

Helminths of the Bluntnose Sixgill Shark, Hexanchus griseus (Bonnaterre, 1788), from the Strait of Messina (Sicily, Southern Italy)
 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Macroscopic Evaluation
2.2. Parasitological Evaluation
2.3. Scanning Electron Microscopy Evaluation
3. Results
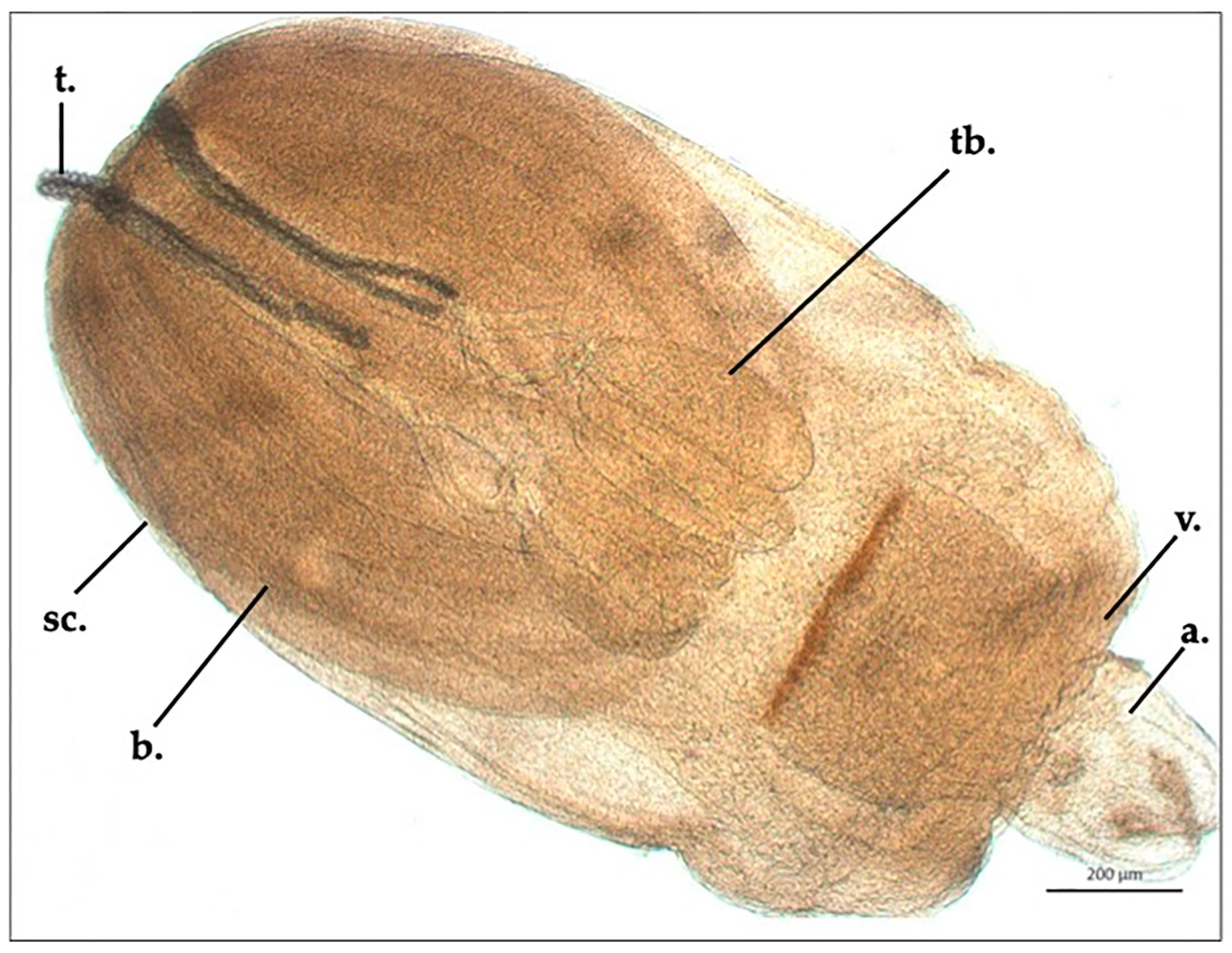
3.1. Nybelinia sp., Larvae (Trypanorhyncha: Tentaculariidae)
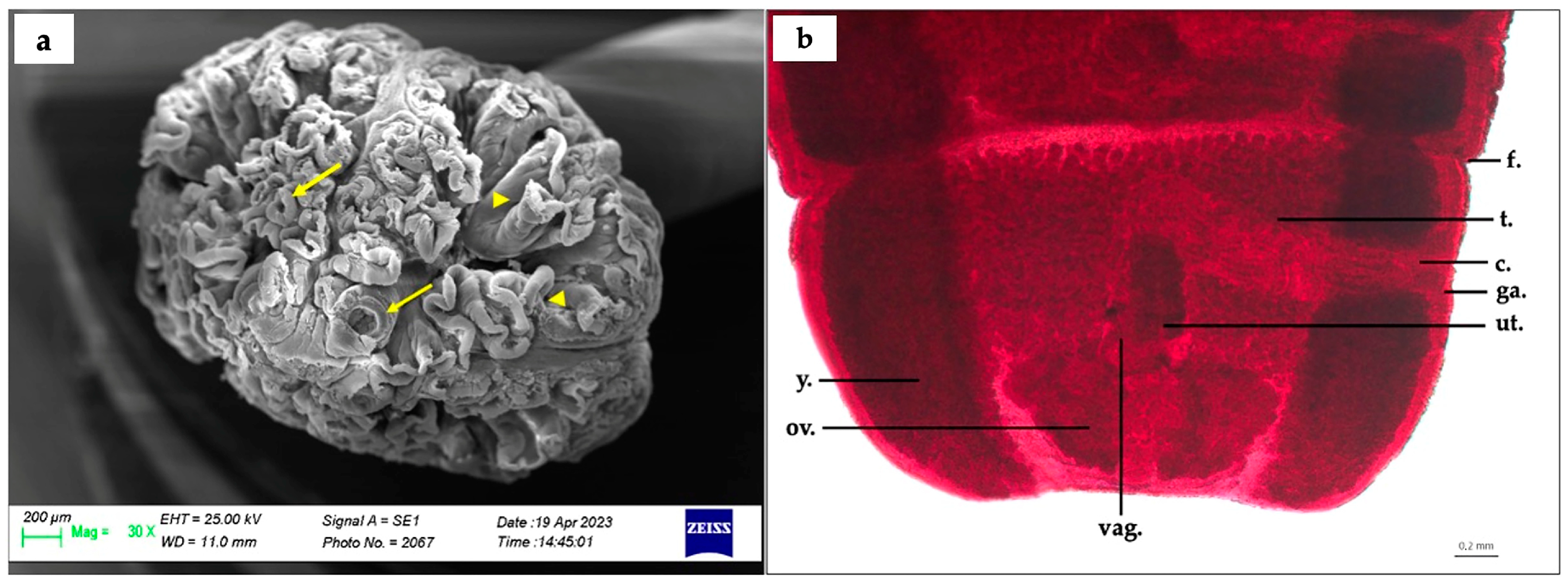
3.2. Phyllobothrium sinuosiceps (Phyllobothriidea: Phyllobothriidae) Adult Specimen
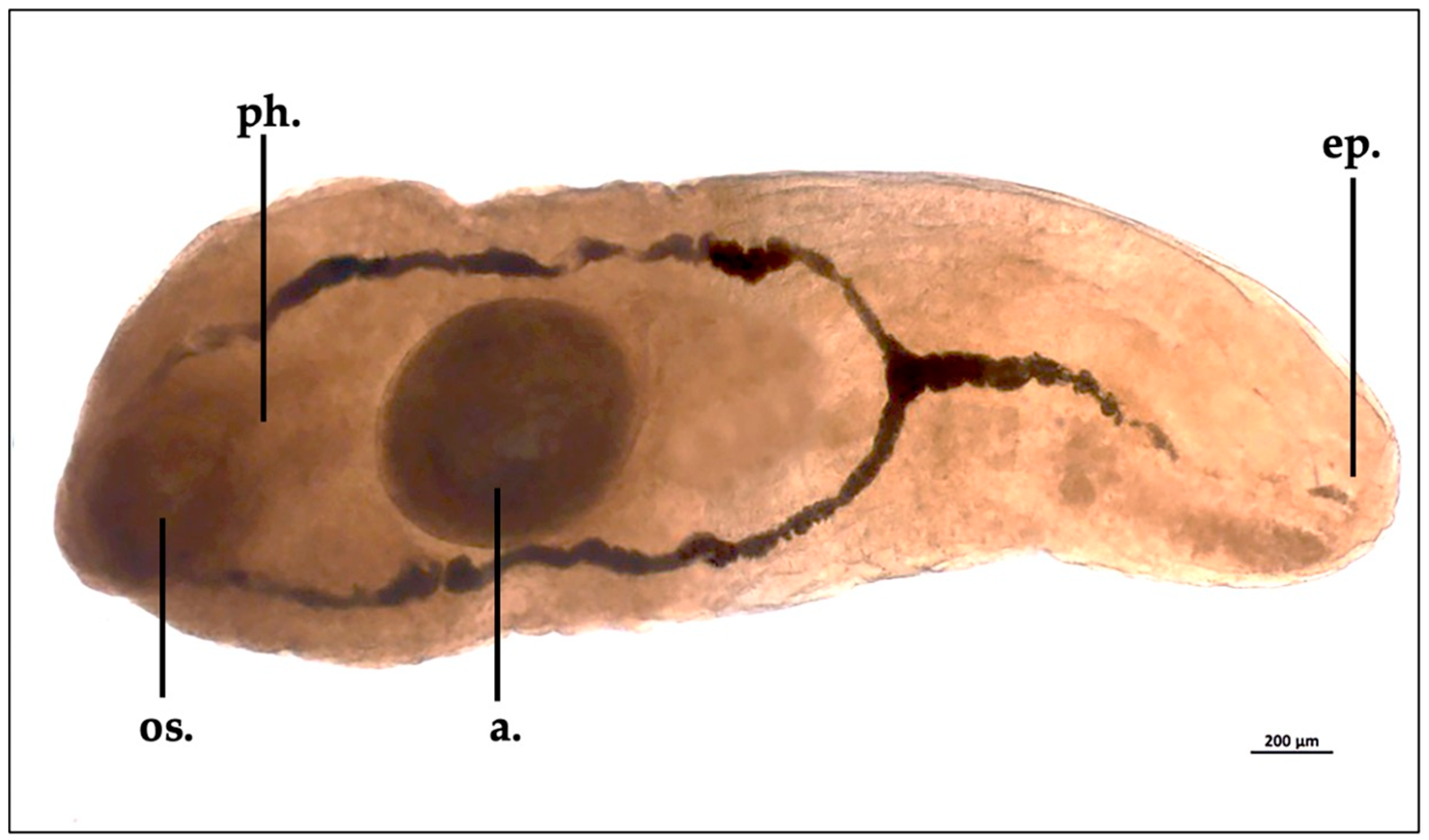
3.3. The trematode Otodistomum veliporum Creplin, 1837 Immature Specimen
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieto, A.; Ralph, G.M.; Comeros-Raynal, M.T.; Heessen, H.J.L.; Rijnsdorp, A.D. European Red List of Marine Fishes; Publications Office of the European Union: Luxembourg, 2015. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References; California Academy of Sciences: San Francisco, CA, USA, 2022. [Google Scholar]
- Barrull, J.; Mate, I.; Bueno, M. Observaciones de tiburones (Chondrichthyes, Euselachii) en aguas de Cataluña, con algunos aspectos generales de su ecología. Sci. Gerund. 1999, 24, 127–151. [Google Scholar]
- Capapé, C.; Hemida, F.; Guélorget, O.; Barrull, J.; Mate, I.; Ben Souissi, J.; Bradaï, M.N. Reproductive biology of the bluntnose sixgill shark Hexanchus griseus (Bonnaterre, 1788) (Chondrichthyes: Hexanchidae) from the Mediterranean Sea: A review. Acta Adriat. 2004, 45, 95–106. [Google Scholar]
- FAO. Species Photographic Plates. Mediterranean Sharks, by Monica Barone, Fabrizio Serena and Mark Dimech. Rome, Italy. 2018. Available online: https://www.fao.org/publications/card/en/c/I8615EN/ (accessed on 26 May 2023).
- Vella, N.; Vella, A. Population genetics of the deep-sea bluntnose sixgill shark, Hexanchus griseus, revealing spatial genetic heterogeneity. Mar. Genom. 2017, 36, 25–32. [Google Scholar] [CrossRef]
- Farrugio, H.; Soldo, A. Sicily Channel/Tunisian Plateau: Status and Conservation of Fisheries. UNEP-MAP-RAC/SPA; Cebrian, D., Requena, S., Eds.; RAC/SPA: Tunis, Tunisia, 2015; p. 76. [Google Scholar]
- Mundy, B.C. Checklist of the fishes of the Hawaiian Archipelago. Bish. Mus. Bull. Zool. 2005, 6, 1–704. [Google Scholar]
- Last, P.R.; Stevens, J.D. Sharks and Rays of Australia. J. Mar. Biol. Assoc. UK 1994, 74, 744. [Google Scholar] [CrossRef]
- Ebert, D.A. Biological Aspects of the Sixgill Shark, Hexanchus griseus. Copeia 1986, 1, 131–135. [Google Scholar] [CrossRef]
- Barrull, J.; Mate, I. Biología de la cañabota Hexanchus griseus (Bonnaterre, 1788) en el Mar Mediterráneo. Bol. Esp. Elasmobranquios 2000, 3, 13–20. [Google Scholar]
- Soldo, A.; Bariche, M.; Buscher, E.; Cook, S.F.; Compagno, L.J.V. Hexanchus griseus. In The IUCN Red List of Threatened Species; IUCN: Geneva, Switzerland, 2016. [Google Scholar]
- Nuez, I.; Giovos, I.; Tiralongo, F.; Penadés-Suay, J.; Ćetković, I.; Di Lorenzo, M.; Kleitou, P.; Bakiu, R.; Bradai, M.; Al Mabruk, S.; et al. Assessing the current status of Hexanchus griseus in the Mediterranean Sea using local ecological knowledge. Mar. Policy 2023, 147, 105378. [Google Scholar] [CrossRef]
- Tserpes, G.; Maravelias, C.D.; Pantazi, M.; Peristeraki, P. Distribution of relatively rare demersal elasmobranchs in the eastern Mediterranean. Estuar. Coast. Shelf Sci. 2013, 117, 48–53. [Google Scholar] [CrossRef]
- Wood, C.L.; Byers, J.E.; Cottingham, K.L.; Altman, I.; Donahue, M.J.; Blakeslee, A.M.H. Parasites alter community structure. Proc. Natl. Acad. Sci. USA 2007, 104, 9335–9339. [Google Scholar] [CrossRef] [PubMed]
- Amundsen, P.A.; Lafferty, K.D.; Knudsen, R.; Primicerio, R.; Klemetsen, A.; Kuris, A.M. Food web topology and parasites in a subarctic lake. J. Anim. Ecol. 2009, 78, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Egusa, T.; Fukushima, K.; Oda, T.; Ohte, N.; Tokuchi, N.; Watanabe, K.; Kanaiwa, M.; Murakami, I.; Lafferty, K.D. Nematomorph parasites indirectly alter the food web and ecosystem function of streams through behavioural manipulation of their cricket hosts. Ecol. Lett. 2012, 15, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.A.; Ball, M.A.; Chubb, J.C. To grow or not to grow? Intermediate and paratenic hosts as helminth life cycle strategies. J. Theor. Biol. 2009, 258, 135–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benesh, D.; Chubb, J.; Parker, G. The trophic transmission vacuum and the evolution of complex helminth life cycles. Proc. Biol. Sci. R. Soc. 2014, 281, 20141462. [Google Scholar] [CrossRef]
- Waeschenbach, A.; Webster, B.L.; Littlewood, D.T. Adding resolution to ordinal level relationships of tapeworms (Platyhelminthes: Cestoda) with large fragments of mtD-NA. Mol. Phyl. Evol. 2012, 63, 834–847. [Google Scholar] [CrossRef]
- Littlewood, D.T.J.; Bray, R.A.; Waeschenbach, A. Phylogenetic patterns of diversity in the cestodes and trematodes. In Parasite Diversity and Diversification: Evolutionary Ecology Meets Phylogenetics, 1st ed.; Morand, S., Krasnov, B.R., Littlewood, D.T.J., Eds.; Cambridge University Press: Cambridge, UK, 2015; pp. 304–319. [Google Scholar] [CrossRef]
- Caira, J.N.; Jensen, K.; Waeschenbach, A.; Olson, P.D.; Littlewood, D.T.J. Orders out of chaos—Molecular phylogenetics reveals the complexity of shark and stingray tapeworm relationships. Int. J. Parasitol. 2014, 44, 55–73. [Google Scholar] [CrossRef] [Green Version]
- Caira, J.N.; Jensen, K. A digest of elasmobranch tapeworms. J. Parasitol. 2014, 100, 373–391. [Google Scholar] [CrossRef]
- Carvajal, J.G. Grillotia dollfusi sp. n. (Cestoda: Trypanorhyncha) from the skate, Raja chilensis, from Chile, and a note on G. heptanchi. J. Parasitol. 1971, 57, 1269–1271. [Google Scholar] [CrossRef]
- Carvajal, J.G. Records of cestodes from Chilean sharks. J. Parasitol. 1974, 60, 29–34. [Google Scholar] [CrossRef]
- Beveridge, I.; Campbell, R.A. A new species of Grillotia Guiart, 1927 (Cestoda: Trypanorhyncha) with redescriptions of congeners and new synonyms. Syst. Parasitol. 2013, 85, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Holzer, A.S.; Bartošová-Sojková, P.; Born-Torrijos, A.; Lövy, A.; Hartigan, A.; Fiala, I. The joint evolution of the Myxozoa and their alternate hosts: A cnidarian recipe for success and vast biodiversity. Mol. Ecol. 2018, 27, 1651–1666. [Google Scholar] [CrossRef] [PubMed]
- Lisnerová, M.; Fiala, I.; Cantatore, D.; Irigoitia, M.; Timi, J.; Pecková, H.; Bartošová-Sojková, P.; Sandoval, C.M.; Luer, C.; Morris, J.; et al. Mechanisms and Drivers for the Establishment of Life Cycle Complexity in Myxozoan Parasites. Biology 2020, 1, 10. [Google Scholar] [CrossRef] [Green Version]
- Lisnerová, M.; Martinek, I.N.; Alama-Bermejo, G.; Bouberlová, K.; Schaeffner, B.C.; Nkabi, N.; Holzer, A.S.; Bartošová-Sojková, P. An ancient alliance: Matching evolutionary patterns of cartilaginous fishes (Elasmobranchii) and chloromyxid parasites (Myxozoa). Infect. Genet. Evol. 2022, 103, 105346. [Google Scholar] [CrossRef]
- Henderson, A.C.; Flannery, K.; Dunne, J. Biological Observations on Shark Species Taken in Commercial Fisheries to the West of Ireland. Biol. Environ. 2003, 103B, 1–7. [Google Scholar] [CrossRef]
- Kay, M.W. Otodistomum plicatum n. sp. (Trematoda, Digenea) from Hexanchus griseus (Bonnaterre). Ohio J. Sci. 1947, 47, 79–83. [Google Scholar]
- Gibson, D.I.; Bray, R.A. The Azygiidae, Hirudineliidae, Ptychogonimidae, Sclerodistomidae and Syncoeliidae of fishes from the North-East Atlantic. Bull. Br. Mus. 1977, 32, 167–245. [Google Scholar]
- Knoff, M.; de São Clemente, S.C.; Pinto, R.M.; Gomes, D.C. Digenea and acanthocephala of elasmobranch fishes from the southern coast of Brazil. Mem. Inst. Oswaldo Cruz. 2001, 96, 1095–1101. [Google Scholar] [CrossRef] [Green Version]
- Oldewage, W.H.; Smale, M.J. Occurrence of piscine parasitic copepods (Crustacea) on sharks taken mainly off Cape Recife, South Africa. S. Afr. J. Mar. Sci. 1993, 13, 309–312. [Google Scholar] [CrossRef] [Green Version]
- Cable, R.M. An Illustrated Laboratory Manual of Parasitology; Burgess: Minniapolis, MN, USA, 1961. [Google Scholar]
- Williams, H.H. The anatomy of Phyllobothrium sinuosiceps sp.nov. (Cestoda: Tetraphyllidea) from Hexanchus griseus (Gmelin) the six gilled shark. Parasitology 1959, 49, 54–69. [Google Scholar] [CrossRef]
- Beveridge, I.; Bray, R.A.; Cribb, T.H.; Justine, J.L. Diversity of trypanorhynch metacestodes in teleost fishes from coral reefs off eastern Australia and New Caledonia. Parasite 2014, 21, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinero, F.R.; Milazzo, C.; Minervino, M.; Marchio, C.; Filice, M.; Bevacqua, L.; Giglio, G.; Leonetti, F.L.; Micarelli, P.; Tripepi, S.; et al. Parasitic Load, Hematological Parameters, and Trace Elements Accumulation in the Lesser Spotted Dogfish Scyliorhinus canicula from the Central Tyrrhenian Sea. Biology 2022, 11, 663. [Google Scholar] [CrossRef] [PubMed]
- Pascual, S.; González, A.F.; Arias, C.; Guerra, A. Biotic relationships of Illex coindetii and Todaropsis eblanae (Cephalopoda, Ommastrephidae) in the Northeastern Atlantic: Evidence from parasites. Sarsia 1996, 81, 265–274. [Google Scholar] [CrossRef]
- Pascual, S.; Abollo, E.; Mladineo, I.; Gestal, C. Metazoa and Related Diseases. In Handbook of Pathogens and Diseases in Cephalopods; Gestal, C., Pascual, S., Guerra, Á., Fiorito, G., Vieites, J.M., Eds.; Springer International Publishing: Cham, Switzerland, 2019; Volume 1, pp. 169–179. [Google Scholar]
- Gestal, C.; Abollo, E.; Arias, C.; Pascual, S.S.E.M. study of the plerocercoid larval Phyllobothrium sp. (Tetraphyllidea, Phyllobothriidae) and Nybelinia lingualis (Trypanothyncha, Tentaculariidae), cestode parasite in Octopus vulgaris (Mollusca, Cephalopoda) off Vigo estuary. Iberus 1998, 16, 125–132. [Google Scholar]
- Mulas, A.; Bellodi, A.; Cau, A.; Gastoni, A.; Locci, I.; Follesa, M.C. Trophic interactions among Chondrichthyans in the central-western Mediterranean Sea. Biol. Mar. Mediterr. 2011, 18, 81–84. [Google Scholar]
- Palm, H.W.; Walter, T. Tentaculariid cestodes (Trypanorhyncha) from the Muséum national d’Histoire naturelle, Paris. Zoosystema 2000, 22, 641–666. [Google Scholar]
- Hart, J.F. Cestoda from Fishes of Puget Sound. II. Tetrarhynchoidea. Trans. Am. Microsc. Soc. 1936, 55, 369. [Google Scholar] [CrossRef]
- Caira, J.N.; Healy, C.J. Elasmobranchs as hosts of metazoan parasites. In Biology of Sharks and Their Relatives, 2nd ed; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2004; Volume 1, pp. 523–551. [Google Scholar]
- Dick, T.A.; Chambers, C.; Isinguzo, I. Cestoidea (Phylum Platyhelminthes). In Fish Diseases and Disorders: Protozoan and Metazoan Infections, 2nd ed; Woo, P.T.K., Ed.; CAB international: London, UK, 2005; Volume 1, pp. 391–416. [Google Scholar]
- Rasmussen, T.; Randhawa, H. Host Diet Influences Parasite Diversity: A Case Study Looking at Tapeworm Diversity among Sharks. Mar. Ecol. Prog. Ser. 2018, 605, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Pramanik, P.B.; Manna, B. Two new species of the genus Phyllobothrium van Beneden, 1850 from the cartilaginous fish Carcharhinus bleekeri Muller & Henle, 1841 at Digha coastal waters, Bay of Bengal, India. J. Nat. Hist. 2009, 5, 64–72. [Google Scholar]
- Sene, A.; Cheikh, B.; Bernard, M. Ultrastructure of spermiogenesis of Phyllobothrium lactuca (Cestoda, Tetraphyllidea, Phyllobothriidae). Folia Parasitol. 1999, 46, 191–198. [Google Scholar]
- Dicken, M.L.; Hussey, N.E.; Christiansen, H.M.; Smale, M.J.; Nkabi, N.; Cliff, G.; Wintner, S.P. Diet and Trophic Ecology of the Tiger Shark (Galeocerdo Cuvier) from South African Waters. PLoS ONE 2017, 12, e0177897. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, P.; Gustinelli, A.; Caffara, M.; Marsella, A.; Toffan, A.; Fioravanti, M.L. Morphological and Molecular Characterization of Two Gastrointestinal Parasites in Mustelus Mustelus (Linnaeus, 1758) from Adriatic Sea. Eur. Zool. J. 2020, 87, 616–623. [Google Scholar] [CrossRef]
- Williams, H.H. The Taxonomy, Ecology and Host-Specificity of Some Phyllobothriidae (Cestoda: Tetraphyllidea), a Critical Revision of Phyllobothrium Beneden, 1849 and Comments on Some Allied Genera. Phil. Trans. R. Soc. Lond. B 1968, 253, 231–307. [Google Scholar] [CrossRef]
- Threlfall, W.; Carvajal, G.L. Otodistomum cestoides (van Beneden, 1871) from two species of skates taken in Chilean waters. Mem. Inst. Oswaldo Cruz. 1986, 81, 341–342. [Google Scholar] [CrossRef] [Green Version]
- Zdzitowiecki, K.; Pisano, E. New records of Digenea infection elasmobranch and teleost fish off Heard Island (Kerguelen, sub-region, sub-Antarctic). Arch. Fish. Mar. Res. 1996, 43, 265–272. [Google Scholar]
- Gibson, D.I.; Margolis, L. Trematoda. In Guide to the Parasites of Fishes of Canada; Margolis, L., Ed.; Department of Fisheries and Oceans: Ottawa, ON, Canada, 1996; ISBN 9780660164038. [Google Scholar]
- Sperone, E.; Milazzo, C. On the Occurrence of the Digenean Otodistomum Veliporum in the Spiracle of the Kitefin Shark Dalatias licha. Acta Adriat. 2018, 59, 137–140. [Google Scholar] [CrossRef] [Green Version]
- Espínola-Novelo, J.F.; Escribano, R.; Oliva, M.E. Metazoan Parasite Communities of Two Deep-Sea Elasmobranchs: The Southern Lanternshark, Etmopterus Granulosus, and the Largenose Catshark, Apristurus Nasutus, in the Southeastern Pacific Ocean. Parasite 2018, 25, 53. [Google Scholar] [CrossRef] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Benedetto, G.; Capparucci, F.; Iaria, C.; Marino, F.; Gaglio, G. Helminths of the Bluntnose Sixgill Shark, Hexanchus griseus (Bonnaterre, 1788), from the Strait of Messina (Sicily, Southern Italy). Animals 2023, 13, 2405. https://doi.org/10.3390/ani13152405
De Benedetto G, Capparucci F, Iaria C, Marino F, Gaglio G. Helminths of the Bluntnose Sixgill Shark, Hexanchus griseus (Bonnaterre, 1788), from the Strait of Messina (Sicily, Southern Italy). Animals. 2023; 13(15):2405. https://doi.org/10.3390/ani13152405
Chicago/Turabian StyleDe Benedetto, Giovanni, Fabiano Capparucci, Carmelo Iaria, Fabio Marino, and Gabriella Gaglio. 2023. "Helminths of the Bluntnose Sixgill Shark, Hexanchus griseus (Bonnaterre, 1788), from the Strait of Messina (Sicily, Southern Italy)" Animals 13, no. 15: 2405. https://doi.org/10.3390/ani13152405