
Scoparia dulcis L. Extract Relieved High Stocking Density-Induced Stress in Crucian Carp (Carassius auratus)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical Reagent
2.2. Preparation of SDE
2.3. Determination of Flavonoid Content
2.4. Experimental Fish and Diets
2.5. Protection of Dietary AE against High Stocking Density-Induced Stress Assay
2.6. Protection of Dietary AE against CuSO4 Exposure-Induced Stress Assay
2.7. Protection of Dietary AE against Trichlorfon Exposure-Induced Stress Assay
2.8. Biochemical Analysis
2.9. Statistical Analysis
3. Results
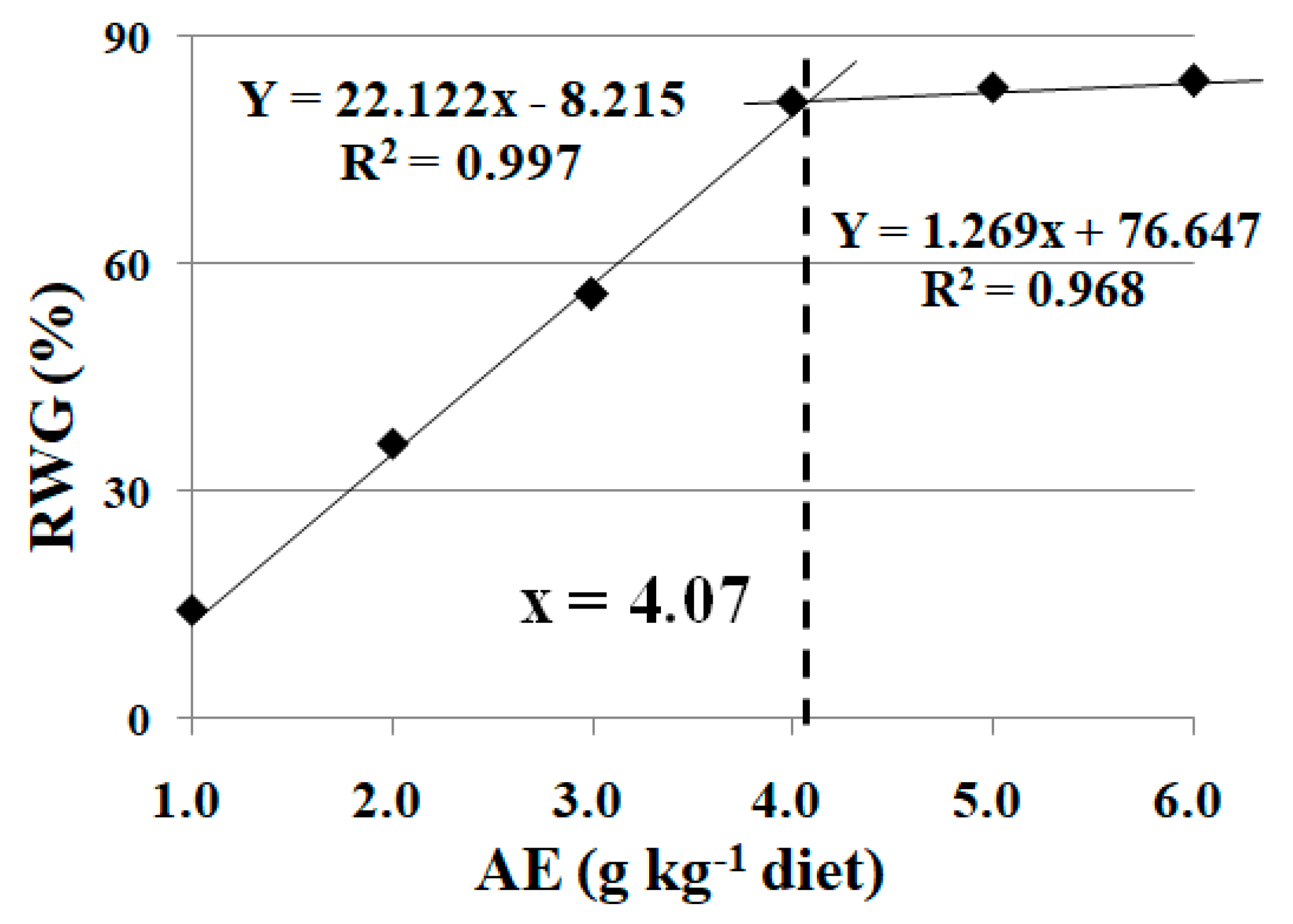
3.1. Effects of Dietary AE on Fish Growth Performance under High Stocking Density
3.2. Effects of Dietary AE on Fish Digestive and Absorptive Enzymatic Activities and Antioxidant Capacity under High Stocking Density
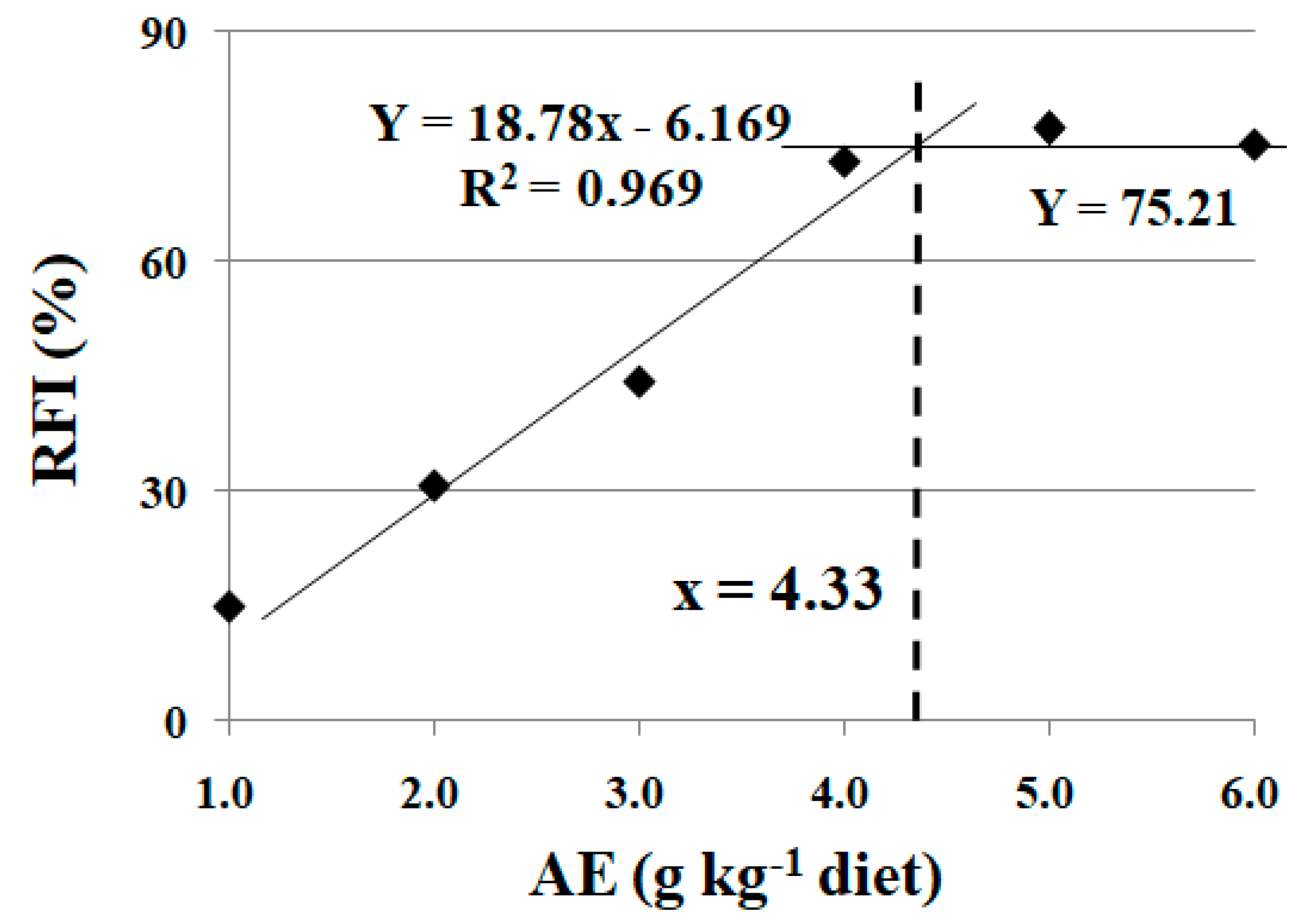
3.3. Effects of Dietary AE on Feed Intake of Fish under Cu Exposure
3.4. Effects of Dietary AE on Digestive and Antioxidant Parameters in Fish under Cu Exposure
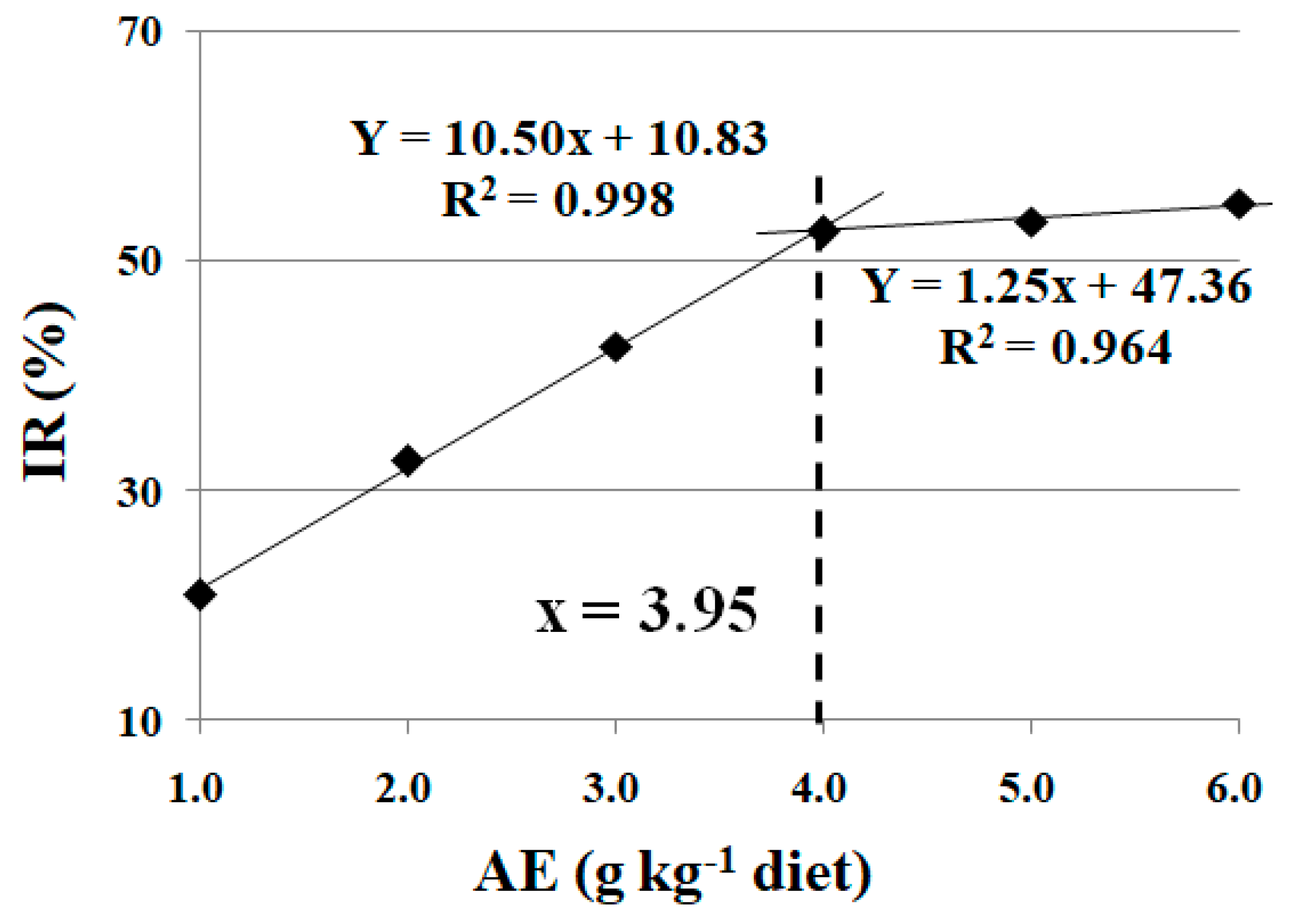
3.5. Effects of Dietary AE on Rollover Rate of Fish under Trichlorfon-Induced Stress
3.6. Effects of Dietary AE on Metabolic Parameters and Antioxidant Status in Fish Muscle under Trichlorfon-Induced Stress
4. Discussion
4.1. Dietary SDE Relieves the Detrimental Effects of High Stocking Density and Cu Exposure on Fish Growth Performance and Feed Intake
4.2. Dietary SDE Relieves the Detrimental Effects of High Stocking Density and Cu Exposure on Fish Digestive Ability
4.3. Dietary SDE Relieves the Detrimental Effects of High Stocking Density and Cu Exposure on Antioxidant Status in Fish Digestive Organs
4.4. Dietary SDE Relieves the Detrimental Effects of Trichlorfon Exposure on Fish Muscle Function and Bioenergetic Homeostasis
4.5. Dietary SDE Relieves the Detrimental Effect of Trichlorfon Exposure on Antioxidant Status in Fish Muscle
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Braun, N.; de Lima, R.L.; Baldisserotto, B.; Dafre, A.L.; de Oliveira Nuñer, A.P. Growth, biochemical and physiological responses of Salminus brasiliensis with different stocking densities and handling. Aquaculture 2010, 301, 22–30. [Google Scholar] [CrossRef]
- Ellis, T.; North, B.; Scott, A.P.; Bromage, N.R.; Porter, M.; Gadd, D. The relationships between stocking density and welfare in farmed rainbow trout. J. Fish Biol. 2002, 61, 493–531. [Google Scholar] [CrossRef]
- Kucukbay, F.Z.; Yazlak, H.; Karaca, I.; Sahin, N.; Tuzcu, M.; Cakmak, M.N.; Sahin, K. The effects of dietary organic or inorganic selenium in rainbow trout (Oncorhynchus mykiss) under crowding conditions. Aquacult. Nutr. 2009, 15, 569–576. [Google Scholar] [CrossRef]
- Chen, G.F.; Feng, L.; Kuang, S.Y.; Liu, Y.; Jiang, J.; Hu, K.; Jiang, W.D.; Li, S.H.; Tang, L.; Zhou, X.Q. Effect of dietary arginine on growth, intestinal enzyme activities and gene expression in muscle, hepatopancreas and intestine of juvenile Jian carp (Cyprinus carpio var. Jian). Br. J. Nutr. 2012, 108, 195–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.C.; Lin, C.H. Toxicity of copper sulfate for survival, growth, molting and feeding of juveniles of the tiger shrimp, Penaeus monodon. Aquaculture 2001, 192, 55–65. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, L.; Jiang, J.; Liu, Y.; Zhou, X.Q. Effects of dietary protein levels on the growth performance, digestive capacity and amino acid metabolism of juvenile Jian carp (Cyprinus carpio var. Jian). Aquac. Res. 2009, 40, 1073–1082. [Google Scholar] [CrossRef]
- Jiang, W.D.; Wu, P.; Kuang, S.Y.; Liu, Y.; Jiang, J.; Hu, K.; Li, S.H.; Tang, L.; Feng, L.; Zhou, X.Q. Myo-inositol prevents copper-induced oxidative damage and changes in antioxidant capacity in various organs and the enterocytes of juvenile Jian carp (Cyprinus carpio var. Jian). Aquat. Toxicol. 2011, 105, 543–551. [Google Scholar] [CrossRef]
- Thomaz, J.M.; Martins, N.D.; Monteiro, D.A.; Rantin, F.T.; Kalinin, A.L. Cardio-respiratory function and oxidative stress biomarkers in Nile tilapia exposed to the organophosphate insecticide trichlorfon (NEGUVON®). Ecotoxicol. Environ. Saf. 2009, 72, 1413–1424. [Google Scholar] [CrossRef]
- Guimara, A.T.B.; Silva de Assis, H.C.; Boeger, W. The effect of trichlorfon on acetylcholinesterase activity and histopathology of cultivated fish Oreochromis niloticus. Ecotoxicol. Environ. Saf. 2006, 68, 57–62. [Google Scholar] [CrossRef]
- Wang, X.; Chang, X.; Zhao, L.; Feng, J.; Li, H.; Liang, J. Trichlorfon exposure in common carp (Cyprinus carpio L.) leads to oxidative stress, neurotoxicity, and immune responses. Aquaculture 2022, 548, 737681. [Google Scholar] [CrossRef]
- Jiang, Z.; Sung, J.; Wang, X.; Zhang, Y.; Wang, Y.; Zhou, H.; Wen, L. A review on the phytochemistry and pharmacology of the herb Scoparia dulcis L. for the potential treatment of metabolic syndrome. RSC Adv. 2021, 11, 31235–31259. [Google Scholar] [CrossRef]
- Zambonino Infante, J.L.; Cahu, C.L. Ontogeny of the gastrointestinal tract of marine fish larvae. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 477–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinan, K.I.; Bene, K.; Zengin, G.; Diuzheva, A.; Jekő, J.; Cziáky, Z.; Picot-Allain, C.M.N.; Mollica, A.; Rengasamy, K.R.; Mahomoodally, M.F. A comparative study of the HPLC-MS profiles and biological efficiency of different solvent leaf extracts of two African plants: Bersama abyssinica and Scoparia dulcis. Int. J. Environ. Health Res. 2021, 31, 285–297. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, Y.; Yao, W.; Ba, Q.; Wang, H. Effects of Cadmium Exposure on the Immune System and Immunoregulation. Front. Immunol. 2021, 12, 695484. [Google Scholar] [CrossRef] [PubMed]
- Adaikpoh, M.A.; Orhue, N.E.J.; Igbe, I. The protective role of Scoparia dulcis on tissue antioxidant defense system of rats exposed to cadmium. Afr. J. Biotechnol. 2010, 6, 1192–1196. [Google Scholar]
- Latha, M.; Pari, L.; Sitasawad, S.; Bhonde, R. Scoparia dulcis, a traditional antidiabetic plant, protects against streptozotocin induced oxidative stress and apoptosis in vitro and in vivo. J. Biochem. Mol. Toxicol. 2004, 18, 261–272. [Google Scholar] [CrossRef]
- Tsai, J.C.; Peng, W.H.; Chiu, T.H.; Lai, S.C.; Lee, C.Y. Anti-inflammatory Effects of Scoparia dulcis L. and Betulinic Acid. Am. J. Chin. Med. 2011, 39, 943–956. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Ma, Y.; Liu, Y.; Wu, M.; Long, J.; Jing, X.; Zhou, S.; Yuan, P.; Jiang, J. Integrated biomarker parameters response to the toxic effects of high stocking density, CuSO4, and trichlorfon on fish and protective role mediated by Angelica sinensis extract. Fish Physiol. Biochem. 2020, 46, 1679–1698. [Google Scholar] [CrossRef]
- Jia, Z.; Tang, M.; Wu, J. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar]
- Li, H.; Lu, L.; Wu, M.; Xiong, X.; Luo, L.; Ma, Y.; Liu, Y. The effects of dietary extract of mulberry leaf on growth performance, hypoxia-reoxygenation stress and biochemical parameters in various organs of fish. Aquac. Rep. 2020, 18, 100494. [Google Scholar] [CrossRef]
- Li, H.; Wu, M.; Jiang, J.; Sun, X.; Chen, L.; Feng, M.; Yuan, D.; Wen, Z.; Qin, C. The extracts of Angelica sinensis restore the digestive and absorptive capacity through improving antioxidant status in digestive organs of fish treated with trichlorfon. Aquac. Res. 2019, 50, 490–504. [Google Scholar] [CrossRef]
- Li, H.; Jiang, W.; Liu, Y.; Jiang, J.; Zhang, Y.; Wu, P.; Zeng, Y.; Zhou, X.; Feng, L. Dietary glutamine improves the function of erythrocytes through its metabolites in juvenile carp (Cyprinus carpio var. Jian). Aquaculture 2017, 474, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Rowland, S.J.; Mifsud, C.; Nixon, M.; Boyd, P. Effects of stocking density on the performance of the Australian freshwater silver perch (Bidyanus bidyanus) in cages. Aquaculture 2006, 253, 301–308. [Google Scholar] [CrossRef]
- Perumal, P.S.; Anaswara, P.V.; Muthuraman, A.; Krishan, S. Therapeutic potency of saponin rich aqueous extract of Scoparia dulcis L. in alloxan induced diabetes in rats. AYU 2014, 35, 211–217. [Google Scholar] [PubMed] [Green Version]
- Gharedaashi, E.; Nekoubin, H.; Imanpoor, M.; Taghizadeh, V. Effect of copper sulfate on the survival and growth performance of Caspian Sea kutum, Rutilus frisii kutum. SpringerPlus 2013, 2, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanty, M.; Adhikari, S.; Mohanty, P.; Sarangi, N. Role of Waterborne Copper on Survival, Growth and Feed Intake of Indian Major Carp, Cirrhinus mrigala Hamilton. Bull. Environ. Contam. Toxicol. 2009, 82, 559–563. [Google Scholar] [CrossRef]
- Gilloteaux, J.; Kashouty, R.; Yono, N. The perinuclear space of pancreatic acinar cells and the synthetic pathway of zymogen in Scorpaena scrofa L.: Ultrastructural aspects. Tissue Cell 2008, 40, 7–20. [Google Scholar] [CrossRef]
- Liu, G.; Ye, Z.; Liu, D.; Zhao, J.; Sivaramasamy, E.; Deng, Y.; Zhu, S. Influence of stocking density on growth, digestive enzyme activities, immune responses, antioxidant of Oreochromis niloticus fingerlings in biofloc systems. Fish Shellfish Immunol. 2018, 81, 416–422. [Google Scholar] [CrossRef]
- Dong, J.; Zhao, Y.Y.; Yu, Y.H.; Sun, N.; Li, Y.D.; Wei, H.; Yang, Z.Q.; Li, X.D.; Li, L. Effect of stocking density on growth performance, digestive enzyme activities, and nonspecific immune parameters of Palaemonetes sinensis. Fish Shellfish Immunol. 2018, 73, 37–41. [Google Scholar] [CrossRef]
- Ni, M.; Liu, M.; Lou, J.; Mi, G.; Yuan, J.; Gu, Z. Stocking density alters growth performance, serum biochemistry, digestive enzymes, immune response, and muscle quality of largemouth bass (Micropterus salmoides) in in-pond raceway system. Fish Physiol. Biochem. 2021, 47, 1243–1255. [Google Scholar] [CrossRef]
- Gonçalves, A.T.; Núñez-Acuña, G.; Détrée, C.; Gallardo-Escárate, C. Coding/non-coding cross-talk in intestinal epithelium transcriptome gives insights on how fish respond to stocking density. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 29, 14–23. [Google Scholar] [CrossRef]
- Wang, T.; Long, X.; Cheng, Y.; Liu, Z.; Yan, S. A Comparison Effect of Copper Nanoparticles versus Copper Sulphate on Juvenile Epinephelus coioides: Growth Parameters, Digestive Enzymes, Body Composition, and Histology as Biomarkers. Int. J. Genom. 2015, 2015, 1–10. [Google Scholar]
- Chairi, H.; Fernández-Diaz, C.; Navas, J.; Manchado, M.; Rebordinos, L.; Blasco, J. In vivo genotoxicity and stress defences in three flatfish species exposed to CuSO4. Ecotoxicol. Environ. Saf. 2010, 73, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Zhao, S.; Chen, G.F.; Jiang, W.D.; Liu, Y.; Jiang, J.; Hu, K.; Li, S.H.; Zhou, X.Q. Antioxidant status of serum, muscle, intestine and hepatopancreas for fish fed graded levels of biotin. Fish Physiol. Biochem. 2013, 40, 499–510. [Google Scholar] [CrossRef]
- Gliszczyńska-Świgło, A. Antioxidant activity of water soluble vitamins in the TEAC (trolox equivalent antioxidant capacity) and the FRAP (ferric reducing antioxidant power) assays. Food Chem. 2006, 96, 131–136. [Google Scholar] [CrossRef]
- Yonar, M.E.; Ispir, U.; Mişe Yonar, S.; Kirici, M. Effect of copper sulphate on the antioxidant parameters in the rainbow trout fry, Oncorhynchus mykiss. Cell. Mol. Biol. 2016, 62, 55–58. [Google Scholar]
- Latha, M.; Pari, L. Modulatory Effect of Scoparia dulcis in Oxidative Stress-Induced Lipid Peroxidation in Streptozotocin Diabetic Rats. J. Med. Food 2003, 6, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Brennan, M.L.; Shen, Z.; MacPherson, J.C.; Schmitt, D.; Molenda, C.E.; Hazen, S.L. Myeloperoxidase functions as a major enzymatic catalyst for initiation of lipid peroxidation at sites of inflammation. J. Biol. Chem. 2002, 277, 46116–46122. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S. Inhibition of mackerel (Scomber scombrus) muscle lipoxygenase by green tea polyphenols. Food Res. Int. 2006, 39, 486–491. [Google Scholar] [CrossRef]
- Ponce, A.M.; Blanco, S.E.; Molina, A.S.; García-Domenech, R.; Gálvez, J. Study of the action of flavonoids on xanthine-oxidase by molecular topology. J. Chem. Inf. Comput. Sci. 2000, 40, 1039–1045. [Google Scholar] [CrossRef]
- Babincová, M.; Sourivong, P. Free Radical Scavenging Activity of Scoparia dulcis Extract. J. Med. Food 2001, 4, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Wankhar, W.; Srinivasan, S.; Rajan, R.; Rathinasamy, S. Evaluating In-Vitro Antioxidant Activity and Gc-Ms Analysis of Scoparia Dulcis Linn (Scrophulariacae). J. App. Pharm. Sci. 2015, 5, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Coulibaly, A.Y.; Kiendrebeogo, M.; Kehoe, P.G.; Sombie, P.A.E.D.; Lamien, C.E.; Millogo, J.F.; Nacoulma, O.G. Antioxidant and Anti-Inflammatory Effects of Scoparia dulcis L. J. Med. Food 2011, 14, 1576–1582. [Google Scholar] [CrossRef] [PubMed]
- Martinez Alvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant defenses in fish: Biotic and abiotic factors. Rev. Fish Biol. Fisher. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Reed, D.J. Glutathione: Toxicological implications. Annu. Rev. Pharmacol. Toxicol. 1990, 30, 603–631. [Google Scholar] [CrossRef] [PubMed]
- Masella, R.; Benedetto, R.D.; Varı, R.; Filesi, C.; Giovannini, C. Novel mechanisms of natural antioxidant compounds in biological systems: Involvement of glutathione and glutathione-related enzymes. J. Nutr. Biochem. 2005, 16, 577–586. [Google Scholar] [CrossRef]
- Latha, M.; Pari, L. Effect of an aqueous extract of Scoparia dulcis on blood glucose, plasma insulin and some polyol pathway enzymes in experimental rat diabetes. Braz. J. Med. Biol. Res. 2004, 37, 577–586. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Liu, G.; Gu, X.; Zhang, T.; Xia, A.; Zheng, Y.; Yin, J.; Han, M.; Jiang, Q. Effects of Quercetin on the Intestinal Microflora of Freshwater Dark Sleeper Odontobutis potamophila. Antioxidants 2022, 11, 2015. [Google Scholar] [CrossRef]
- Singh, M.; Guru, A.; Sudhakaran, G.; Pachaiappan, R.; Mahboob, S.; Al-Ghanim, K.A.; Al-Misned, F.; Juliet, A.; Gobi, M.; Arokiaraj, J. Copper sulfate induced toxicological impact on in-vivo zebrafish larval model protected due to acacetin via anti-inflammatory and glutathione redox mechanism. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 262, 109463. [Google Scholar] [CrossRef]
- Johnston, I.A.; Davison, W.; Goldspink, G. Energy metabolism of carp swimming muscles. J. Comp. Physiol. 1977, 114, 203–216. [Google Scholar] [CrossRef]
- Vieira, L.R.; Sousa, A.; Frasco, M.F.; Lima, I.; Morgado, F.; Guilhermino, L. Acute effects of Benzo[a]pyrene, anthracene and a fuel oil on biomarkers of the common goby Pomatoschistus microps (Teleostei, Gobiidae). Sci. Total Environ. 2008, 395, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Baldissera, M.D.; Souza, C.F.; Parmeggiani, B.; Vendrusculo, R.G.; Ribeiro, L.C.; Muenchen, D.K.; Zeppenfeld, C.C.; Meinhart, A.D.; Wagner, R.; Zanella, R.; et al. Protective effects of diet containing rutin against trichlorfon-induced muscle bioenergetics disruption and impairment on fatty acid profile of silver catfish Rhamdia quelen. Ecotoxicol. Environ. Saf. 2020, 205, 111127. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.P. Amino acids and proteins. In Fish Nutrition; Halver, J.E., Hardy, R.W., Eds.; Academic Press: Cambridge, UK, 2002; pp. 143–181. [Google Scholar]
- Woo, S.J.; Chung, J.K. Effects of trichlorfon on oxidative stress, neurotoxicity, and cortisol levels in common carp, Cyprinus carpio L., at different temperatures. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 229, 108698. [Google Scholar] [CrossRef]
- Pari, L.; Latha, M. Protective role of Scoparia dulcis plant extract on brain antioxidant status and lipidperoxidation in STZ diabetic male Wistar rats. BMC Complement. Altern. Med. 2004, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2017, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Powers, S.K.; Lennon, S.L. Analysis of cellular responses to free radicals: Focus on exercise and skeletal muscle. Proc. Nutr. Soc. 1999, 58, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, P.; Borah, A.; Das, S. Quercetin-induced amelioration of deltamethrin stress in freshwater teleost, Channa punctata: Multiple biomarker analysis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 227, 108626. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Extracts | Flavonoids (mg g−1 Dry Extract) |
|---|---|
| CHE | 30.22 ± 1.68 b |
| EAE | 38.66 ± 1.91 c |
| AE | 67.67 ± 3.36 d |
| AQE | 21.40 ± 1.27 a |
| Ingredients | % | Proximate Analysis 3 | % |
|---|---|---|---|
| Fish meal | 25.0 | Dry matter | 93.16 |
| Soybean meal | 32.0 | Crude protein | 34.83 |
| Wheat flour | 37.0 | Crude lipid | 5.51 |
| DL-methionine | 0.70 | Crude Ash | 5.87 |
| Fish oil | 1.50 | ||
| Sunflower oil | 1.80 | ||
| Vitamin mixture 1 | 1.00 | ||
| Mineral mixture 2 | 1.00 |
| Densities (Fish L−1) + AE (g kg−1 Diet) | IBW (g Fish−1) | FBW (g Fish−1) | WG (g Fish−1) | SGR (% d−1) | FI (g Fish−1) | FE (%) | SR (%) |
|---|---|---|---|---|---|---|---|
| 0.48 + 0 (K) | 8.24 ± 0.32 a | 30.96 ±1.38 d | 22.72 ± 1.30 d | 2.21 ± 0.14 c | 32.14 ± 1.21 d | 70.66 ± 4.22 d | 100.00 ± 0.00 a |
| 0.97 + 0 (Y) | 8.25 ± 0.35 a | 19.74 ± 1.19 a | 11.49 ± 0.81 a | 1.45 ± 0.1 a | 26.98 ± 1.56 a | 42.80 ± 1.45 a | 100.00 ± 0.00 a |
| 0.97 + 1 (E1) | 8.35 ± 0.36 a | 21.45 ± 0.97 a | 13.11 ± 0.58 a | 1.57 ± 0.08 a | 27.86 ± 1.37 ab | 47.17 ± 2.84 ab | 100.00 ± 0.00 a |
| 0.97 + 2 (E2) | 8.34 ± 0.31 a | 23.92 ± 1.52 b | 15.58 ± 1.03 b | 1.75 ± 0.15 b | 28.98 ± 1.30 ab | 53.82 ± 4.13 bc | 100.00 ± 0.00 a |
| 0.97 + 3 (E3) | 8.30 ± 0.41 a | 26.08 ± 1.29 c | 17.78 ± 0.99 c | 1.91 ± 0.11 b | 29.67 ± 1.50 bc | 60.08 ± 4.12 cd | 100.00 ± 0.00 a |
| 0.97 + 4 (E4) | 8.20 ± 0.33 a | 28.85 ± 1.10 d | 20.65 ± 1.38 d | 2.1 ± 0.12 c | 30.90 ± 1.26 cd | 66.89 ± 4.72 d | 100.00 ± 0.00 a |
| 0.97 + 5 (E5) | 8.28 ± 0.40 a | 29.12 ± 1.53 d | 20.84 ± 1.09 d | 2.1 ± 0.14 c | 31.12 ± 1.38 cd | 67.12 ± 2.93 d | 100.00 ± 0.00 a |
| 0.97 + 6 (E6) | 8.16 ± 0.18 a | 29.10 ± 1.42 d | 20.94 ± 1.37 d | 2.12 ± 0.08 c | 31.13 ± 1.48 cd | 67.44 ± 4.02 d | 100.00 ± 0.00 a |
| Densities (Fish L−1) + AE (g kg−1 Diet) | Amylase (U mg−1 Protein) | Lipase (U mg−1 Protein) | ASA (U g−1 Protein) | H2O2 (mmol g−1 Protein) | SOD (U mg−1 rotein) | CAT (U mg−1 Protein) | GSH (mg g−1 Protein) | GR (U g−1 Protein) |
|---|---|---|---|---|---|---|---|---|
| 0.48 + 0 (K) | 1.43 ± 0.07 c | 46.22 ± 2.58 d | 35.91 ± 2.87 a | 16.24 ± 0.86 a | 97.64 ± 7 a | 27.09 ± 1.7 b | 14.24 ± 0.92 c | 14.37 ± 0.88 bc |
| 0.97 + 0 (Y) | 0.91 ± 0.06 a | 30.53 ± 2.64 a | 37.54 ± 1.62 a | 29.62 ± 2.01 c | 100.53 ± 3.4 a | 20.41 ± 1.13 a | 6.42 ± 0.43 a | 10.23 ± 0.9 a |
| 0.97 + 1 (E1) | 0.94 ± 0.07 a | 33.27 ± 2.62 a | 37.49 ± 1.84 a | 28.3 ± 1.56 c | 99.85 ± 7.38 a | 20.5 ± 0.92 a | 6.36 ± 0.26 a | 12.67 ± 0.9 b |
| 0.97 + 2 (E2) | 1.02 ± 0.07 a | 38.26 ± 2.65 b | 46.14 ± 2.71 b | 26.88 ± 1.58 c | 132.88 ± 9.99 c | 25.21 ± 1.43 b | 6.85 ± 0.43 ab | 12.82 ± 0.91 b |
| 0.97 + 3 (E3) | 1.29 ± 0.07 b | 38.63 ± 2.68 bc | 49.17 ± 3.02 bc | 27.02 ± 1.16 c | 134.44 ± 10.92 c | 26.62 ± 1.55 b | 8.4 ± 0.53 b | 14.24 ± 0.92 bc |
| 0.97 + 4 (E4) | 1.4 ± 0.06 bc | 43.37 ± 2.68 cd | 51.87 ± 3.03 c | 19.84 ± 1.1 b | 123.56 ± 6.55 bc | 27.27 ± 1.69 b | 15.01 ± 0.7 c | 14.27 ± 0.92 bc |
| 0.97 + 5 (E5) | 1.45 ± 0.05 c | 43.54 ± 2.6 cd | 47.94 ± 2.97 bc | 15.78 ± 0.59 a | 125.22 ± 7.48 bc | 31.24 ± 2.03 c | 19.49 ± 1.19 d | 15.72 ± 0.89 c |
| 0.97 + 6 (E6) | 1.35 ± 0.09 bc | 40.56 ± 2.7 bc | 45.7 ± 1.66 b | 14.72 ± 1.06 a | 112.61 ± 6.03 ab | 28.64 ± 1.9 bc | 18.61 ± 1.09 d | 15.03 ± 0.92 c |
| AE (g kg−1 Diet) + Cu (mg L−1) | FI (% of Body Weight) |
|---|---|
| 0 + 0.0 (K) | 4.25 ± 0.09 f |
| 0 + 0.7 (Y) | 0.07 ± 0.00 a |
| 1 + 0.7 (E1) | 0.70 ± 0.01 b |
| 2 + 0.7 (E2) | 1.35 ± 0.12 c |
| 3 + 0.7 (E3) | 1.93 ± 0.14 d |
| 4 + 0.7 (E4) | 3.12 ± 0.17 e |
| 5 + 0.7 (E5) | 3.30 ± 0.19 e |
| 6 + 0.7 (E6) | 3.22 ± 0.26 e |
| AE (g kg−1) + Cu (mg L−1) | Amylase (U mg−1 Protein) | Lipase (U g−1 Protein) | MDA (nmol mg−1 Protein) | CAT (U mg−1 Protein) | GSH (mg g−1 Protein) |
|---|---|---|---|---|---|
| 0 + 0.0 (K) | 1.02 ± 0.03 b | 26.95 ± 1.47 a | 12.28 ± 0.71 a | 36.33 ± 1.94 d | 13.87 ± 0.79 e |
| 0 + 0.7 (Y) | 0.78 ± 0.02 a | 28.13 ± 1.53 ab | 21.81 ± 1.18 d | 27.66 ± 1.84 a | 9.2 ± 0.42 a |
| 1 + 0.7 (E1) | 0.98 ± 0.02 b | 29.19 ± 1.47 ab | 19.11 ± 0.85 c | 28.71 ± 1.2 ab | 9.95 ± 0.61 ab |
| 2 + 0.7 (E2) | 1.04 ± 0.05 b | 33.64 ± 1.53 bc | 19.28 ± 1.23 c | 30.38 ± 0.91 abc | 11.48 ± 0.77 cd |
| 3 + 0.7 (E3) | 1.03 ± 0.03 b | 38.24 ± 3.87 c | 15.80 ± 1.00 b | 31.46 ± 1.03 bc | 12.17 ± 0.40 d |
| 4 + 0.7 (E4) | 1.11 ± 0.04 c | 36.9 ± 1.53 c | 13.28 ± 0.87 a | 33.16 ± 1.46 c | 11.32 ± 0.60 cd |
| 5 + 0.7 (E5) | 1.05 ± 0.03 bc | 37.75 ± 4.04 c | 13.98 ± 0.43 ab | 32.87 ± 1.5 c | 10.97 ± 0.48 bcd |
| 6 + 0.7 (E6) | 1.01 ± 0.03 b | 38.31 ± 4.52 c | 14.21 ± 0.52 ab | 31.94 ± 1.37 bc | 10.71 ± 0.41 bc |
| AE (g kg−1 Diet) + Trichlorfon (mg L−1) | Rollover (% of Total) |
|---|---|
| 0 + 0.0 (K) | 0.00 ± 0.00 a |
| 0 + 2.2 (Y) | 100.00 ± 0.00 f |
| 1 + 2.2 (E1) | 79.17 ± 2.89 e |
| 2 + 2.2 (E2) | 67.50 ± 4.33 d |
| 3 + 2.2 (E3) | 57.50 ± 2.50 c |
| 4 + 2.2 (E4) | 47.50 ± 2.50 b |
| 5 + 2.2 (E5) | 46.67 ± 2.89 b |
| 6 + 2.2 (E6) | 45.00 ± 2.50 b |
| AE (g kg−1 Diet) + Trichlorfon (mg L−1) | LDH (U g−1 Protein) | GOT (U g−1 Protein) | GPT (U g−1 Protein) | MDA (nmol mg−1 Protein) | SOD (U mg−1 Protein) | CAT (U mg−1 Protein) | GPx (U mg−1 Protein) |
|---|---|---|---|---|---|---|---|
| 0 + 0.0 (K) | 292.2 ± 24.35 d | 14.94 ± 0.78 c | 16.38 ± 1.02 d | 3.04 ± 0.14 a | 152.36 ± 4.92 cd | 25.05 ± 0.87 d | 381.88 ± 31.25 c |
| 0 + 2.2 (Y) | 207.4 ± 14.37 a | 10.21 ± 0.81 a | 11.44 ± 0.73 a | 4.71 ± 0.23 d | 122.17 ± 5.8 a | 14.08 ± 1.17 a | 236.98 ± 12.07 a |
| 1 + 2.2 (E1) | 207.92 ± 13.85 a | 12.19 ± 1.04 b | 12.69 ± 0.83 ab | 4.64 ± 0.15 d | 129.36 ± 4.61 ab | 16.04 ± 1.13 a | 255.31 ± 20.16 a |
| 2 + 2.2 (E2) | 237.35 ± 14.18 ab | 13.73 ± 0.76 bc | 14.73 ± 0.79 cd | 4.16 ± 0.16 c | 138.02 ± 9.24 bc | 22.23 ± 1.75 bc | 316.32 ± 11.91 b |
| 3 + 2.2 (E3) | 231.91 ± 13.85 ab | 13.42 ± 0.85 bc | 14.66 ± 1.18 cd | 3.7 ± 0.19 b | 147.67 ± 12.19 cd | 21.97 ± 1.86 bc | 313.54 ± 16.91 b |
| 4 + 2.2 (E4) | 269.13 ± 14.57 cd | 16.78 ± 1.24 d | 15.53 ± 0.83 cd | 3.71 ± 0.21 b | 163.65 ± 8.39 d | 23.36 ± 0.78 bcd | 360.37 ± 24.48 c |
| 5 + 2.2 (E5) | 273.24 ± 13.92 cd | 12.39 ± 1.22 b | 15.18 ± 0.52 cd | 3.49 ± 0.15 b | 158.82 ± 6.96 d | 23.56 ± 1.14 cd | 369.1 ± 30.45 c |
| 6 + 2.2 (E6) | 256.3 ± 14.32 bc | 12.75 ± 1.17 b | 14.25 ± 1.16 bc | 3.52 ± 0.2 b | 158.35 ± 11.56 d | 20.92 ± 1.59 b | 356.58 ± 21.22 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, G.; Wu, M.; Li, H.; Xu, J.; Liu, H.; Du, W.; Yang, Q.; Feng, L.; Jiang, J. Scoparia dulcis L. Extract Relieved High Stocking Density-Induced Stress in Crucian Carp (Carassius auratus). Animals 2023, 13, 2522. https://doi.org/10.3390/ani13152522
Chen G, Wu M, Li H, Xu J, Liu H, Du W, Yang Q, Feng L, Jiang J. Scoparia dulcis L. Extract Relieved High Stocking Density-Induced Stress in Crucian Carp (Carassius auratus). Animals. 2023; 13(15):2522. https://doi.org/10.3390/ani13152522
Chicago/Turabian StyleChen, Gangfu, Min Wu, Huatao Li, Jing Xu, Haijing Liu, Wenhao Du, Qihui Yang, Lin Feng, and Jun Jiang. 2023. "Scoparia dulcis L. Extract Relieved High Stocking Density-Induced Stress in Crucian Carp (Carassius auratus)" Animals 13, no. 15: 2522. https://doi.org/10.3390/ani13152522