
Identification of the Keratin-Associated Protein 22-2 Gene in the Capra hircus and Association of Its Variation with Cashmere Traits
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Geographic Location, and Growing Conditions
2.2. Blood and Cashmere Samples of Goats
2.3. Tissue Samples
2.4. Search for a Caprine Homologue of Sheep KRTAP22-2 in the Goat Genome
2.5. Primer Design and Amplification
2.6. Screening for Variation in Caprine KRTAP22-2
2.7. Sequencing and Analysis of Allelic Variants
2.8. In Situ Hybridization Analysis
2.9. Analysis of the Association between Genetic Variation and Cashmere Traits
3. Results
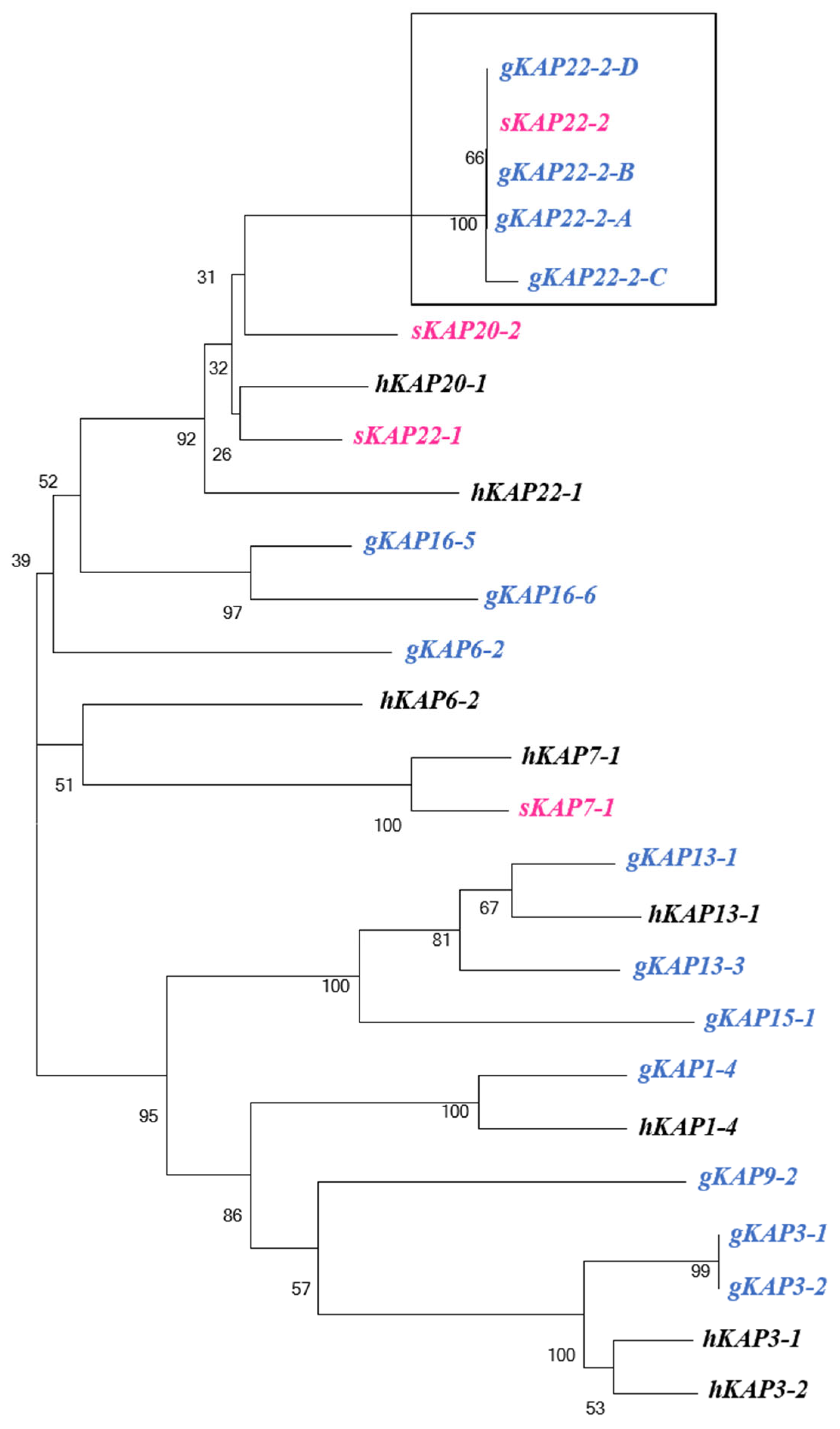
3.1. Identification of KRTAP22-2 in the Goat Genome
3.2. Detection of Allelic Variation in Caprine KRTAP22-2
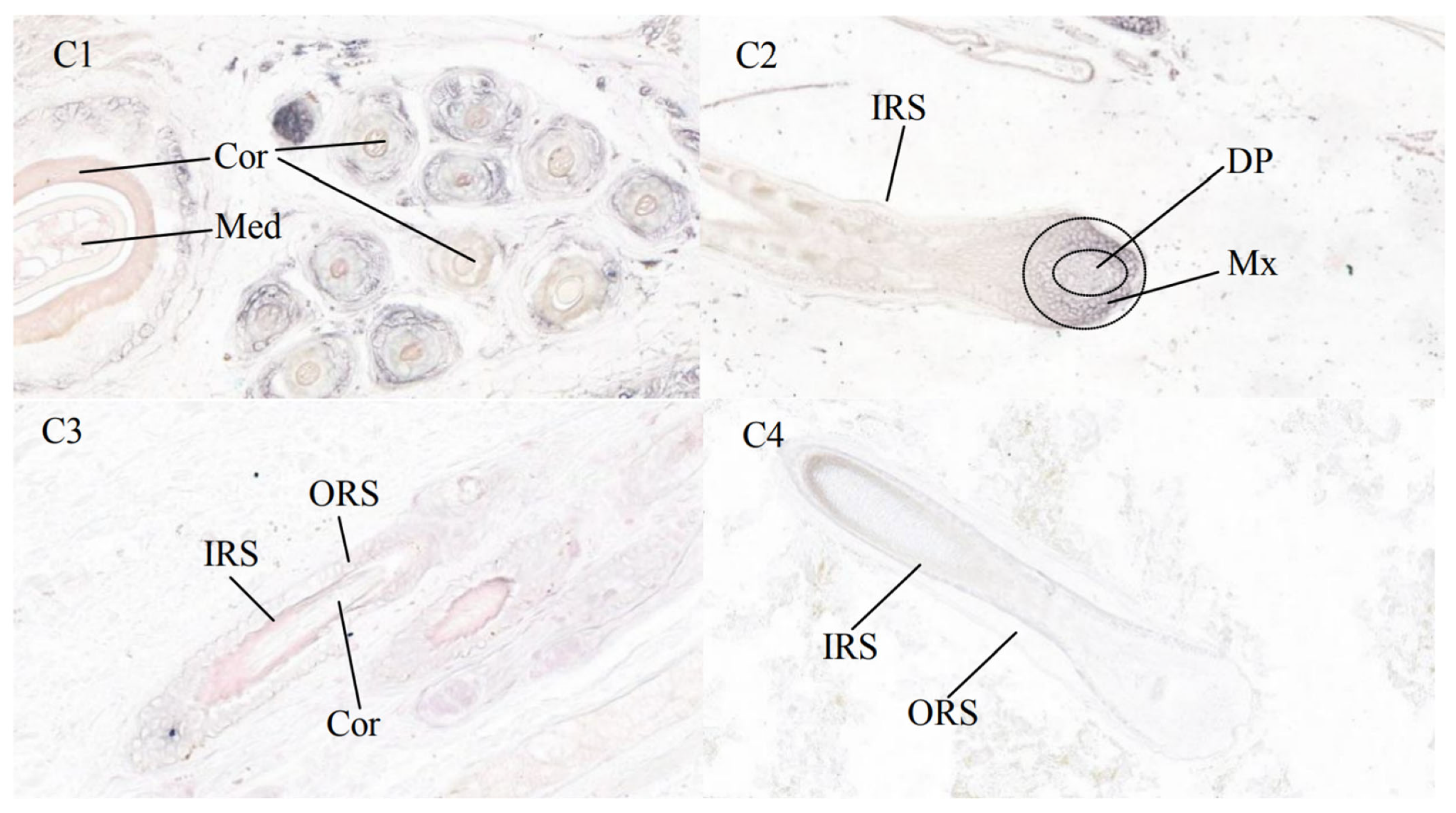
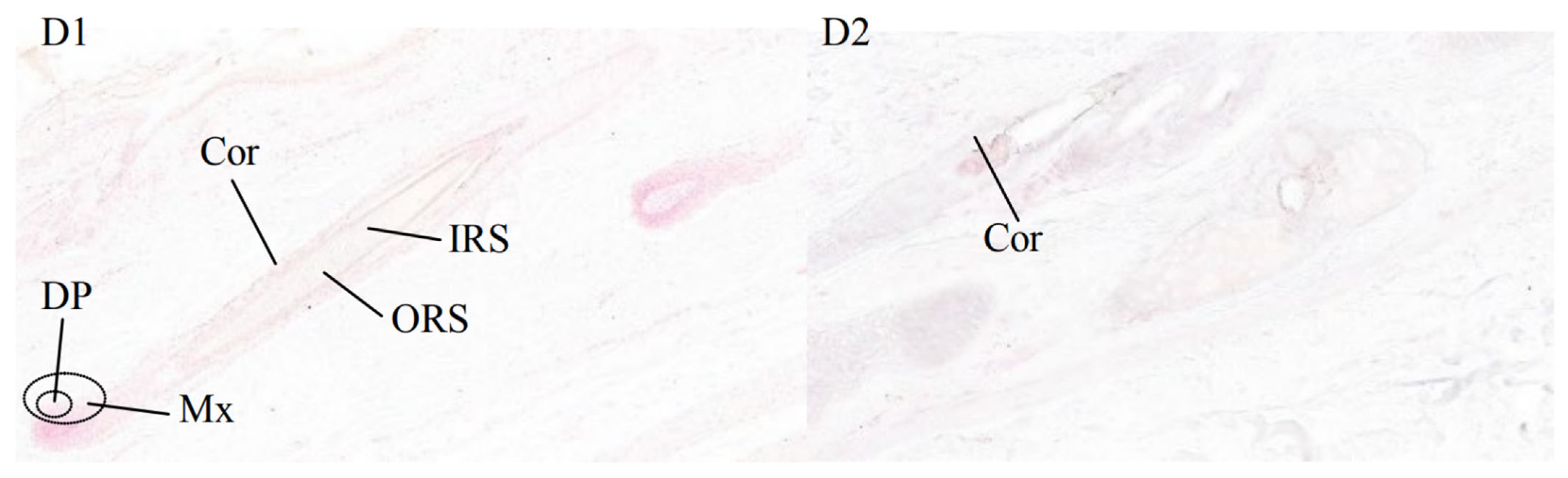
3.3. Results of In Situ Hybridization
3.4. Association Analysis of the KRTAP22-2 Gene with Cashmere Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Z.; Li, H.; Su, R.; Liu, Z.; Li, J. Relationships between cashmere production and other important economic traits of Inner Mongolia cashmere goats. Ital. J. Anim. Sci. 2019, 18, 1154–1160. [Google Scholar] [CrossRef]
- Katoh, K.; Shibayama, M.; Tanabe, T.; Yamauchi, K. Preparation and physicochemical properties of compression-molded keratin films. Biomaterials 2004, 25, 2265–2272. [Google Scholar] [CrossRef]
- Powell, B.C.; Rogers, G.E. The role of keratin proteins and their genes in the growth, structure and properties of hair. EXS 1997, 78, 59–148. [Google Scholar] [CrossRef]
- Strasser, B.; Mlitz, V.; Hermann, M.; Tschachler, E.; Eckhart, L. Convergent evolution of cysteine-rich proteins in feathers and hair. BMC Evol. Biol. 2015, 15, 82. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; McKenzie, G.W.; Yu, Z.; Clerens, S.; Dyer, J.M.; Plowman, J.E.; Wright, M.W.; Arora, R.; Bawden, C.S.; et al. An updated nomenclature for keratin-associated proteins (KAPs). Int. J. Biol. Sci. 2012, 8, 258–264. [Google Scholar] [CrossRef]
- Rechiche, O.; Plowman, J.E.; Harland, D.P.; Lee, T.V.; Lott, J.S. Expression and purification of high sulfur and high glycine-tyrosine keratin-associated proteins (KAPs) for biochemical and biophysical characterization. Protein Expr. Purif. 2018, 146, S1046592817307106. [Google Scholar] [CrossRef]
- Bai, L.; Wang, J.; Zhou, H.; Gong, H.; Tao, J.; Hickford, J.G.H. Identification of Ovine KRTAP28-1 and Its Association with Wool Fibre Diameter. Animals 2019, 9, 142. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.T.; Dyer, J.M.; Hickford, J.G.H. The sheep KAP8-2 gene, a new KAP8 family member that is absent in humans. SpringerPlus 2014, 3, 528. [Google Scholar] [CrossRef]
- Liu, H.; Li, N.; Jia, C.; Zhu, X.; Jia, Z. Effect of the Polymorphisms of Keratin Associated Protein 8.2 Gene on Fibre Traits in Inner Mongolia Cashmere Goats. Asian Australas. J. Anim. Sci. 2007, 20, 821–826. [Google Scholar] [CrossRef]
- Rogers, M.A.; Langbein, L.; Praetzel-Wunder, S.; Giehl, K. Characterization and expression analysis of the hair keratin associated protein KAP26.1. Br. J. Dermatol. 2008, 159, 725–729. [Google Scholar] [CrossRef]
- Rogers, M.A.; Winter, H.; Langbein, L.; Wollschlager, A.; Praetzel-Wunder, S.; Jave-Suarez, L.F.; Schweizer, J. Characterization of human KAP24.1, a cuticular hair keratin-associated protein with unusual amino-acid composition and repeat structure. J. Invest. Derm. 2007, 127, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Schweizer, J. Human KAP genes, only the half of it? Extensive size polymorphisms in hair keratin-associated protein genes. J. Invest. Derm. 2005, 124, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Perutz, M.F. The Role of Aromatic Rings as Hydrogen-Bond Acceptors in Molecular Recognition. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 1993, 345, 105–112. [Google Scholar]
- Fraser, R.D.; Parry, D.A. Filamentous Structure of Hard beta-Keratins in the Epidermal Appendages of Birds and Reptiles. Subcell. Biochem. 2017, 82, 231–252. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G.R.; Hickford, J.G.; Bickerstaffe, R. Polymorphism in two genes for B2 high sulfur proteins of wool. Anim. Genet. 1994, 25, 407–415. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Plowman, J.E.; Dyer, J.M.; Hickford, J.G. Analysis of variation in the ovine ultra-high sulphur keratin-associated protein KAP5-4 gene using PCR-SSCP technique. Electrophoresis 2010, 31, 3545–3547. [Google Scholar] [CrossRef]
- Kiktev, D.A.; Sheng, Z.; Lobachev, K.S.; Petes, T.D. GC content elevates mutation and recombination rates in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2018, 115, E7109–E7118. [Google Scholar] [CrossRef]
- Zhou, H.; Hickford, J.G.; Fang, Q. A two-step procedure for extracting genomic DNA from dried blood spots on filter paper for polymerase chain reaction amplification. Anal. Biochem. 2006, 354, 159–161. [Google Scholar] [CrossRef]
- Byun, S.O.; Fang, Q.; Zhou, H.; Hickford, J.G. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal. Biochem. 2009, 385, 174–175. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Hickford, J.G. Diversity of the glycine/tyrosine-rich keratin-associated protein 6 gene (KAP6) family in sheep. Mol. Biol. Rep. 2011, 38, 31–35. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Forrest, R.H.; Li, S.; Wang, J.; Dyer, J.M.; Luo, Y.; Hickford, J.G. Wool Keratin-Associated Protein Genes in Sheep-A Review. Genes 2016, 7, 24. [Google Scholar] [CrossRef]
- Kimchi-Sarfaty, C. A “silent” polymorphism in the MDR1 gene changes substrate specificity. Science 2007, 315, 525–528. [Google Scholar] [CrossRef]
- Andken, B.B.; Lim, I.; Benson, G.; Vincent, J.J.; Ferenc, M.T.; Heinrich, B.; Jarzylo, L.A.; Man, H.Y.; Deshler, J.O. 3’-UTR SIRF: A database for identifying clusters of whort interspersed repeats in 3’ untranslated regions. BMC Bioinform. 2007, 8, 274. [Google Scholar] [CrossRef] [PubMed]
- Daverio, M.S.; Anello, M.; Alcolea Ersinger, V.; Alvarez, S.; Frank, E.; Vidal-Rioja, L.; Di Rocco, F. Identification of llama KRTAP7-1 and KRTAP8-1 fiber genes and polymorphism screening. Small Rumin. Res. 2019, 175, 149–154. [Google Scholar] [CrossRef]
- Ullah, F.; Jamal, S.M.; Ekegbu, U.J.; Haruna, I.L.; Zhou, H.T.; Hickford, J.G.H. Polymorphism in the ovine keratin-associated protein gene KRTAP7-1 and its association with wool characteristics. J. Anim. Sci. 2020, 98, skz381. [Google Scholar] [CrossRef]
- Ullah, F.; Jamal, S.M.; Zhou, H.; Hickford, J.G.H. Variation in the KRTAP6-3 gene and its association with wool characteristics in Pakistani sheep breeds and breed-crosses. Trop. Anim. Health Prod. 2020, 52, 3035–3043. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Jamal, S.M.; Zhou, H.T.; Hickford, J.G.H. Variation in ovine KRTAP8-1 affects mean staple length and opacity of wool fiber. Anim. Biotechnol. 2021, 34, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.M.; Gad-Allah, A.A.; Albetar, E.M. Genetic variation in the ovine KAP22-1 gene and its effect on wool traits in Egyptian sheep. Arch. Anim. Breed. 2022, 65, 293–300. [Google Scholar] [CrossRef]
- Shanaz, S.; Ganai, T.A.; Ahmad, S.F.; Khan, A.A.; Shabir, N.; Shah, R.; Mir, S.; Ganai, N.A. Molecular characterization, polymorphism and association of Keratin-Associated Protein 13.1 (KAP13.1) gene in Changthangi goats. Biol. Rhythm. Res. 2020, 51, 915–924. [Google Scholar] [CrossRef]
- Sallam, A.M. Association analysis of the ovine KAP6-1 gene and wool traits in Barki sheep. Anim. Biotechnol. 2020, 32, 733–739. [Google Scholar] [CrossRef]
- Michalik, M.; Orwick-Rydmark, M.; Habeck, M.; Alva, V.; Arnold, T.; Linke, D. An evolutionarily conserved glycine-tyrosine motif forms a folding core in outer membrane proteins. PLoS ONE 2017, 12, e0182016. [Google Scholar] [CrossRef]
- Rogers, M.A.; Langbein, L.; Winter, H.; Ehmann, C.; Praetzel, S.; Schweizer, J. Characterization of a first domain of human high glycine-tyrosine and high sulfur keratin-associated protein (KAP) genes on chromosome 21q22.1. J. Biol. Chem. 2002, 277, 48993–49002. [Google Scholar] [CrossRef]
- Rogers, G.E. Hair follicle differentiation and regulation. Int. J. Dev. Biol. 2004, 48, 163–170. [Google Scholar] [CrossRef]
- Rogers, M.A.; Langbein, L.; Praetzel-Wunder, S.; Winter, H.; Schweizer, J. Human hair keratin-associated proteins (KAPs). Int. Rev. Cytol. 2006, 251, 209–263. [Google Scholar] [CrossRef]
- Jin, M.; Cao, Q.; Wang, R.; Piao, J.; Zhao, F.; Piao, J. Molecular characterization and expression pattern of a novel Keratin-associated protein 11.1 gene in the Liaoning cashmere goat (Capra hircus). Asian-Australas. J. Anim. Sci. 2017, 30, 328–337. [Google Scholar] [CrossRef]
- Powell, B.C.; Nesci, A.; Rogers, G.E. Regulation of keratin gene expression in hair follicle differentiation. Ann. N. Y. Acad. Sci. 1991, 642, 1725577. [Google Scholar] [CrossRef]
- Shimomura, Y.; Aoki, N.; Rogers, M.A.; Langbein, L.; Ito, M. hKAP1.6 and hKAP1.7, two novel human high sulfur keratin-associated proteins are expressed in the hair follicle cortex. J. Investig. Dermatol. 2002, 118, 226–231. [Google Scholar] [CrossRef]
- Rogers, M.A.; Langbein, L.; Winter, H.; Ehmann, C.; Praetzel, S.; Korn, B.; Schweizer, J. Characterization of a cluster of human high/ultrahigh sulfur keratin-associated protein genes embedded in the type I keratin gene domain on chromosome 17q12-21. J. Biol. Chem. 2001, 276, 19440–19451. [Google Scholar] [CrossRef]
- Shimomura, Y.; Aoki, N.; Schweizer, J.; Langbein, L.; Rogers, M.A.; Winter, H.; Ito, M. Polymorphisms in the human high sulfur hair keratin-associated protein 1, KAP1, gene family. J. Biol. Chem. 2002, 277, 45493–45501. [Google Scholar] [CrossRef]
- Shimomura, Y.; Aoki, N.; Rogers, M.A.; Langbein, L.; Schweizer, J.; Ito, M. Characterization of human keratin-associated protein 1 family members. J. Investig. Derm. Symp. Proc. 2003, 8, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Wuliji, T.; Davis, G.H.; Dodds, K.G.; Turner, P.R.; Andrews, R.N.; Bruce, G.D. Production performance, repeatability and heritability estimates for live weight, fleece weight and fiber characteristics of alpacas in New Zealand. Small Rumin. Res. 2000, 37, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Iman, N.Y.; Johnson, C.L.; Russell, W.C.; Stobart, R.H. Estimation of genetic parameters for wool fiber diameter measures. J. Anim. Sci. 1992, 70, 1110–1115. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Projict | Variant | Absent | Present | p Value | ||
|---|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | |||||
| Cashmere yield(g) | A | 410 ± 3.3 | 3 | 415 ± 11.4 | 324 | 0.733 |
| B | 410 ± 3.1 | 281 | 420 ± 15.1 | 46 | 0.523 | |
| Mean fiber diameter(µm) | A | 13.6 ± 0.03 | 3 | 13.6 ± 0.11 | 324 | 0.899 |
| B | 13.5± 0.03 | 281 | 13.8 ± 0.14 | 46 | 0.041 | |
| Cashmere fiber length(cm) | A | 4.2 ± 0.04 | 3 | 4.1 ± 0.12 | 324 | 0.254 |
| B | 4.2 ± 0.03 | 281 | 4.4 ± 0.16 | 46 | 0.149 | |
| Projict | Mean ± SE | p Value | ||
|---|---|---|---|---|
| AA (n = 281) | AB (n = 43) | AC (n = 18) | ||
| Cashmere yield(g) | 405.7 ± 4.5 | 434.2 ± 17.3 | 444.1 ± 22.3 | 0.134 |
| Mean fiber diameter(µm) | 13.5 ± 0.04 b | 14.1 ± 0.21 a | 13.6 ± 0.16 b | 0.009 |
| Crimped fiber length(cm) | 4.2 ± 0.05 | 4.2 ± 0.18 | 4.4 ± 0.24 | 0.150 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Cao, J.; Zhao, F.; He, Z.; Sun, H.; Wang, J.; Liu, X.; Li, S. Identification of the Keratin-Associated Protein 22-2 Gene in the Capra hircus and Association of Its Variation with Cashmere Traits. Animals 2023, 13, 2806. https://doi.org/10.3390/ani13172806
Chen Z, Cao J, Zhao F, He Z, Sun H, Wang J, Liu X, Li S. Identification of the Keratin-Associated Protein 22-2 Gene in the Capra hircus and Association of Its Variation with Cashmere Traits. Animals. 2023; 13(17):2806. https://doi.org/10.3390/ani13172806
Chicago/Turabian StyleChen, Zhanzhao, Jian Cao, Fangfang Zhao, Zhaohua He, Hongxian Sun, Jiqing Wang, Xiu Liu, and Shaobin Li. 2023. "Identification of the Keratin-Associated Protein 22-2 Gene in the Capra hircus and Association of Its Variation with Cashmere Traits" Animals 13, no. 17: 2806. https://doi.org/10.3390/ani13172806