
Eriocheir sinensis feminization-1c (Fem-1c) and Its Predicted miRNAs Involved in Sexual Development and Regulation

1
School of Marine Sciences, Ningbo University, Ningbo 315020, China
2
Laboratory for Marine Biology and Biotechnology, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China
3
DECAPODA Biology Science and Technology Co., Ltd. (Lianyungang), Lianyungang 222000, China
*
Author to whom correspondence should be addressed.
Animals 2023, 13(11), 1813; https://doi.org/10.3390/ani13111813
Submission received: 30 March 2023
/
Revised: 28 May 2023
/
Accepted: 29 May 2023
/
Published: 30 May 2023
(This article belongs to the Special Issue Crustaceans Biology, Physiology, Reproduction and Development)
Abstract
:Simple Summary
The Fem-1c gene in Eriocheir sinensis (Henri Milne Edwards, 1854) (EsFem-1c) is related to sex differentiation; however, its functional mechanism is less reported. In this study, qRT-PCR results of EsFem-1c from the intersex crab implied that EsFem-1c plays a role in crab androgenic gland (AG) development. RNAi and scanning electron microscope observations revealed that EsFem-1c influenced sexual development in E. sinensis. A dual-luciferase reporter assay demonstrated that tcf-miR-315-5p binding to the 3′UTR sequence effectively inhibited the translation of the EsFem-1c gene, and tcf-miR-307 binding to the alternative splicing region in the 3′UTR sequence could increase EsFem-1c expression. In summary, this study provides a better understanding of the molecular regulation mechanism of EsFem-1c.
Abstract
Feminization-1c (Fem-1c) is important for sex differentiation in the model organism Caenorhabditis elegans. In our previous study, the basic molecular characteristics of the Fem-1c gene (EsFem-1c) in Eriocheir sinensis (Henri Milne Edwards, 1854) were cloned to determine the relationship with sex differentiation. In this study, the genomic sequence of EsFem-1c contained five exons and four introns, with an exceptionally long 3′UTR sequence. The qRT-PCR results of EsFem-1c demonstrated lower tissue expression in the androgenic gland of the intersex crab than the normal male crab, implying that EsFem-1c plays a role in crab AG development. RNA interference experiments and morphological observations of juvenile and mature crabs indicated that EsFem-1c influences sexual development in E. sinensis. A dual-luciferase reporter assay disclosed that tcf-miR-315-5p effectively inhibits the translation of the EsFem-1c gene, influencing male development. An intriguing finding was that miRNA tcf-miR-307 could increase EsFem-1c expression by binding to the alternative splicing region with a length of 248 bp (ASR-248) in the 3′UTR sequence. The present research contributes to a better understanding of the molecular regulation mechanism of EsFem-1c and provides a resource for future studies of the miRNA-mediated regulation of sexual development and regulation in E. sinensis.
1. Introduction
The Feminization-1 (Fem-1) gene is a well-known key regulator to control male sexual development in the cascade sex determination pathway of Caenorhabditis elegans; together with the Cullin-2 protein, and by ubiquitin-mediated proteolysis, it directs the proteasome-mediated degradation of the Transformer-1 (TRA-1) protein [1,2,3,4]. Doniach (1984) and Kimble (1984) demonstrated that a Fem-1 mutation in C. elegans can result in a sex switch from male to hermaphrodite [5,6]. Fem-1 influences courtship and sex determination in Drosophila melanogaster [7]. Several mammalian and invertebrate species have been found to carry Fem-1 family (Fem-1a, -1b, and -1c) genes [8,9,10]. They are distinguished by the conserved existence of ankyrin repeat domains, and are assumed to have different biological activities [11].
Recently, Fem-1 genes in several crustaceans have been studied. It was observed that the sexually dimorphic expression pattern is related to the development of sexual phenotype and the formation of germ cells [12]. Mrfem-1 and Mnfem-1 genes have specific strong positive signals in the ovary of Macrobrachium rosenbergii and Macrobrachium nipponense, and Mnfem-1 can interact with proteins containing ankyrin, cathepsin L, insulinase or ubiquitin, indicating that Mnfem-1 and Mrfem-1 could function in prawn sexual development and ovarian development [13,14]. The fem-1c gene has also been identified in Hyriopsis cumingii and E. sinensis differently regulating gonadal differentiation development. The HcFem-1c gene in Hyriopsis cumingii maintains testicular function and induces sperm discharge [2]. EsFem-1c transcripts are abundant in muscle for Eriocheir sinensis, in addition to expression in gonads [15]. Although these findings provide some insights into Fem-1c’s role in crustacean sex development and differentiation, they fail to elucidate its mechanisms.
As known, 3′ untranslated regions (3′UTRs) of messenger RNAs (mRNAs) notoriously regulate mRNA-regulation processes by serving as hubs for post-transcriptional control, such as stability, localization, and translation [16]. Functional elements in the 3′UTR sequences, such as Musashi binding elements (MBEs) and K boxes, play important roles in gene regulation [17,18]. The 3′UTR is also well-known as a target binding site for microRNA (miRNA), which regulates mRNA expression after transcription [19]. After forming the miRNA-induced silencing complex (miRISC), miRISC binds to specific mRNA and activates the regulation function [20]. In addition, alternative cleavage and polyadenylation (APA) and natural antisense transcript (NAT) have been demonstrated to influence 3′UTRs functions [21]. APA is an RNA transcription regulation mechanism that allows mRNA with different 3′ ends to be generated from a single gene using polyadenylation (poly(A)) sites selectively, resulting in alternative transcript and protein isoforms [22,23,24]. The reverse strand of endogenous encoding or non-encoding molecules transcribes NAT [25]. An NAT of Pvfem-1 is only present in the oocyte nucleus of the subadult female Pacific white shrimp, Penaeus vannamei, indicating a possible post-transcriptional mechanism in ovary development [26].
MiRNAs play a role in crustacean immunity, reproduction, cell proliferation, and apoptosis. Many studies on E. sinensis have explained how miRNAs target genes to control the post-transcriptional regulatory processes. For example, miR-2 and miR-133 can down-regulate cooperatively the 3′ UTR of the cyclin B gene in E. sinensis via cytoplasmic polyadenylation element (CPE) and miRNA-binding sites [27]. Moreover, miR-133b regulates the role of SAP30 in the spermatogenesis of E. sinensis by regulating histone deacetylation levels to ensure normal spermatogenesis and the sperm decondensation nuclear mechanism [28]. Furthermore, during the White Spot Syndrome Virus (WSSV) challenge in E. sinensis, MiR-7 suppresses Myd88-ILF2-(IL-16L) signaling pathways and increases WSSV replication [29]. The miR-17-3p is highly regulated during spermatogonia differentiation and development in E. sinensis [30]. The miR-305-5p, miR-263a-5p, and miR-7-5p play major regulatory roles in the metamorphosis of E. sinensis [31]. In E. sinensis, miR-34 and let-7b can down-regulate insulin-like androgenic gland hormone (IAG) gene expression, whereas miR-9-5p, let-7, and miR-8915 can down-regulate Double-sex (Dsx) gene expression [32].
The Chinese mitten crab, E. sinensis (Henri Milne Edwards, 1854), is an economically important aquatic animal in China. In previous studies, Fem-1 genes were discovered to play important roles in the early gonadal development of E. sinensis based on genomic and transcriptomic data [15]. In this study, we assessed the expression profiles of EsFem-1c in normal and intersex crabs (genetic female crabs with androgenic gland). RNA interference (RNAi) was used to investigate the effects of EsFem-1c on the gender characteristics of juvenile and mature crabs. The function of potential miRNAs binding to the 3′UTR of EsFem-1c was identified and analyzed by constructing the small RNAs (sRNAs) sequencing database. Finally, it was found that the gender-biased expression pattern of the alternative spliced 3′UTR with a length of 248 bp (ASR-248) in the EsFem-1c gene was the basis for regulating the function of EsFem-1c.
2. Materials and Methods
2.1. Collection of Animals and Samples
E. sinensis was collected from crab hatcheries in Jiangsu province, China. Because the number of experimental individuals and types of target tissue samples for different experiments differed, the crabs were kept conventionally in our laboratory for further experimentation and sampling.
After anesthetizing the mature normal crabs (body weight: 94.27 ± 8.14 g) and the intersex crabs (body weight: 91.32 ± 6.29 g) on ice for 5 min, different tissues including muscle, ovaries or testes, and androgenic glands were dissected and quickly frozen in liquid nitrogen for subsequent RNA extraction.
We selected female crabs for ovary sampling based on the description by Gu and He (1997), which were in stages Ⅱ, Ⅲ, and Ⅳ [33]. Testis samples were obtained from male crabs at various stages of development, including spermatocyte, spermatid, and sperm stages [34]. Furthermore, the androgenic gland tissues were sampled at the proliferation, synthesis, and secretion stages [35].
Male and female reproductive systems were collected to construct the miRNAs database. Ovary (O) and spermathecal (SA) samples were collected from female crabs. In contrast, testis (T), vas deferens (VD), accessory sex glands (ASG), ejaculatory ducts (ED), seminal vesicles (SV), and androgenic glands (AG) were sampled from male crabs.
All animal treatments and experimental procedures in the present study were carried out in accordance with Ningbo University’s Guide for the Use of Experimental Animals.
2.2. DNA, RNA, and miRNAs Preparation and cDNA Template Synthesis
Using the Marine Animal DNA kit (TIANGEN, Beijing, China), the genomic DNA of muscles was extracted from female and male crabs as directed. Total RNA was extracted from various samples using TRIZOL® reagent (Invitrogen, Waltham, MA, USA) according to manufacturer’s instructions. The cDNA template was synthesized from 1 µg of total RNA using the PrimeScript™ RT reagent Kit (with gDNA Eraser) (TaKaRa, Shiga, Japan). Moreover, according to the protocol, miRNAs were extracted from the reproduction system of male and female crabs using the PureLink miRNA Isolation Kit (Invitrogen, USA). Then, 2 µg miRNA was used to synthesize the miRNA cDNA template using the miRNA First Strand cDNA Synthesis Kit (Vazyme, Nanjing, China).
2.3. Cloning and Analysis of Fem-1c 3′UTR Sequence from Eriocheir Sinensis
To determine the sequence accuracy of EsFem-1c 3′UTR for future studies, the polymerase chain reaction (PCR) template and specific primers were utilized to perform the PCR (Table 1). Primer Premier software (version 6.0, Premier Biosoft, San Francisco, CA, USA) was used to design the primers [36].
A 25.0 µL mixture of approximately 0.5 µL of each primer (10 mM), 0.5 µL Ex Taq polymerase (TaKaRa, Japan), 1.0 µL cDNA template, 2.5 µL 10 × Ex Taq polymerase buffer, 2.0 µL dNTP, and 18.0 µL double-distilled water was used for the PCR. The PCR program included pre-denaturation at 95 °C for 3 min, denaturation at 95 °C for 30 s, annealing at 58 °C for 30 s, and extension at 72 °C for 80 s for 35 cycles. The products separated by gel electrophoresis were sequenced by Youkang (Yongkang, China). The sequences were trimmed of vector contamination using the VecScreen tool (https://www.ncbi.nlm.nih.gov/tools/vecscreen/ (accessed on 1 April 2023)) [37].
Poly (A) signal Miner (http://dnafsminer.bic.nus.edu.sg/PolyA.html (accessed on 1 April 2023)) was utilized to detect the position of the poly (A) signal [38]. The elements in 3′UTRs were analyzed using UTRScan (http://itbtools.ba.itb.cnr.it/utrscan (accessed on 1 April 2023)) [39].
2.4. Synthesis of dsRNA for Silencing EsFem-1c Gene by RNA Interference In Vivo
Specific primers cloned and cleaved a 406 bp fragment of the EsFem-1c gene (GenBank: KR108012) and a fragment of the EGFP gene (GenBank: U55761) into the pGEM@-T Easy Vector (Promega Biotech, Beijing, China) (Table 1). After sequencing, double-stranded RNAs (dsRNAs) including dsRNA EGFP and dsRNA EsFem-1c were synthesized using the TranscriptAidTM T7 high-yield transcription kit following the manufacturer’s protocol (Thermo Scientific Inc., San Jose, CA, USA). Nanodrop2000 (Thermo Scientific Inc., USA) was used to measure the concentration.
2.5. Target-Silencing EsFem-1c Gene in Juvenile and Mature Crabs
In this experiment, dsRNA EsFem-1c was injected into the mature mitten crabs (body weight 94.27 ± 8.14 g) and juvenile Ⅰ crabs (average body weight 10 mg).
There were three treatment groups in mature crabs: the blank control group (saline solution), the negative control group (dsRNA EGFP), and the treated group (dsRNA EsFem-1c). In mature crabs, the injection dose was 1 µg/g body weight. The dsRNA was injected into the coelom through the arthrodial membrane between the third and fourth pleopod by a 100 μL micro syringe (Shanghai Anting, Shanghai, China). After 24 h, mature crabs were sampled for qRT-PCR, including the eyestalk ganglion and ovary in females and testis and AG in male crabs. To analyze the molecular function of dsRNA EsFem-1c, IAG (GenBank ID: KU724192.1) and crustacean female sex hormone 1 (CFSH-1) (GenBank ID: OP351640) genes of E. sinensis were detected [40].
The blank control group (BC) and the treated group (dsRNA EsFem-1c) were conducted on juvenile Ⅰ crabs at 1 µg/g of body weight. The injection position was also at the arthrodial membrane between the third and fourth pleopod using the IM11-2 and M-152 micro injection system (NARISHIGE, Tokyo, Japan). Approximately 15 days after, the whole body was sampled at the juvenile Ⅲ stage and held in 1× phosphate-buffered saline (1× PBS) solution and 4% paraformaldehyde.
2.6. The Morphological Observation of Juvenile Crab
The general anatomical structures of the juvenile crab were observed under the stereomicroscope (Olympus, Tokyo, Japan). Scanning electron microscope (SEM) (Hitachi, Tokyo, Japan) was used to obtain micrographs of juvenile crabs. The juvenile crabs’ samples were prepared by the standard process described by Zhu et al. [40]. The samples for observation were prepared through ethanol gradient dehydration and tert-butyl alcohol replacement. After being frozen for 20 h at −20 °C, the samples were dried for 24 h in the Martin Christ Alpha 1—4LD plus freeze dryer (Martin Christ, Osterode am Harz, Germany) before being sprayed with gold by a Hitachi E-1010 ion sputtering device (Hitachi, Japan). The length of male penis was obtained by the software Image J and converted according to the scale of SEM images.
2.7. Small RNA Libraries Construction and Sequencing
RNA (1 µg) was used to isolate sRNA and construct libraries using the Illumina TruSeq small RNA sample preparation Kit (Illumina, San Diego, CA, USA). The sRNA was linked to the 5′ and 3′ adapters before being reverse-transcribed into cDNA. The cDNA library was evaluated using an Agilent Bioanalyzer 2100 system (Agilent Technologies, Santa Clara, CA, USA) after PCR amplification. Finally, the sequencing was performed via the Illumina Hiseq 2500 platform (LC-BIO, Hangzhou, China).
2.8. Bioinformatics Analysis of Sequencing Reads
After removing the low-quality reads, reads with 5′ adapter and ambiguous nucleotides, and reads without 3′ adapter or inserted tag from raw data using ACGT101-miR software (LC Sciences, Houston, TX, USA), the clean reads with lengths of 18–26 nt were processed for subsequent bioinformatics analyses. The selected clean reads were filtered in Rfam, Repbase, and other common RNA family databases, before being mapped to the genome of the mitten crab [41] and then to the miRBase 22.0 database [42] to identify novel or known miRNAs. The secondary structures, minimum free energy (mfe) value, and base bias of all identified miRNA on the first or each position were analyzed.
The miRNA read counts were normalized to norm values to measure miRNA expression [43]. Differential expression miRNAs (DEMs) among male and female reproduction were indicated as p-value and Log2 (Fold change). The threshold for significant differential expression was defined as p-value < 0.05. The miRanda (v3.3a) [44] and TargetScan (v5.0) [45] were employed to predict the target genes of the identified miRNAs and clarify their functions. Furthermore, the Gene Ontology (GO) enrichment analysis [46] was used to determine the biological functions of the predicted target genes. Moreover, the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was used to identify metabolic or signal transduction pathways [47].
2.9. Quantitative Real-Time PCR
The specific primers were used for quantitative real-time PCR (qRT-PCR) in the ABI 7500 system (Applied Biosystems, Waltham, MA, USA) with reverse transcription cDNA products of mRNA and miRNA (Table 2). A standard curve was used to measure amplification efficiency and ascertain primer specificity. Each sample was repeated in triplicates.
The qRT-PCR for mRNA was used to assay the transcript expression profiles of target genes in different cDNA templates using the TB Green Premix kit (2×) (TaKaRa, Japan). To standardize the mRNA expression, Es-β-actin (GenBank ID: ATO74508.1) was used as the reference gene. The reaction mixture and cycling conditions have been described in our previous studies [40]. EsFem-1c-qF/R had an amplification efficiency of 102.253%. EsCFSH-1-qF/R had an amplification efficiency of 105.895%. EsIAG-qF/R had an amplification efficiency of 101.054%, and the amplification efficiency of Es-β-actin-qF/R was 100.451%.
The qRT-PCR was used to determine the expression of miRNAs using the 2 × Taq Pro Universal SYBR qPCR Master Mix Kit (Vazyme, China). A 20.0 µL PCR mixture containing 10.0 µL of 2 × Taq Mix, 2.0 µL of diluted cDNA, 0.4 µL of miRNA specific primer, 0.4 µL of Universal Reverse Q primer, and 7.2 µL of RNase-free water was detected under the following conditions: denaturation at 95 °C for 10 s, annealing and extension at 60 °C for 30 s and 72 °C for 15 s, 40 cycles, respectively. To standardize miRNA expression, a single stranded U6 primer was used as an internal control [32].
2.10. Identification and Analysis of miRNAs Targeting EsFem-1c Gene
The binding sites located in the 3′UTR sequence of the EsFem-1c gene were detected using miRanda [44], RNA22 [48], and RNAHybird [49] based on the miRNA target prediction results. The analogs of miRNAs were synthesized chemically by General Biol. (General Biol., China). The present study used these analogs of predicted miRNAs to further investigate their effectiveness.
2.11. Luciferase 3′UTR Reporter Assay
To determine whether miRNAs can regulate the EsFem-1c gene, the interactions between them were examined using the dual-luciferase reporter assay in vitro.
The pGL4.10 [luc2] vector (Promega, China) was optimized to become the pGL4.10 [luc2]-basic plasmid (pGL-B), and the Xba I restriction site was inserted into the downstream of the firefly luciferase coding region via PCR amplification using specific primers (Table 3). Following enzyme digestion and purification, the 3′UTR sequence of EsFem-1c was cloned into the pGL-B to construct the pGL4.10 [luc2]-Fem1c-wild (Fem1c-WT). Similarly, the Mut Express® Ⅱ Fast Mutagenesis Kit (Vazyme, China) and the specific primers were used to construct the pGL4.10 [luc2]-Fem1c-mutant (Fem1c-MUT) (Table 3).
The cell line HEK 293T was obtained from the Chinese Academy of Science cell bank (Shanghai, China) and cultured in Dulbecco’s Modified Eagle Medium (DMEM; Hyclone, Logan, UT, USA) supplemented with 1% 100 × Penicillin-streptomycin-neomycin solution (100 × PSN; Gibco, Shanghai, China), and 10% fetal bovine serum (FBS; Thermo Fisher Scientific, China) at 37 °C and 5% CO2 [50]. Cells were seeded into the 24-well plates at 1.5 × 105 cells per well. When the cells were approximately 70–80% confluent, the co-transfection was conducted with 30 ng miRNAs mimics or the negative control (NC), 40 ng pGL4.74 [hRluc/TK] plasmid, and 200 ng Fem1c-WT or Fem1c-MUT plasmid, using Lipofectamine 3000 reagent (Thermo Fisher Scientific, Shanghai, China) according to the manufacturer’s protocol. Each sample was transfected in triplicate. Cells were incubated in serum- and antibiotic-free DMEM for 6 h, then 48 h transfection.
After harvesting cell lysates, luciferase activity was detected using the Dual-Luciferase Reporter Assay System kit (Promega, Beijing, China). The luc2/hRluc signal ratio was used to normalize the transfection efficiency between samples and calculate the relative luciferase measurement. The experiments were repeated thrice.
2.12. Statistical Analysis
3. Results
3.1. The Bioinformatics Analysis of EsFem-1c mRNA Sequence
The whole sequence of EsFem-1c was submitted to the NCBI database (GenBank number: KR108012) after being obtained from the E. sinensis genomic database [15]. There were five exons and four introns in the EsFem-1c sequence (Figure 1). The coding region measured 5976 base pairs (bp) and included a 1929 bp open read frame (ORF). The start codon was preceded by a 5′UTR of 205 bp, and a 3′UTR of 3914 bp followed the termination codon. The N-terminal region contained six consecutive ankyrin repeats (ANKs), while the C-terminal region contained two ANKs. The bioinformatics software forecast analysis results revealed one potential poly-adenylation signal (PAS; AATAAA) located in the 19 bp upstream of the poly (A) tract, and the analysis by the UTRScan software identified three K-boxes motifs (XTGTGATX) and six Musashi binding element sites (MBE; G/AT1-3AGT).
3.2. Expression Patterns Analysis of the EsFem-1c in Various Tissues of the Normal and Intersex Crabs
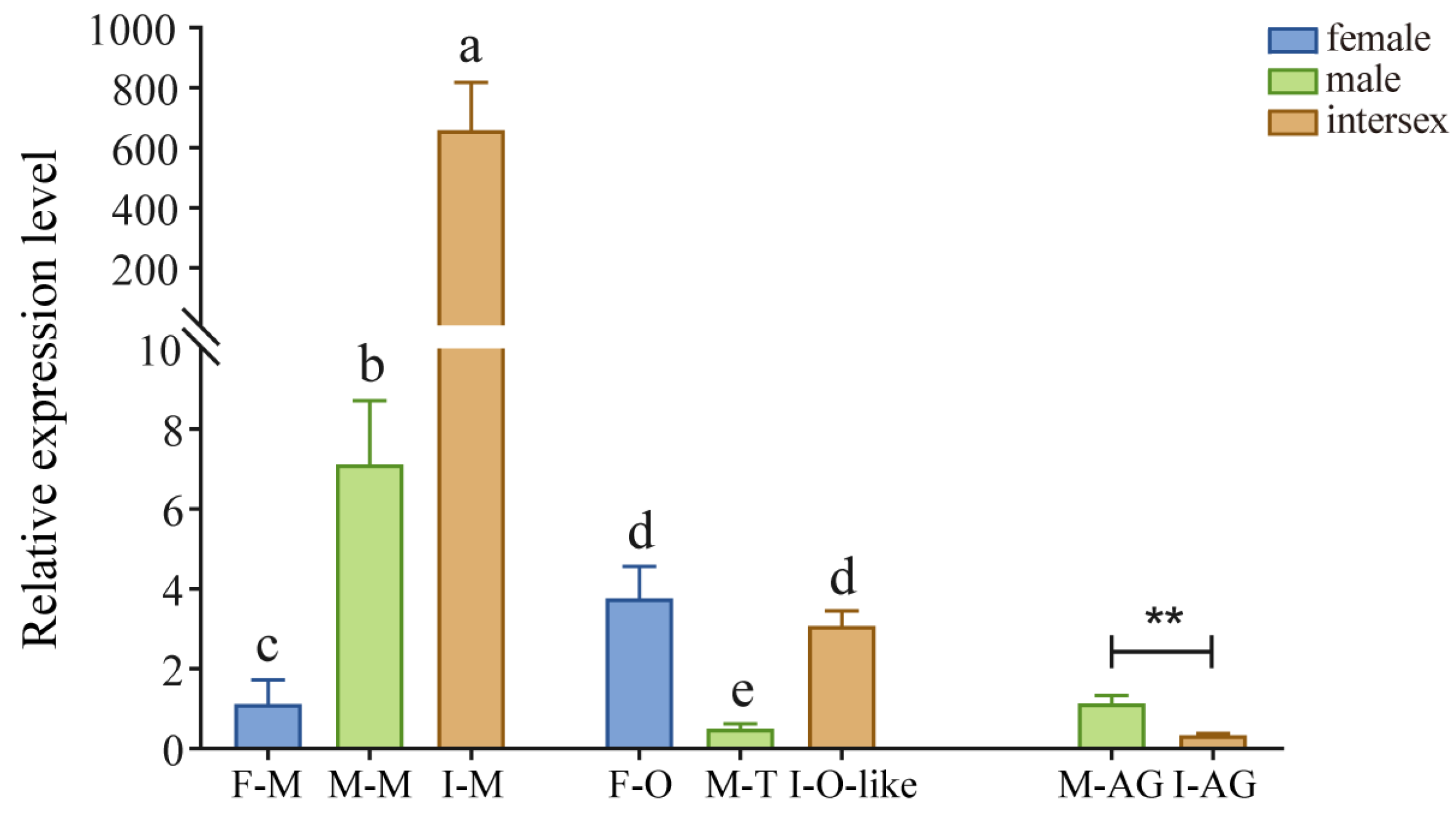
EsFem-1c gene expression was found in normal and intersex crab tissues, such as muscle, ovary, testes, and AG (Figure 2). EsFem-1c expression patterns in intersex crabs differed from those in normal female and male crabs. The gender of intersex crabs used in our experiment was distinguished to be female by our specific primers.
Notably, EsFem-1c expression was significantly higher in intersex crab muscles (p < 0.05), and the expression in normal male crabs was significantly higher than in normal female crabs (p < 0.05). Nevertheless, little difference existed between the normal female ovary and ovary-like tissue in intersex crabs. They were significantly higher (p < 0.05) than normal male testis. Regarding AG, EsFem-1c expression was significantly lower in intersex crabs than normal male crabs (p < 0.05).
3.3. The Expression Profiles of Sex Genes and Sexual Characteristics Change after Targeted-Silencing EsFem-1c in E. sinensis
Using dsRNA EsFem-1c, we investigated the roles of EsFem-1c in mature crabs and juveniles.
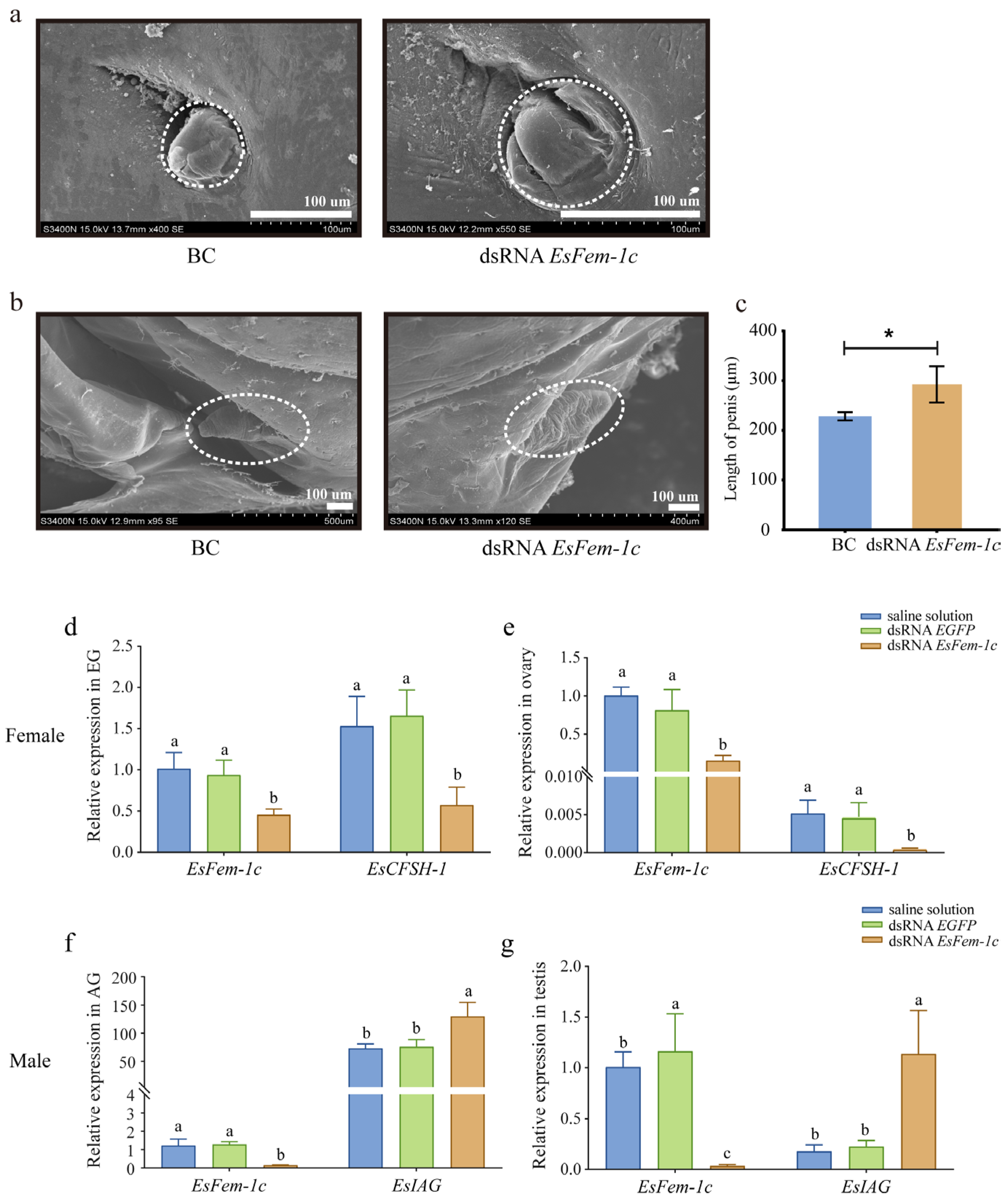
After being injected with dsRNA EsFem-1c, the crabs’ phenotypes, especially gonopore’s cover in the female crab and the AG in the male crab, changed from juvenile Ⅰ to Ⅲ, and the abnormal cleft structure of the gonopore’s cover was observed in female crabs at juvenile Ⅲ (Figure 3a). The appearance of the male penis did not change significantly when developed to juvenile Ⅲ (Figure 3b); however, the length became significantly longer than the control group (p < 0.05) (Figure 3c).
After target-silencing EsFem-1c, the expression levels of EsIAG and EsCFSH-1 in mature crabs differed from the control group. In female crab EGs and ovaries, the expression of EsCFSH-1 decreased significantly, with EsFem-1c silencing efficiencies of 65.1% and 80.2% (p < 0.05), respectively (Figure 3d,e). The silencing efficiency of dsRNA EsFem-1c in male crabs was 71.6% and 89.7% in AG and testis, respectively, while EsIAG in AGs and testes demonstrated an increased expression (p < 0.05) (Figure 3f,g).
3.4. The Expression of EsFem-1c in the Reproduction Tissues
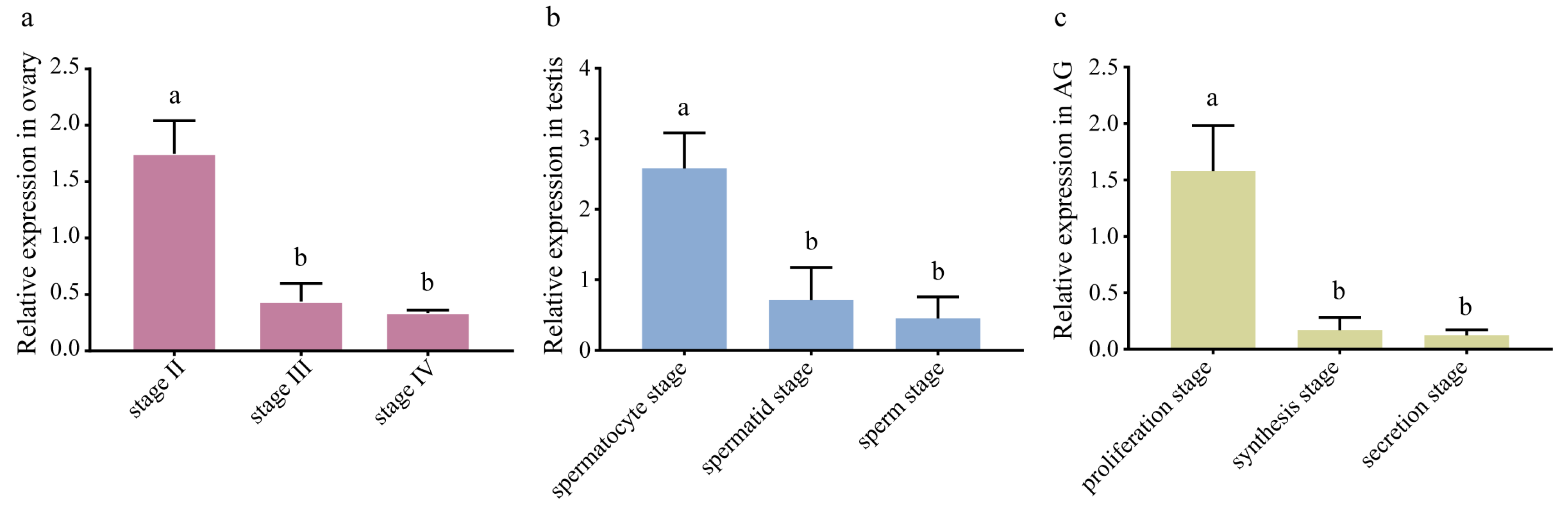
EsFem-1c mRNA expression was detected in the ovary, testis, and AG at various developmental stages. According to qRT-PCR results (Figure 4), EsFem-1c expression decreased with ovarian development from Stage Ⅱ to Ⅳ (p < 0.05). The mRNA transcript of EsFem-1c from the spermatocyte stage of the testis and proliferation stage of AG revealed a downward trend (p < 0.05) (Figure 4). Interestingly, the peak level of EsFem-1c in gonadal developmental stages all appeared during the early stages of development (Figure 4).
3.5. Overview of Small RNAs Sequencing in the Reproduction System of E. sinensis
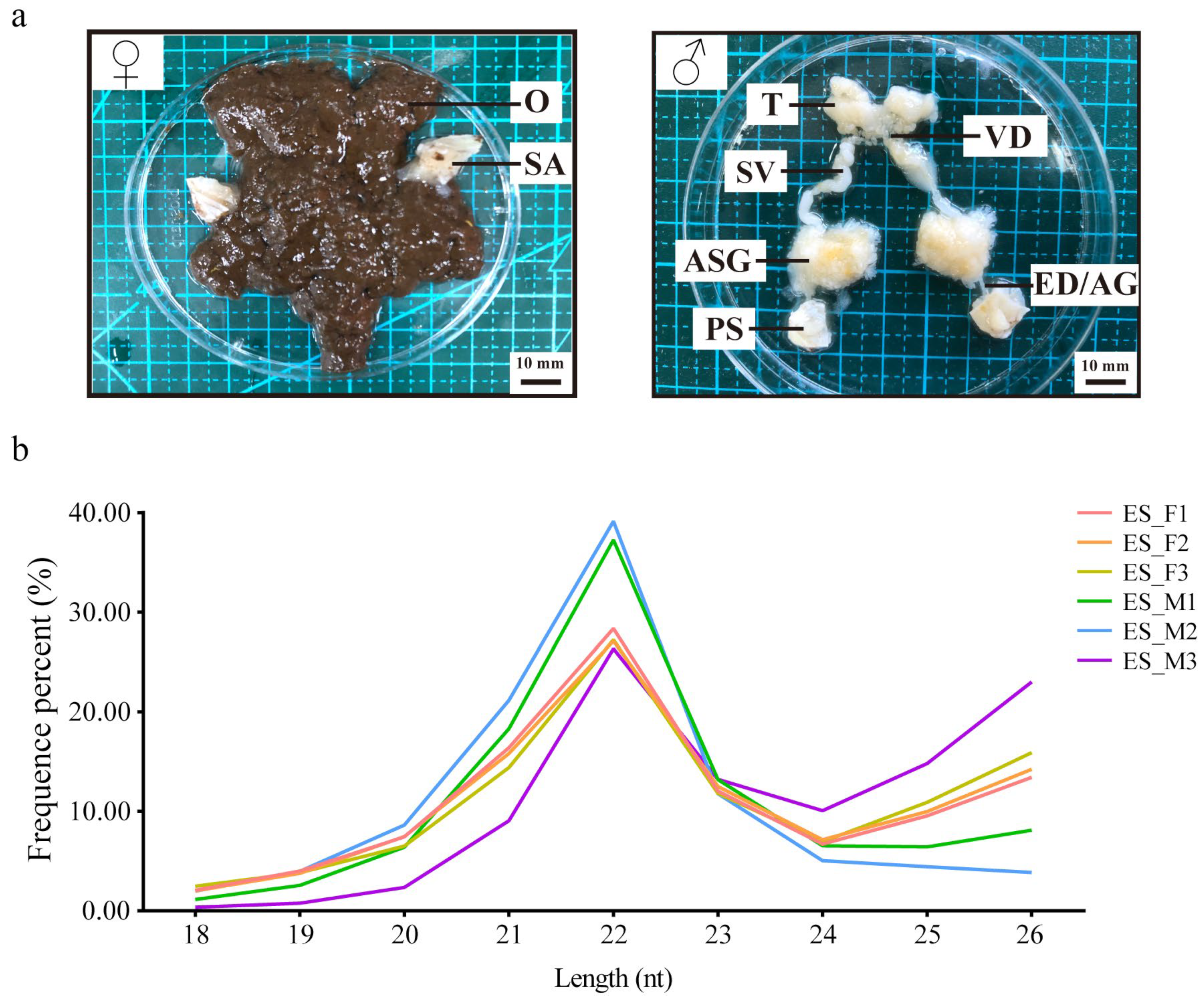
The six libraries (ES_F1, ES_F2, ES_F3, ES_M1, ES_M2, and ES_M3) were constructed using the E. sinensis reproduction system (Figure 5a). After removing the reads with the 3ADT and length filter and junk reads, 9,576,534, 7,908,841, 8,466,316, 11,347,130, 12,064,581, and 8,643,956 clean reads were obtained, which were then compared and filtered using databases (mRNA, RFam, and Repbase) to obtain sRNAs (Table S1). The length distribution of sRNAs indicated similar trends in females and males, with the peak at 22 nt representing a typical characteristic of miRNAs (Figure 5b). By searching the genome of E. sinensis using the miRBase 22.0 and miRDeep2 programs, 154 known miRNAs and 611 novel miRNAs were identified. Nucleotide bias analysis revealed that uridine (U) was dominant at the first position of the known miRNAs, and U and guanosine (G) were predominant at each position of the reads (Figure S1).
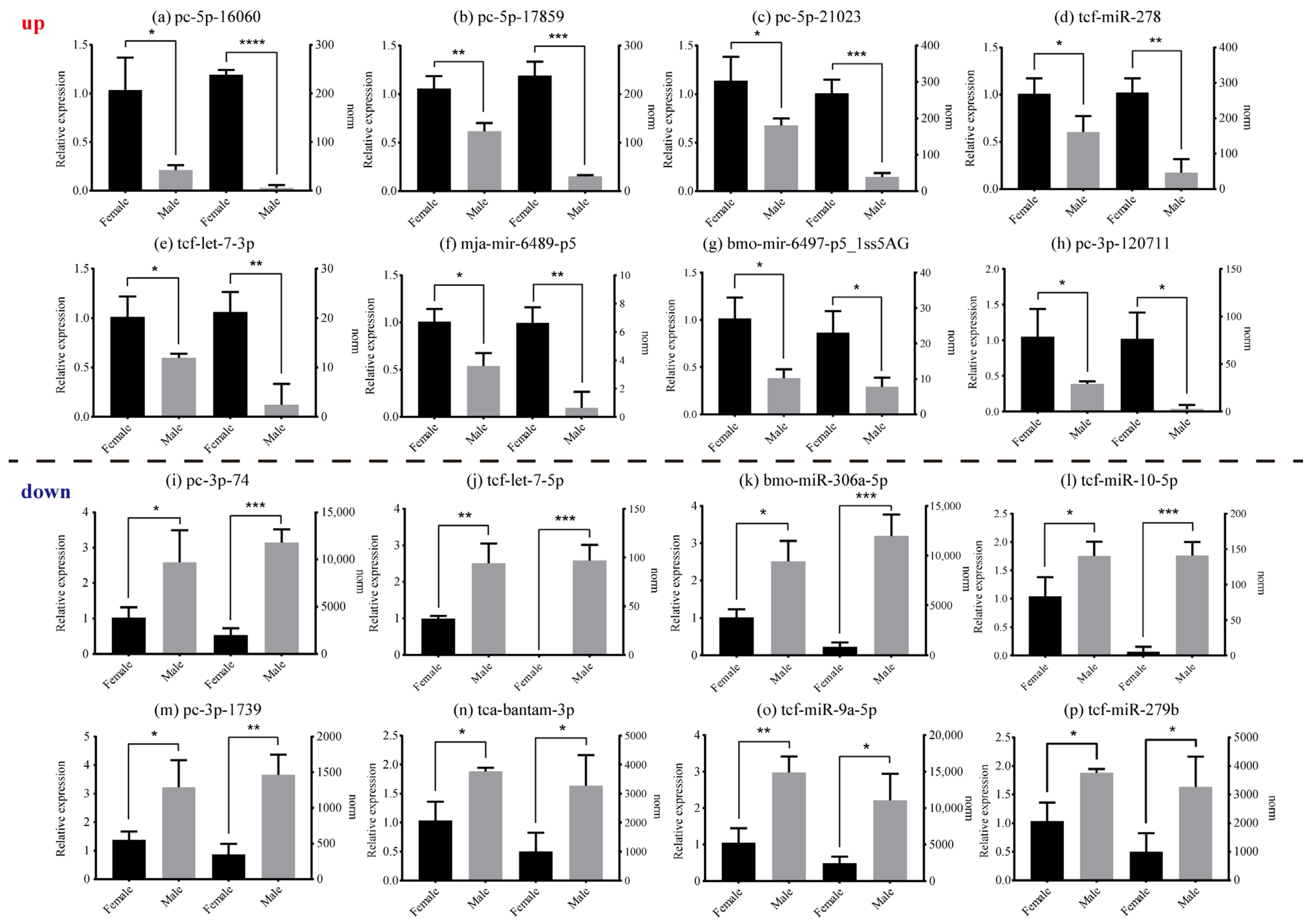
Based on differential expression analysis of miRNAs, the male and female reproduction tissues of E. sinensis expressed 43 significantly DE miRNAs (DEMs) (p < 0.05) (Table S2). Among these, 20 miRNAs presented significant up-regulation (female > male), while 23 miRNAs indicated down-regulation (male > female). PC-5p-16060 was the most significantly up-regulated miRNAs and PC-3p-74 was the most significantly down-regulated miRNAs. DEMs results were also used to generate a volcano plot depicting differential expression levels (Figure S2a), and cluster analysis was used to estimate miRNA expression cluster profiles in different groups (Figure S2b). As displayed in Figure 6, the qRT-PCR assay demonstrated that the results of the 16 miRNAs agreed with the norm values from the miRNA libraries.
The genes predicted to be targets of miRNAs were further classified using GO and KEGG pathway enrichment annotation analyses to investigate the potential pathways that these miRNAs might regulate. After analyzing the GO annotation results, target genes were identified to have the highest cornucopian gene counts in the deoxyribonucleotide biosynthetic process term (GO: 0009263) in the biological process, the integral component of the plasma membrane (GO: 0005887) in the cellular component, and the molecular function (GO: 0003674) in the molecular function (Figure S3a). Moreover, KEGG pathway enrichment analysis depicted that target genes statistically focused on three significant pathways, including the circadian rhythm (map04710), insulin resistance (map04931), and longevity regulating pathway (map04211) (Figure S3b).
3.6. The Interaction between the Identified miRNAs and EsFem-1c Gene by Dual-Luciferase Reporter Assay
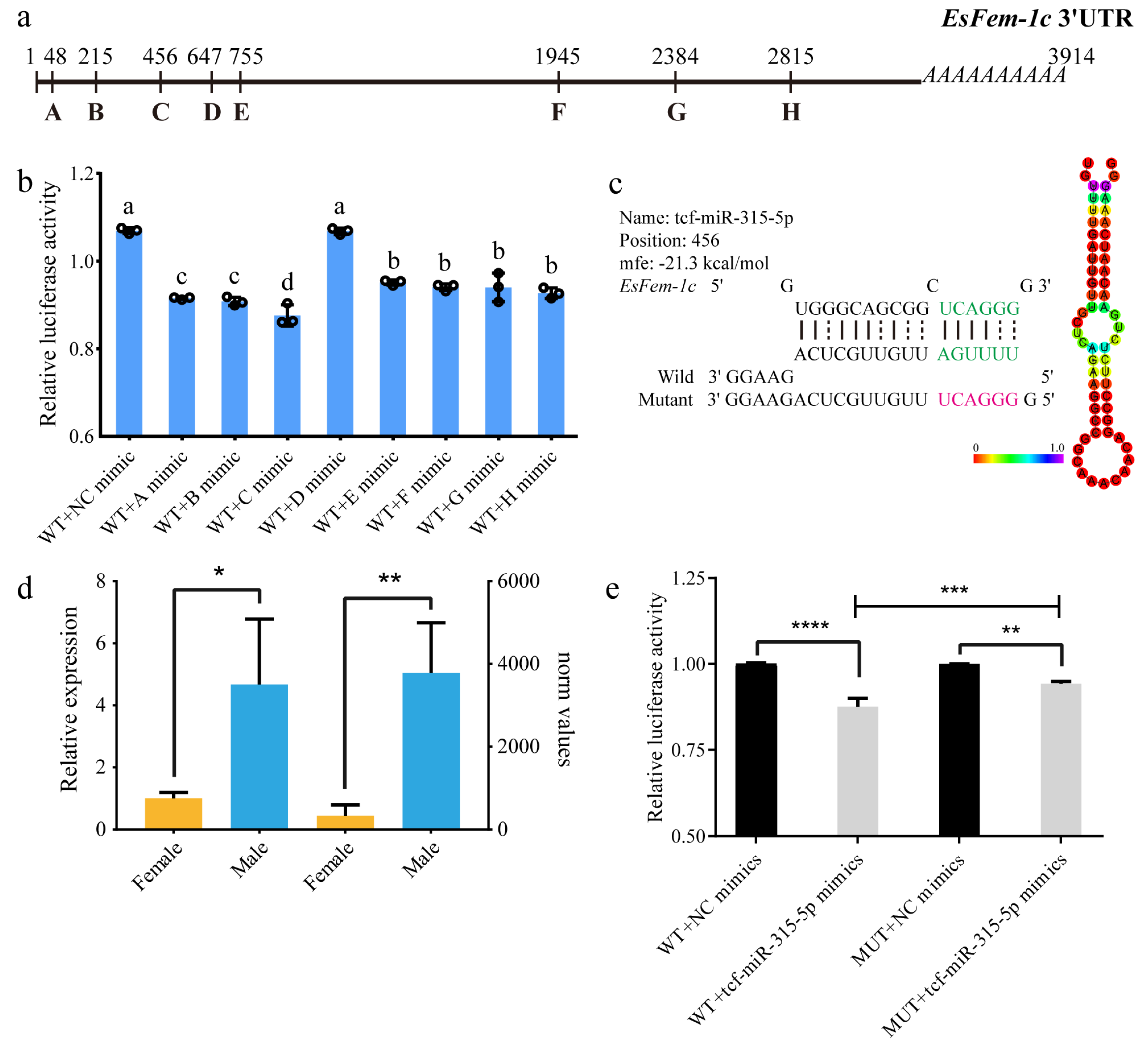
After analysis by using the miRanda, RNA22, and RNAHybird programs, eight miRNAs were predicted to bind to the 3′UTR of EsFem-1c (Figure 7a). The miRNAs including tcf-let-7-3p (A), bmo-mir-6497-p5 (B), tcf-miR-315-5p (C), tcf-miR-7 (D), and tcf-miR-281-3p (E) were found near the termination codon in the 3′UTR sequence. PC-3p-120711 (F), tcf-let-7-5p (G), and tca-bantam-3p (H) were identified near the poly (A) site. The lowest free energy values were observed in bmo-mir-6497-p5 and tcf-let-7-5p (Table S3). In terms of DEMs, female crabs had significantly higher expression of tcf-let-7-3p, bmo-mir-6497-p5, and PC-3p-120711, whereas males had higher expression of tcf-miR-315-5p and tcf-let-7-5p. There was no sex bias expression of tcf-miR-7, tca-bantam-3p, and tcf-miR-281-3p.
To determine whether these eight miRNAs could regulate EsFem-1c, the relative luciferase activity of these miRNAs was measured in vitro using the dual-luciferase reporter assay. While the tcf-miR-7 mimic did not significantly influence luciferase activity, the tcf-let-7-3p, bmo-mir-6497-p5, and tcf-miR-315-5p mimics resulted in an approximate 23%, 26%, and 32% decrease in luciferase activity compared to the NC group (Figure 7b).
The miRNA tcf-miR-315-5p was selected for further analysis and verification, because it significantly reduced luciferase activity. The GC% of tcf-miR-315-5p was 46.70%, and the mfe value was -21.30 kcal/mol (Figure 7c). The analysis of norm values and relative expression revealed that males had significantly higher levels of tcf-miR-315-5p than females (p < 0.05) (Figure 7d). As illustrated in Figure 7e, the results of co-transfecting displayed that the fluorescence intensity of cells treated with Fem1c-WT plasmid and tcf-miR-315-5p mimic was significantly decreased compared with that of the controls (p < 0.05), indicating that tcf-miR-315-5p inhibited the expression of the EsFem-1c gene by targeting its 3′UTR. Co-transfecting with Fem1c-MUT plasmid and tcf-miR-315-5p mimic also significantly decreased the luciferase activity (p < 0.05). However, the luciferase activity in the Fem1c-MUT plasmid group was higher than that in the Fem1c-WT plasmid group (p < 0.05) (Figure 7e).
3.7. The Analysis of the Alternative Splicing Region in 3′UTR of EsFem-1c
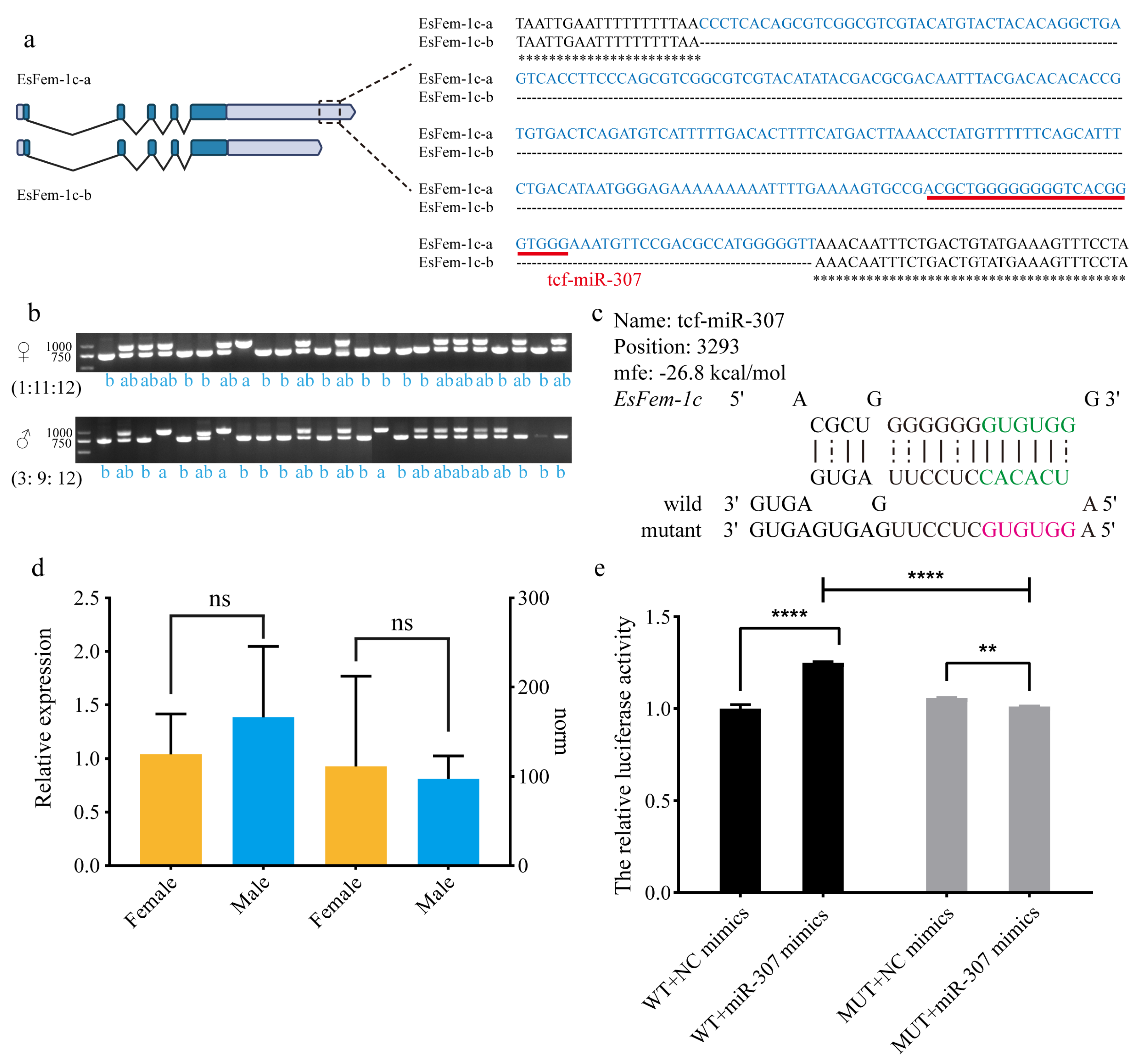
To validate the accuracy of the 3′UTR of EsFem-1c sequence, PCR amplification was performed in the muscle of 48 crabs (female: male = 1:1) with specific primers. There were three types of products: single band at 991 bp (a), single band at 743 bp (b), and double bands at 991 bp and 743 bp (ab). Figure 8b indicates that the ratio of a: ab: b in females differed from that in males. The female was 1:11:12, and the male was 3:9:12, with the female having a low single band at 991 bp. Sanger sequencing of these products was performed to confirm the findings, revealing that ASR-248 existed upstream of the poly (A) site, where tcf-miR-307 was observed (Figure 8a). The mfe of tcf-miR-307 was −26.0 kcal/mol, and the expression pattern revealed no sex bias (Figure 8c,d).
Co-transfection of the Fem1c-WT vector and tcf-miR-307 mimics significantly increased the relative activity of luciferase compared to the NC group (p < 0.05), as revealed in Figure 8e. The luciferase activity of the Fem1c-MUT vector and tcf-miR-307 mimics was significantly lower than that of the NC group (p < 0.05). Moreover, luciferase activity was lower in the Fem1c-MUT plasmid group than in the Fem1c-WT plasmid group (p < 0.05). These findings indicate that tcf-miR-307 could promote ASR-248 function.
4. Discussion
Several previous studies have demonstrated that Fem-1c has a broad expression abundance throughout the entire gonadal developmental process; for example, the expression level of Lmfem-1c gradually increases during testis development in Locusta migratoria manilensis [9]. The highest expression of EsFem-1c exists in the early development stage of gonads in E. sinensis, which is similar to that found in Cherax quadricarinatus, Crassostrea gigas, and Penaeus vannamei [26,53,54], indicating that EsFem-1c is involved in the development of oocyte and spermatogenesis. In contrast, tissue distribution analysis in the present study revealed that the expression level of the EsFem-1c gene in the normal ovary was higher than that in the normal testis, consistent with the result in H. cumingii [2], indicating that EsFem-1c plays a critical role in female gonads. Interestingly, EsFem-1c gene expression in intersex crab muscle is significantly higher than in male and female crabs. We can assume that the EsFem-1c gene has a regulatory pathway that allows excessive expression to move to other muscle tasks, such as molting or muscle growth and breakdown. Furthermore, EsFem-1c has a higher expression level in normal male AG than in intersex AG, whereas MnFem-1b is present in the AG of M. nipponense [12], indicating a potential role in male phenotype.
The length of the penis grows, and EsIAG expression increases in male mitten crabs after knocking down the EsFem-1c. IAG regulates male differentiation in decapods. A similar report indicates that the knocking down of MrIAG negatively regulates MrFem-1 in the post-larvae of M. rosenbergii [55]. Based on the function of EsIAG, these findings suggest that EsFem-1c regulates male sexual development [56]. The present study also confirms the promoting effect of EsFem-1c on female sexual development by regulating EsCFSH-1, which has influenced the sexual characteristics of female crabs [40]. Therefore, EsFem-1c is thought to be an upstream regulator of EsIAG and EsCFSH-1, which regulate sex differentiation in E. sinensis.
GO enrichment and KEGG pathway analyses of the target genes predicted to bind to 43 DEMs revealed that they were significantly enriched in many pathways, including insulin, MAPK, Wnt, TGF, and the oxytocin signaling pathway, all of which have been associated with reproductive function [32]. For example, PC-3p-375 and bmo-miR-306a-5p were predicted to target the glycogen synthase kinase 3 (GSK3) gene in the insulin signaling pathway. tcf-miR-10-5p and PC-3p-241774 targeted C-jun-amino-terminal kinase-interacting protein 4 (JIP-4) and neurofibromin (NF). These genes were the key regulators of the MAPK pathway. The PC-3p-795 targeted Ras-related protein acted on the Wnt pathway. Our results agree with the analysis of miRNAs during larval metamorphic and gonadal development in E. sinensis [31].
Tcf-miR-315-5p expression is abundant in males, as in Pteromalus puparum [57], indicating that tcf-miR-315-5p may regulate male development. In addition, the miR-315 has been studied to act as one subset in Drosophila melanogaster sperm storage and the post-mating responses [58,59], as well as to regulate Nilaparvata lugens sex determination and differentiation [60]. The dual-luciferase reporter assay in the present study yielded a similar conclusion regarding the inhibition function of tcf-miR-315-5p on the EsFem-1c gene.
Furthermore, EsFem-1c expression decreased gradually with the development of the testis and AG. It can be speculated that tcf-miR-315-5p, as a key miRNA, is responsible for the repression of testis and AG development by inhibiting EsFem-1c expression. This is similar to the fact that miRNA let-7d and miR-34 are proven to control the testicular differentiation during gonadal development progress by inhibiting EsIAG gene expression [32].
ASR-248 identified in 3′UTR provides a new understanding of how 3′UTR influences the mRNA expression of EsFem-1c in E. sinensis. There are two theories regarding its function. The first can be used as the NAT. ASR-248 transcribes from 3′UTR, the non-protein-coding region. Considering the various roles of NAT, ASR-248 may influence the function of EsFem-1c by altering mRNA stability, masking miRNA-binding sites, and forming endogenous siRNAs [25,61]. An NAT observed in the Pvfem-1 in P. vannamei suggests a negative post-transcriptional regulation to control female sexual trait differentiation [26]. The second possibility is APA. By forming an APA complex, APA can eventually produce several mRNA polyadenylation isoforms [24]. In this study, ASR-248 caused variations in the transcription products of EsFem-1c, which may influence the positive effect of miRNA tcf-miR-307. We can infer that different types of EsFem-1c transcripts may have different sex differentiation mechanisms, such as the dose effect of functional proteins. A similar mode has been reported on regulating Dsx gene expression in three insects, D. melanogaster [62], Megaselia scalaris, and Anopheles gambiae [63].
5. Conclusions
In conclusion, current research indicates that EsFem-1c is important in regulating sex differentiation and controlling sexual traits. The miRNA tcf-miR-315-5p functions as a suppressed regulator to influence the function of the 3′UTR of EsFem-1c. The miRNA tcf-miR-307 positively affects the EsFem-1c gene by binding to gender-biased ASR-248 in the 3′UTR. The findings of this study should help future research into the sex-related functional mechanism of EsFem-1c in E. sinensis.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ani13111813/s1. Figure S1: The nucleotide bias at first and each position of known miRNAs in female and male crabs; Figure S2: Differential miRNAs (DEMs) expression in E. sinensis female and male reproductive systems; Figure S3: GO and KEGG classification for predicted target genes of DEMs; Table S1: Overview of miRNA-seq data; Table S2: The DE miRNAs between female and male reproductive tissues of E. sinensis; Table S3: The miRNAs targeted to 3′UTR of EsFem-1c.
Author Contributions
D.Z., Z.C. and T.F. designed the study; D.Z., N.M., R.H. and W.L. performed the experiments; D.Z. and T.F. analyzed the data; Z.C. fundraising and supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32072964) and the National Key R and D Program of China (2022YFD2400104-02).
Institutional Review Board Statement
The committee on the Ethics of Animal Experiments of Ningbo University (No. SYXK20190005) was established in 2019, but the committee authorities were only for rabbits, mice, and rats, and not aquatic animals. In this study, mature and juvenile crabs were purchased from commercial farms, and all experimental operations involving animals complied with the requirements of the governing regulation for the use of experimental animals in Zhejiang Province (Zhejiang provincial government order No. 263, released on 17 August 2009, effective from 1 October 2010) and were performed according to the Experimental Animal Management Law of China and met the animal operation standards of the industry.
Informed Consent Statement
Not applicable.
Data Availability Statement
The miRNAseq data have been submitted to the BioProject under the accession number PRJNA942040. Upon reasonable request, the datasets used and analyzed during the present study are also available from the corresponding author.
Acknowledgments
We would like to thank the anonymous reviewers for their helpful remarks and suggestions.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Starostina, N.G.; Lim, J.M.; Schvarzstein, M.; Wells, L.; Spence, A.M.; Kipreos, E.T. A CUL-2 ubiquitin ligase containing three FEM proteins degrades TRA-1 to regulate C. elegans sex determination. Dev. Cell. 2007, 13, 127–139. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, C.; Guo, P.; Wang, G.; Li, J. Molecular characterization and expression of the feminization-1c (fem-1c) in the freshwater mussel (Hyriopsis cumingii). Aquac. Fish. 2018, 3, 6–13. [Google Scholar] [CrossRef]
- Zanetti, S.; Puoti, A. Sex Determination in the Caenorhabditis elegans Germline. In Germ Cell Development in C. elegans; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2013; pp. 41–69. [Google Scholar] [CrossRef]
- Chen, X.; Liao, S.; Makaros, Y.; Guo, Q.; Zhu, Z.; Krizelman, R.; Dahan, K.; Tu, X.; Yao, X.; Koren, I.; et al. Molecular basis for arginine C-terminal degron recognition by Cul2(FEM1) E3 ligase. Nat. Chem. Biol. 2021, 17, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Kimble, J.; Edgar, L.; Hirsh, D. Specification of male development in Caenorhabditis elegans the fem genes. Dev. Biol. 1984, 105, 234–239. [Google Scholar] [CrossRef]
- Doniach, T.; Hodgkin, J. A sex-determining gene, fem-1, required for both male and hermaphrodite development in Caenorhabditis elegans. Dev. Biol. 1984, 106, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Thies, M.; Berke, B. A role for the Fem-1 gene of Drosophila melanogaster in adult courtship. bioRxiv 2020, 19, 911693. [Google Scholar] [CrossRef]
- Ventura-Holman, T.; Lu, D.; Si, X.; Izevbigie, E.B.; Maher, J.F. The Fem1c genes: Conserved members of the Fem1 gene family in vertebrates. Gene 2003, 314, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; You-Jin, H.; Bin, C.; Feng-Ling, S.; Peng, W.; Zheng-Bo, H. Cloning and expression analysis of fem-1 genes from the oriental migratory locust, Locusta migratoria manilensis (Orthoptera: Locustidae). Acta Entomol. Sin. 2013, 56, 729–737. [Google Scholar] [CrossRef]
- Shi, J.-T.; Li, Z.; Gui, J.-F.; Zhou, L. The cloning and expression analysis of zebrafish Fem-1c, a member of Fem-1 family. Ata Hydrobiol. Sin. 2015, 39, 459–467. [Google Scholar] [CrossRef]
- Du, H.-N.; Hu, H.-Y. Ankyrin Repeat Mediated Protein-protein Interaction. Prog. Biochem. Biophys. 2002, 29, 6–9. [Google Scholar] [CrossRef]
- Rahman, N.M.; Fu, H.; Qiao, H.; Jin, S.; Bai, H.; Zhang, W.; Jiang, F.W.; Liang, G.; Sun, S.; Gong, Y.; et al. Molecular cloning and expression analysis of Fem1b from oriental river prawn Macrobrachium nipponense. Genet. Mol. Res. 2016, 15, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.-Y.; Liu, Z.-Q.; Lin, J.-Y.; Li, J.-L.; Qiu, G.-F. Molecular characterization of a novel ovary-specific gene fem-1 homolog from the oriental river prawn, Macrobrachium nipponense. Gene 2016, 575, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.-X.; Liu, X.; Ye, B.-Q.; Liu, Y.; Tan, S.-P.; Ma, K.-Y.; Qiu, G.-F. Molecular characterization of ovary-specific gene Mrfem-1 and siRNA-mediated regulation on targeting Mrfem-1 in the giant freshwater prawn, Macrobrachium rosenbergii. Gene 2020, 754, 144891. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Cui, Z.; Hui, M.; Liu, Y.; Li, Y. Molecular characterization and expression profile of three Fem-1 genes in Eriocheir sinensis provide a new insight into crab sex-determining mechanism. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2015, 189, 6–14. [Google Scholar] [CrossRef]
- Mayr, C. Regulation by 3′-Untranslated Regions. Annu. Rev. Genet. 2017, 51, 171–194. [Google Scholar] [CrossRef]
- Schneider, A.B.; Wolfinger, M.T. Musashi binding elements in Zika and related Flavivirus 3′UTRs: A comparative study in silico. Sci. Rep. 2019, 9, 6911. [Google Scholar] [CrossRef]
- Lai, E.C.; Burks, C.; Posakony, J.W. The K box, a conserved 3′ UTR sequence motif, negatively regulates accumulation of enhancer of split complex transcripts. Development 1998, 125, 4077–4088. [Google Scholar] [CrossRef]
- Bae, B.; Miura, P. Emerging Roles for 3′ UTRs in Neurons. Int. J. Mol. Sci. 2020, 21, 3413. [Google Scholar] [CrossRef]
- Frederick, P.M.; Simard, M.J. Regulation and different functions of the animal microRNA-induced silencing complex. Wiley Interdiscip. Rev. RNA 2022, 13, e1701. [Google Scholar] [CrossRef]
- Mayr, C. What Are 3′ UTRs Doing? Cold Spring Harb. Perspect. Biol. 2019, 11, a034728. [Google Scholar] [CrossRef]
- Sadek, J.; Omer, A.; Hall, D.; Ashour, K.; Gallouzi, I.E. Alternative polyadenylation and the stress response. Wiley Interdiscip. Rev. RNA 2019, 10, e1540. [Google Scholar] [CrossRef]
- Garaulet, D.L.; Zhang, B.; Wei, L.; Li, E.; Lai, E.C. miRNAs and Neural Alternative Polyadenylation Specify the Virgin Behavioral State. Dev. Cell. 2020, 54, 410–423. [Google Scholar] [CrossRef]
- Ren, F.; Zhang, N.; Zhang, L.; Miller, E.; Pu, J.J. Alternative Polyadenylation: A new frontier in post transcriptional regulation. Biomark. Res. 2020, 8, 67. [Google Scholar] [CrossRef]
- Faghihi, M.A.; Wahlestedt, C. Regulatory roles of natural antisense transcripts. Nat. Rev. Mol. Cell. Biol. 2009, 10, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Torres, P.; Ventura-Lopez, C.; Llera-Herrera, R.; Ibarra, A.M. A natural antisense transcript of the fem-1 gene was found expressed in female gonads during the characterization, expression profile, and cellular localization of the fem-1 gene in Pacific white shrimp Penaeus vannamei. Gene 2019, 706, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.-N.; Shi, L.-L.; Liu, Z.-Q.; Qiu, G.-F. Global analysis of the ovarian microRNA transcriptome: Implication for miR-2 and miR-133 regulation of oocyte meiosis in the Chinese mitten crab, Eriocheir sinensis (Crustacea: Decapoda). BMC Genom. 2014, 15, 547. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Tang, Y.; Sun, L.; Li, S.; Liu, H.; Chen, Z.; Li, G. SAP30 targeted by miR-133b was involved in the process of nuclear decondensation in Chinese mitten crab (Eriocheir sinensis) sperm. Aquac. Rep. 2023, 29, 101540. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, W.; Xu, Z.; Pan, J.; Zhao, Z.; Ren, Q. Eriocheir Sinensis microRNA-7 targets crab Myd88 to enhance white spot syndrome virus replication. Fish Shellfish Immunol. 2018, 79, 274–283. [Google Scholar] [CrossRef]
- He, L.; Wang, Y.-L.; Li, Q.; Yang, H.-D.; Duan, Z.-L.; Wang, Q. Profiling microRNAs in the testis during sexual maturation stages in Eriocheir sinensis. Anim. Reprod. Sci. 2015, 162, 52–61. [Google Scholar] [CrossRef]
- Chen, M.; Yue, Y.; He, J.; Li, P.; Yan, J.; Wang, F.; Cai, F.; Zhou, K. Screening and identification of microRNAs during larval metamorphic development of Chinese mitten crab Eriocheir sinensis. Aquac. Res. 2020, 51, 2322–2335. [Google Scholar] [CrossRef]
- Luo, B.-Y.; Xiong, X.-Y.; Liu, X.; He, X.Y.; Qiu, G.-F. Identification and characterization of sex-biased and differentially expressed miRNAs in gonadal developments of the Chinese mitten crab, Eriocheir sinensis. Mol. Reprod. Dev. 2021, 88, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; He, L. Histological and cytological observation on the development cycle of crab (Eriocheir sinensis) ovary. Oceanol. Et Limnol. Sin. 1997, 28, 138–145. [Google Scholar] [CrossRef]
- Wang, Y.L.; Sun, W.J.; He, L.; Li, Q.; Wang, Q. Morphological alterations of all stages of spermatogenesis and acrosome reaction in Chinese mitten crab Eriocheir sinensis. Cell Tissue Res. 2015, 360, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Qiu, G.; Wu, P.; Yundong, L. Structure and function of the androgenic gland in Eriocheir sinensis. J. Fish. China 2000, 24, 99–104. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST A tool to design target-specific. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Alzohairy, A.M. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar] [CrossRef]
- Yalamanchili, H.K.; Alcott, C.E.; Ji, P.; Wagner, E.J.; Zoghbi, H.Y.; Liu, Z. PolyA-miner: Accurate assessment of differential alternative poly-adenylation from 3′ Seq data using vector projections and non-negative matrix factorization. Nucleic Acids Res. 2020, 48, e69. [Google Scholar] [CrossRef]
- Grillo, G.; Turi, A.; Licciulli, F.; Mignone, F.; Liuni, S.; Banfi, S.; Gennarino, V.A.; Horner, D.S.; Pavesi, G.; Picardi, E. UTRdb and UTRsite (RELEASE 2010): A collection of sequences and regulatory motifs of the untranslated regions of eukaryotic mRNAs. Nucleic Acids Res. 2010, 38, D75–D80. [Google Scholar] [CrossRef]
- Zhu, D.; Feng, T.; Mo, N.; Han, R.; Lu, W.; Shao, S.; Cui, Z. New insights for the regulatory feedback loop between type 1 crustacean female sex hormone (CFSH-1) and insulin-like androgenic gland hormone (IAG) in the Chinese mitten crab (Eriocheir sinensis). Front. Physiol. 2022, 13, 1054773. [Google Scholar] [CrossRef]
- Cui, Z.; Liu, Y.; Yuan, J.; Zhang, X.; Ventura, T.; Ma, K.-Y.; Sun, S.; Song, C.; Zhan, D.; Yang, Y.; et al. The Chinese mitten crab genome provides insights into adaptive plasticity and developmental regulation. Nat. Commun. 2021, 12, 2395. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shahid, M.Q.; Wu, J.; Wang, L.; Liu, X.; Lu, Y. Comparative Small RNA Analysis of Pollen Development in Autotetraploid and Diploid Rice. Int. J. Mol. Sci. 2016, 17, 499. [Google Scholar] [CrossRef]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human MicroRNA targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T. Gene Ontology tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef]
- Miranda, K.C.; Huynh, T.; Tay, Y.; Ang, Y.S.; Tam, W.L.; Thomson, A.M.; Lim, B.; Rigoutsos, I. A pattern-based method for the identification of MicroRNA binding sites and their corresponding heteroduplexes. Cell 2006, 126, 1203–1217. [Google Scholar] [CrossRef]
- Rehmsmeier, M.; Steffen, P.; Hochsmann, M.; Giegerich, R. Fast and effective prediction of microRNA/target duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef]
- Zhang, D.; Shi, B.; Shao, P.; Shao, C.; Wang, C.; Li, J.; Liu, X.; Ma, X.; Zhao, X. The identification of miRNAs that regulate ovarian maturation in Cynoglossus semilaevis. Aquaculture 2022, 555, 738250. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Stoline, M.R. The Status of Multiple Comparisons Simultaneous Estimation of All Pairwise Comparisons in One-Way ANOVA Designs. Am. Stat. 1981, 35, 134–141. [Google Scholar] [CrossRef]
- Zhou, Z.-Y.; Qi, L.; Hong, Y.; Lingfeng, K. Cloning and Expression analysis of Fem-1 gene of Pacific Oyster (Crassostrea gigas). Period. Ocean Univ. China 2018, 48, 45–54. [Google Scholar] [CrossRef]
- Zheng, J.; Chen, L.; Jia, Y.; Chi, M.; Li, F.; Cheng, S.; Liu, S.; Liu, Y.; Gu, Z. Genomic structure, expression, and functional characterization of the Fem-1 gene family in the redclaw crayfish, Cherax quadricarinatus. Gen. Comp. Endocrinol. 2022, 316, 113961. [Google Scholar] [CrossRef]
- Qian, H.; Ma, K.; Feng, J.; Guo, Z.; Gong, J.; Chen, H.; Bai, H.; Qiu, G. Transcriptome analysis of the post-larvae of giant freshwater prawn (Macrobrachium rosenbergii) after IAG gene knockdown with microRNA interference. General Comp. Endocrinol. 2022, 325, 114054. [Google Scholar] [CrossRef]
- Fu, C.; Li, F.; Wang, L.; Wu, F.; Wang, J.; Fan, X.; Liu, T. Molecular characteristics and abundance of insulin-like androgenic gland hormone and effects of RNA interference in Eriocheir sinensis. Anim. Reprod. Sci. 2020, 215, 106332. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Wang, B.; Li, K.; Xiong, S.; Ye, X.; Wang, J.; Zhang, J.; Yan, Z.; Wang, F.; Song, Q.; et al. Identification and characterization of miRNAs in an endoparasitoid wasp, Pteromalus puparum. Arch. Insect Biochem. Physiol. 2020, 103, e21633. [Google Scholar] [CrossRef] [PubMed]
- Mohorianu, I.; Fowler, E.K.; Dalmay, T.; Chapman, T. Control of seminal fluid protein expression via regulatory hubs in Drosophila melanogaster. Proc. Biol. Sci. 2018, 285, 20181681. [Google Scholar] [CrossRef] [PubMed]
- Laudanski, K.; Soh, J.; DiMeglio, M.; Sullivan, K.E. Prolonged Transcriptional Consequences in Survivors of Sepsis. Int. J. Mol. Sci. 2021, 22, 5422. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, J.; Zhan, A.; Wang, Y.; Ma, X.; Jie, W.; Cao, Z.; Omar, M.A.A.; He, K.; Li, F. Identification and Analysis of MicroRNAs Associated with Wing Polyphenism in the Brown Planthopper, Nilaparvata lugens. Int. J. Mol. Sci. 2020, 21, 9754. [Google Scholar] [CrossRef]
- Faghihi, M.A.; Zhang, M.; Huang, J.; Modarresi, F.; Van der Brug, M.P.; Nalls, M.A.; Cookson, M.R.; St-Laurent, G., 3rd; Wahlestedt, C. Evidence for natural antisense transcript-mediated inhibition of microRNA function. Genome Biol. 2010, 11, R56. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Baker, B.S. Regulation of sex-specific RNA splicing at the Drosophila doublesex gene: Cis-acting mutations in exon sequences alter sex-specific RNA splicing patterns. Genes Dev. 1990, 4, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Kobayashi, K.; Watanabe, H.; Iguchi, T. Environmental Sex Determination in the Branchiopod Crustacean Daphnia magna: Deep Conservation of a Doublesex gene in the Sex-Determining Pathway. PLoS Genet. 2011, 7, e1001345. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Nucleotide characteristics and deduced amino acid sequence of EsFem-1c. The EsFem-1c gene had five exons and four introns in its entire genomic structure. The green blanks enclose the five exons. The open reading frame (ORF) is the region between the initiation codon (highlighted in green) and the termination codon (highlighted in red, the amino acid is indicated by one asterisk), and the eight ANK motifs are highlighted in grey. The polyadenylation signal (PAS) is presented in red at the end of the 3′UTR. The K-box motif regions are enclosed in the yellow frame rectangle, while the Musashi binding element (MBE) motifs are in the blue frame rectangle.
Figure 1.
Nucleotide characteristics and deduced amino acid sequence of EsFem-1c. The EsFem-1c gene had five exons and four introns in its entire genomic structure. The green blanks enclose the five exons. The open reading frame (ORF) is the region between the initiation codon (highlighted in green) and the termination codon (highlighted in red, the amino acid is indicated by one asterisk), and the eight ANK motifs are highlighted in grey. The polyadenylation signal (PAS) is presented in red at the end of the 3′UTR. The K-box motif regions are enclosed in the yellow frame rectangle, while the Musashi binding element (MBE) motifs are in the blue frame rectangle.

Figure 2.
EsFem-1c tissue expression profiles in normal and intersex mitten crabs. This experiment used β-actin mRNA expression levels as an internal reference. F-M, M-M, and I-M represent the muscle of females, males, and intersex, respectively; M-AG and I-AG represent the androgenic gland of males and intersex, respectively; F-O represents normal ovary of females; M-T represents normal testis of males; I-O-like represents ovary-like tissue of intersex. Vertical bars represent the mean ± standard error of the mean (SEM) (n = 4). Asterisks and different lower-case letters indicated significant differences at p < 0.05.
Figure 2.
EsFem-1c tissue expression profiles in normal and intersex mitten crabs. This experiment used β-actin mRNA expression levels as an internal reference. F-M, M-M, and I-M represent the muscle of females, males, and intersex, respectively; M-AG and I-AG represent the androgenic gland of males and intersex, respectively; F-O represents normal ovary of females; M-T represents normal testis of males; I-O-like represents ovary-like tissue of intersex. Vertical bars represent the mean ± standard error of the mean (SEM) (n = 4). Asterisks and different lower-case letters indicated significant differences at p < 0.05.

Figure 3.
The analysis after injecting with the dsRNA EsFem-1c. (a,b) The altered characteristics of female gonopores and male penises in juvenile crabs. (c) A comparison of penis length in normal crabs treated with dsRNA EsFem-1c in juvenile Ⅲ stage. BC, blank control group. Data are presented as the mean ± SEM of separated individuals (n = 6). Asterisks indicate significant differences at p < 0.05. (d,e) Silencing efficiency of EsFem-1c and the expression changes of EsCFSH-1 gene in female crab eyestalk ganglion (EG) and ovary. (f,g) Silencing efficiency of EsFem-1c and the expression changes of EsIAG gene in male crab androgenic gland (AG) and testis. Data are presented as the mean ± SEM of separated individuals (n = 6). For the expression of different genes in each group, different lower-case letters indicated significant differences at p < 0.05.
Figure 3.
The analysis after injecting with the dsRNA EsFem-1c. (a,b) The altered characteristics of female gonopores and male penises in juvenile crabs. (c) A comparison of penis length in normal crabs treated with dsRNA EsFem-1c in juvenile Ⅲ stage. BC, blank control group. Data are presented as the mean ± SEM of separated individuals (n = 6). Asterisks indicate significant differences at p < 0.05. (d,e) Silencing efficiency of EsFem-1c and the expression changes of EsCFSH-1 gene in female crab eyestalk ganglion (EG) and ovary. (f,g) Silencing efficiency of EsFem-1c and the expression changes of EsIAG gene in male crab androgenic gland (AG) and testis. Data are presented as the mean ± SEM of separated individuals (n = 6). For the expression of different genes in each group, different lower-case letters indicated significant differences at p < 0.05.

Figure 4.
The expression of EsFem-1c in the reproduction tissues during developmental stages. (a) EsFem-1c expression level in the ovaries at stages Ⅱ, Ⅲ, and Ⅳ. (b) EsFem-1c expression level in the testis at the spermatocyte, spermatid, and sperm stages. (c) EsFem-1c expression level in AGs at the proliferation, synthesis, and secretion stages. Data are presented as the mean ± SEM of seperated individuals (n = 4). Statistical significance was accepted at p < 0.05 and indicated by different lower-case letters.
Figure 4.
The expression of EsFem-1c in the reproduction tissues during developmental stages. (a) EsFem-1c expression level in the ovaries at stages Ⅱ, Ⅲ, and Ⅳ. (b) EsFem-1c expression level in the testis at the spermatocyte, spermatid, and sperm stages. (c) EsFem-1c expression level in AGs at the proliferation, synthesis, and secretion stages. Data are presented as the mean ± SEM of seperated individuals (n = 4). Statistical significance was accepted at p < 0.05 and indicated by different lower-case letters.

Figure 5.
Basic information on six small RNA libraries of E. sinensis. (a) Female and male mitten crab reproduction systems. The female crab had ovary (O) and spermathecal (SA). The testis (T), vas deferens (VD), seminal vesicle (SV), accessory sex glands (ASG), ejaculatory duct (ED), androgenic gland (AG), and penis (PS) were all present in male crabs. (b) Sequence length distribution of six sRNA libraries. ES_F1, ES_F2, and ES_F3 were the three female libraries, while ES_M1, ES_M2, and ES_M3 were the three male libraries.
Figure 5.
Basic information on six small RNA libraries of E. sinensis. (a) Female and male mitten crab reproduction systems. The female crab had ovary (O) and spermathecal (SA). The testis (T), vas deferens (VD), seminal vesicle (SV), accessory sex glands (ASG), ejaculatory duct (ED), androgenic gland (AG), and penis (PS) were all present in male crabs. (b) Sequence length distribution of six sRNA libraries. ES_F1, ES_F2, and ES_F3 were the three female libraries, while ES_M1, ES_M2, and ES_M3 were the three male libraries.

Figure 6.
The relative expression level of 16 expressed miRNAs detected by qRT-PCR (left) and norm values derived from sequencing data (right). (a–h) The up-regulated group; the expression level of these eight miRNAs were higher in females than in males. (i–p) The down-regulated group; the expression level of these eight miRNAs were higher in males than in females. The norm values are normalized from miRNA read counts to count the miRNA expression. The values are presented as means ± SEM (n = 3). Statistical significance was accepted at p < 0.05 and marked by different asterisks.
Figure 6.
The relative expression level of 16 expressed miRNAs detected by qRT-PCR (left) and norm values derived from sequencing data (right). (a–h) The up-regulated group; the expression level of these eight miRNAs were higher in females than in males. (i–p) The down-regulated group; the expression level of these eight miRNAs were higher in males than in females. The norm values are normalized from miRNA read counts to count the miRNA expression. The values are presented as means ± SEM (n = 3). Statistical significance was accepted at p < 0.05 and marked by different asterisks.

Figure 7.
An analysis of the miRNAs predicted to target the 3′UTR of EsFem-1c gene. (a) Software-detected predicted binding position of eight miRNAs in the 3′UTR of EsFem-1c. (b) The relative luciferase activities of miRNAs tested by the dual-luciferase assay in HEK293T cells. A, tcf-let-7-3p; B, bmo-mir-6497-p5; C, tcf-miR-315-5p; D, tcf-miR-7; E, tcf-miR-281-3p; F, PC-3p-120711; G, tcf-let-7-5p; H, tca-bantam-3p. (c) The tcf-miR-315-5p miRNA sequence and secondary structure. (d) qRT-PCR analysis and norm values of the miRNA tcf-miR-315-5p revealed that this miRNA was highly expressed in males E. sinensis. (e) The relative luciferase activities in the wild and mutant assay. Values represent mean ± SEM of independent biological replicates (n = 3). The lower-case letters and asterisks indicate statistically significant differences at p < 0.05.
Figure 7.
An analysis of the miRNAs predicted to target the 3′UTR of EsFem-1c gene. (a) Software-detected predicted binding position of eight miRNAs in the 3′UTR of EsFem-1c. (b) The relative luciferase activities of miRNAs tested by the dual-luciferase assay in HEK293T cells. A, tcf-let-7-3p; B, bmo-mir-6497-p5; C, tcf-miR-315-5p; D, tcf-miR-7; E, tcf-miR-281-3p; F, PC-3p-120711; G, tcf-let-7-5p; H, tca-bantam-3p. (c) The tcf-miR-315-5p miRNA sequence and secondary structure. (d) qRT-PCR analysis and norm values of the miRNA tcf-miR-315-5p revealed that this miRNA was highly expressed in males E. sinensis. (e) The relative luciferase activities in the wild and mutant assay. Values represent mean ± SEM of independent biological replicates (n = 3). The lower-case letters and asterisks indicate statistically significant differences at p < 0.05.

Figure 8.
Analysis of the 248 bp region in the 3′UTR of EsFem-1c. (a) Software-detected the 248 bp sequence and binding sites of miRNA tcf-miR-307 in the 3′UTR of EsFem-1c. (b) PCR results of male and female crabs revealed three types of products: a single at 991 bp (a), a single at 743 bp (b), and a double at 991 bp and 743 bp (ab). (c) Information on the miRNA sequence and secondary structure of the miRNA tcf-miR-307. (d) qRT-PCR analysis and norm values of the tcf-miR-307 revealed that miRNA had no sex bias between the female and male E. sinensis. (e) The relative luciferase activities in the wild and mutant assay. The data are presented as mean ± SEM (n = 3). Statistical significance was accepted at p < 0.05 and indicated by different asterisks.
Figure 8.
Analysis of the 248 bp region in the 3′UTR of EsFem-1c. (a) Software-detected the 248 bp sequence and binding sites of miRNA tcf-miR-307 in the 3′UTR of EsFem-1c. (b) PCR results of male and female crabs revealed three types of products: a single at 991 bp (a), a single at 743 bp (b), and a double at 991 bp and 743 bp (ab). (c) Information on the miRNA sequence and secondary structure of the miRNA tcf-miR-307. (d) qRT-PCR analysis and norm values of the tcf-miR-307 revealed that miRNA had no sex bias between the female and male E. sinensis. (e) The relative luciferase activities in the wild and mutant assay. The data are presented as mean ± SEM (n = 3). Statistical significance was accepted at p < 0.05 and indicated by different asterisks.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primer sequences used to clone and synthesize dsRNA of target genes.
| Primer | Sequence (5′–3′) | Applications |
|---|---|---|
| EsFem-1c-F1 | CCAGTCATGAAGAGCAGCC | Gene cloning |
| EsFem-1c-R1 | ATGGCCCAGAAGTGCTTG | |
| EsFem-1c-248-F2 | CAGACTGCTGGGGCTTACAA | |
| EsFem-1c-248-R2 | TTCATACATACCGGCCAGCC | |
| EsFem-1c-dsF | AGATCCAGACGAGATGCGAATG | Synthesis of dsRNA |
| EsFem-1c-dsR | CCGTCCTTCACCGCCAC | |
| EGFP-dsF | CACAAGTTCAGCGTGTCCG | |
| EGFP-dsR | AACCACTACCTGAGCACCCA | |
| Primer-T7 | TAATACGACTCACTATAGGG | |
| Primer-SP6 | ATTTAGGTGACACTATAG |
Table 2.
Primer sequences used for qRT-PCR.
| Primer | Sequence (5′–3′) | Applications |
|---|---|---|
| EsFem-1c-qF | CTCAGTCCTGTTCCCTGCATT | qRT-PCR for mRNA |
| EsFem-1c-qR | AGGGCTGGCCGGTATGTAT | |
| Es-β-actin-qF | GCATCCACGAGACCACTTACA | |
| Es-β-actin-qR | CTCCTGCTTGCTGATCCACATC | |
| EsCFSH-1-qF | ATACGTTGAGCGCCAGATCC | |
| EsCFSH-1-qR | CAGAGCCACACATACGGAGC | |
| EsIAG-qF | GCTCCTACAAGCAGCACCC | |
| EsIAG-qR | AGGGTCTTCCAGATGGATCG | |
| tca-bantam-3p | TGAGATCATTGTGAAAGCTGATT | qRT-PCR for miRNAs |
| tcf-miR-281-3p | CTGTCATGGAGTTGCTCTCTTT | |
| tcf-miR-307 | TCACAACCTCCTTGAGTGAGT | |
| tcf-miR-7 | TGGAAGACTAGTGATTTTGTTGTT | |
| bmo-mir-6497-p5 | TCGGGATAAGGATTGGCTC | |
| tcf-miR-315-5p | TTTTGATTGTTGCTCAGAAGG | |
| tcf-let-7-5p | TGAGGTAGTAGGTTGTGTGGTT | |
| tcf-let-7-3p | CTGTACAACTTGCTAACTTTCC | |
| PC-3p-120711 | TGTGGTTGAGCAAAAAGGG | |
| PC-5p-16060 | TCGATCCCCGGCACCTCCA | |
| PC-3p-74 | TGACTAGAGATTCACACTCAT | |
| tcf-miR-278_R | TCGGTGGGATTCTCGTCCG | |
| mja-mir-6489-p5 | CGGACTGGCGCTCTTGGA | |
| tcf-miR-10-5p | TACCCTGTAGATCCGAATTTG | |
| PC-5p-21023 | GGTGGAAAGAGATTCAGTCG | |
| PC-5p-17859 | GATGGGTGTGTCTCTGGTGC | |
| PC-3p-1739 | TAGCACCATGTGAATTCAGTAC | |
| bmo-miR-306a-5p | TCAGGTACTGTGTGACTCTG | |
| tcf-miR-9a-5p | TCTTTGGTGATCTAGCTGTATG | |
| tcf-miR-279b | TGACTAGATCCATACTCATCT | |
| U6-R | AACGCTTCACGAATTTGCGT |
Table 3.
Primer sequences for constructing wild and mutant plasmids of EsFem-1c 3′UTR.
| Primer | Sequence (5′–3′) | Applications |
|---|---|---|
| Fem-1c-miF | GGAGCTCTAACCCTGGAGACAGGGATGAA | Wild plasmids |
| Fem-1c-miR | CTAGCTAGCCTCGCTGGACAGACAAGATGG | |
| Fem-1c-248-miF | GGAGCTCGTTGGACAGAGGTTGCTGTTTT | |
| Fem-1c-248-miR | CTAGCTAGCGGCCCAGAAGTGCTTGATGTT | |
| Fem-1c-MF1 | TGAGGGGGGACTAAATGTCATTTCAATTTTGTGTTGC | Mutant plasmids |
| Fem-1c-MR1 | CATTTAGTCCCCCCTCAAACCTCTCAACAATCACAT | |
| Fem-1c-MF2 | AAAAGTGCCGGGTGTGACGCTGGGGGGGGTCACG | |
| Fem-1c-MR2 | GTCACACCCGGCACTTTTCAAAATTTTTTTTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhu, D.; Feng, T.; Mo, N.; Han, R.; Lu, W.; Cui, Z. Eriocheir sinensis feminization-1c (Fem-1c) and Its Predicted miRNAs Involved in Sexual Development and Regulation. Animals 2023, 13, 1813. https://doi.org/10.3390/ani13111813
AMA Style
Zhu D, Feng T, Mo N, Han R, Lu W, Cui Z. Eriocheir sinensis feminization-1c (Fem-1c) and Its Predicted miRNAs Involved in Sexual Development and Regulation. Animals. 2023; 13(11):1813. https://doi.org/10.3390/ani13111813
Chicago/Turabian StyleZhu, Dandan, Tianyi Feng, Nan Mo, Rui Han, Wentao Lu, and Zhaoxia Cui. 2023. "Eriocheir sinensis feminization-1c (Fem-1c) and Its Predicted miRNAs Involved in Sexual Development and Regulation" Animals 13, no. 11: 1813. https://doi.org/10.3390/ani13111813
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.