
The Influence of Age, Sex and Season on Andean Condor Ranging Behavior during the Immature Stage
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Species
2.3. Bird Tagging
2.4. Data Analysis
2.4.1. Data Processing
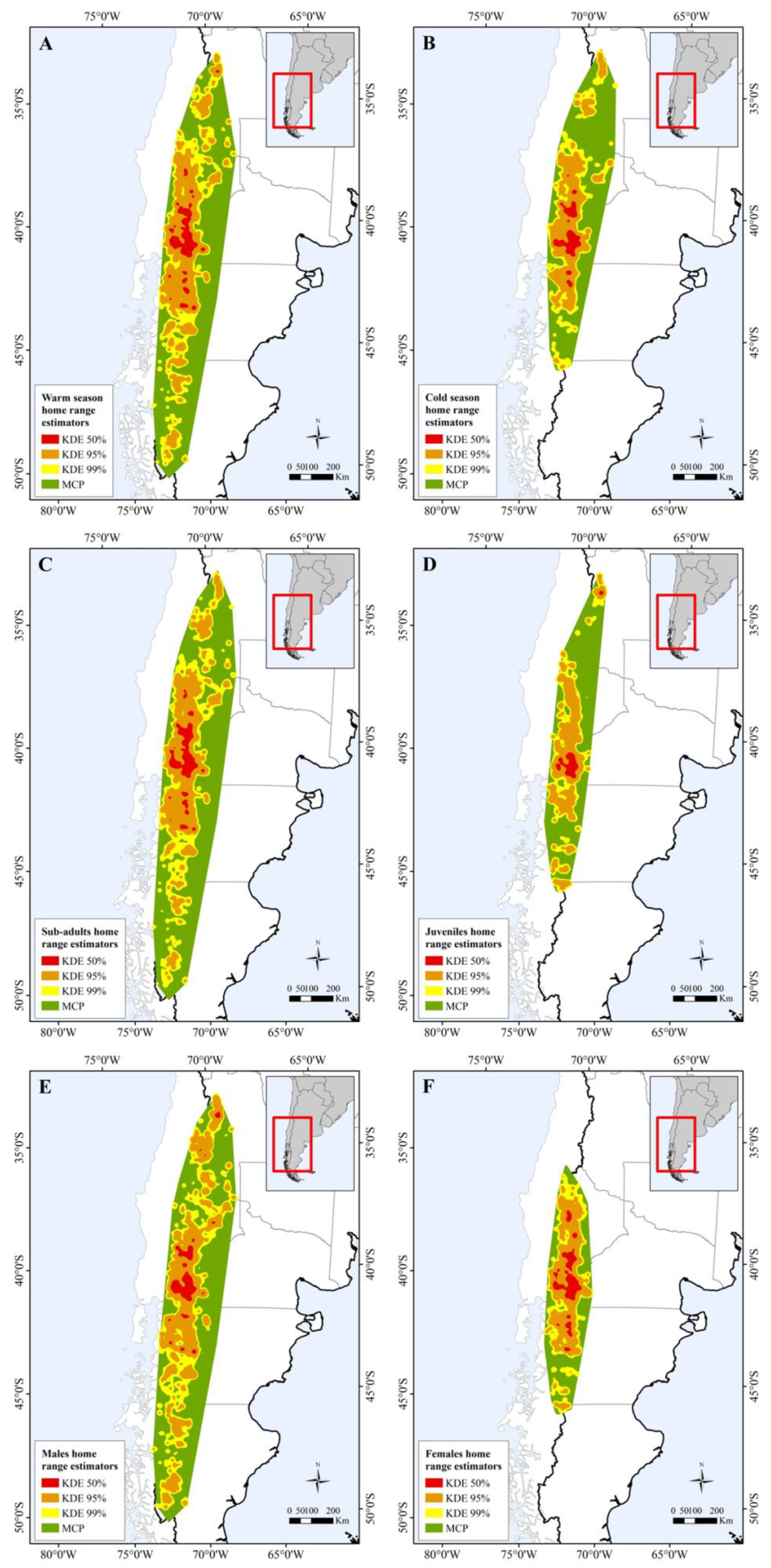
2.4.2. Home Range Size
2.4.3. Flight Distances
2.5. Statistical Analysis
3. Results
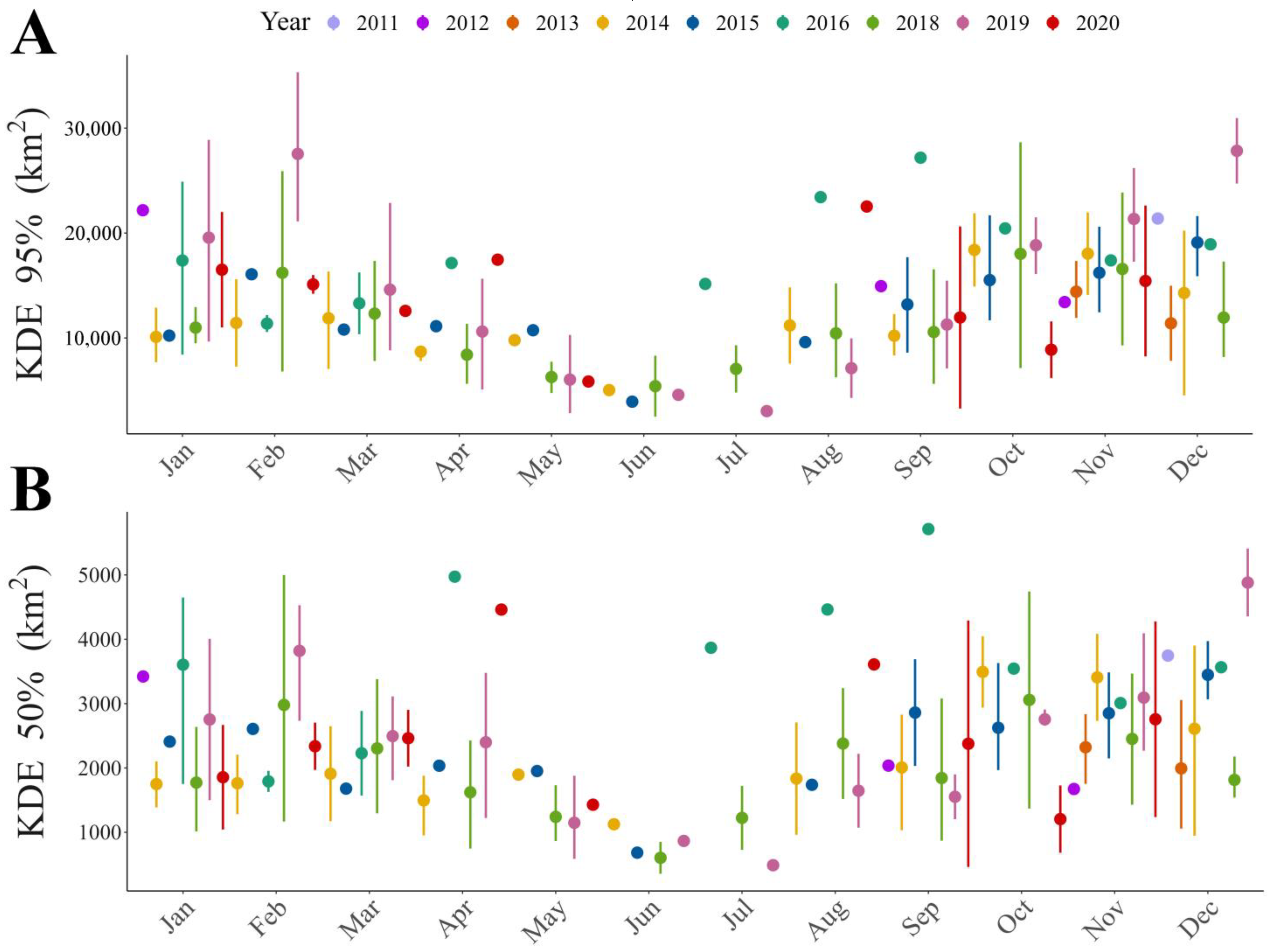
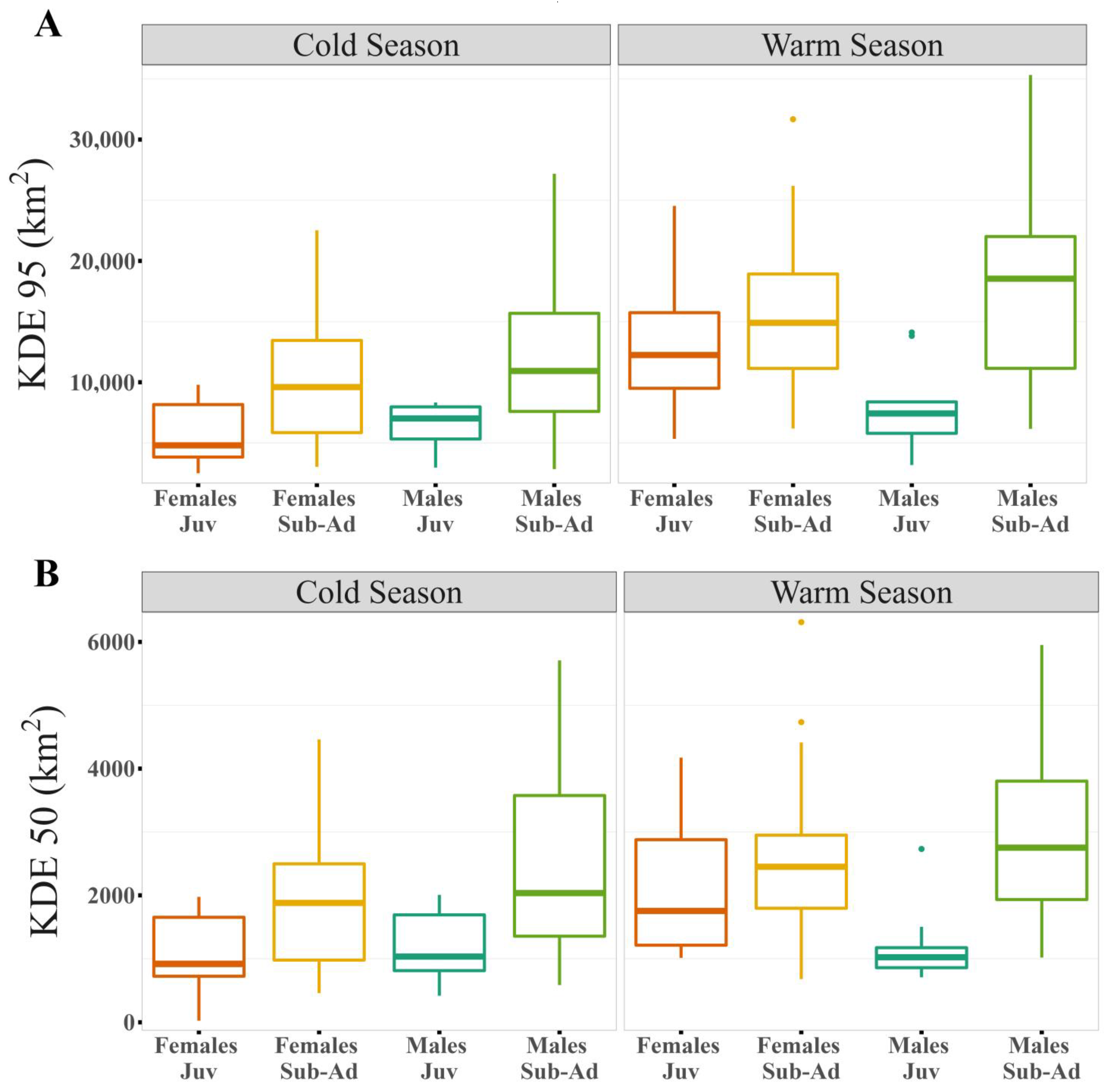
3.1. Home Range Size
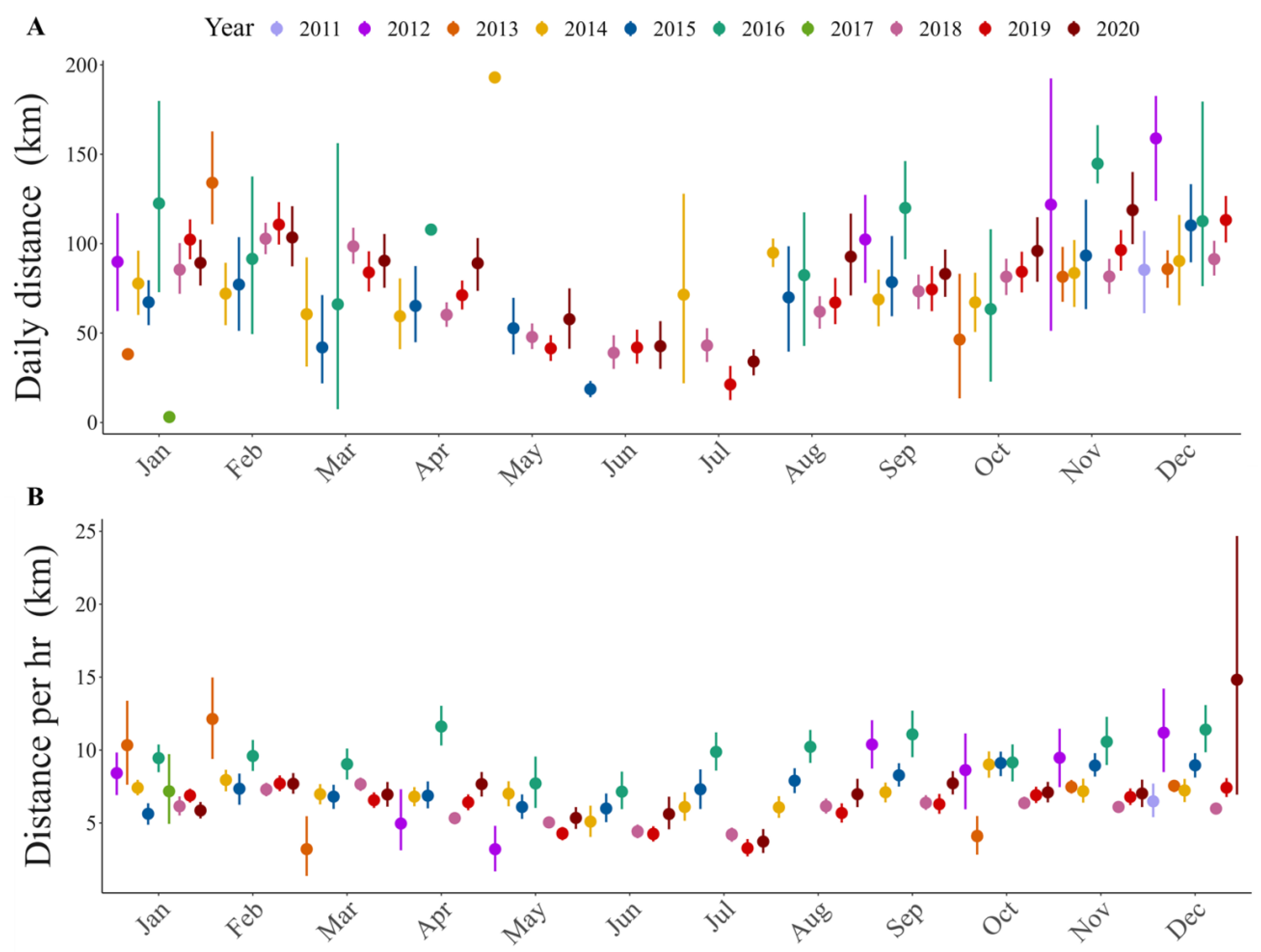
3.2. Flight Distances
4. Discussion
Condors vs. Other Vulture Movements
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bowler, D.E.; Benton, T.G. Causes and Consequences of Animal Dispersal Strategies: Relating Individual Behaviour to Spatial Dynamics. Biol. Rev. 2005, 80, 205–225. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, P.J. Mating Systems, Philopatry and Dispersal in Birds and Mammals. Anim. Behav. 1980, 28, 1140–1162. [Google Scholar] [CrossRef]
- Ronce, O. How Does It Feel to Be like a Rolling Stone? Ten Questions about Dispersal Evolution. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 231–253. [Google Scholar] [CrossRef]
- Gil, J.A.; Díez, Ó. Dispersión Juvenil Del Quebrantahuesos En Los Pirineos [Juvenile Dispersal of Bearded Vulture in the Pyrenees]. Quercus 1993, 91, 13–16. (In Spanish) [Google Scholar]
- Walls, S.S.; Kenward, R.E. Movements of Radio-Tagged Buzzards Buteo buteo in Early Life. IBIS 1998, 140, 561–568. [Google Scholar] [CrossRef]
- Forsman, E.D.; Anthony, R.G.; Reid, J.A.; Loschl, P.J.; Sovern, S.G.; Taylor, M.; Biswell, B.L.; Ellingson, A.; Meslow, E.C.; Miller, G.S. Natal and Breeding Dispersal of Northern Spotted Owls. Wildl. Monogr. 2002, 149, 1–35. [Google Scholar]
- Snyder, N.F.; Schmitt, N.J. California Condor: Gymnogyps californianus. In The Birds of North America; Poole, A., Gill, F., Eds.; The Academy of Natural Sciences and The American Ornithologists’ Union: Philadelphia, PA, USA; Washington, DC, USA, 2002. [Google Scholar]
- Vasilakis, D.P.; Poirazidis, K.S.; Elorriaga, J.N. Range Use of a Eurasian Black Vulture (Aegypius monachus) Population in the Dadia-Lefkimi-Soufli National Park and the Adjacent Areas, Thrace, NE Greece. J. Nat. Hist. 2008, 42, 355–373. [Google Scholar] [CrossRef]
- Alarcón, P.A.E.; Lambertucci, S.A. A Three-Decade Review of Telemetry Studies on Vultures and Condors. Mov. Ecol. 2018, 6, 13. [Google Scholar] [CrossRef]
- McCaslin, H.M.; Caughlin, T.T.; Heath, J.A. Long-Distance Natal Dispersal Is Relatively Frequent and Correlated with Environmental Factors in a Widespread Raptor. J. Anim. Ecol. 2020, 89, 2077–2088. [Google Scholar] [CrossRef]
- Penteriani, V.; Ferrer, M.; Delgado, M.d.M. Floater Strategies and Dynamics in Birds, and Their Importance in Conservation Biology: Towards an Understanding of Nonbreeders in Avian Populations. Anim. Conserv. 2011, 14, 233–241. [Google Scholar] [CrossRef]
- Cagnacci, F.; Boitani, L.; Powell, R.A.; Boyce, M.S. Animal Ecology Meets GPS-Based Radiotelemetry: A Perfect Storm of Opportunities and Challenges. Philos. Trans. R. Soc. B 2010, 365, 2157–2162. [Google Scholar] [CrossRef] [Green Version]
- Penteriani, V.; Delgado, M.d.M. Thoughts on Natal Dispersal. J. Raptor Res. 2009, 43, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Krüger, S.; Reid, T.; Amar, A. Differential Range Use between Age Classes of Southern African Bearded Vultures Gypaetus barbatus. PLoS ONE 2014, 9, e114920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margalida, A.; Pérez-García, J.M.; Afonso, I.; Moreno-Opo, R. Spatial and Temporal Movements in Pyrenean Bearded Vultures (Gypaetus barbatus): Integrating Movement Ecology into Conservation Practice. Sci. Rep. 2016, 6, 35746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morant, J.; Arrondo, E.; Sánchez-Zapata, J.A.; Donázar, J.A.; Cortés-Avizanda, A.; De La Riva, M.; Blanco, G.; Martínez, F.; Oltra, J.; Carrete, M.; et al. Large-Scale Movement Patterns in a Social Vulture Are Influenced by Seasonality, Sex, and Breeding Region. Ecol. Evol. 2023, 13, e9817. [Google Scholar] [CrossRef]
- Morrison, J.L.; Wood, P.B. Broadening Our Approaches to Studying Dispersal in Raptors. J. Raptor Res. 2009, 43, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Clobert, J.; Le Galliard, J.-F.; Cote, J.; Meylan, S.; Massot, M. Informed Dispersal, Heterogeneity in Animal Dispersal Syndromes and the Dynamics of Spatially Structured Populations. Ecol. Lett. 2009, 12, 197–209. [Google Scholar] [CrossRef]
- Clobert, J.; Baguette, M.; Benton, T.G.; Bullock, J.M. Dispersal Ecology and Evolution; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Walters, J.R. Dispersal Behavior: An Ornithological Frontier. Condor 2000, 102, 479–481. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Both, C.; Van Noordwijk, A.J.; Rutten, A.L.; Drent, P.J. Natal Dispersal and Personalities in Great Tits (Parus major). Proc. R. Soc. London. Ser. B Biol. Sci. 2003, 270, 741–747. [Google Scholar] [CrossRef] [Green Version]
- Aebischer, A.; Nyffeler, P.; Arlettaz, R. Wide-Range Dispersal in Juvenile Eagle Owls (Bubo bubo) across the European Alps Calls for Transnational Conservation Programmes. J. Ornithol. 2010, 151, 1. [Google Scholar] [CrossRef] [Green Version]
- Delgado, M.d.M.; Penteriani, V.; Nams, V.O. How Fledglings Explore Surroundings from Fledging to Dispersal. A Case Study with Eagle Owls Bubo bubo. Ardea 2009, 97, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Kouba, M.; Bartoš, L.; Štastný, K. Differential Movement Patterns of Juvenile Tengmalms Owls (Aegolius funereus) during the Post-Fledging Dependence Period in Two Years with Contrasting Prey Abundance. PLoS ONE 2013, 8, e67034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolaus, M.; Wang, X.; Lamers, K.P.; Ubels, R.; Both, C. Unravelling the Causes and Consequences of Dispersal Syndromes in a Wild Passerine. Proc. R. Soc. B Biol. Sci. 2022, 289, 20220068. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.D.M.; Ramsden, D.; Roper, S.; Cresswell, B.; Skuse, J. Juvenile Barn Owl Tyto alba Dispersal: A Radio-Tracking Study of Roost Site Selection in Relation to Landscape Features. Bird Study 2021, 68, 245–257. [Google Scholar] [CrossRef]
- del Hoyo, J.; Elliot, A.; Sargatal, J. Handbook of the Birds of the World. New World Vultures to Guineafowl, 2nd ed.; Lynx edicions: Barcelona, Spain, 1994; ISBN 84-87334-15-6. [Google Scholar]
- Bustamante, J.; Hiraldo, F. Post-Fledging Dependence Period and Maturation of Flight Skills in the Black Kite Milvus migrans. Bird Study 1989, 36, 199–204. [Google Scholar] [CrossRef]
- Bustamante, J.; Negro, J.J. The Post-Fledging Dependence Period in the Lesser Kestrel Falco naumanni. J. Raptor Res. 1994, 28, 158–163. [Google Scholar]
- Krüger, S.; Amar, A. Insights into Post-Fledging Dispersal of Bearded Vultures Gypaetus barbatus in Southern Africa from GPS Satellite Telemetry. Bird Study 2017, 64, 125–131. [Google Scholar] [CrossRef]
- Williams, H.J.; Shepard, E.L.C.; Holton, M.D.; Alarcón, P.A.E.; Wilson, R.P.; Lambertucci, S.A. Physical Limits of Flight Performance in the Heaviest Soaring Bird. Proc. Natl. Acad. Sci. USA 2020, 117, 17884–17890. [Google Scholar] [CrossRef]
- Bamford, A.J.; Diekmann, M.; Monadjem, A.; Mendelsohn, J. Ranging Behaviour of Cape Vultures Gyps coprotheres from an Endangered Population in Namibia. Bird Conserv. Int. 2007, 17, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Plaza, P.I.; Lambertucci, S.A. Ecology and Conservation of a Rare Species: What Do We Know and What May We Do to Preserve Andean Condors? Biol. Conserv. 2020, 251, 108782. [Google Scholar] [CrossRef]
- Birdlife International Vultur Gryphus. The IUCN Red List of Threatened Species 2020: E.T22697641A181325230. Available online: https://doi.org/10.2305/IUCN.UK.2020-3.RLTS.T22697641A181325230.en (accessed on 12 March 2021).
- Guido, J.M.; Alarcón, P.A.E.; Donázar, J.A.; Hiraldo, F.; Lambertucci, S.A. The Use of Biosphere Reserves by a Wide-Ranging Avian Scavenger Indicates Its Significant Potential for Conservation. Environ. Conserv. 2019, 47, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, A.L. Regiones Fitogeográficas Argentinas; Enciclopedia Argentina de agricultura y jardinería [Argentine Phytogeographic Regions; Argentine encyclopedia of agriculture and gardening]; ACME: Buenos Aires, Argentina, 1976. (In Spanish) [Google Scholar]
- Paruelo, J.M.; Beltran, A.; Jobbagy, E.; Sala, O.E.; Golluscio, R.A. The Climate of Patagonia: General Patterns and Controls on Biotic Processes. Ecol. Austral 1998, 8, 85–101. [Google Scholar]
- Ballejo, F.; Lambertucci, S.A.; Trejo, A.; De Santis, L.J. Trophic Niche Overlap among Scavengers in Patagonia Supports the Condor-Vulture Competition Hypothesis. Bird Conserv. Int. 2018, 28, 390–402. [Google Scholar] [CrossRef] [Green Version]
- Lambertucci, S.A.; Navarro, J.; Zapata, J.A.S.; Hobson, K.A.; Alarcón, P.A.; Wiemeyer, G.; Blanco, G.; Hiraldo, F.; Donázar, J.A. Tracking Data and Retrospective Analyses of Diet Reveal the Consequences of Loss of Marine Subsidies for an Obligate Scavenger, the Andean Condor. Proc. R. Soc. B 2018, 285, 20180550. [Google Scholar] [CrossRef] [PubMed]
- Lambertucci, S.A.; Trejo, A.; Di Martino, S.; Sánchez-Zapata, J.A.; Donázar, J.A.; Hiraldo, F. Spatial and Temporal Patterns in the Diet of the Andean Condor: Ecological Replacement of Native Fauna by Exotic Species. Anim. Conserv. 2009, 12, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Ferguson-Lees, J.; Christie, D.A. Raptors of the World; Houghton Mifflin Harcourt: Boston, MA, USA, 2001. [Google Scholar]
- Wallace, R.B.; Reinaga, A.; Piland, N.; Piana, R.; Vargas, F.H.; Zegarra, R.E.; Alvarado, S.; Kohn, S.; Lambertucci, S.A.; Alarcón, P.; et al. Defining Spatial Conservation Priorities for the Andean Condor (Vultur gryphus). J. Raptor Res. 2022, 56, 1–16. [Google Scholar] [CrossRef]
- Lambertucci, S.A.; Mastrantuoni, O.A. Breeding Behavior of a Pair of Free-Living Andean Condors. J. Field Ornithol. 2008, 79, 147–151. [Google Scholar] [CrossRef]
- Lambertucci, S.A. Biología y Conservación Del Cóndor Andino (Vultur gryphus) En Argentina [Biology and Conservation of the Andean Condor (Vultur gryphus) in Argentina]. El Hornero 2007, 22, 149–158. (In Spanish) [Google Scholar]
- Alarcón, P.A.; Morales, J.M.; Donázar, J.A.; Sánchez-Zapata, J.A.; Hiraldo, F.; Lambertucci, S.A. Sexual-Size Dimorphism Modulates the Trade-off between Exploiting Food and Wind Resources in a Large Avian Scavenger. Sci. Rep. 2017, 7, 11461. [Google Scholar] [CrossRef] [Green Version]
- Perrig, P.L.; Lambertucci, S.A.; Donadio, E.; Padró, J.; Pauli, J.N. Monitoring Vultures in the 21st Century: The Need for Standardized Protocols. J. Appl. Ecol. 2019, 56, 796–801. [Google Scholar] [CrossRef] [Green Version]
- Wallace, M.P.; Temple, S.A. Impacts of the 1982–1983 El Niño on Population Dynamics of Andean Condors in Peru. Biotropica 1988, 20, 144–150. [Google Scholar] [CrossRef]
- Hofman, M.P.G.; Hayward, M.W.; Heim, M.; Marchand, P.; Rolandsen, C.M.; Mattisson, J.; Urbano, F.; Heurich, M.; Mysterud, A.; Melzheimer, J.; et al. Right on Track? Performance of Satellite Telemetry in Terrestrial Wildlife Research. PLoS ONE 2019, 14, e0216223. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Fieberg, J.; Börger, L. Could You Please Phrase “Home Range” as a Question? J. Mammal. 2012, 93, 890–902. [Google Scholar] [CrossRef]
- Walter, W.D.; Onorato, D.P.; Fischer, J.W. Is There a Single Best Estimator? Selection of Home Range Estimators Using Area-under-the-Curve. Mov. Ecol. 2015, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bivand, R.; Keitt, T.; Rowlingson, B.; Pebesma, E.; Sumner, M.; Hijmans, R.; Rouault, E.; Bivand, M.R. Package ‘Rgdal.’ Bindings for the Geospatial Data Abstraction Library. Available online: https://cran.r-project.org/web/packages/rgdal/index.html (accessed on 15 October 2017).
- Calenge, C. Home Range Estimation in R: The AdehabitatHR Package; Office National de la Classe et de la Faune Sauvage: Auffargis, France, 2011. [Google Scholar]
- Laver, P. ABODE: Kernel Home Range Estimation for ArcGIS, Using VBA and ArcObjects. User Man. Beta Version 2005, 2, 62. [Google Scholar]
- Laver, P.N.; Kelly, M.J. A Critical Review of Home Range Studies. J. Wildl. Manag. 2008, 72, 290–298. [Google Scholar] [CrossRef]
- Powell, R.A.; Mitchell, M.S. What Is a Home Range? J. Mammal. 2012, 93, 948–958. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: New York, NY, USA, 2009. [Google Scholar]
- Pinheiro, J.; Bates, D. Mixed-Effects Models in S and S-PLUS; Springer Science & Business Media: Berlin/Heidelberg, Germany; New York, NY, USA, 2000. [Google Scholar]
- Alarcón, P.A.E. Movimiento Animal y Patrones Emergentes de Uso Del Espacio: Hacia Una Interpretación Mecanística de La Ecología Del Cóndor Andino (Vultur gryphus) [Animal Movement and Emerging Patterns of Space Use: Towards a Mechanistic Interpretation of the Ecology of the Andean Condor (Vultur gryphus)]; Universidad Nacional del Comahue: San Carlos de Bariloche, Argentina, 2016. (In Spanish) [Google Scholar]
- Lambertucci, S.A.; Alarcón, P.A.; Hiraldo, F.; Sanchez-Zapata, J.A.; Blanco, G.; Donázar, J.A. Apex Scavenger Movements Call for Transboundary Conservation Policies. Biol. Conserv. 2014, 170, 145–150. [Google Scholar] [CrossRef] [Green Version]
- López-López, P.; Gil, J.A.; Alcántara, M. Post-Fledging Dependence Period and Onset of Natal Dispersal in Bearded Vultures (Gypaetus barbatus): New Insights from GPS Satellite Telemetry. J. Raptor Res. 2014, 48, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Martens, F.R.; Pfeiffer, M.B.; Downs, C.T.; Venter, J.A. Post-Fledging Movement and Spatial Ecology of the Endangered Cape Vulture (Gyps coprotheres). J. Ornithol. 2018, 159, 913–922. [Google Scholar] [CrossRef] [Green Version]
- Kane, A.; Wolter, K.; Neser, W.; Kotze, A.; Naidoo, V.; Monadjem, A. Home Range and Habitat Selection of Cape Vultures Gyps coprotheres in Relation to Supplementary Feeding. Bird Study 2016, 63, 387–394. [Google Scholar] [CrossRef]
- Margalida, A.; Carrete, M.; Hegglin, D.; Serrano, D.; Arenas, R.; Donázar, J.A. Uneven Large-Scale Movement Patterns in Wild and Reintroduced Pre-Adult Bearded Vultures: Conservation Implications. PLoS ONE 2013, 8, e65857. [Google Scholar] [CrossRef] [Green Version]
- Phipps, W.L.; Willis, S.G.; Wolter, K.; Naidoo, V. Foraging Ranges of Immature African White-Backed Vultures (Gyps africanus) and Their Use of Protected Areas in Southern Africa. PLoS ONE 2013, 8, e52813. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-Y.; Kokko, H. Sex-Biased Dispersal: A Review of the Theory. Biol. Rev. 2019, 94, 721–736. [Google Scholar] [CrossRef] [Green Version]
- Trochet, A.; Courtois, E.A.; Stevens, V.M.; Baguette, M.; Chaine, A.; Schmeller, D.S.; Clobert, J.; Wiens, J.J. Evolution of Sex-Biased Dispersal. Q. Rev. Biol. 2016, 91, 297–320. [Google Scholar] [CrossRef]
- Rivers, J.W.; Johnson, J.M.; Haig, S.M.; Schwarz, C.J.; Burnett, L.J.; Brandt, J.; George, D.; Grantham, J. An Analysis of Monthly Home Range Size in the Critically Endangered California Condor Gymnogyps californianus. Bird Conserv. Int. 2014, 24, 492–504. [Google Scholar] [CrossRef] [Green Version]
- Donázar, J.A.; Travaini, A.; Ceballos, O.; Rodríguez, A.; Delibes, M.; Hiraldo, F. Effects of Sex-Associated Competitive Asymmetries on Foraging Group Structure and Despotic Distribution in Andean Condors. Behav. Ecol. Sociobiol. 1999, 45, 55–65. [Google Scholar] [CrossRef]
- Marinero, N.V.; Cailly-Arnulphi, V.B.; Lambertucci, S.A.; Borghi, C.E. Pigmentation and Not Only Sex and Age of Individuals Affects Despotism in the Andean Condor. PLoS ONE 2018, 13, e0205197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padró, J.; Pauli, J.N.; Perrig, P.L.; Lambertucci, S.A. Genetic Consequences of Social Dynamics in the Andean Condor: The Role of Sex and Age. Behav. Ecol. Sociobiol. 2019, 73, 100. [Google Scholar] [CrossRef]
- Morales, J.M.; Moorcroft, P.R.; Matthiopoulos, J.; Frair, J.L.; Kie, J.G.; Powell, R.A.; Merrill, E.H.; Haydon, D.T. Building the Bridge between Animal Movement and Population Dynamics. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2289–2301. [Google Scholar] [CrossRef] [Green Version]
- Nathan, R. An Emerging Movement Ecology Paradigm. Proc. Natl. Acad. Sci. USA 2008, 105, 19050–19051. [Google Scholar] [CrossRef] [Green Version]
- Buechley, E.R.; McGrady, M.J.; Çoban, E.; Şekercioğlu, Ç.H. Satellite Tracking a Wide-Ranging Endangered Vulture Species to Target Conservation Actions in the Middle East and East Africa. Biodivers. Conserv. 2018, 27, 2293–2310. [Google Scholar] [CrossRef]
- Kang, J.-H.; Hyun, B.-R.; Kim, I.K.; Lee, H.; Lee, J.-K.; Hwang, H.-S.; Eom, T.-K.; Rhim, S.-J. Movement and Home Range of Cinereous Vulture Aegypius monachus during the Wintering and Summering Periods in East Asia. Turk. J. Zool. 2019, 43, 305–313. [Google Scholar] [CrossRef]
- Peshev, H.; Stoynov, E.; Parvanov, D.; Grozdanov, A. Seasonal and Spatial Dynamics of the Population of the Griffon Vulture Gyps fulvus (Hablizl, 1783) (Aves: Accipitridae) in Southwestern Bulgaria. Acta Zool. Bulg. 2018, 2018 (Suppl. 12), 67–75. [Google Scholar]
- Sherub, S.; Fiedler, W.; Duriez, O.; Wikelski, M. Bio-Logging, New Technologies to Study Conservation Physiology on the Move: A Case Study on Annual Survival of Himalayan Vultures. J. Comp. Physiol. A 2017, 203, 531–542. [Google Scholar] [CrossRef] [Green Version]
- Harel, R.; Duriez, O.; Spiegel, O.; Fluhr, J.; Horvitz, N.; Getz, W.M.; Bouten, W.; Sarrazin, F.; Hatzofe, O.; Nathan, R. Decision-Making by a Soaring Bird: Time, Energy and Risk Considerations at Different Spatio-Temporal Scales. Philos. Trans. R. Soc. B 2016, 371, 20150397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urios, V.; López-López, P.; Limiñana, R.; Godino, A. Ranging Behaviour of a Juvenile Bearded Vulture (Gypaetus barbatus meridionalis) in South Africa Revealed by GPS Satellite Telemetry. Ornis Fenn. 2010, 87, 114–118. [Google Scholar]
- Perrig, P.L.; Lambertucci, S.A.; Cruz, J.; Alarcón, P.A.E.; Plaza, P.I.; Middleton, A.D.; Blanco, G.; Sánchez-Zapata, J.A.; Donázar, J.A.; Pauli, J.N. Identifying Conservation Priority Areas for the Andean condor in Southern South America. Biol. Conserv. 2020, 243, 108494. [Google Scholar] [CrossRef]
- Arrondo, E.; Moleón, M.; Cortés-Avizanda, A.; Jiménez, J.; Beja, P.; Sánchez-Zapata, J.A.; Donázar, J.A. Invisible Barriers: Differential Sanitary Regulations Constrain Vulture Movements across Country Borders. Biol. Conserv. 2018, 219, 46–52. [Google Scholar] [CrossRef]
- Pauli, J.N.; Donadio, E.; Lambertucci, S.A. The Corrupted Carnivore: How Humans Are Rearranging the Return of the Carnivore-Scavenger Relationship. Ecology 2018, 99, 2122–2124. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Variables | Est. Value | Lower | Upper | Std. Error | t-Value | p-Value | |
|---|---|---|---|---|---|---|---|---|
| KDE 95% | (Intercept) | 3.843 | 3.695 | 3.991 | 0.076 | 50.460 | 0.000 | * |
| Season (warm) | 0.243 | 0.110 | 0.377 | 0.069 | 3.530 | 0.001 | * | |
| Age (sub-adult) | 0.069 | −0.108 | 0.245 | 0.086 | 0.796 | 0.435 | ||
| Sex (male) | −0.094 | −0.304 | 0.116 | 0.101 | −0.930 | 0.366 | ||
| Season (warm): Age (sub-adult) | −0.002 | −0.146 | 0.141 | 0.074 | −0.028 | 0.978 | ||
| Season (warm): Sex (male) | −0.096 | −0.215 | 0.024 | 0.062 | −1.554 | 0.123 | ||
| Age (sub-adult): Sex (male) | 0.227 | 0.017 | 0.438 | 0.103 | 2.211 | 0.039 | * | |
| KDE 50% | (Intercept) | 2.977 | 2.805 | 3.149 | 0.088 | 33.640 | 0.000 | * |
| Season (warm) | 0.305 | 0.140 | 0.470 | 0.085 | 3.577 | 0.001 | * | |
| Age (sub-adult) | 0.205 | 0.000 | 0.409 | 0.100 | 2.045 | 0.054 | ||
| Sex (male) | −0.073 | −0.315 | 0.168 | 0.116 | −0.630 | 0.538 | ||
| Season (warm): Age (sub-adult) | −0.112 | −0.289 | 0.065 | 0.092 | −1.224 | 0.223 | ||
| Season (warm): Sex (male) | −0.134 | −0.281 | 0.013 | 0.076 | −1.763 | 0.080 | ||
| Age (sub-adult): Sex (male) | 0.245 | 0.006 | 0.483 | 0.117 | 2.093 | 0.049 | * |
| Model | Variables | Est. Value | Lower | Upper | Std. Error | t-Value | p-Value | |
|---|---|---|---|---|---|---|---|---|
| Hourly Distance | (Intercept) | 2.940 | 2.805 | 3.075 | 0.069 | 42.746 | 0.000 | * |
| Season (warm) | −0.098 | −0.148 | −0.047 | 0.026 | −3.781 | 0.000 | * | |
| Age (sub-adult) | 0.203 | 0.085 | 0.322 | 0.060 | 3.371 | 0.001 | * | |
| Sex (male) | −0.093 | −0.292 | 0.106 | 0.096 | −0.968 | 0.343 | ||
| Season (warm): Age (sub-adult) | −0.015 | −0.069 | 0.040 | 0.028 | −0.527 | 0.598 | ||
| Season (warm): Sex (male) | 0.081 | 0.037 | 0.126 | 0.023 | 3.567 | 0.000 | * | |
| Age (sub-adult): Sex (male) | −0.033 | −0.199 | 0.134 | 0.085 | −0.384 | 0.701 | ||
| Daily Distance | (Intercept) | 4.528 | 4.457 | 4.599 | 0.036 | 125.178 | 0.000 | * |
| Season (warm) | 0.219 | 0.191 | 0.248 | 0.014 | 15.211 | 0.000 | * | |
| Age (sub-adult) | 0.137 | 0.081 | 0.193 | 0.028 | 4.808 | 0.000 | * | |
| Sex (male) | 0.038 | −0.039 | 0.115 | 0.037 | 1.041 | 0.312 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guido, J.M.; Cecchetto, N.R.; Plaza, P.I.; Donázar, J.A.; Lambertucci, S.A. The Influence of Age, Sex and Season on Andean Condor Ranging Behavior during the Immature Stage. Animals 2023, 13, 1234. https://doi.org/10.3390/ani13071234
Guido JM, Cecchetto NR, Plaza PI, Donázar JA, Lambertucci SA. The Influence of Age, Sex and Season on Andean Condor Ranging Behavior during the Immature Stage. Animals. 2023; 13(7):1234. https://doi.org/10.3390/ani13071234
Chicago/Turabian StyleGuido, Jorgelina María, Nicolás Rodolfo Cecchetto, Pablo Ignacio Plaza, José Antonio Donázar, and Sergio Agustín Lambertucci. 2023. "The Influence of Age, Sex and Season on Andean Condor Ranging Behavior during the Immature Stage" Animals 13, no. 7: 1234. https://doi.org/10.3390/ani13071234