
Different Methods for Seminal Plasma Removal and Sperm Selection on the Quality and Fertility of Collared Peccary Sperm
 , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bioethics and Chemicals
2.2. Semen Collection
2.3. Experimental Design
2.3.1. Experiment I: Swim-Up
2.3.2. Experiment II: Percoll® Gradient
2.3.3. Experiment III: Heterologous IVF
2.4. Sperm Evaluations
2.4.1. Motility Patterns
2.4.2. Morphology and Functional Integrity of the Membrane
2.4.3. Viability and Mitochondrial Activity
2.4.4. Integrity of the DNA
2.4.5. Oxidative Status
2.5. Heterologous IVF
2.5.1. Oocyte Collection and In Vitro Maturation
2.5.2. Co-Incubation and Embryo Development
2.6. Statistical Analysis
3. Results
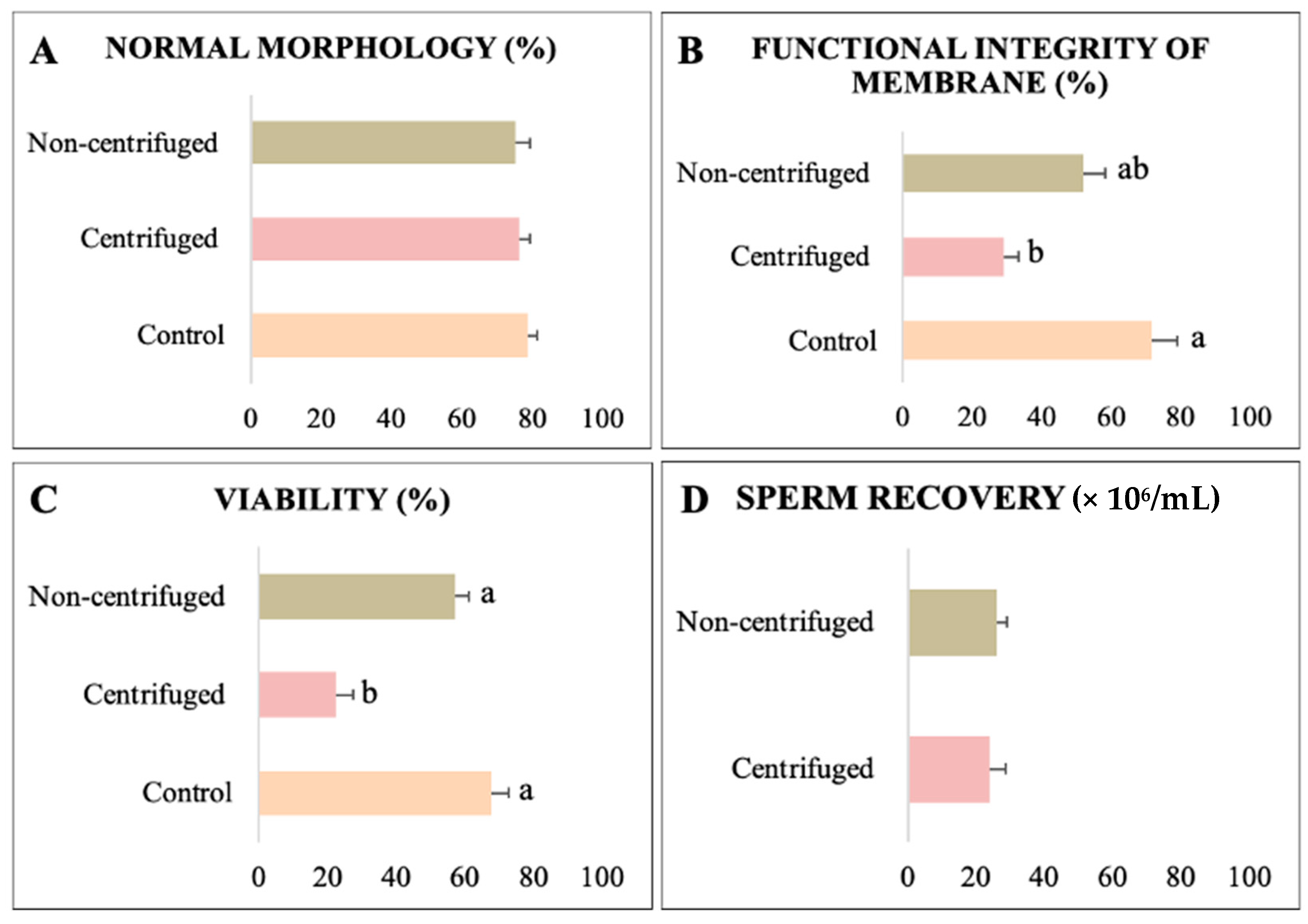
3.1. Experiment I: Swim-Up
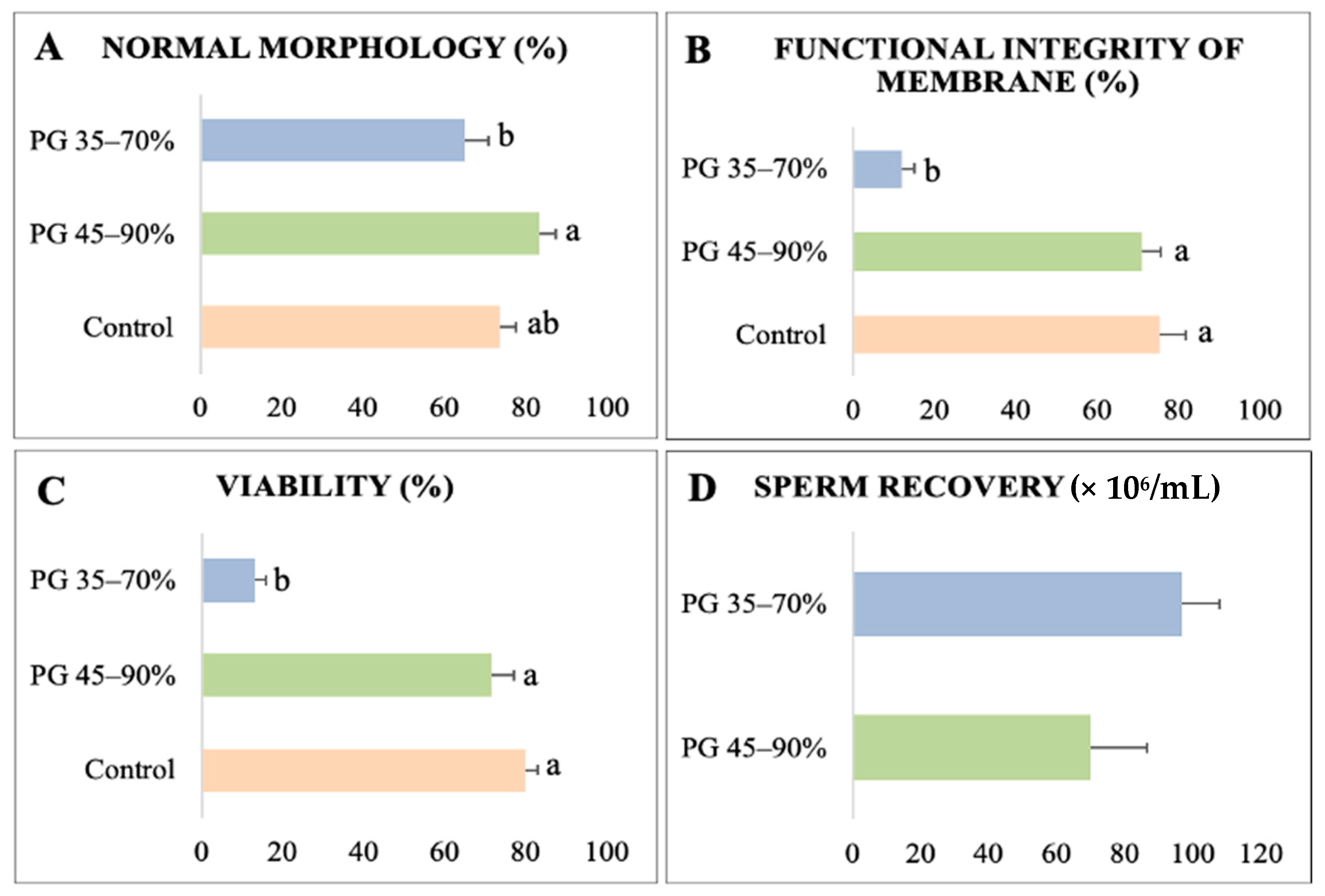
3.2. Experiment II: Percoll® Gradient
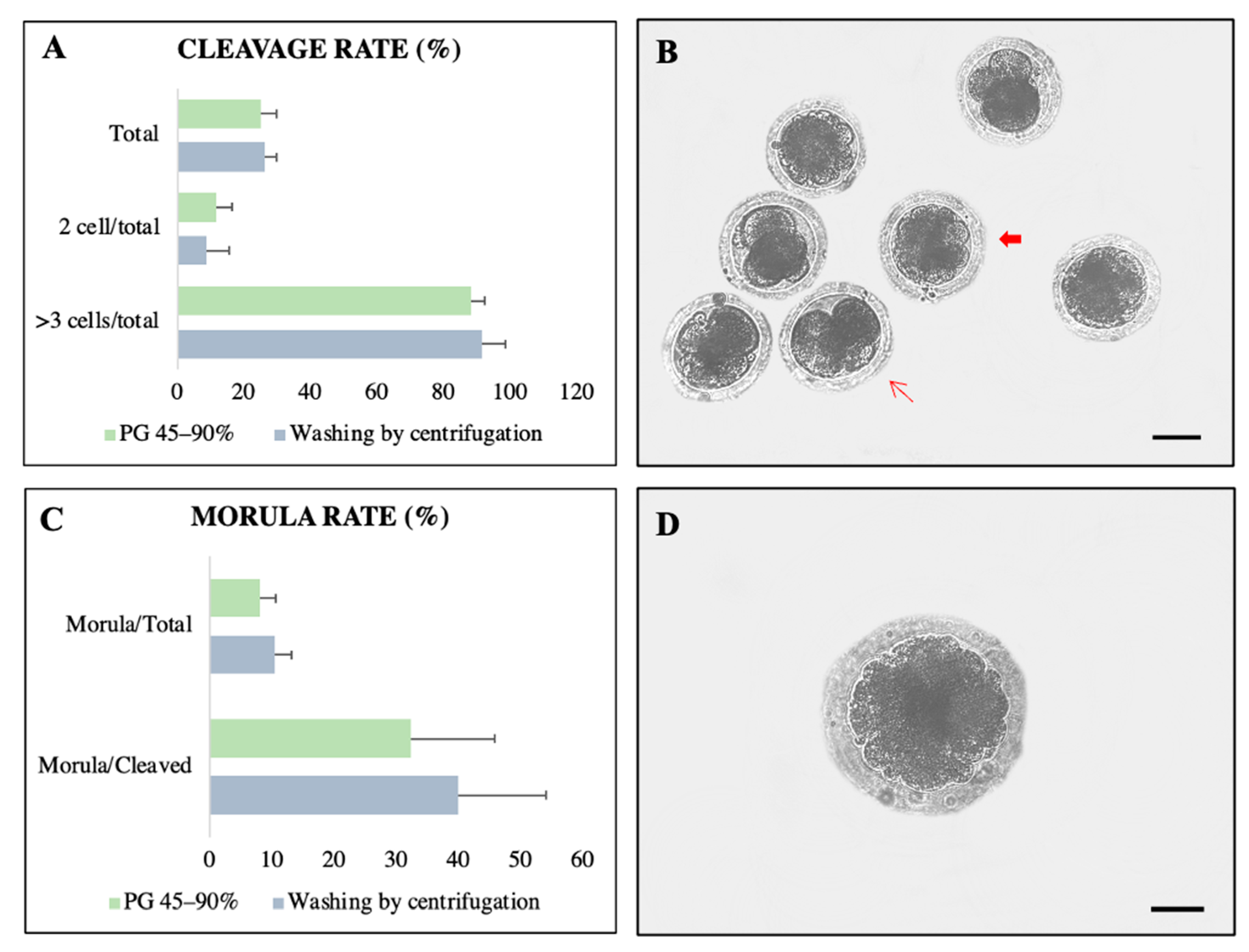
3.3. Experiment III: Heterologous IVF
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gongora, J.; Reyna-Hurtado, R.; Beck, H.; Taber, A.; Altrichter, M.; Keuroghlian, A. Pecari tajacu. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2011. [Google Scholar] [CrossRef]
- Desbiez, A.L.J.; Keuroghlian, A.; Beisiegel, B.M.; Medici, E.P.; Gatti, A.; Pontes, A.R.M.; Campos, C.B.; Tófoli, C.F.; Júnior, E.A.M.; Azevedo, F.C.; et al. Avaliação do risco de extinção do cateto Pecari tajacu Linnaeus, 1758, no Brasil. Bio Brasil 2012, 1, 74–83. [Google Scholar]
- Pérez-Irineo, G.; Santos-Moreno, A. Abundance, herd size, activity pattern and occupancy of ungulates in Southeastern Mexico. Anim. Biol. 2016, 66, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Scognamillo, D.; Maxit, I.E.; Sunquist, M.; Polisar, J. Coexistence of jaguar (Panthera onca) and puma (Puma concolor) in a mosaic landscape in the Venezuelan llanos. J. Zool. 2003, 259, 269–279. [Google Scholar] [CrossRef]
- Morais, B.H.S.; Cardoso, D.L.; Costa, J.S.; Mayor, P.; Albuquerque, N.I.; Chisté, R.C.; Guimarães, D.A.A. Use of wildlife as an alternative protein source: Collared peccary meat. Meat Sci. 2022, 192, 108895. [Google Scholar] [CrossRef] [PubMed]
- Castelo, T.S.; Bezerra, F.S.; Lima, G.L.; Alves, H.M.; Oliveira, I.R.; Santos, E.A.; Peixoto, G.C.X.; Silva, A.R. Effect of centrifugation and sugar supplementation on the semen cryopreservation of captive collared peccaries (Tayassu tajacu). Cryobiology 2010, 61, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.A.S.; Sousa, P.C.; Peixoto, G.C.X.; Simão, B.R.; Oliveira, M.F.; Silva, A.R. Establishing the hypoosmotic swelling test for sperm analysis in collared peccaries (Pecari tajacu). Arq. Bras. Med. Vet. Zootec. 2013, 65, 1257–1260. [Google Scholar] [CrossRef] [Green Version]
- Souza, A.L.P.; Lima, G.L.; Peixoto, G.C.X.; Oliveira, M.F.; Silva, A.R. Use of Aloe vera–based extender for chilling and freezing collared peccary (Pecari tajacu) semen. Theriogenology 2016, 85, 1432–1438. [Google Scholar] [CrossRef]
- Moreira, S.S.; Santos, C.S.; Castelo, T.S.; Bezerra, L.G.P.; Praxedes, É.C.G.; Matos, T.M.; Souza-Junior, J.B.F.; Feijó, F.M.C.; Comizzoli, P.; Silva, A.R. Investigating the need for antibiotic supplementation to the extender used for semen cryopreservation in collared peccaries. Front. Vet. Sci. 2022, 9, 954921. [Google Scholar] [CrossRef]
- Moreira, S.S.; Silva, A.M.; Pereira, A.G.; Santos, R.P.; Dantas, M.R.; Souza-Júnior, J.B.; Snoeck, P.P.N.; Silva, A.R. Effect of detergents based on sodium dodecyl sulfate on functional metrics of frozen–thawed collared peccary (Pecari tajacu) semen. Animals 2023, 13, 451. [Google Scholar] [CrossRef]
- Santos, E.A.; Sousa, P.C.; Martins, J.A.M.; Moreira, R.A.; Monteiro-Moreira, A.C.O.; Moreno, F.B.M.B.; Oliveira, M.F.; Moura, A.A.; Silva, A.R. Protein profile of the seminal plasma of collared peccaries (Pecari tajacu Linnaeus, 1758). Reproduction 2014, 147, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Moreira, S.S.; Silva, A.M.; Praxedes, É.C.; Campos, L.B.; Santos, C.S.; Souza, A.L.; Pereira, A.F.; Souza-Júnior, J.B.; Costa, L.L.; Silva, A.R. Composition of collared peccary seminal plasma and sperm motility kinetics in semen obtained during dry and rainy periods in a semiarid biome. Anim. Reprod. Sci. 2019, 211, 91–101. [Google Scholar] [CrossRef]
- Peixoto, G.C.; Lima, G.L.; Maia, K.M.; Souza, A.L.P.; Castelo, T.S.; Paiva, A.L.; Paula, V.V.; Oliveira, M.F.; Brito, A.B.; Domingues, S.F.S.; et al. Single injection of eCG/hCG leads to successful estrous synchronization in the collared peccary (Pecari tajacu Linnaeus, 1758). Anim. Reprod. Sci. 2019, 208, 106112. [Google Scholar] [CrossRef]
- Santos, M.V.O.; Silva, A.R.; Pereira, A.F. Embryo production by in vitro fertilization in wild ungulates: Progress and perspectives. Ann. Anim. Sci. 2022, 22, 1151–1162. [Google Scholar] [CrossRef]
- Morrell, J.M. Effect of colloid centrifugation on boar sperm quality during storage and function in in vitro fertilization. Theriogenology 2019, 137, 122–126. [Google Scholar] [CrossRef]
- Deori, S.; Ntallaris, T.; Wallgren, M.; Morrell, J.M.; Johannisson, A. Comparison of single layer centrifugation and magnetic activated cell sorting for selecting viable boar spermatozoa after thawing. Livest. Sci. 2022, 257, 104853. [Google Scholar] [CrossRef]
- Arias, M.E.; Andara, K.; Briones, E.; Felmer, R. Bovine sperm separation by swim-up and density gradients (Percoll and BoviPure): Effect on sperm quality, function, and gene expression. Reprod. Biol. 2017, 17, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Sekhon, L.; Kim, Y.; Agarwal, A. The role of oxidative stress and antioxidants in assisted reproduction. Curr. Womens Health Rev. 2010, 6, 227–238. [Google Scholar] [CrossRef]
- Campos, L.B.; Silva, M.A.; Bezerra, J.A.B.; Castelo, T.S.; Peixoto, G.C.X.; Silva, A.R. Survival of collared peccaries (Pecari tajacu) sperm after freezing-thawing using different extenders. Acta Sci. Vet. 2014, 42, 1217. [Google Scholar]
- Navarro-Serna, S.; París-Oller, E.; Simonik, O.; Romar, R.; Gadea, J. Replacement of Albumin by preovulatory oviductal fluid in swim-up sperm preparation method modifies boar sperm parameters and improves in vitro penetration of oocytes. Animals 2021, 11, 1202. [Google Scholar] [CrossRef]
- Pavaneli, A.P.P.; Passarelli, M.S.; Freitas, F.V.; Ravagnani, G.M.; Torres, M.A.; Martins, S.M.M.K.; Yeste, M.; Andrade, A.F.C. Removal of seminal plasma prior to liquid storage of boar spermatozoa: A practice that can improve their fertilizing ability. Theriogenology 2019, 125, 79–86. [Google Scholar] [CrossRef]
- Pavaneli, A.P.P.; Recuero, S.; Chaves, B.R.; Garcia-Bonavila, E.; Llavanera, M.; Pinart, E.; Bonet, S.; Andrade, A.F.C.; Yeste, M. The presence of seminal plasma during liquid storage of pig spermatozoa at 17 °C modulates their ability to elicit in vitro capacitation and trigger acrosomal exocytosis. Int. J. Mol. Sci. 2020, 21, 4520. [Google Scholar] [CrossRef] [PubMed]
- Cabello, Y.; Belchín, P.; González-Martínez, M.; López-Fernández, C.; Johnston, S.; Gosálvez, J. The efficacy of novel centrifugation-free sperm selection (Io-Lix) on sperm parameters and ICSI reproductive outcomes. Reprod. Biomed. Online 2023, 46, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, M.; Yoshioka, K.; Hikono, H.; Iwagami, G.; Suzuki, C.; Kikuchi, K. Centrifugation on Percoll density gradient enhances motility, membrane integrity and in vitro fertilizing ability of frozen–thawed boar sperm. Zygote 2013, 23, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Olivares, C.C.S.; Fonseca, J.F.; Camargo, L.S.A.; Souza-Fabjan, J.M.G.; Rodrigues, A.L.R.; Brandão, F.Z. Comparison of different methods of goat sperm selection and capacitation for optimization of assisted reproductive technologies. Small Rumin. Res. 2015, 127, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Vega-Hidalgo, J.; Rodriguez, M.; Dipaz-Berrocal, D.; Rivas, J.; Huayhua, C.; Mellisho, E. Sperm selection techniques in cattle: Microfilter device versus conventional methods. Andrologia 2022, 54, e14585. [Google Scholar] [CrossRef] [PubMed]
- Fishel, S.; Jackson, P.; Webster, J.; Faratian, B. Endotoxins in culture medium for human in vitro fertilization. Fertil. Steril. 1988, 49, 108–111. [Google Scholar] [CrossRef]
- Matás, C.; Vieira, L.; García-Vázquez, F.A.; Avilés-López, K.; López-Úbeda, R.; Carvajal, J.A.; Gadeia, J. Effects of centrifugation through three different discontinuous Percoll gradients on boar sperm function. Anim. Reprod. Sci. 2011, 127, 62–72. [Google Scholar] [CrossRef]
- Anel-Lopez, L.; Ortega-Ferrusola, C.; Álvarez, M.; Borragán, S.; Chamorro, C.; Peña, F.J.; Morrell, L.; Anel, L.; Paz, P. Improving sperm banking efficiency in endangered species through the use of a sperm selection method in brown bear (Ursus arctos) thawed sperm. BMC Vet. Res. 2017, 13, 200. [Google Scholar] [CrossRef] [Green Version]
- Mehta, A.; Sigman, M. Identification and preparation of sperm for ART. Urol. Clin. 2014, 41, 169–180. [Google Scholar] [CrossRef]
- Souza, A.L.P.; Castelo, T.S.; Queiroz, J.P.A.F.; Barros, I.O.; Paula, V.V.; Oliveira, M.F.; Silva, A.R. Evaluation of anesthetic protocol for the collection of semen from captive collared peccaries (Tayassu tajacu) by electroejaculation. Anim. Reprod. Sci. 2009, 16, 370–375. [Google Scholar] [CrossRef]
- Waberski, D.; Magnus, F.; Ardon, F.; Petrunkin, A.M.; Weitze, K.F.; Petersen, E.T. Binding of boar spermatozoa to oviductal epithelium in vitro in relation to sperm morphology and storage time. Reproduction 2006, 131, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Mun, S.-E.; Sim, B.-W.; Yoon, S.-B.; Jeong, P.-S.; Yang, H.-J.; Choi, S.-A.; Park, H.-O.; Kim, Y.-H.; Kang, P.; Kang, K.-J.; et al. Dual effect of fetal bovine serum on 21 early development depends on stage-specific reactive oxygen species demands in pigs. PLoS ONE 2017, 12, 22 e0175427. [Google Scholar] [CrossRef] [Green Version]
- Sousa, P.C.; Santos, E.A.A.; Souza, A.L.P.; Lima, G.L.; Barros, F.F.P.C.; Oliveira, M.F.; Silva, A.R. Sperm morphological and morphometric evaluation in captive collared peccaries (Pecari tajacu). Pesqui. Vet. Bras. 2013, 33, 924–930. [Google Scholar] [CrossRef] [Green Version]
- Celeghini, E.C.C.; Arruda, R.P.; Andrade, A.F.C.; Nascimento, J.; Raphael, C.F. Practical techniques for bovine sperm simulta- neous fluorimetric assessment of plasma, acrosomal and mitochondrial membranes. Reprod. Domest. Anim. 2007, 42, 479–488. [Google Scholar] [CrossRef]
- Castro, L.S.; Assis, P.M.; Siqueira, A.F.; Hamilton, T.R.; Mendes, C.M.; Losano, J.D.; Nichi, M.; Visintin, J.A.; Assumpção, M.E.O.A. Sperm oxidative stress is detrimental to embryo development: A dose-dependent study model and a new and more sensitive oxidative status evaluation. Oxid. Med. Cell. Longev. 2016, 2016, 8213071. [Google Scholar] [CrossRef] [Green Version]
- Martins, C.F.; Dode, M.N.; Bao, S.N.; Rumpf, R. The use of the acridine orange test and the TUNEL assay to assess the integrity of freeze-dried bovine spermatozoa DNA. Gen. Mol. Res. 2007, 6, 94–104. [Google Scholar]
- Pang, Y.W.; Sun, Y.Q.; Jiang, X.L.; Huang, Z.Q.; Zhao, S.J.; Du, W.H.; Hao, H.S.; Zhao, X.M.; Zhu, H.B. Protective effects of melatonin on bovine sperm characteristics and subsequent in vitro embryo development. Mol. Reprod. Dev. 2016, 83, 993–1002. [Google Scholar] [CrossRef]
- Gambini, A.; Stéfano, A.; Jarazo, J.; Buemo, C.; Karlanian, F.; Salamone, D.F. Embryo aggregation does not improve the development of interspecies somatic cell nuclear transfer embryos in the horse. Theriogenology 2016, 86, 1081–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rath, D.; Long, C.R.; Dobrinsky, J.R.; Welch, G.R.; Schreier, L.L.; Johnson, L.A. In vitro production of sexed embryos for gender preselection: High-speed sorting of X-chromosome-bearing sperm to produce pigs after embryo transfer. J. Anim. Sci. 1999, 77, 3346–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, X.G.; Cai, M.M.; Zhang, Y.T.; Liu, Y.; Gao, Z.L.; Song, J.; Liu, Z.H. Effect of Astragalus polysaccharide addition to thawed boar sperm on in vitro fertilization and embryo development. Theriogenology 2018, 121, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.A.; Santos, M.V.O.; Nascimento, L.E.; Lira, G.P.O.; Praxedes, É.A.; Oliveira, M.F.; Silva, A.R.; Pereira, A.F. Production of collared peccary (Pecari tajacu Linnaeus, 1758) parthenogenic embryos following different oocyte chemical activation and in vitro maturation conditions. Theriogenology 2020, 142, 320–327. [Google Scholar] [CrossRef]
- Liu, D.Y.; Clarke, C.N.; Baker, H.W.G. Relationship between sperm motility assessed with the Hamilton-Thorn motility analyzer and fertilization rates in vitro. J. Androl. 1991, 12, 231–239. [Google Scholar]
- Holt, C.; Holt, W.V.; Moore, H.D.; Reed, H.C.; Curnock, R.M. Objectively measured boar sperm motility parameters correlate with the outcomes of on-farm inseminations: Results of two fertility trials. J. Androl. 1997, 18, 312–323. [Google Scholar]
- Broekhuijse, M.L.W.J.; Šoštarić, E.; Feitsma, H.; Gadella, B.M. Application of computer-assisted semen analysis to explain variations in pig fertility. J. Anim. Sci. 2012, 90, 779–789. [Google Scholar] [CrossRef]
- Henkel, R.R.; Schill, W.B. Sperm preparation for ART. Reprod. Bioland Endocrinol. 2003, 1, 108. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Li, R.; Lv, Y.; Zeng, W. Melatonin protects rabbit spermatozoa from cryo-damage via decreasing oxidative stress. Cryobiology 2019, 88, 1–8. [Google Scholar] [CrossRef]
- Martínez-Pastor, F.; Lacalle, E.; Martínez-Martínez, S.; Fernández-Alegre, E.; Álvarez-Fernández, L.; Martinez-Alborcia, M.J.; Bolarin, A.; Morrell, J.M. Low density Porcicoll separates spermatozoa from bacteria and retains sperm quality. Theriogenology 2021, 165, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.S.; Campos, L.B.; Praxedes, É.C.G.; Moreira, S.S.J.; Souza-Júnior, J.B.F.; Comizzoli, P.; Feijó, F.M.C.; Silva, A.R. Influence of antibiotics on bacterial load and sperm parameters during short-term preservation of collared peccary semen. Anim. Reprod. 2021, 18, e20210021. [Google Scholar] [CrossRef] [PubMed]
- Ben, L.; Yan, C.; Yu, S.J. Effect of swim-up and percoll treatment on sperm quality and in vitro embryo development in yak. J. Integr. Agric. 2013, 12, 2235–2242. [Google Scholar] [CrossRef] [Green Version]
- Cruz, R.S.; Giuliano, S.M.; Gambarotta, M.C.; Morrell, J.M.; Abraham, M.C.; Miragaya, M.H.; Carretero, M.I. Comparison of different methods of sperm selection of llama raw semen. Anim. Reprod. Sci. 2016, 173, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Pradieé, J.; Sánchez-Calabuig, M.J.; Castaño, C.; O’Brien, E.; Esteso, M.C.; Beltrán-Breña, P.; Maillo, V.; Santiago-Moreno, J.; Rizos, D. Fertilizing capacity of vitrified epididymal sperm from Iberian ibex (Capra pyrenaica). Theriogenology 2017, 108, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Campos, L.B.; Peixoto, G.C.X.; Silva, A.M.; Souza, A.L.P.; Castelo, T.S.; Maia, K.M.; Pereira, A.F.; Silva, A.R. Estimating the binding ability of collared peccary (Pecari tajacu Linnaeus, 1758) sperm using heterologous substrates. Theriogenology 2017, 92, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Matás, C.; Coy, P.; Romar, R.; Marco, M.; Gadea, J.; Ruiz, S. Effect of sperm preparation method on in vitro fertilization in pigs. Reproduction 2003, 125, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Cunha, A.T.; Carvalho, J.O.; Guimarães, A.L.; Leme, L.O.; Caixeta, F.M.; Viana, J.H.; Dode, M.A. Bovine epididymal spermatozoa treatment for in vitro fertilization: Heparin accelerates fertilization and enables a reduction in coincubation time. PLoS ONE 2019, 14, e0209692. [Google Scholar] [CrossRef]
- Bezerra, L.G.P.; Souza, A.L.P.; Silva, H.V.R.; Vasconcelos, F.R.; Moura, A.D.A.A.; Pereira, A.F.; Oliveira, M.F.; Silva, A.R. Ultrastructural description of fresh and frozen/thawed sperm derived from collared peccaries (Pecari tajacu Linnaeus, 1758). Microsc. Res. Tech. 2018, 81, 1301–1309. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CASA | Control (No Separation) | Swim-Up | |
|---|---|---|---|
| Centrifuged (without SP) | Non-Centrifuged (with SP) | ||
| Motility (%) | 90.4 ± 2.4 a | 31.4 ± 8.1 b | 51.9 ± 6.4 b |
| Progressive motile (%) | 68.6 ± 3.9 a | 17.7 ± 3.8 b | 28.9 ± 3.8 b |
| Velocity average pathway (VAP; µm/s) | 74.0 ± 4.5 a | 47.2 ± 5.1 b | 47.9 ± 1.9 b |
| Velocity straight line (VSL; μm/s) | 54.2 ± 6.1 a | 37.6 ± 4.3 b | 37.6 ± 2.0 b |
| Curvilinear velocity (VCL; μm/s) | 137.6 ± 5.3 a | 91.1 ± 9.1 b | 88.4 ± 3.5 b |
| Amplitude of lateral head (ALH; μm) | 6.1 ± 0.2 a | 5.1 ± 0.2 b | 4.8 ± 0.3 b |
| Beat cross frequency (BCF; Hz) | 36.3 ± 0.5 a | 35.3 ± 1.4 a | 35.9 ± 1.0 a |
| Straightness (STR; %) | 74.0 ± 3.1 a | 74.7 ± 1.5 a | 73.4 ± 2.3 a |
| Linearity (LIN; %) | 42.3 ± 3.2 a | 43.7 ± 1.8 a | 43.3 ± 2.5 a |
| Rapid (%) | 80.1 ± 2.9 a | 21.1 ± 5.2 b | 35.1 ± 4.2 b |
| Medium (%) | 9.9 ± 1.3 a | 10.3 ± 3.8 a | 17.1 ± 2.6 a |
| Slow (%) | 2.7 ± 0.6 a | 6.1 ± 1.4 a | 6.1 ± 1.8 a |
| Static (%) | 7.0 ± 2.2 a | 62.1 ± 8.6 b | 41.7 ± 8.0 b |
| CASA | Control (No Separation) | Percoll® Gradient | |
|---|---|---|---|
| PG 45–90% | PG 35–70% | ||
| Motility (%) | 95.0 ± 1.8 a | 87.5 ± 3.6 a | 8.2 ± 2.1 b |
| Progressive motile (%) | 70.8 ± 6.0 a | 75.3 ± 5.2 a | 3.3 ± 1.4 b |
| Velocity average pathway (VAP; µm/s) | 69.7 ± 7.5 b | 96.8 ± 9.4 a | 40.7 ± 6.1 b |
| Velocity straight line (VSL; μm/s) | 56.0 ± 6.6 b | 88.4 ± 8.9 a | 27.3 ± 7.0 b |
| Curvilinear velocity (VCL; μm/s) | 121.4 ± 11.4 a | 129.6 ±7.5 a | 95.4 ± 12.0 a |
| Amplitude of lateral head (ALH; μm) | 5.4 ± 0.4 a | 4.4 ± 0.3 a | 6.2 ± 0.6 b |
| Beat cross frequency (BCF; Hz) | 36.4 ± 0.9 a | 36.0 ± 1.9 a | 38.8 ± 2.2 a |
| Straightness (STR; %) | 76.0 ± 2.2 a | 87.2 ± 1.4 a | 64.7 ± 6.2 b |
| Linearity (LIN; %) | 46.0 ± 3.1 b | 66.0 ± 4.7 a | 35.7 ± 6.7 b |
| Rapid (%) | 81.0 ± 6.2 a | 79.7 ± 5.3 a | 5.2 ± 1.6 b |
| Medium (%) | 14.0 ± 4.9 a | 7.7 ± 1.8 ab | 2.8 ± 0.7 b |
| Slow (%) | 2.2 ± 0.3 a | 2.7 ± 0.7 a | 2.3 ± 0.6 a |
| Static (%) | 3.0 ± 1.6 a | 10.0 ± 3.0 b | 89.8 ± 2.6 c |
| CASA | Fresh Control | Percoll® Gradient | Washing by Centrifugation |
|---|---|---|---|
| Motility (%) | 96.8 ± 2.0 a | 66.3 ± 10.2 a | 89.0 ± 5.4 a |
| Progressive motile (%) | 59.5 ± 12.1 a | 47.5 ± 7.9 a | 56.0 ± 12.5 a |
| Velocity average pathway (VAP; µm/s) | 83.0 ± 8.7 a | 105.3 ± 7.3 a | 96.0 ± 8.8 a |
| Velocity straight line (VSL; μm/s) | 61.7 ± 9.3 b | 98.7 ± 7.2 a | 72.6 ± 9.3 ab |
| Curvilinear velocity (VCL; μm/s) | 157.3 ± 9.1 a | 139.4 ± 10.6 a | 166.7 ± 9.3 a |
| Amplitude of lateral head (ALH; μm) | 7.3 ± 0.2 b | 4.2 ± 0.6 a | 7.2 ± 0.6 b |
| Beat cross frequency (BCF; Hz) | 34.4 ± 0.7 a | 36.2 ± 2.5 a | 33.1 ± 1.7 a |
| Straightness (STR; %) | 71.3 ± 3.7 b | 92.0 ± 3.2 a | 72.5 ± 3.6 b |
| Linearity (LIN; %) | 40.3 ± 3.9 b | 72.3 ± 4.7 a | 46.0 ± 4.7 b |
| Rapid (%) | 62.5 ± 12.3 a | 48.8 ± 8.5 a | 59.5 ± 12.6 a |
| Medium (%) | 11.8 ± 4.0 a | 4.0 ± 4.0 a | 7.0 ± 1.5 a |
| Slow (%) | 22.3 ± 6.7 a | 8.5 ± 2.7 a | 22.3 ± 6.2 a |
| Static (%) | 3.3 ± 2.0 a | 39.0 ± 11.2 b | 11.0 ± 5.4 a |
| Normal morphology (%) | 79.7 ± 7.4 a | 82.0 ± 6.4 a | 82.0 ± 6.4 a |
| Selection Technique | Matured Oocytes, % | Sperm Bound to Matured Oocytes | Sperm Bound to Non-Matured Oocytes | Sperm Bound to Total Oocytes | Second Polar Body, % | Two Pronuclei, % | Fertilized Oocytes | |
|---|---|---|---|---|---|---|---|---|
| Total, % | Matured, % | |||||||
| Percoll gradient | 65.6 ± 1.2 (40/61) | 4.9 ± 2.8 (196/40) | 9.3 ± 4.3 (196/21) | 3.2 ± 1.6 (196/61) | 9.8 ± 3.1 (6/61) | 4.9 ± 2.7 (3/61) | 14.8 ± 4.9 (9/61) | 22.5 ± 10.8 (9/40) |
| Washing by centrifugation | 52.5 ± 1.5 (32/61) | 3.2 ± 1.9 (103/32) | 3.6 ± 4.7 (103/29) | 1.7 ± 1.3 (103/61) | 3.3 ± 2.4 (2/61) | 9.8 ± 3.6 (6/61) | 13.1 ± 4.0 (8/61) | 25.0 ± 9.5 (8/32) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, M.V.O.; Silva, A.M.; Aquino, L.V.C.; Oliveira, L.R.M.; Moreira, S.S.J.; Oliveira, M.F.; Silva, A.R.; Pereira, A.F. Different Methods for Seminal Plasma Removal and Sperm Selection on the Quality and Fertility of Collared Peccary Sperm. Animals 2023, 13, 1955. https://doi.org/10.3390/ani13121955
Santos MVO, Silva AM, Aquino LVC, Oliveira LRM, Moreira SSJ, Oliveira MF, Silva AR, Pereira AF. Different Methods for Seminal Plasma Removal and Sperm Selection on the Quality and Fertility of Collared Peccary Sperm. Animals. 2023; 13(12):1955. https://doi.org/10.3390/ani13121955
Chicago/Turabian StyleSantos, Maria V. O., Andréia M. Silva, Leonardo V. C. Aquino, Lhara R. M. Oliveira, Samara S. J. Moreira, Moacir F. Oliveira, Alexandre R. Silva, and Alexsandra F. Pereira. 2023. "Different Methods for Seminal Plasma Removal and Sperm Selection on the Quality and Fertility of Collared Peccary Sperm" Animals 13, no. 12: 1955. https://doi.org/10.3390/ani13121955