
Anthropogenic Neighborhood Impact on Bacterial and Fungal Communities in Polar Bear Feces
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
- The Franz Josef Land archipelago (3 individuals). The samples were collected at the beginning of the ice melting period (summer). The bears usually prey on adult seals and cubs in dens in between the islands in the southern part of the archipelago during this period.
- Novaya Zemlya archipelago, Severny Island, Cape Zhelaniya (8 individuals). The samples were collected during the ice-free period (summer–autumn), during which the bears were fasting and feeding opportunistically on the remains of a walrus washed ashore, grass, and kelp.
- Novaya Zemlya archipelago, Yuzhny Island, Belushya Bay, in the area of a large village of Belushya Guba (5 individuals). The samples were collected when the Barents Sea coast was free of landfast ice (autumn). The bears were feeding on human food waste landfill.
2.2. Microbiome Studies
3. Results
3.1. State Conditions of the Polar Bears
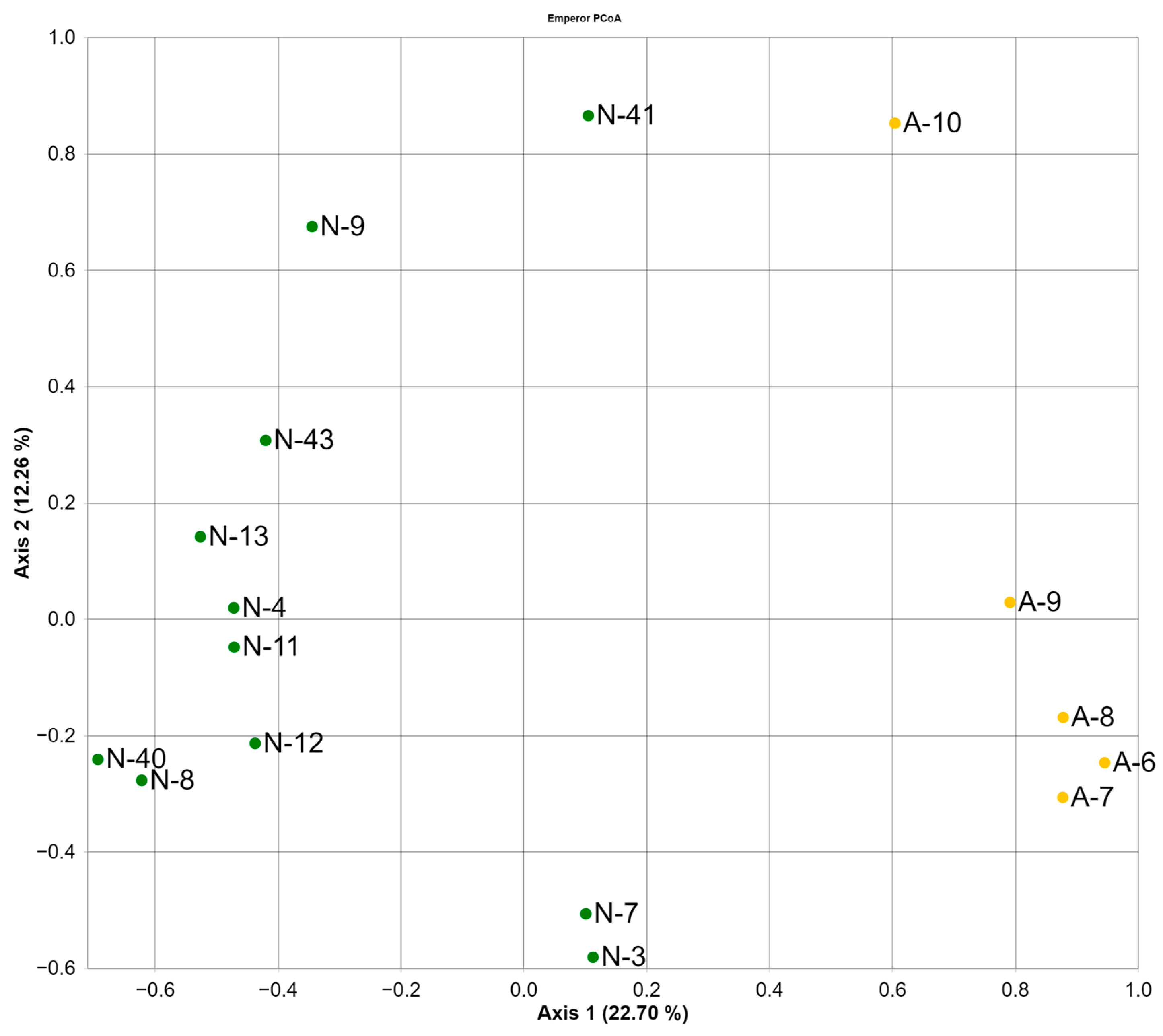
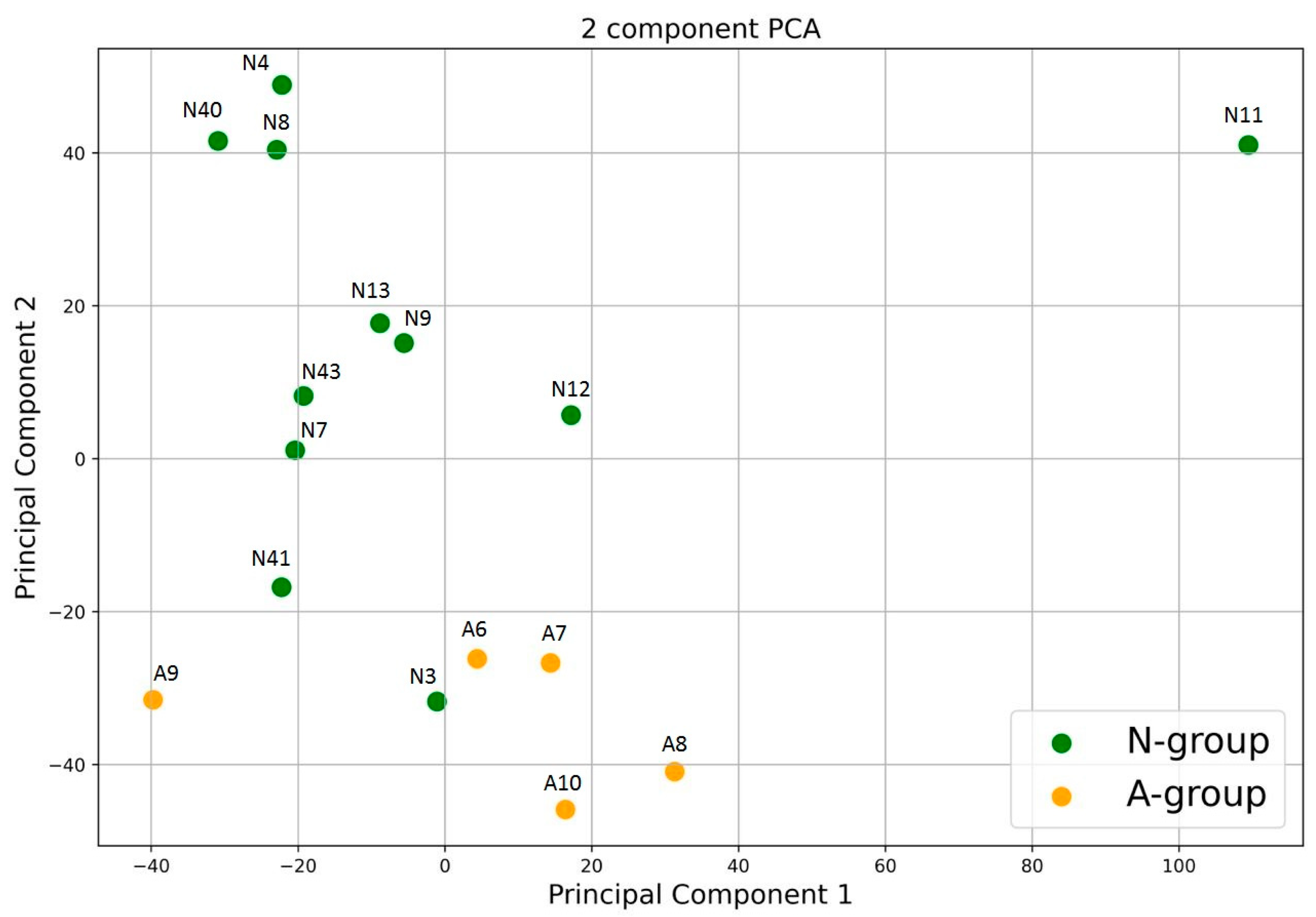
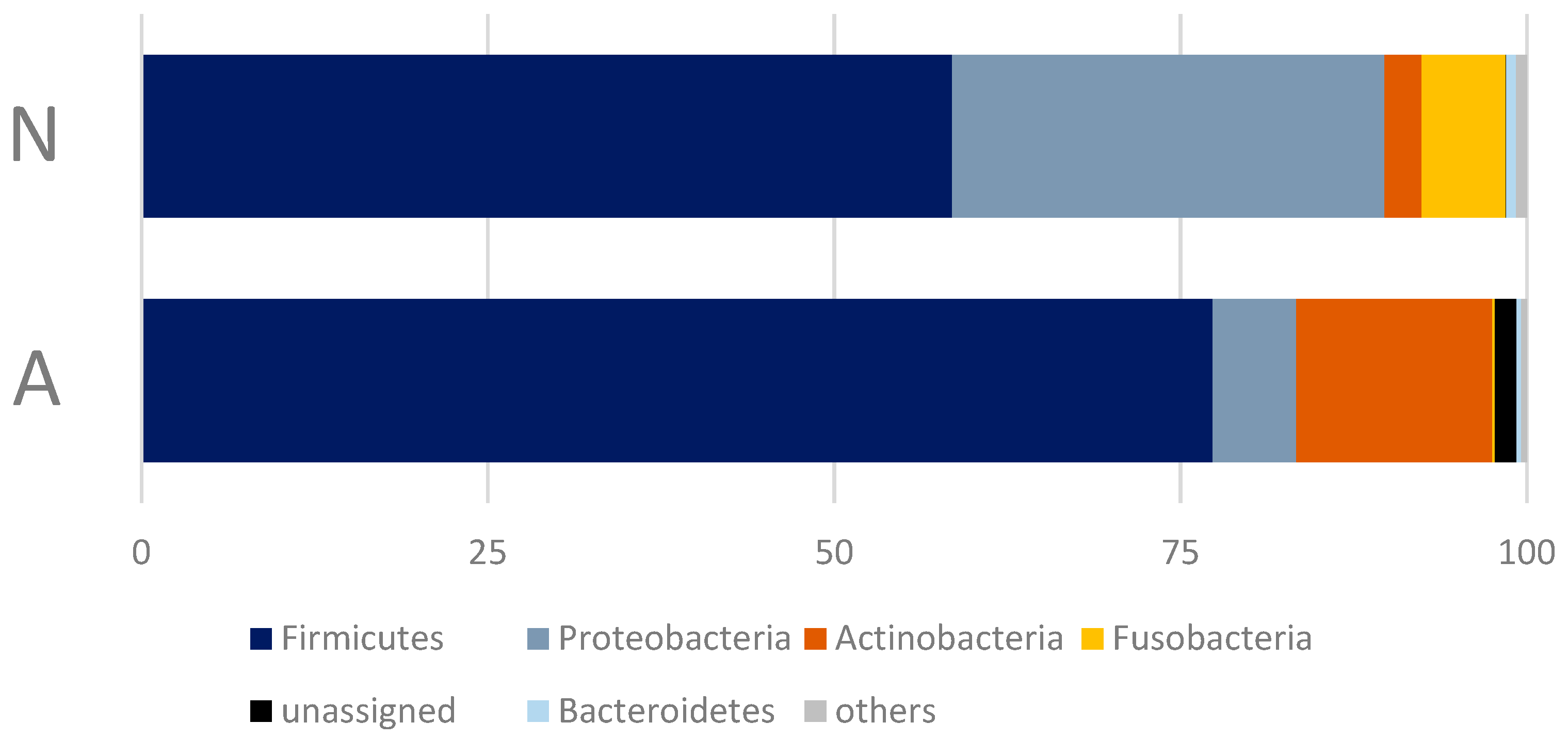
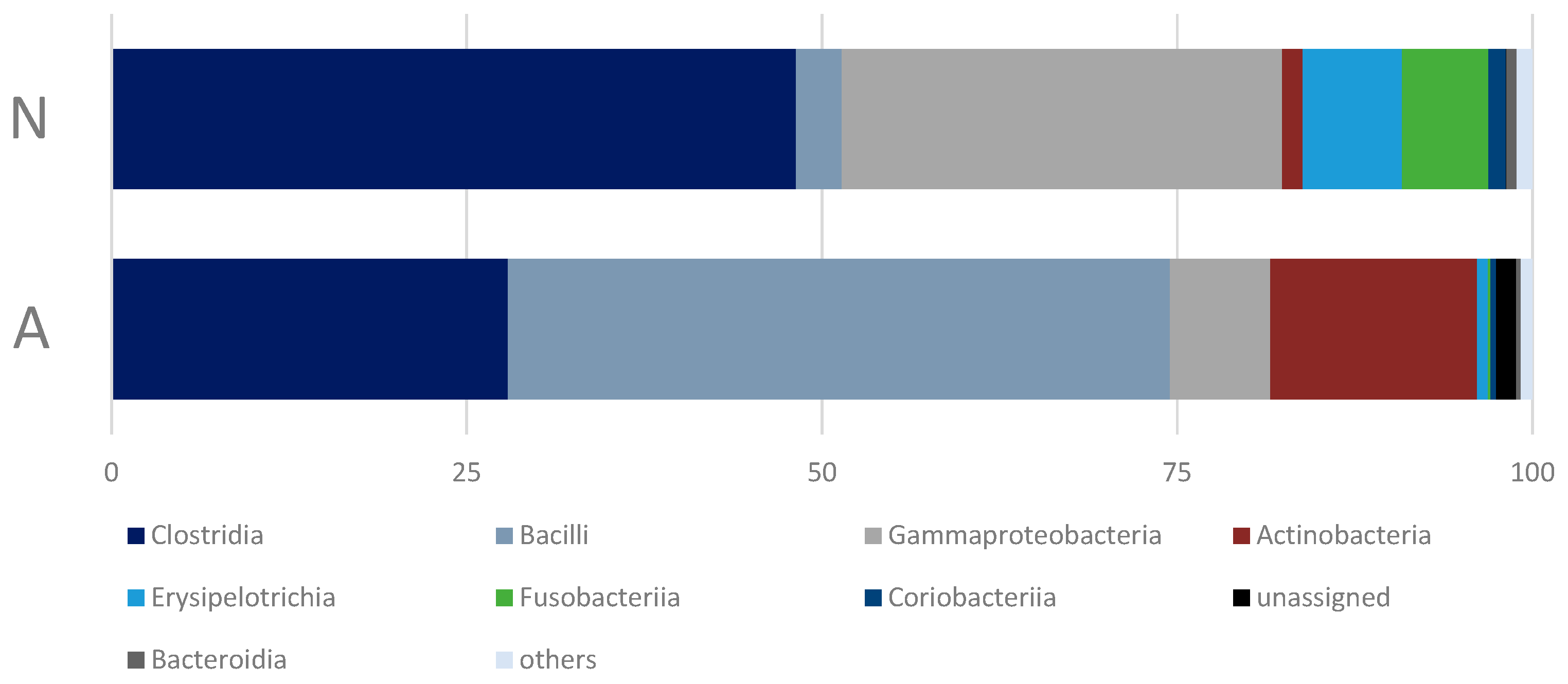
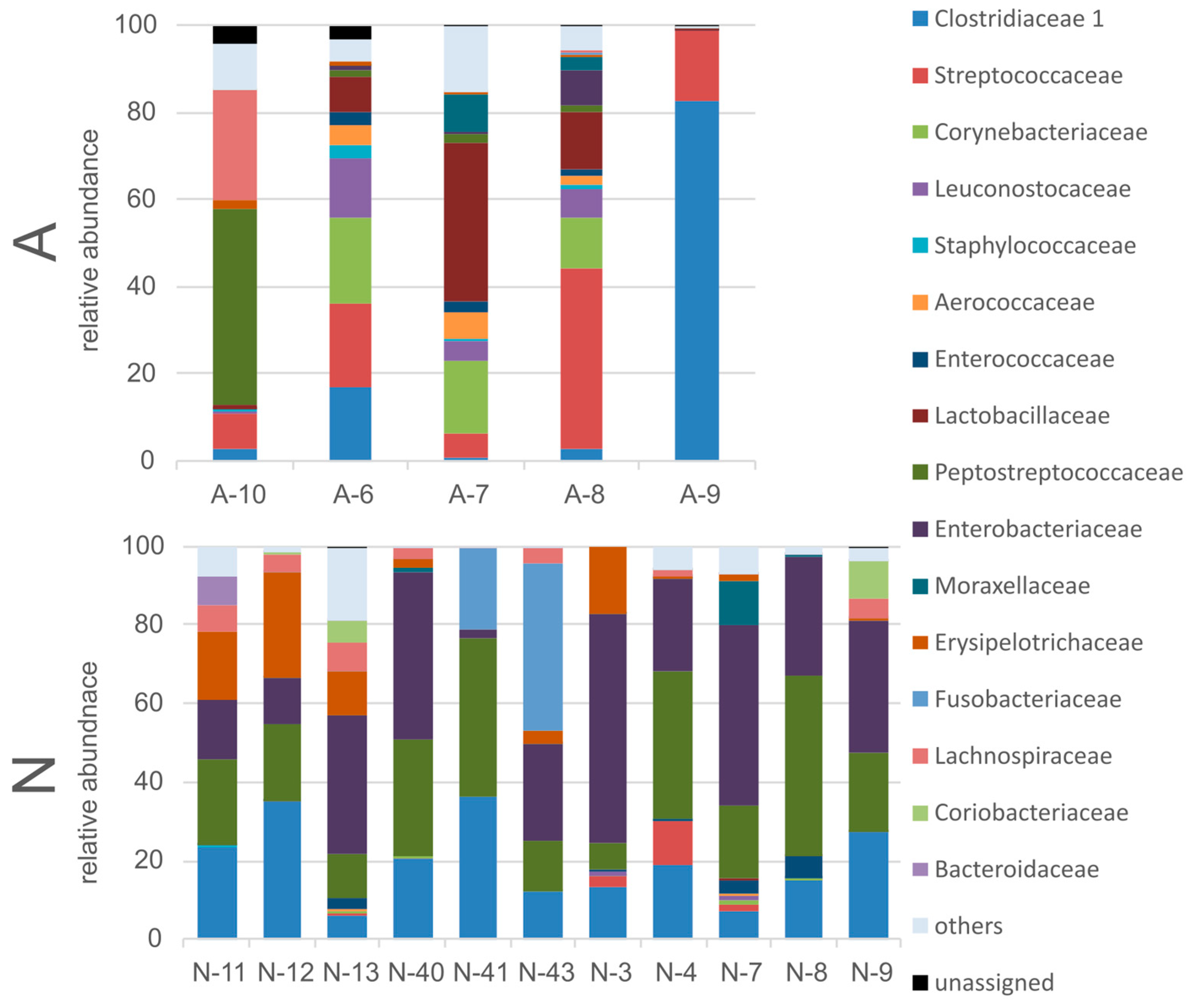
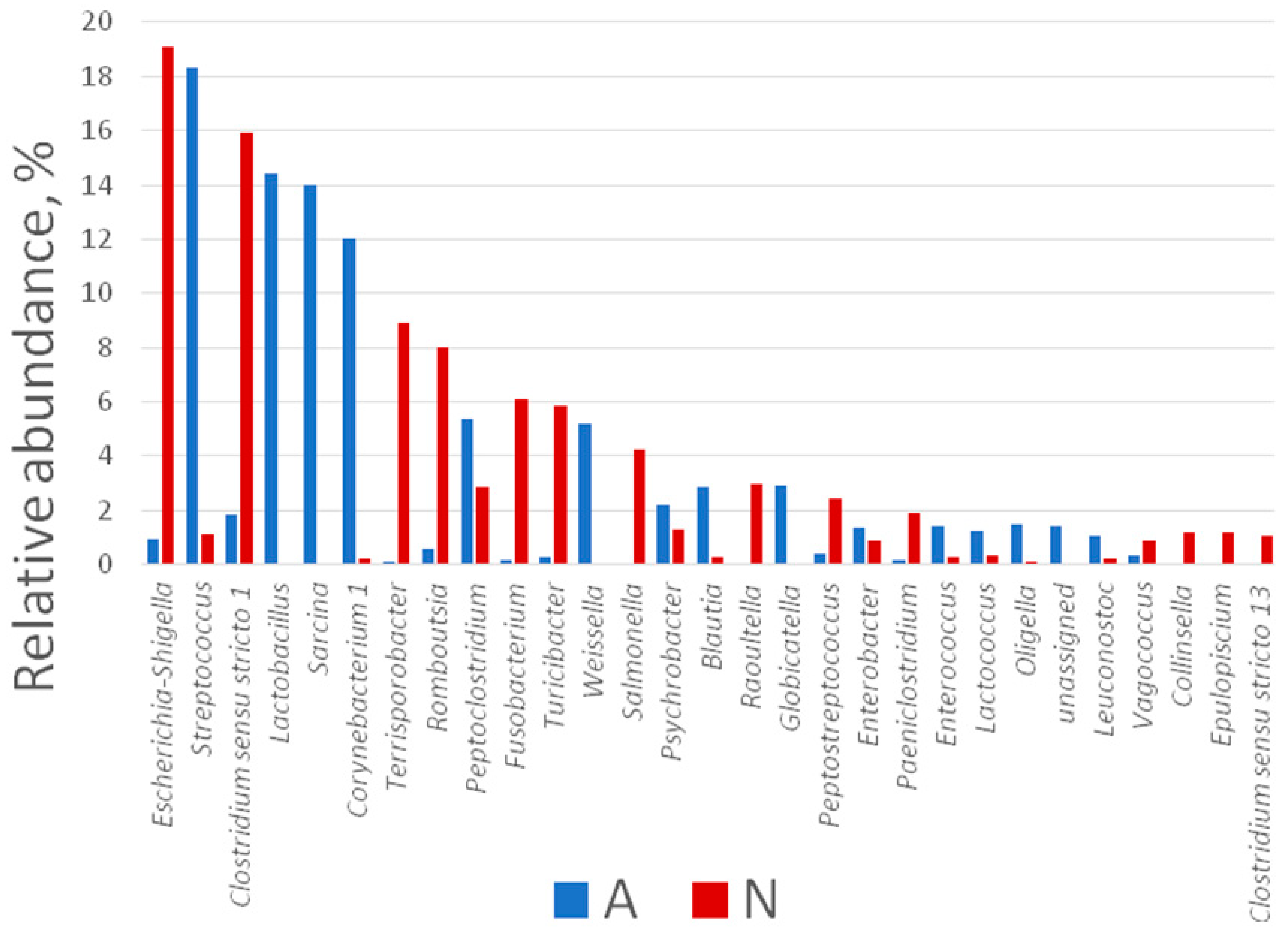
3.2. Gut Bacterial Community of the Polar Bears
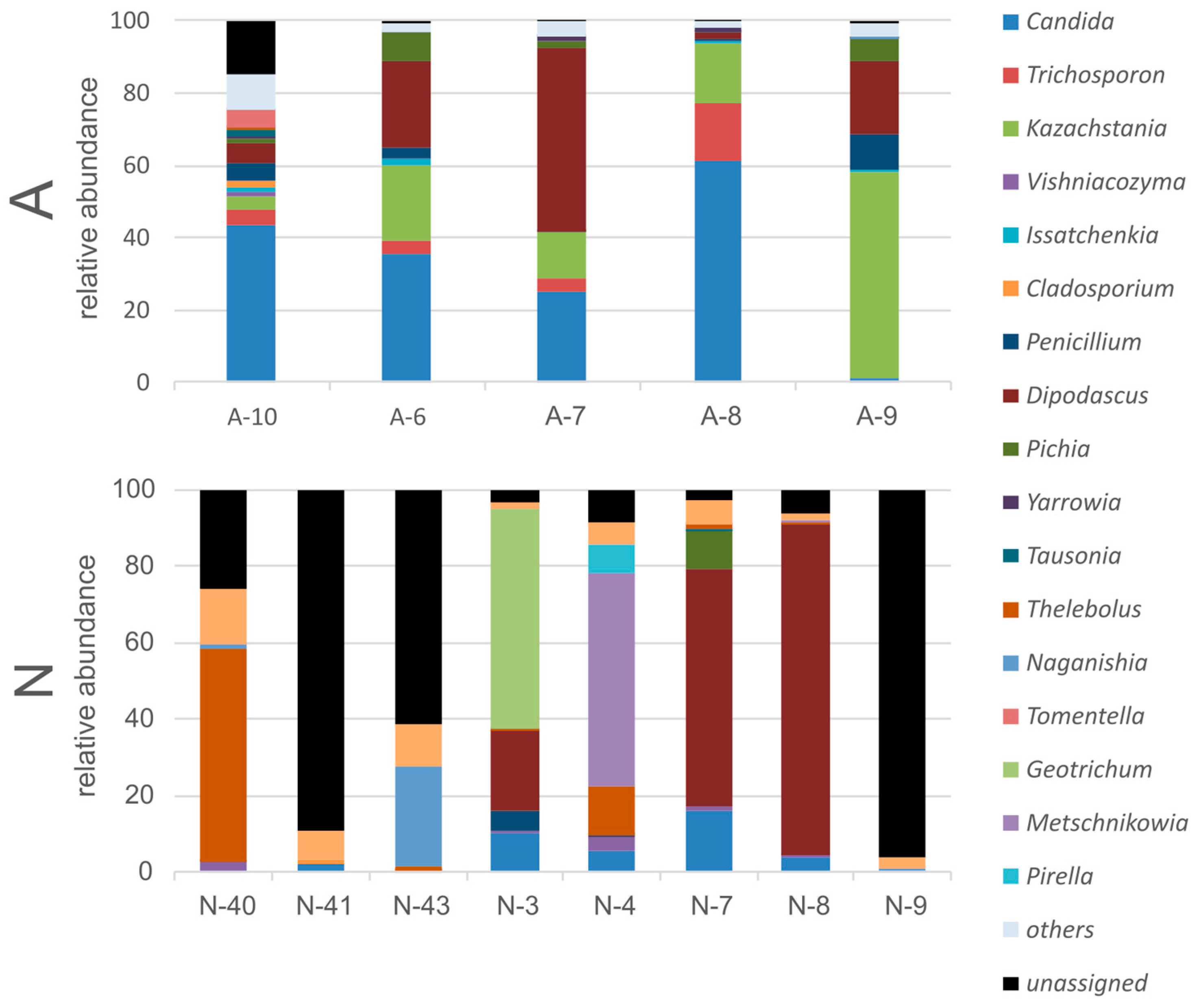
3.3. Gut Fungal Community of the Polar Bears
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Amstrup, S.C.; DeMaster, D. Polar bear, Ursus maritimus. In Wild Mammals of North America: Biology, Management, and Conservation, 2nd ed.; Feldman, G.A., Thompson, B.C., Chapman, J.A., Eds.; John Hopkins University Press: Baltimore, MD, USA, 2003; Volume 2, pp. 587–610. [Google Scholar]
- Stirling, I.; Lunn, N.J.; Iacozza, J. Long-term trends in the population ecology of polar bears in western Hudson Bay in relation to climatic change. Arctic 1999, 52, 294–306. [Google Scholar] [CrossRef]
- Derocher, A.E.; Lunn, N.J.; Stirling, I. Polar Bears in a Warming Climate. Integr. Comp. Biol. 2004, 44, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Regehr, E.V.; Hunter, C.M.; Caswell, H.; Amstrup, S.C.; Stirling, I. Survival and breeding of polar bears in the southern Beaufort Sea in relation to sea ice. J. Anim. Ecol. 2010, 79, 117–127. [Google Scholar] [CrossRef]
- Laidre, K.; Atkinson, S.; Regehr, E.; Stern, H.; Born, E.; Wiig, Ø.; Lunn, N.; Dyck, M. Interrelated ecological impacts of climate change on an apex predator. Ecol. Appl. 2020, 30, e02071. [Google Scholar] [CrossRef] [PubMed]
- Årthun, M.; Eldevik, T.; Smedsrud, L.H.; Skagseth, Ø.; Ingvaldsen, R.B. Quantifying the influence of Atlantic heat on Barents Sea ice variability and retreat. J. Clim. 2012, 25, 4736–4743. [Google Scholar] [CrossRef] [Green Version]
- Stern, H.; Laidre, K. Sea-ice indicators of polar bear habitat. Cryosphere 2016, 10, 2027–2041. [Google Scholar] [CrossRef] [Green Version]
- Schliebe, S.; Rode, K.; Gleason, J.; Wilder, J.; Proffitt, K.; Evans, T.; Miller, S. Effects of sea ice extent and food availability on spatial and temporal distribution of polar bears during the fall open-water period in the Southern Beaufort Sea. Polar Biol. 2008, 31, 999–1010. [Google Scholar] [CrossRef]
- Derocher, A.E.; Wiig, Ø.; Bangjord, G. Predation of Svalbard reindeer by polar bears. Polar Biol. 2000, 23, 675–678. [Google Scholar] [CrossRef]
- Gormezano, L.J.; Rockwell, R.F. What to eat now? Shifts in polar bear diet during the ice-free season in western Hudson Bay. Ecol. Evol. 2013, 3, 3509–3523. [Google Scholar] [CrossRef]
- Iversen, M.; Aars, J.; Haug, T.; Alsos, I.G.; Lydersen, C.; Bachmann, L.; Kovacs, K.M. The diet of polar bears (Ursus maritimus) from Svalbard, Norway, inferred from scat analysis. Polar Biol. 2013, 36, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Tartu, S.; Bourgeon, S.; Aars, J.; Andersen, M.; Ehrich, D.; Thiemann, G.W.; Welker, J.M.; Routti, H. Geographical area and life history traits influence diet in an arctic marine predator. PLoS ONE 2016, 11, e0155980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavan, J. Observation of polar bear (Ursus maritimus) feeding on Svalbard reindeer (Rangifer tarandus platyrhyncus)—Exceptional behaviour or upcoming trend? Czech Polar Rep. 2018, 8, 243–248. [Google Scholar] [CrossRef]
- Stempniewicz, L.; Kulaszewicz, I.; Aars, J. Yes, they can: Polar bears Ursus maritimus successfully hunt Svalbard reindeer Rangifer tarandus platyrhynchus. Polar Biol. 2021, 44, 2199–2206. [Google Scholar] [CrossRef]
- Ivanov, E.A.; Mizin, I.A.; Kirilov, A.G.; Platonov, N.G.; Mordvintsev, I.N.; Naidenko, S.V.; Rozhnov, V.V. Observations of intraspecific killing, cannibalism, and aggressive behavior among polar bears (Ursus maritimus) in the eastern Barents Sea and the Kara Sea. Polar Biol. 2020, 43, 2121–2127. [Google Scholar] [CrossRef]
- Franz, M.; Whyte, L.; Atwood, T.C.; Laidre, K.L.; Roy, D.; Watson, S.E.; Góngora, E.; McKinney, M.A. Distinct gut microbiomes in two polar bear subpopulations inhabiting different sea ice ecoregions. Sci. Rep. 2022, 12, 522. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.E.; Hauffe, H.C.; Bull, M.J.; Atwood, T.C.; McKinney, M.A.; Pindo, M.; Perkins, S.E. Global change-driven use of onshore habitat impacts polar bear faecal microbiota. ISME J. 2019, 13, 2916–2926. [Google Scholar] [CrossRef] [Green Version]
- Whiteman, J.P.; Harlow, H.J.; Durner, G.M.; Regehr, E.V.; Amstrup, S.C.; Ben-David, M. Heightened immune system function in polar bears using terrestrial habitats. Physiol. Biochem. Zool. 2019, 92, 1–11. [Google Scholar] [CrossRef]
- Perez, J.C.; Johnson, A.D. Regulatory circuits that enable proliferation of the fungus Candida albicans in a mammalian host. PLoS Pathog. 2013, 9, e1003780. [Google Scholar] [CrossRef] [Green Version]
- Fiers, W.D.; Gao, I.H.; Iliev, I.D. Gut mycobiota under scrutiny: Fungal symbionts or environmental transients? Curr. Opin. Microbiol. 2019, 50, 79–86. [Google Scholar] [CrossRef]
- Perez, J.C. Candida albicans dwelling in the mammalian gut. Curr. Opin. Microbiol. 2019, 52, 41–46. [Google Scholar] [CrossRef]
- Derocher, A.E.; Wiig, Ø. Postnatal growth in body length and mass of polar bears (Ursus maritimus) at Svalbard. J. Zool. 2006, 256, 343–349. [Google Scholar] [CrossRef]
- Stirling, I.; Thiemann, G.W.; Richardson, E. Quantitative support for a subjective fatness index for immobilized polar bears. J. Wild. Manag. 2008, 72, 568–574. [Google Scholar] [CrossRef]
- Cattet, M.R.L.; Caulkett, N.A.; Obbard, M.E.; Stenhouse, G.B. A body-condition index for ursids. Can. J. Zool. 2002, 80, 1156–1161. [Google Scholar] [CrossRef] [Green Version]
- Manning, T.H. Age Determination in the Polar Bear Ursus maritimus Phipps; Occasional Paper No. 5; Canadian Wildlife Service: Ottawa, ON, Canada, 1973; pp. 3–12. [Google Scholar]
- Callahan, B.; McMurdie, P.; Rosen, M.; Han, A.; Johnson, A.; Holmes, S. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [Green Version]
- Sanders, J.; Beichman, A.; Roman, J.; Scott, J.J.; Emerson, D.; McCarthy, J.J.; Girguis, P.R. Baleen whales host a unique gut microbiome with similarities to both carnivores and herbivores. Nat. Commun. 2015, 6, 8285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [Green Version]
- Hale, V.L.; Tan, C.L.; Niu, K.; Yang, Y.; Knight, R.; Zhang, Q.; Cui, D.; Amato, K.R. Diet versus phylogeny: A comparison of gut microbiota in captive colobine monkey species. Microb. Ecol. 2018, 75, 515–527. [Google Scholar] [CrossRef]
- Sugden, S.; Sanderson, D.; Ford, K.; Stein, L.Y.; St. Clair, C.C. An altered microbiome in urban coyotes mediates relationships between anthropogenic diet and poor health. Sci. Rep. 2020, 10, 22207. [Google Scholar] [CrossRef]
- Sehnal, L.; Brammer-Robbins, E.; Wormington, A.M.; Blaha, L.; Bisesi, J.; Larkin, I.; Martyniuk, C.J.; Simonin, M.; Adamovsky, O. Microbiome composition and function in aquatic vertebrates: Small organisms making big impacts on aquatic animal health. Front. Microbiol. 2021, 12, 567408. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.L.; McLaughlin, R.W.; Zheng, J.S.; Hao, Y.J.; Fan, F.; Tian, R.M.; Wang, D. Microbial communities in different regions of the gastrointestinal tract in East Asian finless porpoises (Neophocaena asiaeorientalis sunameri). Sci. Rep. 2018, 8, 14142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwab, C.; Cristescu, B.; Northrup, J.M.; Stenhouse, G.B.; Gänzle, M. Diet and environment shape fecal bacterial microbiota composition and enteric pathogen load of grizzly bears. PLoS ONE 2011, 6, e27905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herwig, R.P.; Staley, J.T.; Nerini, M.K.; Braham, H.W. Baleen whales: Preliminary evidence for forestomach microbial fermentation. Appl. Environ. Microbiol. 1984, 47, 421–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecherskii, M.V.; Kuznetsova, T.A.; Naumova, E.I.; Kostina, N.V.; Gorlenko, M.V.; Golichenkov, M.V.; Umarov, M.M. Role of Microbiocenosis of the Gastrointestinal Tract in the Nutrition of Grouse. Biol. Bull. 2014, 41, 251–254. [Google Scholar] [CrossRef]
- Pollock, C.G.; Rohrbach, B.; Ramsay, E.C. Fungal dermatitis in captive pinnipeds. J. Zoo Wildl. Med. 2000, 31, 374–378. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual Number | Location | Sex | Age Class | Weight, kg | Body Length, cm | Axillary Girth, cm | Subjective Fatness Index | Body Condition Index (BCI) |
|---|---|---|---|---|---|---|---|---|
| N-3 | NZ | F | subad | 123 | 166 | 100 | 4 | −0.56 |
| N-4 | NZ | M | cub | 41.5 | 115 | 70 | 3 | −0.38 |
| N-7 | NZ | F | subad | 132 | 180 | 98 | 2.5 | −1.38 |
| N-8 | NZ | F | subad | 95 | 162 | 87 | 3 | −1.41 |
| A-7 | NZ | F | ad | 235 | 194 | 132 | 4 | 0.23 |
| A-9 | NZ | F | ad | 270 * | 187 | 4 | ||
| A-6 | NZ | F | ad | 245 | 185 | 140 | 4 | 1.07 |
| A-10 | NZ | F | ad | 208 | 190 | 125 | 4 | −0.05 |
| A-8 | NZ | M | ad | 562 | 230 | 190 | 4 | 1.81 |
| N-9 | NZ | M | ad | 400 * | 242 | 4 | ||
| N-40 | FJL | F | subad | 191 | 181 | 118 | 3 | 0.24 |
| N-41 | FJL | F | ad | 299 | 197 | 138 | 4 | 1.11 |
| N-43 | FJL | M | subad | 199 | 180 | 122 | 3 | 0.51 |
| N-11 | NZ | M | subad | 235 | 192 | 125 | 3 | 0.36 |
| N-12 | NZ | M | ad | 247 | 210 | 127 | 2 | −0.67 |
| N-13 | NZ | F | ad | 147 | 180 | 147 | 2 | −0.88 |
| Alpha-Diversity Index | A-Bears | N-Bears | Mann–Whitney U Test, p-Levels |
|---|---|---|---|
| Chao1 | 92.30 ± 31.11 | 59.50 ± 37.35 | 0.078 |
| Shannon | 3.78 ± 1.52 | 4.24 ± 0.53 | 0.257 |
| Simpson | 0.78 ± 0.23 | 0.84 ± 0.05 | 0.486 |
| OTU | 95.00 ± 31.91 | 55.59 ± 28.19 | 0.497 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vecherskii, M.V.; Kuznetsova, T.A.; Khayrullin, D.R.; Stepankov, A.A.; Artemieva, S.M.; Chukmasov, P.V.; Ivanov, E.A.; Mizin, I.A.; Mordvintsev, I.N.; Platonov, N.G.; et al. Anthropogenic Neighborhood Impact on Bacterial and Fungal Communities in Polar Bear Feces. Animals 2023, 13, 2067. https://doi.org/10.3390/ani13132067
Vecherskii MV, Kuznetsova TA, Khayrullin DR, Stepankov AA, Artemieva SM, Chukmasov PV, Ivanov EA, Mizin IA, Mordvintsev IN, Platonov NG, et al. Anthropogenic Neighborhood Impact on Bacterial and Fungal Communities in Polar Bear Feces. Animals. 2023; 13(13):2067. https://doi.org/10.3390/ani13132067
Chicago/Turabian StyleVecherskii, Maksim V., Tatiana A. Kuznetsova, David R. Khayrullin, Aleksandr A. Stepankov, Svetlana M. Artemieva, Pavel V. Chukmasov, Evgeny A. Ivanov, Ivan A. Mizin, Ilya N. Mordvintsev, Nikita G. Platonov, and et al. 2023. "Anthropogenic Neighborhood Impact on Bacterial and Fungal Communities in Polar Bear Feces" Animals 13, no. 13: 2067. https://doi.org/10.3390/ani13132067