
Cocaine- and Amphetamine-Regulated Transcript (CART) Peptide Is Co-Expressed with Parvalbumin, Neuropeptide Y and Somatostatin in the Claustrum of the Chinchilla

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Preparation
2.2. Immunohistochemistry
2.3. Image Analysis
2.4. Cell Counting and Statistical Analyses
3. Results
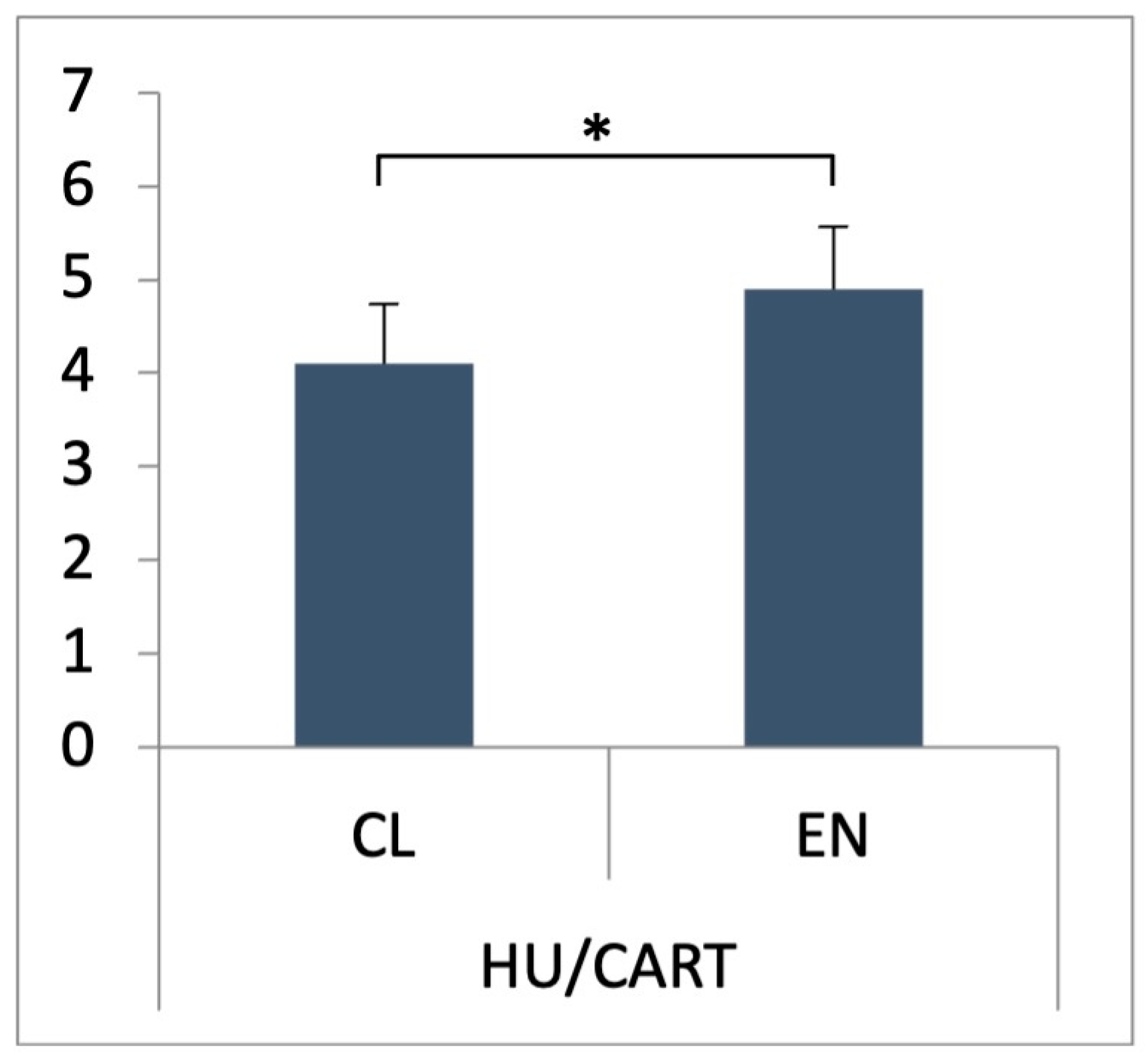
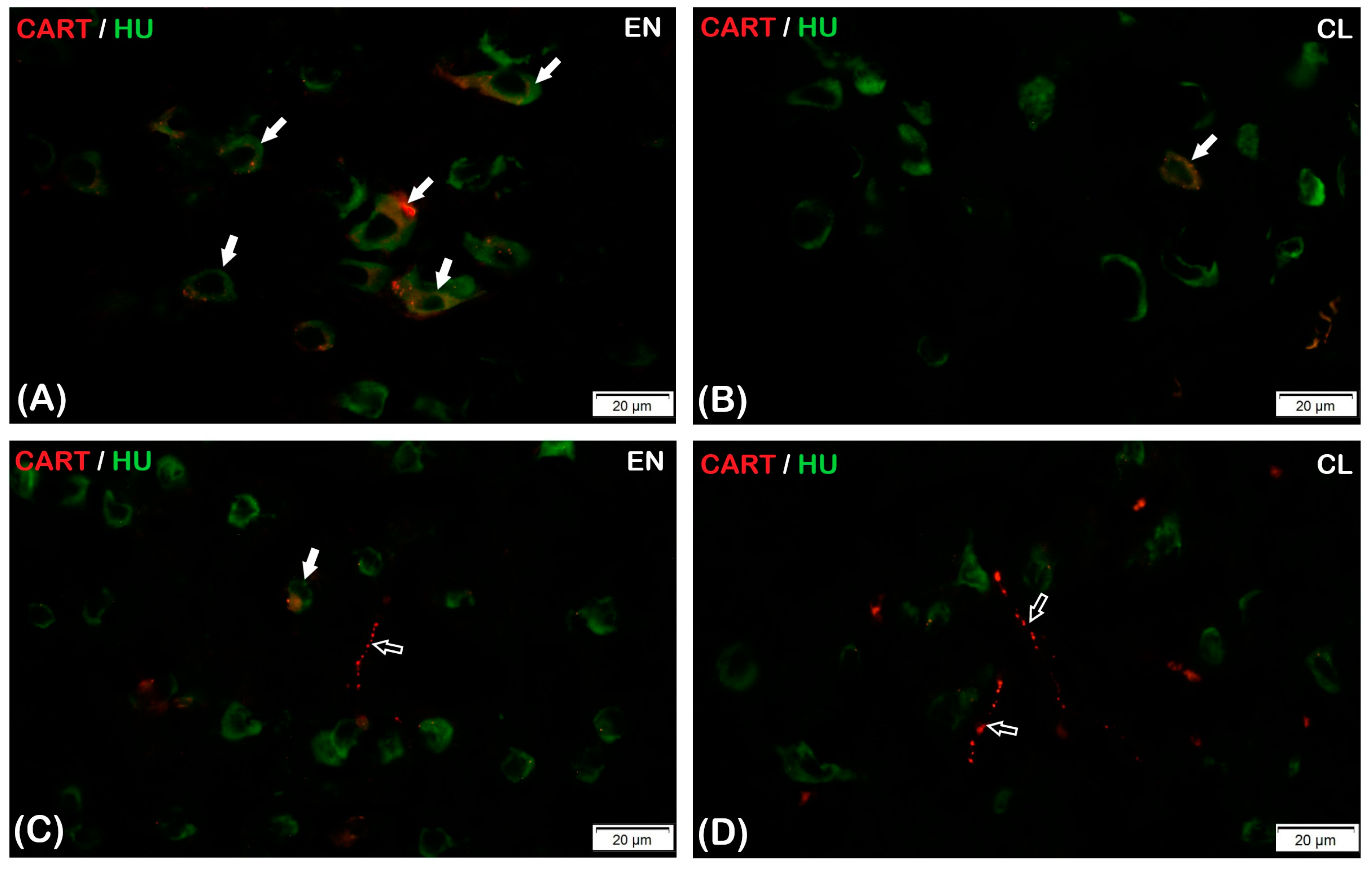
3.1. CART Expression Pattern
3.2. Co-Localization Studies
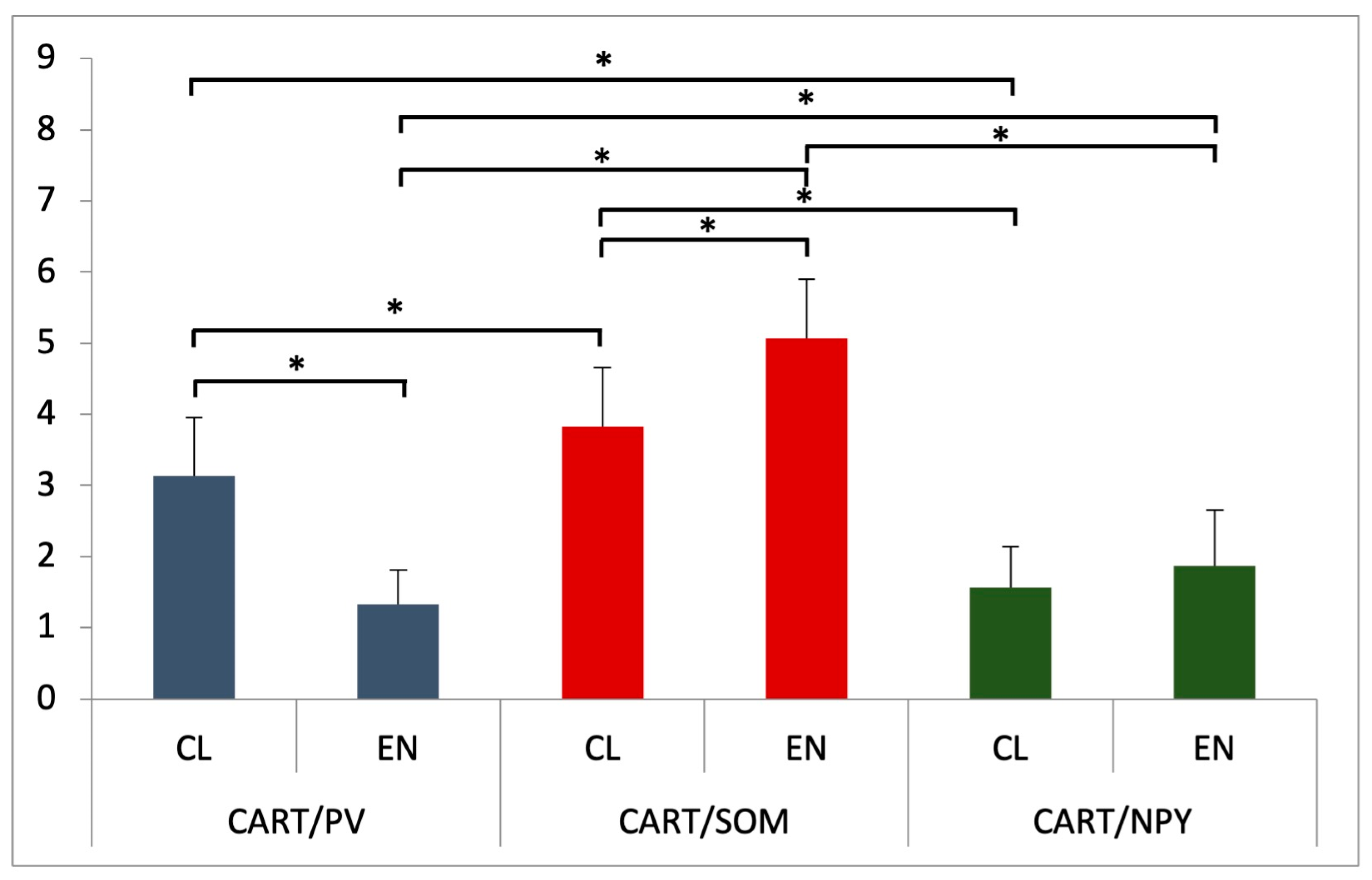
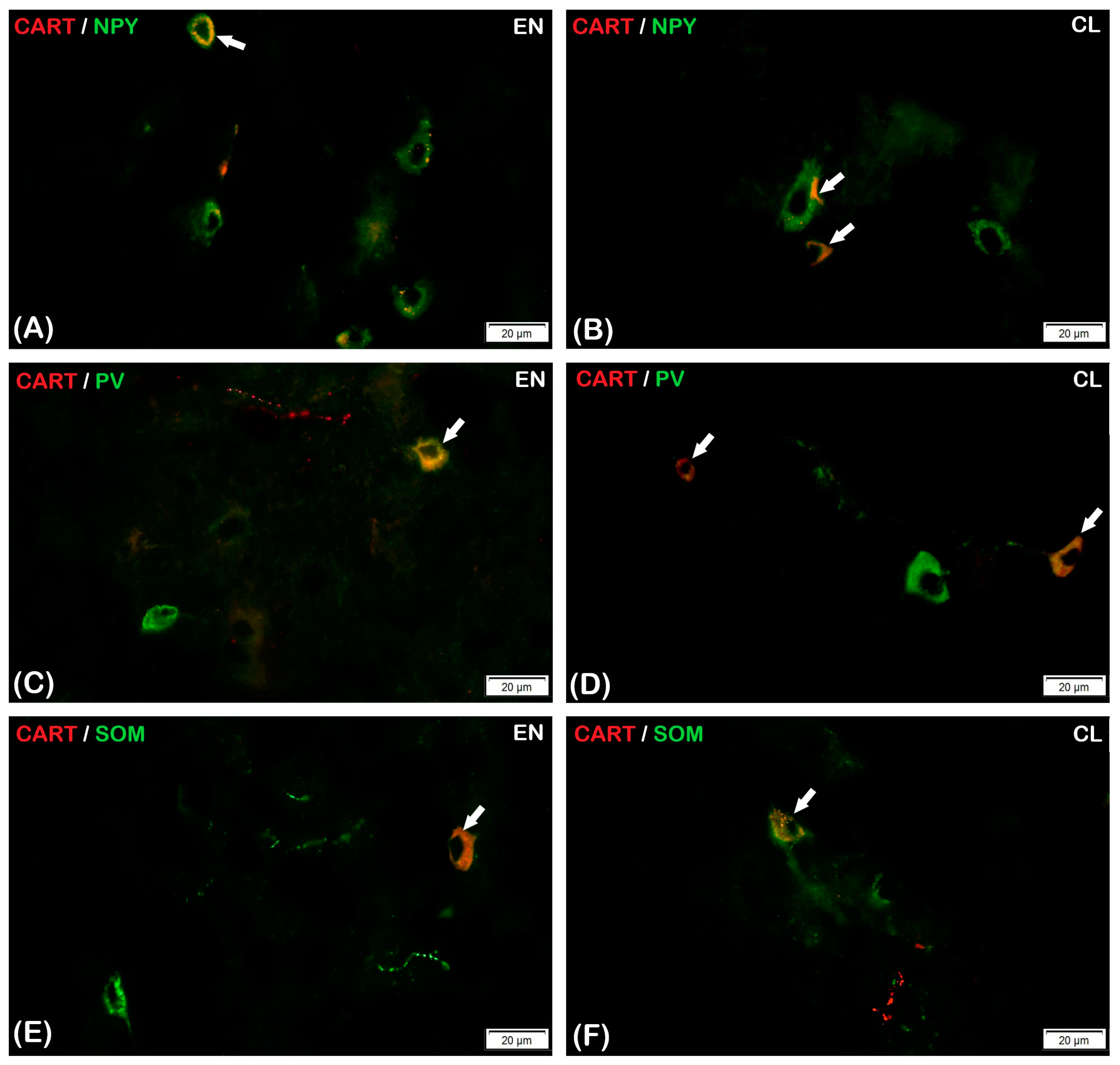
3.2.1. Parvalbumin
3.2.2. Somatostatin
3.2.3. Neuropeptide Y
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Douglass, J.; McKinzie, A.A.; Couceyro, P. PCR Differential Display Identifies a Rat Brain MRNA That Is Transcriptionally Regulated by Cocaine and Amphetamine. J. Neurosci. Off. J. Soc. Neurosci. 1995, 15 Pt 2, 2471–2481. [Google Scholar] [CrossRef] [Green Version]
- Równiak, M.; Robak, A.; Bogus-Nowakowska, K.; Wasilewska, B.; Najdzion, J.; Majewski, M. The Cocaine- and Amphetamine-Regulated Transcript (CART) Immunoreactivity in the Amygdala of the Pig: CART in the Pig Amygdala. Anat. Histol. Embryol. 2010, 39, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, B.C.; Cavalcante, J.C.; Elias, C.F. Expression of Cocaine- and Amphetamine-Regulated Transcript in the Rat Forebrain during Postnatal Development. Neuroscience 2011, 195, 201–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.H.; Cho, Y.H.; Kim, H.Y.; Cha, S.H.; Ryu, H.; Jang, W.; Shin, K.H. Increase in Cocaine- and Amphetamine-Regulated Transcript (CART) in Specific Areas of the Mouse Brain by Acute Caffeine Administration. Neuropeptides 2015, 50, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Szalak, R.; Matysek, M.; Kukula-Koch, W.; Rycerz, K.; Zacharko-Siembida, A.; Arciszewski, M.B. Colocalization Pattern of Cocaine- and Amphetamine-Regulated Transcript Peptide and Parvalbumin Immunoreactivity in the Hippocampus Proper of the Chinchilla. Pol. J. Vet. Sci. 2020, 23, 89–96. [Google Scholar] [CrossRef]
- Shcherbina, L.; Edlund, A.; Esguerra, J.L.S.; Abels, M.; Zhou, Y.; Ottosson-Laakso, E.; Wollheim, C.B.; Hansson, O.; Eliasson, L.; Wierup, N. Endogenous Beta-Cell CART Regulates Insulin Secretion and Transcription of Beta-Cell Genes. Mol. Cell. Endocrinol. 2017, 447, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Mozel, S.; Matysek, M.; Zacharko-Siembida, A.; Szalak, R.; Krzysiak, M.; Arciszewski, M.B. Expression of Cocaine- and Amphetamine-Regulated Transcript in the Pancreas of the European Bison. Med. Weter. 2019, 75, 669–672. [Google Scholar] [CrossRef]
- Czajkowska, M.; Całka, J. Neurochemistry of Enteric Neurons Following Prolonged Indomethacin Administration in the Porcine Duodenum. Front. Pharmacol. 2020, 11, 564457. [Google Scholar] [CrossRef]
- Kay, J.N.; De La Huerta, I.; Kim, I.-J.; Zhang, Y.; Yamagata, M.; Chu, M.W.; Meister, M.; Sanes, J.R. Retinal Ganglion Cells with Distinct Directional Preferences Differ in Molecular Identity, Structure, and Central Projections. J. Neurosci. 2011, 31, 7753–7762. [Google Scholar] [CrossRef] [Green Version]
- Upadhya, M.A.; Nakhate, K.T.; Kokare, D.M.; Singru, P.S.; Subhedar, N.K. Cocaine- and Amphetamine-Regulated Transcript Peptide Increases Spatial Learning and Memory in Rats. Life Sci. 2011, 88, 322–334. [Google Scholar] [CrossRef]
- Lau, J.; Herzog, H. CART in the Regulation of Appetite and Energy Homeostasis. Front. Neurosci. 2014, 8, 313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, K.; Jin, J.; Zhu, X.; Yu, L.; Wang, S.; Qian, L.; Han, L.; Xu, Y. CART Modulates Beta-Amyloid Metabolism-Associated Enzymes and Attenuates Memory Deficits in APP/PS1 Mice. Neurol. Res. 2017, 39, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.C.; Hand, L.J.; Letherby, B.; Huckstep, K.L.; Campbell, E.J.; Lawrence, A.J. Cocaine and Amphetamine Regulated Transcript (CART) Signalling in the Central Nucleus of the Amygdala Modulates Stress-Induced Alcohol Seeking. Neuropsychopharmacology 2021, 46, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Schwaller, B. The Continuing Disappearance of “Pure” Ca2+ Buffers. Cell. Mol. Life Sci. 2009, 66, 275–300. [Google Scholar] [CrossRef] [PubMed]
- Malva, J.O.; Xapelli, S.; Baptista, S.; Valero, J.; Agasse, F.; Ferreira, R.; Silva, A.P. Multifaces of Neuropeptide Y in the Brain–Neuroprotection, Neurogenesis and Neuroinflammation. Neuropeptides 2012, 46, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Neves, J.; Pereira de Almeida, L.; Cavadas, C. Neuropeptide Y (NPY) as a Therapeutic Target for Neurodegenerative Diseases. Neurobiol. Dis. 2016, 95, 210–224. [Google Scholar] [CrossRef]
- Liguz-Lecznar, M.; Urban-Ciecko, J.; Kossut, M. Somatostatin and Somatostatin-Containing Neurons in Shaping Neuronal Activity and Plasticity. Front. Neural Circuits 2016, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Nocera, S.; Simon, A.; Fiquet, O.; Chen, Y.; Gascuel, J.; Datiche, F.; Schneider, N.; Epelbaum, J.; Viollet, C. Somatostatin Serves a Modulatory Role in the Mouse Olfactory Bulb: Neuroanatomical and Behavioral Evidence. Front. Behav. Neurosci. 2019, 13, 61. [Google Scholar] [CrossRef] [Green Version]
- Saiz-Sanchez, D.; Ubeda-Bañon, I.; Flores-Cuadrado, A.; Gonzalez-Rodriguez, M.; Villar-Conde, S.; Astillero-Lopez, V.; Martinez-Marcos, A. Somatostatin, Olfaction, and Neurodegeneration. Front. Neurosci. 2020, 14, 96. [Google Scholar] [CrossRef] [Green Version]
- Kowiański, P.; Moryś, J.M.; Dziewiątkowski, J.; Wójcik, S.; Sidor-Kaczmarek, J.; Moryś, J. NPY-, SOM- and VIP-Containing Interneurons in Postnatal Development of the Rat Claustrum. Brain Res. Bull. 2008, 76, 565–571. [Google Scholar] [CrossRef]
- Kowiański, P.; Dziewiątkowski, J.; Moryś, J.M.; Majak, K.; Wójcik, S.; Edelstein, L.R.; Lietzau, G.; Moryś, J. Colocalization of Neuropeptides with Calcium-Binding Proteins in the Claustral Interneurons during Postnatal Development of the Rat. Brain Res. Bull. 2009, 80, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Pirone, A.; Castagna, M.; Granato, A.; Peruffo, A.; Quilici, F.; Cavicchioli, L.; Piano, I.; Lenzi, C.; Cozzi, B. Expression of Calcium-Binding Proteins and Selected Neuropeptides in the Human, Chimpanzee, and Crab-Eating Macaque Claustrum. Front. Syst. Neurosci. 2014, 8, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szalak, R.; Matysek, M.; Krawczyk, A.; Olejarska, A.; Lalak, R.; Arciszewski, M.B. Immunoreactivity to the Parvalbumin and Calbindin D28k in the Claustrum and Endopiriform Nucleus of the Chinchilla. Med. Weter. 2015, 71, 615–618. [Google Scholar]
- Druga, R.; Salaj, M.; Edelstein, L. Calretinin-Immunoreactive Neurons in the Claustrum of the Guinea Pig. Claustrum 2017, 2, 1273650. [Google Scholar] [CrossRef] [Green Version]
- Corder, K.M.; Li, Q.; Cortes, M.A.; Bartley, A.F.; Davis, T.R.; Dobrunz, L.E. Overexpression of Neuropeptide Y Decreases Responsiveness to Neuropeptide Y. Neuropeptides 2020, 79, 101979. [Google Scholar] [CrossRef]
- Torgerson, C.M.; Irimia, A.; Goh, S.Y.M.; Van Horn, J.D. The DTI Connectivity of the Human Claustrum: Claustrum Connectivity. Hum. Brain Mapp. 2015, 36, 827–838. [Google Scholar] [CrossRef]
- Smith, J.B.; Alloway, K.D. Functional Specificity of Claustrum Connections in the Rat: Interhemispheric Communication between Specific Parts of Motor Cortex. J. Neurosci. 2010, 30, 16832–16844. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.B.; Liang, Z.; Watson, G.D.R.; Alloway, K.D.; Zhang, N. Interhemispheric Resting-State Functional Connectivity of the Claustrum in the Awake and Anesthetized States. Brain Struct. Funct. 2017, 222, 2041–2058. [Google Scholar] [CrossRef] [PubMed]
- Remedios, R.; Logothetis, N.K.; Kayser, C. Unimodal Responses Prevail within the Multisensory Claustrum. J. Neurosci. 2010, 30, 12902–12907. [Google Scholar] [CrossRef] [Green Version]
- Baizer, J.S.; Sherwood, C.C.; Noonan, M.; Hof, P.R. Comparative Organization of the Claustrum: What Does Structure Tell Us about Function? Front. Syst. Neurosci. 2014, 8, 117. [Google Scholar] [CrossRef] [Green Version]
- Druga, R. The Structure and Connections of the Claustrum. In The Claustrum; Elsevier: Amsterdam, The Netherlands, 2014; pp. 29–84. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Smith, J.B.; Alloway, K.D.; Hof, P.R.; Orman, R.; Reser, D.H.; Watakabe, A.; Watson, G.D.R. The Relationship between the Claustrum and Endopiriform Nucleus: A Perspective towards Consensus on Cross-Species Homology. J. Comp. Neurol. 2019, 527, 476–499. [Google Scholar] [CrossRef] [PubMed]
- Suckow, M.A.; Stevens, K.A.; Wilson, R.P. The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar] [CrossRef]
- Kuchinka, J. Morphometry and Variability of the Brain Arterial Circle in Chinchilla (Chinchilla laniger, Molina). Anat. Rec. 2017, 300, 1472–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koylu, E.O.; Couceyro, P.R.; Lambert, P.D.; Kuhar, M.J. Cocaine- and Amphetamine-Regulated Transcript Peptide Immunohistochemical Localization in the Rat Brain. J. Comp. Neurol. 1998, 391, 115–132. [Google Scholar] [CrossRef]
- Abraham, H.; Covasa, M.; Hajnal, A. Cocaine- and Amphetamine-Regulated Transcript Peptide Immunoreactivity in the Brain of the CCK-1 Receptor Deficient Obese OLETF Rat. Exp. Brain Res. 2009, 196, 545–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najdzion, J.; Wasilewska, B.; Bogus-Nowakowska, K.; Robak, A. The Cocaine- and Amphetamine-Regulated Transcript, Calbindin, Calretinin and Parvalbumin Immunoreactivity in the Medial Geniculate Body of the Guinea Pig. J. Chem. Neuroanat. 2014, 59–60, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Wasilewska, B.; Najdzion, J.; Równiak, M.; Bogus-Nowakowska, K.; Hermanowicz, B.; Kolenkiewicz, M.; Żakowski, W.; Robak, A. Cocaine- and Amphetamine-Regulated Transcript and Calcium Binding Proteins Immunoreactivity in the Subicular Complex of the Guinea Pig. Ann. Anat.-Anat. Anz. 2016, 204, 51–62. [Google Scholar] [CrossRef]
- Lambert, P.D.; Couceyro, P.R.; McGirr, K.M.; Dall Vechia, S.E.; Smith, Y.; Kuhar, M.J. CART Peptides in the Central Control of Feeding and Interactions with Neuropeptide Y. Synapse 1998, 29, 293–298. [Google Scholar] [CrossRef]
- Larsen, P.J.; Seier, V.; Fink-Jensen, A.; Holst, J.J.; Warberg, J.; Vrang, N. Cocaine- and Amphetamine-Regulated Transcript Is Present in Hypothalamic Neuroendocrine Neurones and Is Released to the Hypothalamic-Pituitary Portal Circuit: CART as a Hypothalamic-Pituitary Neurotransmitter. J. Neuroendocrinol. 2003, 15, 219–226. [Google Scholar] [CrossRef]
- Dall Vechia, S.; Lambert, P.D.; Couceyro, P.C.; Kuhar, M.J.; Smith, Y. CART Peptide Immunoreactivity in the Hypothalamus and Pituitary in Monkeys: Analysis of Ultrastructural Features and Synaptic Connections in the Paraventricular Nucleus. J. Comp. Neurol. 2000, 416, 291–308. [Google Scholar] [CrossRef]
- Menyhért, J.; Wittmann, G.; Lechan, R.M.; Keller, É.; Liposits, Z.; Fekete, C. Cocaine- and Amphetamine-Regulated Transcript (CART) Is Colocalized with the Orexigenic Neuropeptide Y and Agouti-Related Protein and Absent from the Anorexigenic α-Melanocyte-Stimulating Hormone Neurons in the Infundibular Nucleus of the Human Hypothalamus. Endocrinology 2007, 148, 4276–4281. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PRIMARY ANTIBODIES | ||||
|---|---|---|---|---|
| Antigen | Code | Host Species | Dilution | Supplier |
| HU C/D | A-21271 | mouse | 1:400 | Thermo Fisher Scientific, Waltham, MA, USA |
| CART | H-003-61 | rabbit | 1:10,000 | Phoenix Pharmaceuticals, Burlingame, CA, USA |
| PV | PV 235 | mouse | 1:2000 | Swant AG, Burgdorf, Switzer-land |
| SOM | 8330-0009 | rat | 1:100 | Bio-rad, Hercules, CA, USA |
| NPY | PA1-27980 | guinea pig | 1:700 | Thermo Fisher Scientific, Waltham, MA, USA |
| SECONDARY ANTIBODIES | ||||
| Antigen | Code | Host Species | Dilution | Supplier |
| Alexa Fluor 488, goat anti-mouse IgG | A-11029 | mouse | 1:800 | Thermo Fisher Scientific, Waltham, MA, USA |
| Alexa Fluor 594, donkey anti-rabbit IgG | A-21207 | rabbit | 1:800 | Thermo Fisher Scientific, Waltham, MA, USA |
| Alexa Fluor 488, goat anti-rat IgG | A-11006 | rat | 1:800 | Thermo Fisher Scientific, Waltham, MA, USA |
| Alexa Fluor 488, goat anti-guinea pig IgG | A-11073 | guinea pig | 1:800 | Thermo Fisher Scientific, Waltham, MA, USA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szalak, R.; Matysek, M.; Mozel, S.; Arciszewski, M.B. Cocaine- and Amphetamine-Regulated Transcript (CART) Peptide Is Co-Expressed with Parvalbumin, Neuropeptide Y and Somatostatin in the Claustrum of the Chinchilla. Animals 2023, 13, 2177. https://doi.org/10.3390/ani13132177
Szalak R, Matysek M, Mozel S, Arciszewski MB. Cocaine- and Amphetamine-Regulated Transcript (CART) Peptide Is Co-Expressed with Parvalbumin, Neuropeptide Y and Somatostatin in the Claustrum of the Chinchilla. Animals. 2023; 13(13):2177. https://doi.org/10.3390/ani13132177
Chicago/Turabian StyleSzalak, Radosław, Małgorzata Matysek, Sylwia Mozel, and Marcin B. Arciszewski. 2023. "Cocaine- and Amphetamine-Regulated Transcript (CART) Peptide Is Co-Expressed with Parvalbumin, Neuropeptide Y and Somatostatin in the Claustrum of the Chinchilla" Animals 13, no. 13: 2177. https://doi.org/10.3390/ani13132177