3260. [Google Scholar] [CrossRef]
- Varmuzova, K.; Kubasova, T.; Davidova-Gerzova, L.; Sisak, F.; Havlickova, H.; Sebkova, A.; Faldynova, M.; Rychlik, I. Composition of gut microbiota influences resistance of newly hatched chickens to Salmonella Enteritidis infection. Front. Microbiol. 2016, 7, 957. [Google Scholar] [CrossRef]
- Mon, K.K.Z.; Zhu, Y.; Chanthavixay, G.; Kern, C.; Zhou, H. Integrative analysis of gut microbiome and metabolites revealed novel mechanisms of intestinal Salmonella carriage in chicken. Sci. Rep. 2020, 10, 4809. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Lei, C.; Wang, Y.; Chen, Y.; Zhang, X.; Kang, Z.; Zhai, X.; Ye, X.; Wang, H. Distribution of Salmonella Enteritidis in internal organs and variation of cecum microbiota in chicken after oral challenge. Microb. Pathog 2018, 122, 174–179. [Google Scholar] [CrossRef]
- Robinson, K.; Assumpcao, A.L.F.V.; Arsi, K.; Erf, G.F.; Donoghue, A.; Jesudhasan, P.R.R. Effect of Salmonella Typhimurium colonization on microbiota maturation and blood leukocyte populations in broiler chickens. Animals 2022, 12, 2867. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Li, G.; Shahid, M.S.; Gan, L.; Fan, H.; Lv, Z.; Yan, S.; Guo, Y. L-arginine supplementation ameliorates inflammatory response and alters gut microbiota composition in broiler chickens infected with Salmonella enterica serovar Typhimurium. Poult. Sci. 2020, 99, 1862–1874. [Google Scholar] [CrossRef]
- Khan, S.; Chousalkar, K.K. Salmonella Typhimurium infection disrupts but continuous feeding of Bacillus based probiotic restores gut microbiota in infected hens. J. Anim. Sci. Biotechnol. 2020, 11, 29. [Google Scholar] [CrossRef]
- Rivera-Chávez, F.; Zhang, L.F.; Faber, F.; Lopez, C.A.; Byndloss, M.X.; Olsan, E.E.; Xu, G.; Velazquez, E.M.; Lebrilla, C.B.; Winter, S.E.; et al. Depletion of Butyrate-Producing Clostridia from the Gut Microbiota Drives an Aerobic Luminal Expansion of Salmonella. Cell Host Microbe 2016, 19, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.; Ma, B.; Zhai, X.; Zhang, A.; Lei, C.; Zuo, L.; Yang, X.; Zhou, C.; Wang, H. Florfenicol enhances colonization of a Salmonella enterica serovar Enteritidis floR mutant with major alterations in the intestinal microbiota and metabolome in neonatal chickens. Appl. Environ. Microbiol. 2021, 87, e01681-21. [Google Scholar] [CrossRef]
- Bescucci, D.M.; Montina, T.; Boras, V.E.; Inglis, C.D. Infection by Salmonella enterica serovar Typhimurium DT104 modulates immune responses, the metabolome, and the function of the enteric microbiota in neonatal broiler chickens. Pathogens 2022, 11, 1257. [Google Scholar] [CrossRef]
- Bogomolnaya, L.; Talamantes, M.; Rocha, J.; Nagarajan, A.; Zhu, W.; Spiga, L.; Winter, M.G.; Konganti, K.; Adams, L.G.; Winter, S.; et al. Taxonomic and metagenomic analyses define the development of the microbiota in the chick. mBio 2022, 14, e02444. [Google Scholar] [CrossRef] [PubMed]
- Konashi, S.; Takahashi, S.; Ashiba, Y. Effects of dietary essential amno acids deficiencies on immunological variables in broiler chickens. Br. J. Nutr. 2000, 83, 449–456. [Google Scholar] [PubMed]
- Steeb, B.; Claudi, B.; Burton, N.A.; Tienz, P.; Schmidt, A.; Farhan, H.; Mazé, A.; Bumann, D. Parallel exploitation of diverse host nutrients enhances Salmonella virulence. PLoS Pathog. 2013, 9, e1003301. [Google Scholar] [CrossRef]
- Tattoli, I.; Sorbara, M.T.; Vuckovic, D.; Ling, A.; Soares, F.; Carneiro, L.A.; Yang, C.; Emili, A.; Philpott, D.J.; Girardin, S.E. Amino acid starvation induced by invasive bacterial pathogens triggers an innate host defense program. Cell Host Microbe 2012, 11, 563–575. [Google Scholar] [CrossRef]
- Troha, K.; Ayres, J.S. Metabolic adaptations to infections at the organismal level. Trend Immunol. 2020, 41, 113–125. [Google Scholar] [CrossRef]
Figure 1.
Effect of Salmonella infection on neurotrophin signaling pathway in cecum. All peptides encircled are significantly dephosphorylated during Salmonella infection when compared to non-infected cecal tissue.
Figure 1.
Effect of Salmonella infection on neurotrophin signaling pathway in cecum. All peptides encircled are significantly dephosphorylated during Salmonella infection when compared to non-infected cecal tissue.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kogut, M.H.; Fernandez Miyakawa, M.E. Phenotype Alterations in the Cecal Ecosystem Involved in the Asymptomatic Intestinal Persistence of Paratyphoid Salmonella in Chickens. Animals 2023, 13, 2824. https://doi.org/10.3390/ani13182824
AMA Style
Kogut MH, Fernandez Miyakawa ME. Phenotype Alterations in the Cecal Ecosystem Involved in the Asymptomatic Intestinal Persistence of Paratyphoid Salmonella in Chickens. Animals. 2023; 13(18):2824. https://doi.org/10.3390/ani13182824
Chicago/Turabian StyleKogut, Michael H., and Mariano Enrique Fernandez Miyakawa. 2023. "Phenotype Alterations in the Cecal Ecosystem Involved in the Asymptomatic Intestinal Persistence of Paratyphoid Salmonella in Chickens" Animals 13, no. 18: 2824. https://doi.org/10.3390/ani13182824
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.