
Epithelial-to-Mesenchymal Transition and Phenotypic Marker Evaluation in Human, Canine, and Feline Mammary Gland Tumors
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Collection
2.2. RNA Extraction and Real-Time Polymerase Chain Reaction
2.3. Immunohistochemistry
2.4. Statistical Analysis
3. Results
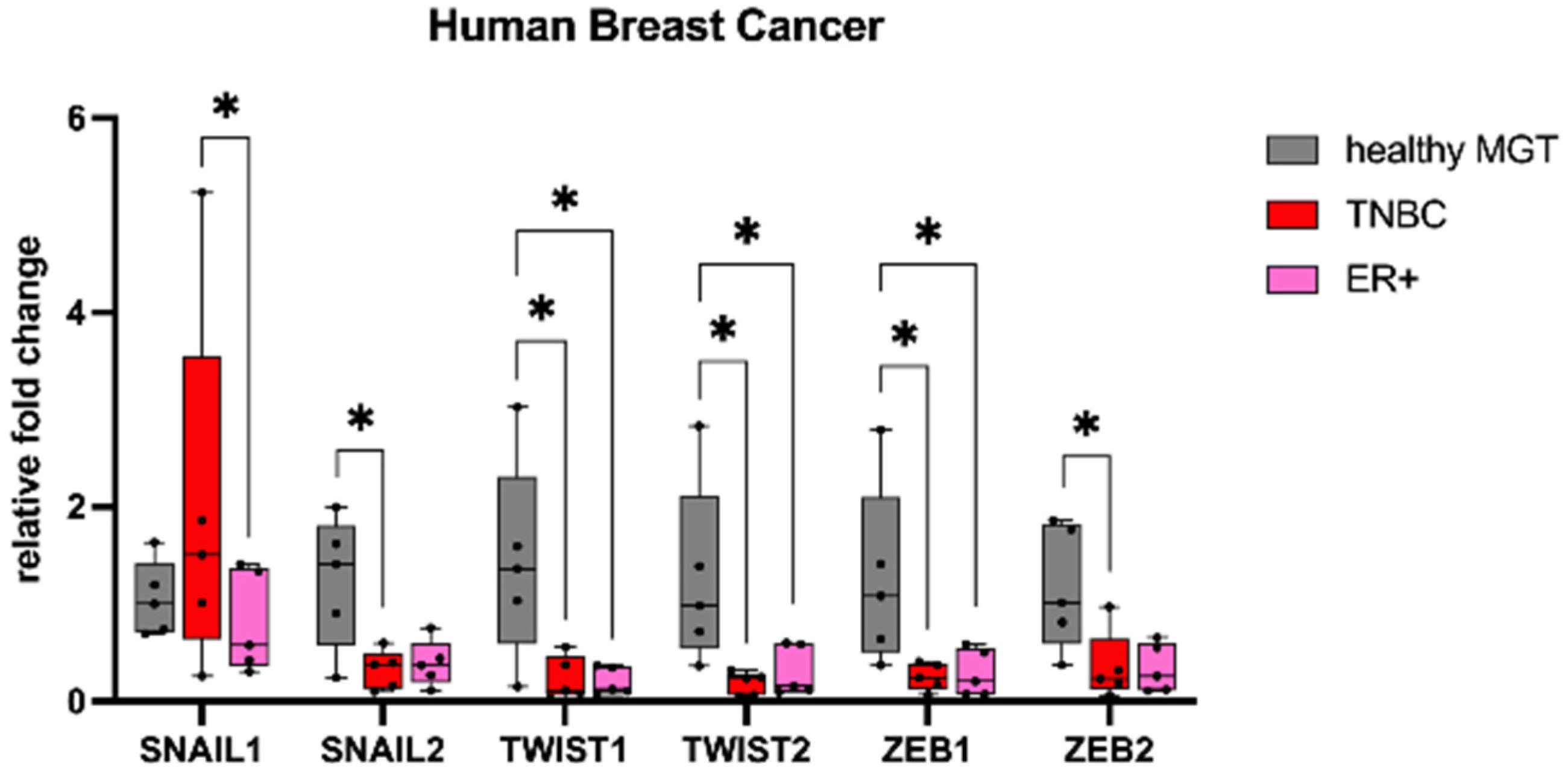
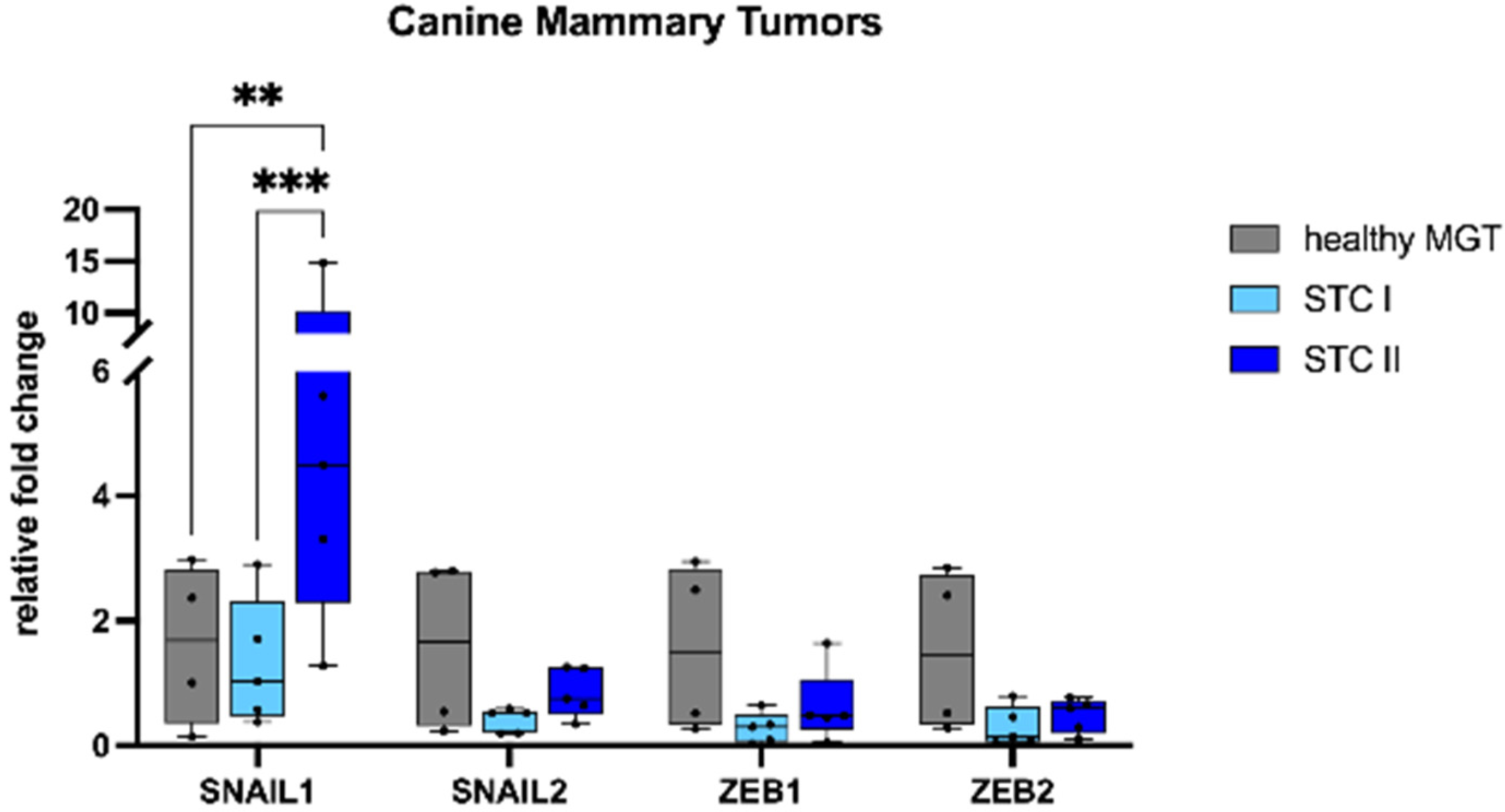
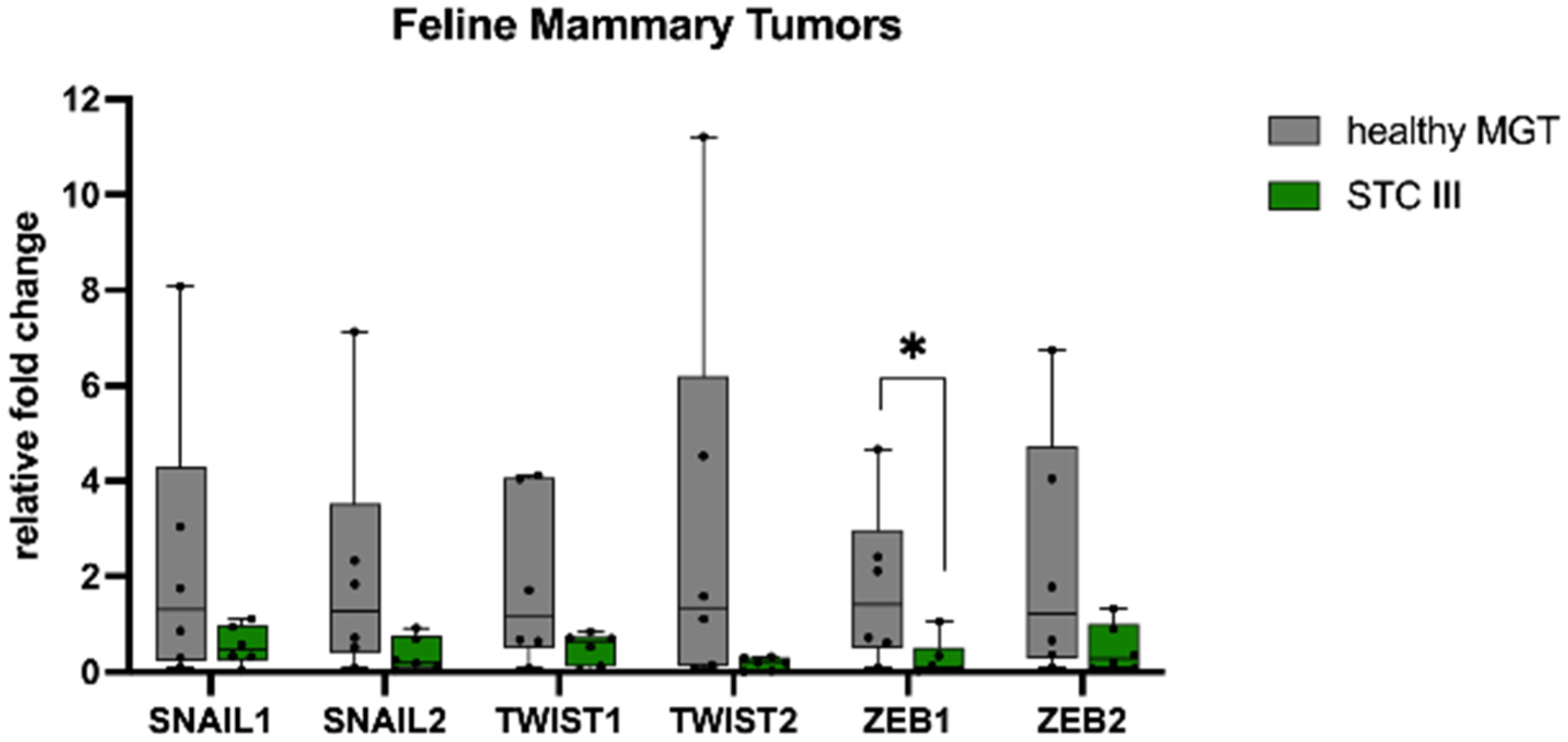
3.1. Gene Expression
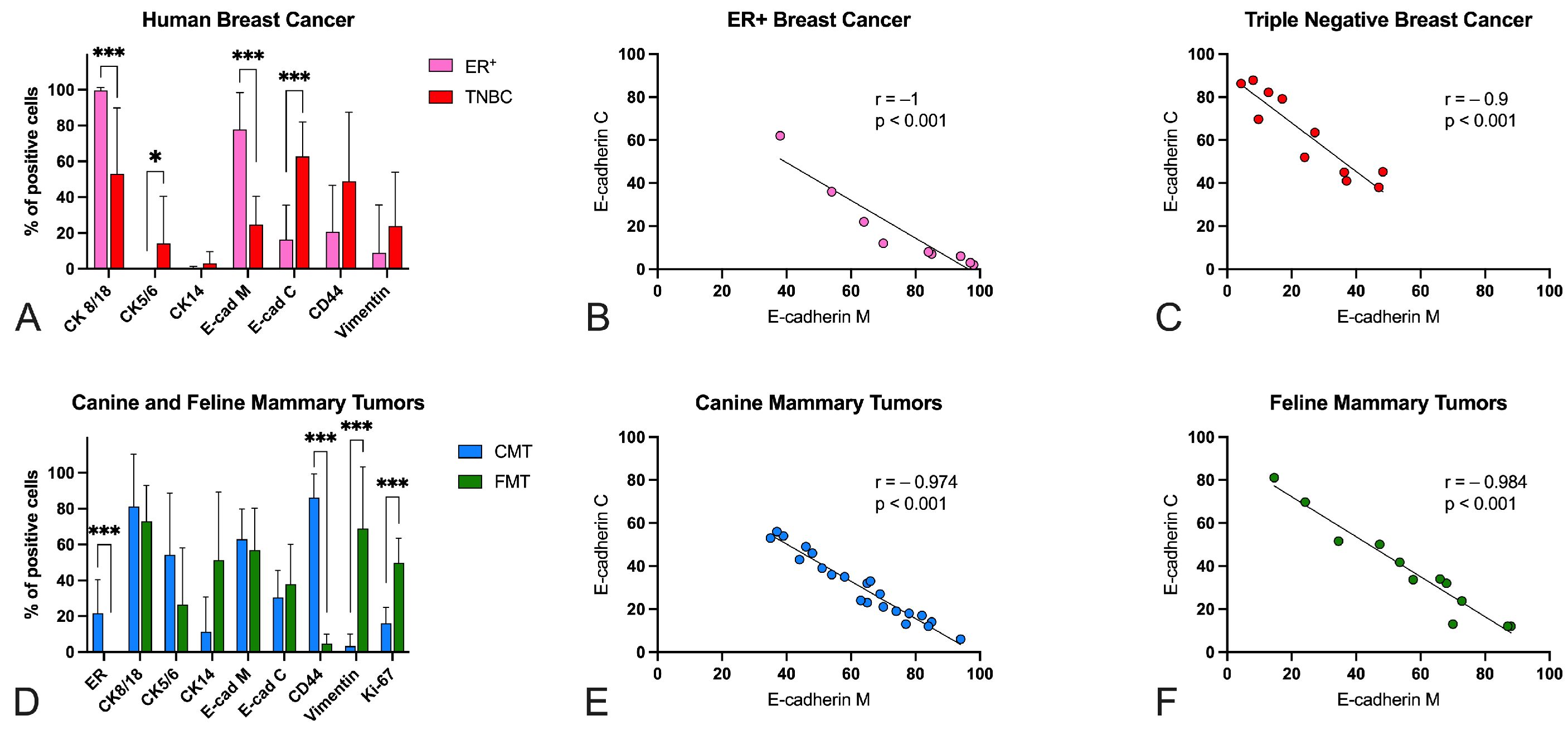
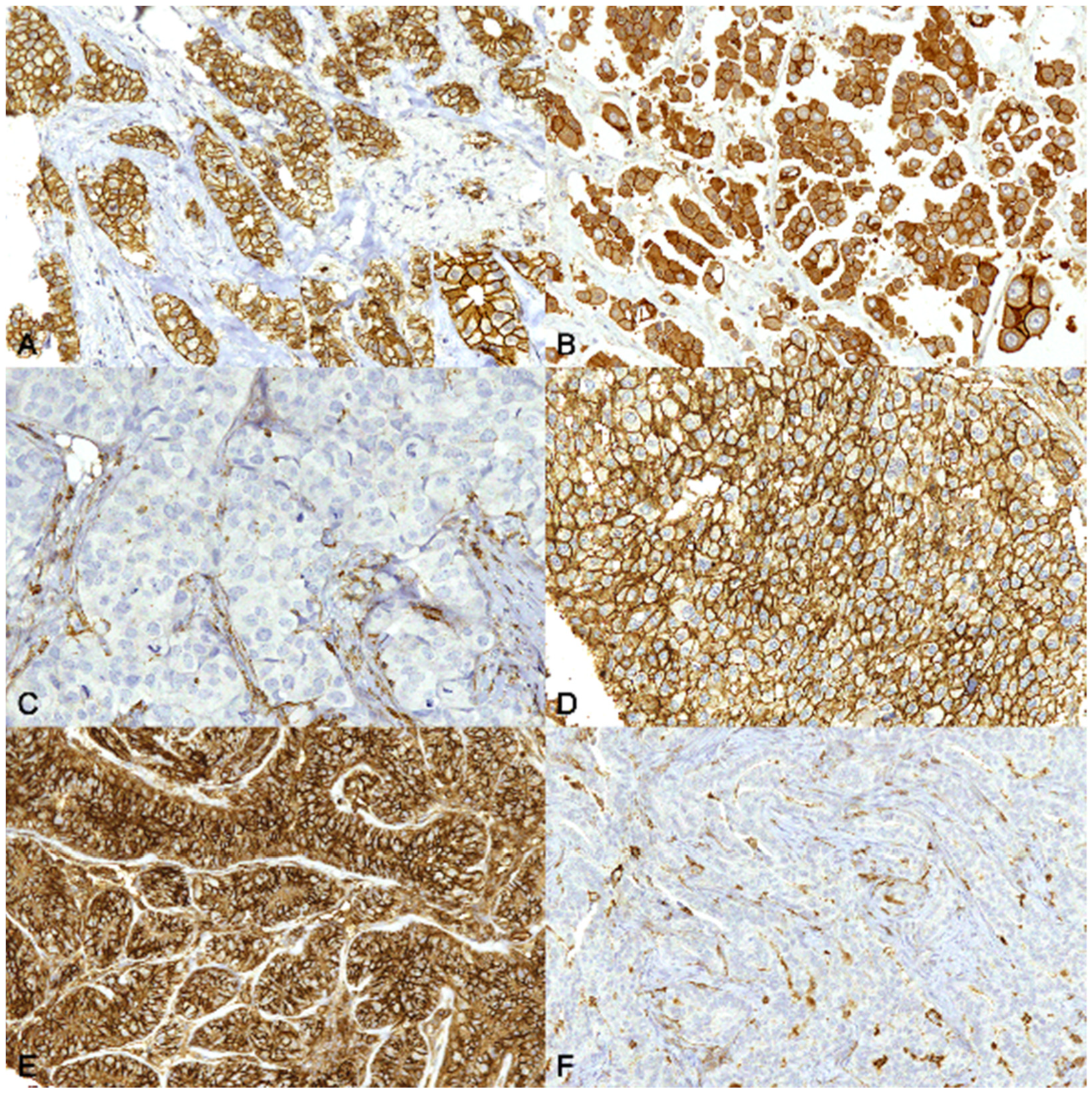
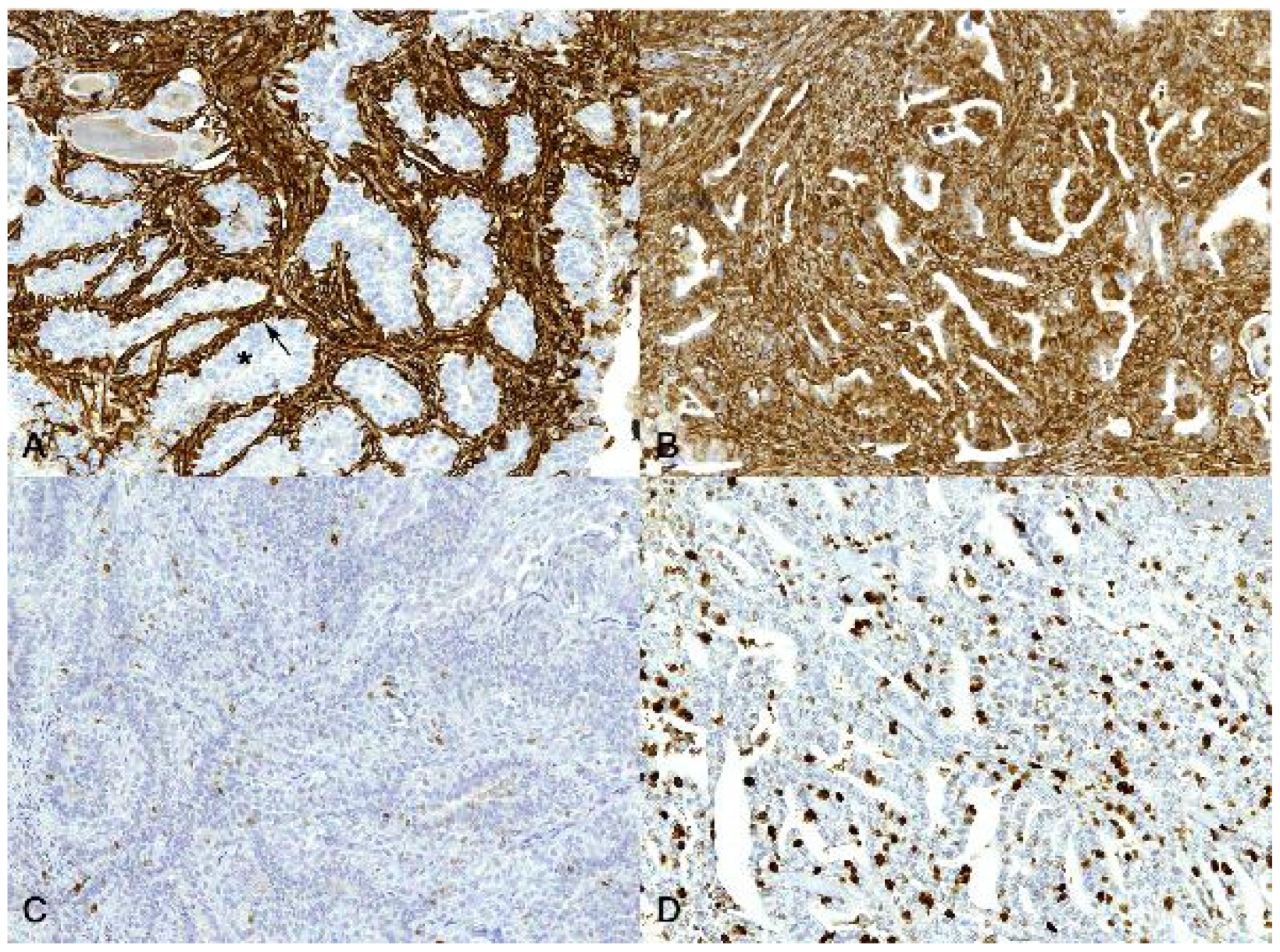
3.2. Immunohistochemistry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Dyba, T.; Randi, G.; Bettio, M.; Gavin, A.; Visser, O.; Bray, F. Cancer incidence and mortality patterns in Europe: Estimates for 40 countries and 25 major cancers in 2018. Eur. J. Cancer 2018, 103, 356–387. [Google Scholar] [CrossRef] [PubMed]
- Sleeckx, N.; de Rooster, H.; Veldhuis Kroeze, E.; Van Ginneken, C.; Van Brantegem, L. Canine Mammary Tumours, an Overview. Reprod. Domest. Anim. 2011, 46, 1112–1131. [Google Scholar] [CrossRef]
- Zappulli, V.; De Zan, G.; Cardazzo, B.; Bargelloni, L.; Castagnaro, M. Feline mammary tumours in comparative oncology. J. Dairy Res. 2005, 72, 98. [Google Scholar] [CrossRef]
- Provenzano, E.; Ulaner, G.A.; Chin, S.F. Molecular Classification of Breast Cancer. PET Clin. 2018, 13, 325–338. [Google Scholar] [CrossRef]
- Shi, Y.; Jin, J.; Ji, W.; Guan, X. Therapeutic landscape in mutational triple negative breast cancer. Mol. Cancer 2018, 17, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldschmidt, M.H.; Peña, L.; Rasotto, R.; Zappulli, V. Classification and grading of canine mammary tumors. Vet. Pathol. 2011, 48, 117–131. [Google Scholar] [CrossRef]
- Sorenmo, K. Canine mammary gland tumors. Vet. Clin. Small Anim. 2003, 33, 573–596. [Google Scholar] [CrossRef] [PubMed]
- Misdorp, W.; Else, R.; Hellme’n, E. Histological Classification of Mammary Tumors of the Dog and the Cat; World Health Organization: Geneva, Switzerland, 1999. [Google Scholar]
- Zappulli, V.; Rasotto, R.; Caliari, D.; Mainenti, M.; Peña, L.; Goldschmidt, M.H.; Kiupel, M. Prognostic Evaluation of Feline Mammary Carcinomas: A Review of the Literature. Vet. Pathol. 2015, 52, 46–60. [Google Scholar] [CrossRef]
- Rasotto, R.; Caliari, D.; Castagnaro, M.; Zanetti, R.; Zappulli, V. An Immunohistochemical Study of HER-2 Expression in Feline Mammary Tumours. J. Comp. Pathol. 2011, 144, 170–179. [Google Scholar] [CrossRef]
- Caliari, D.; Zappulli, V.; Rasotto, R.; Cardazzo, B.; Frassineti, F.; Goldschmidt, M.H.; Castagnaro, M. Triple-negative vimentin-positive heterogeneous feline mammary carcinomas as a potential comparative model for breast cancer. BMC Vet. Res. 2014, 10, 185. [Google Scholar] [CrossRef] [Green Version]
- Wiese, D.A.; Thaiwong, T.; Yuzbasiyan-Gurkan, V.; Kiupel, M. Feline mammary basal-like adenocarcinomas: A potential model for human triple-negative breast cancer (TNBC) with basal-like subtype. BMC Cancer 2013, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, C.; Ferreira, F. Tumor microenvironment of human breast cancer, and feline mammary carcinoma as a potential study model. Biochim. Biophys. Acta Rev. Cancer 2021, 1876, 188587. [Google Scholar] [CrossRef] [PubMed]
- Gameiro, A.; Urbano, A.C.; Ferreira, F. Emerging biomarkers and targeted therapies in feline mammary carcinoma. Vet. Sci. 2021, 8, 164. [Google Scholar] [CrossRef] [PubMed]
- Dagher, E.; Simbault, L.; Abadie, J.; Loussouarn, D.; Campone, M.; Nguyen, F. Identification of an immune-suppressed subtype of feline triple-negative basal-like invasive mammary carcinomas, spontaneous models of breast cancer. Tumor Biol. 2020, 42, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Adega, F.; Borges, A.; Chaves, R. Cat Mammary Tumors: Genetic Models for the Human Counterpart. Vet. Sci. 2016, 3, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polyak, K. Breast cancer: Origins and evolution. J. Clin. Investig. 2007, 117, 3155–3163. [Google Scholar] [CrossRef]
- Sorenmo, K.U.; Rasotto, R.; Zappulli, V.; Goldschmidt, M.H. Development, anatomy, histology, lymphatic drainage, clinical features, and cell differentiation markers of canine mammary gland neoplasms. Vet. Pathol. 2011, 48, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasotto, R.; Goldschmidt, M.H.; Castagnaro, M.; Carnier, P.; Caliari, D.; Zappulli, V. The dog as a natural animal model for study of the mammary myoepithelial basal cell lineage and its role in mammary carcinogenesis. J. Comp. Pathol. 2014, 151, 166–180. [Google Scholar] [CrossRef]
- Prat, A.; Perou, C.M. Deconstructing the molecular portraits of breast cancer. Mol. Oncol. 2011, 5, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.; Fritz, A.J.; Zaidi, S.K.; van Wijnen, A.J.; Nickerson, J.A.; Imbalzano, A.N.; Lian, J.B.; Stein, J.L.; Stein, G.S. Epithelial-to-mesenchymal transition and cancer stem cells contribute to breast cancer heterogeneity. J. Cell. Physiol. 2018, 23, 9136–9144. [Google Scholar] [CrossRef] [PubMed]
- Kisoda, S.; Mouri, Y.; Kitamura, N.; Yamamoto, T.; Miyoshi, K.; Kudo, Y. The role of partial-EMT in the progression of head and neck squamous cell carcinoma. J. Oral Biosci. 2022, 64, 76–182. [Google Scholar] [CrossRef] [PubMed]
- Kröger, C.; Afeyan, A.; Mraz, J.; Eaton, E.N.; Reinhardt, F.; Khodor, Y.L.; Thiru, P.; Bierie, B.; Ye, X.; Burge, C.B.; et al. Acquisition of a hybrid E/M state is essential for tumorigenicity of basal breast cancer cells. Proc. Natl. Acad. Sci. USA 2019, 116, 7353–7362. [Google Scholar] [CrossRef] [Green Version]
- Norgard, R.J.; Pitarresi, J.R.; Maddipati, R.; Aiello-Couzo, N.M.; Balli, D.; Li, J.; Yamazoe, T.; Wengyn, M.D.; Millstein, I.D.; Folkert, I.W.; et al. Calcium signaling induces a partial EMT. EMBO Rep. 2021, 22, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Ginzinger, D.G. Gene quantification using real-time quantitative PCR: An emerging technology hits the mainstream. Exp. Hematol. 2002, 30, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, M.; Sakai, K.; Maeda, S.; Uchida, K.; Nakayama, H. Assessment of Human Epidermal Growth Factor Receptor 2 Expression in Canine Urothelial Carcinoma of the Urinary Bladder. Vet. Pathol. 2018, 56, 369–376. [Google Scholar] [CrossRef]
- Wolff, A.C.; McShane, L.M.; Hammond, M.E.H.; Allison, K.H.; Fitzgibbons, P.; Press, M.F.; Harvey, B.E.; Mangu, P.B.; Bartlett, J.M.S.; Hanna, W.; et al. Human epidermal growth factor receptor 2 testing in breast cancer: American Society of Clinical Oncology/College of American Pathologists Clinical Practice Guideline Focused Update. Arch. Pathol. Lab. Med. 2018, 142, 1364–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Natl. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; El-naggar, S.; Darling, D.S.; Higashi, Y.; Douglas, C. ZEB1 links epithelial-mesenchymal transition and cellular senescence. Development 2008, 135, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Nieto, M.A. The snail superfamily of zinc-finger transcription factors. Nat. Rev. Mol. Cell Biol. 2002, 3, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Mani, S.A.; Donaher, J.L.; Ramaswamy, S.; Itzykson, R.A.; Come, C.; Savagner, P.; Gitelman, I.; Richardson, A.; Weinberg, R.A.; et al. Twist, a master regulator of morphogenesis, plays an essential role in tumor metastasis. Cell 2004, 117, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Peinado, H.; Olmeda, D.; Cano, A. Snail, ZEB and bHLH factors in tumour progression: An alliance against the epithelial phenotype? Nat. Rev. Cancer 2007, 7, 415–428. [Google Scholar] [CrossRef]
- Côme, C.; Magnino, F.; Bibeau, F.; Barbara, P.D.S.; Becker, K.F.; Theillet, C.; Savagner, P. Human Cancer Biology Snail and Slug Play Distinct Roles during Breast Carcinoma Progression. Hum. Cancer Biol. 2006, 12, 5395–5403. [Google Scholar] [CrossRef] [Green Version]
- Moody, S.E.; Perez, D.; Pan, T.; Sarkisian, C.J.; Portocarrero, C.P.; Sterner, C.J.; Notorfrancesco, K.L.; Cardiff, R.D.; Chodosh, L.A. The transcriptional repressor Snail promotes mammary tumor recurrence. Cancer Cell 2005, 8, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Geradts, J.; Garcia, A.; Herreros, D.; Su, Z.; Burchette, J.; Broadwater, G.; Bachelder, R.E. Nuclear Snail1 and nuclear ZEB1 protein expression in invasive and intraductal human breast carcinomas. Hum. Pathol. 2011, 42, 1125–1131. [Google Scholar] [CrossRef] [Green Version]
- Im, K.; Kim, J.; Kim, N.; Yu, C.; Hur, T.; Sur, J. Possible role of snail expression as a prognostic factor in canine mammary neoplasia. J. Comp. Pathol. 2012, 147, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Zidar, N.; Gale, N.; Kojc, N.; Volavsek, M.; Cardesa, A.; Alos, L.; Hofler, H.; Blechschmidt, K.; Becker, K.-F. Cadherin–catenin complex and transcription factor Snail-1 in spindle cell carcinoma of the head and neck. Virchows Arch. 2008, 453, 267–274. [Google Scholar] [CrossRef]
- Komatsu, T.; Iwano, H.; Ebisawa, M.; Watabe, A.; Endo, Y.; Hirayama, K.; Taniyama, H.; Kadosawa, T. Pathological classification of canine mammary tumor based on quantifying mRNA levels of hormonal receptors, SATB1, and Snail in tissue and fine needle biopsy samples. J. Vet. Med. Sci. 2012, 74, 719–726. [Google Scholar] [CrossRef] [Green Version]
- Tan, R.; Wang, L.; Song, J.; Li, J.; He, T. Expression and significance of Twist, estrogen receptor, and E-cadherin in human breast cancer cells and tissues. J. Cancer Res. Ther. 2017, 13, 707–714. [Google Scholar]
- Montserrat, N.; Gallardo, A.; Escuin, D.; Catasus, L.; Prat, J.; Gutiérrez-Avignó, F.J.; Peiró, G.; Barnadas, A.; Lerma, E. Repression of E-cadherin by SNAIL, ZEB1, and TWIST in invasive ductal carcinomas of the breast: A cooperative effort? Hum. Pathol. 2011, 42, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Baptista, C.S.; Santos, S.; Laso, A.; Bastos, E.; Ávila, S.; Guedes-Pinto, H.; Gärtner, F.; Gut, I.G.; Castrillo, J.L.; Chaves, R. Sequence variation and mRNA expression of the TWIST1 gene in cats with mammary hyperplasia and neoplasia. Vet. J. 2012, 191, 203–207. [Google Scholar] [CrossRef]
- Eger, A.; Aigner, K.; Sonderegger, S.; Dampier, B.; Oehler, S.; Schreiber, M.; Berx, G.; Cano, A.; Beug, H.; Foisner, R. DeltaEF1 is a transcriptional repressor of E-cadherin and regulates epithelial plasticity in breast cancer cells. Oncogene 2005, 24, 2375–2385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aigner, K.; Dampier, B.; Descovich, L.; Mikula, M.; Sultan, A.; Schreiber, M.; Mikulits, W.; Brabletz, T.; Strand, D.; Obrist, P.; et al. The transcription factor ZEB1 (dEF1) promotes tumour cell dedifferentiation by repressing master regulators of epithelial polarity. Oncogene 2007, 26, 6979–6988. [Google Scholar] [CrossRef] [Green Version]
- Xavier, P.L.P.; Cordeiro, Y.G.; Rochetti, A.L.; Sangalli, J.R.; Zuccari, D.A.P.C.; Silveira, J.C.; Bressan, F.F.; Fukumasu, H. ZEB1 and ZEB2 transcription factors are potential therapeutic targets of canine mammary cancer cells. Vet. Comp. Oncol. 2018, 16, 596–605. [Google Scholar] [CrossRef]
- Isenmann, S.; Arthur, A.; Zannettino, A.; Turner, J.; Shi, S.; Glackin, C.; Gronthos, S. TWIST family of basic helix-loop-helix transcription factors mediate human mesenchymal stem cell growth and commitment. Stem Cells 2009, 27, 2457–2468. [Google Scholar] [CrossRef]
- Pettersson, A.T.; Mejhert, N.; Jernås, M.; Carlsson, L.M.S.; Dahlman, I.; Laurencikiene, J.; Arner, P.; Rydén, M. Twist1 in Human White Adipose Tissue and Obesity. J. Clin. Endocrinol. Metab. 2011, 96, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Díaz, V.M.; de Herreros, A.G. F-box proteins: Keeping the epithelial-to-mesenchymal transition (EMT) in check. Semin. Cancer Biol. 2016, 36, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Díaz, V.; Vinas-Castells, R.; de Herreros, A.G. Regulation of the protein stability of EMT transcription factors Regulation of the protein stability of EMT transcription factors. Cell Adh. Migr. 2014, 8, 418–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Škovierová, H.; Okajčeková, T.; Strnádel, J.; Vidomanova, E.; Halasova, E. Molecular regulation of epithelial-to-mesenchymal transition in tumorigenesis (Review). Int. J. Mol. Med. 2018, 41, 1187–1200. [Google Scholar] [CrossRef] [Green Version]
- Celià-Terrassa, T.; Jolly, M.K. Cancer stem cells and epithelial-to-mesenchymal transition in cancer metastasis. Cold Spring Harb. Perspect. Med. 2020, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sterneck, E.; Poria, D.K.; Balamurugan, K. Slug and E-Cadherin: Stealth Accomplices? Front. Mol. Biosci. 2020, 7, 138. [Google Scholar] [CrossRef]
- Subbalakshmi, A.R.; Sahoo, S.; Biswas, K.; Jolly, M.K. A Computational Systems Biology Approach Identifies SLUG as a Mediator of Partial Epithelial-Mesenchymal Transition (EMT). Cells Tissues Organs 2021, 211, 109–122. [Google Scholar] [CrossRef]
- Hashmi, A.A.; Naz, S.; Hashmi, S.K.; Hussain, Z.F.; Irfan, M.; Bakar, S.M.A.; Faridi, N.; Khan, A.; Edhi, M.M. Cytokeratin 5/6 and cytokeratin 8/18 expression in triple negative breast cancers: Clinicopathologic significance in South-Asian population. BMC Res. Notes 2018, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, S.; Poornima, R.; Challa, V.R.; Goud, Y.G.B. Triple Negative Breast Cancer—Our Experience and Review. Indian J. Surg. Oncol. 2012, 3, 12–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelmegeed, S.M.; Mohammed, S. Canine mammary tumors as a model for human disease (Review). Oncol. Lett. 2018, 15, 8195–8205. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Wu, Y.; Yao, J.; Wang, Y.; Yu, Y.; Rychahou, P.G.; Evers, B.M.; Zhou, B.P. G9a interacts with Snail and is critical for Snail-mediated E-cadherin repression in human breast cancer. J. Clin. Investig. 2012, 122, 1469–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheel, C.; Weinberg, R.A. Cancer stem cells and epithelial-mesenchymal transition: Concepts and molecular links. Semin. Cancer Biol. 2012, 22, 396–403. [Google Scholar] [CrossRef]
- Kowalski, P.J.; Rubin, M.A.; Kleer, C.G. E-cadherin expression in primary carcinomas of the breast and its distant metastases. Breast Cancer Res. 2003, 5, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Chunthapong, J.; Seftor, E.A.; Khalkhali-Ellis, Z.; Seftor, R.E.B.; Amir, S.; Lubaroff, D.M.; Heidger, P.M.; Hendrix, M.J.C. Dual roles of E-cadherin in prostate cancer invasion. J. Cell. Biochem. 2004, 91, 649–661. [Google Scholar] [CrossRef]
- Khoursheed, M.A.; Mathew, T.C.; Makar, R.R.; Louis, S.; Asfar, S.K.; Al-Sayer, H.M.; Dashti, H.M.; Al-Bader, A. Expression of E-cadherin in human colorectal cancer. R. Coll. Surg. Edinb. Irel. 2003, 1, 86–91. [Google Scholar] [CrossRef]
- Liu, J.B.; Feng, C.Y.; Deng, M.; Ge, D.F.; Liu, D.C.; Mi, J.Q.; Feng, X.S. E-cadherin expression phenotypes associated with molecular subtypes in invasive non-lobular breast cancer: Evidence from a retrospective study and meta-analysis. World J. Surg. Oncol. 2017, 15, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.G.; Hiscox, S. Beta-catenin—Cell adhesion and beyond (Review). Int. J. Oncol. 1997, 11, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Zappulli, V.; De Cecco, S.; Trez, D.; Caliari, D.; Aresu, L.; Castagnaro, M. Immunohistochemical Expression of E-Cadherin and β-Catenin in Feline Mammary Tumours. J. Comp. Pathol. 2012, 147, 161–170. [Google Scholar] [CrossRef]
- Restucci, B.; Maiolino, P.; Martano, M.; Esposito, G.; Filippis, D.D.E.; Borzacchiello, G.; Muzio, L.L.O. Expression of b-Catenin, E-cadherin and APC in Canine Mammary Tumors. Anticancer Res. 2007, 27, 3083–3090. [Google Scholar]
- Bruner, H.C.; Derksen, P.W.B. Loss of E-Cadherin-dependent cell-cell adhesion and the development and progression of cancer. Cold Spring Harb. Perspect. Biol. 2018, 10, a029330. [Google Scholar] [CrossRef] [Green Version]
- Sneath, R.J.S.; Mangham, D.C. The normal structure and function of CD44 and its role in neoplasia. J. Clin. Pathol. Mol. Pathol. 1998, 51, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Vos, M.C.; Hollemans, E.; Ezendam, N.; Feijen, H.; Boll, D.; Pijlman, B.; Van Der Putten, H.; Klinkhamer, P.; Van Kuppevelt, T.H.; Van Der Wurff, A.A.M.; et al. MMP-14 and CD44 in Epithelial-to-Mesenchymal Transition (EMT) in ovarian cancer. J. Ovarian Res. 2016, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, G.L.; Song, L.N.; Deng, Z.F.; Chen, Y.; Ma, L.J. Prognostic value of CD44V6 expression in breast cancer: A meta-analysis. Onco-Targets Ther. 2018, 11, 5451–5457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, J.; Pollán, M.; Ruibal, A.; Jiménez, E.; Lucas, A.R.; Núñez, M.I.; Sánchez, J.; Tejerina, A. Histologic grade and cd44 are independent predictors of axillary lymph node invasion in early (t1) breast cancer. Tumor Biol. 1999, 20, 319–330. [Google Scholar] [CrossRef]
- Lyzak, J.S.; Yaremko, M.L.; Recant, W.; Baunoch, D.A.; Joseph, L. Role of CD44 in nonpalpable T1a and T1b breast cancer. Hum. Pathol. 1997, 28, 772–778. [Google Scholar] [CrossRef]
- Klingbeil, P.; Natrajan, R.; Everitt, G.; Vatcheva, R.; Marchio, C.; Palacios, J.; Buerger, H.; Reis-Filho, J.S.; Isacke, C.M. CD44 is overexpressed in basal-like breast cancers but is not a driver of 11p13 amplification. Breast Cancer Res. Treat. 2010, 120, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Damasceno, K.A.; Ferreira, E.; Estrela-Lima, A.; Bosco, Y.; Silva, L.P.; Barros, A.L.B.; Bertagnolli, A.C.; Cassali, G.D. Relationship between the expression of versican and EGFR, HER-2, HER-3 and CD44 in matrixproducing tumours in the canine mammary gland. Histol. Histopathol. 2016, 31, 675–688. [Google Scholar] [CrossRef]
- Paltian, V.; Alldinger, S.; Baumgärtner, W.; Wohlsein, P. Expression of CD44 in Canine Mammary Tumours. J. Comp. Pathol. 2009, 141, 237–247. [Google Scholar] [CrossRef]
- Regidor, P.; Callies, R.; Regidor, M.; Gunthert, U.; Zoller, M.; Schindler, A. Expression of the CD44 variant isoforms 6 and 4/5 in breast cancer. Correlation with established prognostic parameters. Arch. Gynecol. Obstet. 1966, 258, 125–135. [Google Scholar] [CrossRef]
- Hebbard, L.; Steffen, A.; Zawadzki, V.; Fieber, C.; Howells, N.; Moll, J.; Ponta, H.; Hofmann, M.; Sleeman, J. CD44 expression and regulation during mammary gland development and function. J. Cell Sci. 2000, 113, 2619–2630. [Google Scholar] [CrossRef]
- Herrlich, P.; Morrison, H.; Sleeman, J.; Orian-Rousseau, V.; Konig, H.; Weg-Remers, S.; Ponta, H. CD44 Acts Both as a Growth- and Invasiveness-Promoting Molecule and as a Tumor-Suppressing Cofactor. Ann. N. Y. Acad. Sci. 2000, 910, 106–118. [Google Scholar] [CrossRef]
- Sarli, G.; Sassi, F.; Brunetti, B.; Rizzo, A.; Diracca, L.; Benazzi, C. Lymphatic vessels assessment in feline mammary tumours. BMC Cancer 2007, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Green, K.J.; Bohringer, M.; Gocken, T.; Jones, J.C. Intermediate filament associated proteins. Adv. Protein Chem. 2005, 70, 143–202. [Google Scholar] [CrossRef]
- Gjerdrum, C.; Tiron, C.; Høiby, T.; Stefansson, I.; Haugen, H.; Sandal, T.; Collett, K.; Li, S.; McCormack, E.; Gjertsen, B.T.; et al. Axl is an essential epithelial-to-mesenchymal transition-induced regulator of breast cancer metastasis and patient survival. Proc. Natl. Acad. Sci. USA 2010, 107, 1124–1129. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, N.; Tokunaga, E.; Kitao, H.; Hisamatsu, Y.; Taketani, K.; Akiyoshi, S.; Okada, S.; Aishima, S.; Morita, M.; Maehara, Y. Vimentin as a poor prognostic factor for triple-negative breast cancer. J. Cancer Res. Clin. Oncol. 2013, 139, 739–746. [Google Scholar] [CrossRef]
- Hemalatha, A.; Suresh, T.N.; Harendra Kumar, M.L. Expression of vimentin in breast carcinoma, its correlation with Ki67 and other histopathological parameters. Indian J. Cancer 2013, 50, 189–194. [Google Scholar] [CrossRef]
- Rismanchi, S.; Yadegar, O.; Muhammadnejad, S.; Amanpour, S.; Taghizadeh-Jahed, M.; Muhammadnejad, A. Expression of vimentin filaments in canine malignant mammary gland tumors: A simulation of clinicopathological features of human breast cancer. Biomed. Rep. 2014, 2, 725–728. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Tokunaga, E.; Inoue, Y.; Yamashita, N.; Saeki, H.; Okano, S.; Kitao, H.; Oki, E.; Oda, Y.; Maehara, Y. Impact of Expression of Vimentin and Axl in Breast Cancer. Clin. Breast Cancer 2016, 16, 520–526. [Google Scholar] [CrossRef]
- Raposo-Ferreira, T.M.M.; Brisson, B.K.; Durham, A.C.; Laufer-Amorim, R.; Kristiansen, V.; Puré, E.; Volk, S.W.; Sorenmo, K. Characteristics of the Epithelial-Mesenchymal Transition in Primary and Paired Metastatic Canine Mammary Carcinomas. Vet. Pathol. 2018, 55, 622–633. [Google Scholar] [CrossRef] [Green Version]
- Valdivia, G.; Alonso-Diez, Á.; Pérez-Alenza, D.; Peña, L. From Conventional to Precision Therapy in Canine Mammary Cancer: A Comprehensive Review. Front. Vet. Sci. 2021, 8, 623800. [Google Scholar] [CrossRef]
- Kaszak, I.; Ruszczak, A.; Kanafa, S.; Kacprzak, K.; Król, M.; Jurka, P. Current biomarkers of canine mammary tumors. Acta Vet. Scand. 2018, 60, 66. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, F.; Peña, L.; Ibisch, C.; Loussouarn, D.; Gama, A.; Rieder, N.; Belousov, A.; Campone, M.; Abadie, J. Canine invasive mammary carcinomas as models of human breast cancer. Part 1: Natural history and prognostic factors. Breast Cancer Res. Treat. 2018, 167, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Abadie, J.; Nguyen, F.; Loussouarn, D.; Peña, L.; Gama, A.; Rieder, N.; Belousov, A.; Bemelmans, I.; Jaillardon, L.; Ibisch, C.; et al. Canine invasive mammary carcinomas as models of human breast cancer. Part 2: Immunophenotypes and prognostic significance. Breast Cancer Res. Treat. 2018, 167, 459–468. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HUMAN | DOG | CAT | |
|---|---|---|---|
| SNAIL1 | F: 5′-TTCTCACTGCCATGGAATTCC-3′ R: 5′-GCAGAGGACACAGAACCAGAAA-3′ | F: 5′-ACTGCAGCCGTGCCTTTG-3′ R: 5′-AAGGTTCGGGAACAGGTCTTG-3′ | F: 5′-CACCTGTTTCATGGGCAATTT-3′ R: 5′-CATCGGTCAGGCTGAAGCA-3′ |
| SNAIL2 | F: 5′-GCACACTGAGTGACGCAATCA-3′ R: 5′-AGCACAGGAGAAAATGCCTTTG-3′ | F: 5′-TTTTCTGGGCTGGCCAAA-3′ R: 5′-CGCCCAGGCTCACGTATT-3′ | F: 5′-TGCAGACCCATTCGGATGT-3′ R: 5′-CAGCAGCCAGATTCCTCATGT-3′ |
| TWIST1 | F: 5′-GTCTAGAGACTCTGGAGCTGGATAACT-3′ R: 5′-CGCCCTGTTTCTTTGAATTTG-3′ | - | F: 5′-TTAGAAGAGCAGAACCCAAAT-3′ R: 5′-CTGCCCGTCTGGGAATCA-3′ |
| TWIST2 | F: 5′-AGGACGGTCCCCACATAGG-3′ R: 5′-ACATAAGACCCAGAAGAAAAATCCA-3′ | F: 5′-CAGACACGGTCCCCACACA-3′ R: 5′-AACCCAGAAGAAAAGATCCAAACA-3′ | F: 5′-GGAAACGCGACGCTGAGT-3′ R: 5′-GGAAGCCACAGATGCACTTTG-3′ |
| ZEB1 | F: 5′-GATGATGAATGCGAGTCAGATGC-3′ R: 5′-ACAGCAGTGTCTTGTTGTTGT-3′ | F: 5′-AAAATGAGCAAAACCATGATCCTAA-3′ R: 5′-CCCTGCCTCTGGTCCTCTTC-3′ | F: 5′-CCCACACGACCACAGATAAGG-3′ R: 5′-TGAATTCATAATCCACAGGTTCA-3′ |
| ZEB2 | F: 5′-CCAGCTCGAGCGGCATA-3′ R: 5′-GCCACACTCTGTGCATTTGAA-3′ | F: 5′-TTACCCAGGTCGCCCGTAA-3′ R: 5′-TTAGCCTGAGCGGAGGATCA-3′ | F: 5′-CACGATCCAGACCGCAGTTA-3′ R: 5′-GTCGCGTTCCTCCAGTTTTC-3′ |
| ACTB | F: 5′-TGGCACCACACCTTCTACAA-3′ R: 5′-CCAGAGGCGTACAGGGATAG-3′ | F: 5′-TGGCACCACACCTTCTACAA-3′ R: 5′-CCAGAGGCGTACAGGGATAG-3′ | F: 5′-TGGCACCACACCTTCTACAA-3′ R: 5′-CCAGAGGCGTACAGGGATAG-3′ |
| IHC (Mean ± SD) | HBC ER+ | HBC TNBC | CMT STC I | CMT STC II | FMT STC III |
|---|---|---|---|---|---|
| ER | 81.7 ± 18.4 | negative | 30.4 ± 17.4 ** | 13.1 ± 16.4 ** | negative |
| PR | 57.5 ± 42.6 | negative | NP | NP | negative |
| ERBB2 | 0 or 1+ # | negative | negative | negative | negative |
| CK8/18 | 99.5 ± 1.6 *** | 53.0 ± 36.9 *** | 91.7 ± 10.4 | 70.8 ± 38.0 | 73.0 ± 19.9 |
| CK5/6 | Negative * | 14.2 ± 26.22 * | 54.9 ± 32.9 | 53.6 ± 37.5 | 26.4 ± 31.8 |
| CK14 | 0.4 ± 0.8 | 3.2 ± 6.3 | 9.6 ± 11.7 | 13.1 ± 25.4 | 51.3 ± 37.9 |
| CD44 | 20.7 ± 25.8 | 48.9 ± 38.6 | 85.7 ± 14.5 | 86.8 ± 12.4 | 4.7 ± 5.4 |
| E-cad M | 77.8 ± 20.5 *** | 24.7 ± 15.7 *** | 58.9 ± 18.8 | 66.9 ± 14.8 | 57.0 ± 23.3 |
| E-cad C | 16.4 ± 19.1 *** | 62.7 ± 19.3 *** | 35.1 ± 15.7 | 25.8 ± 13.6 | 37.9 ± 22.3 |
| Vimentin | 8.9 ± 26.7 | 23.8 ± 30.1 | 4.2 ± 9.3 | 2.5 ± 3.1 | 68.9 ± 34.3 |
| Ki-67 | NP | NP | 14.0 ± 7.0 | 18.4 ± 10.4 | 49.6 ± 13.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sammarco, A.; Gomiero, C.; Beffagna, G.; Cavicchioli, L.; Ferro, S.; Michieletto, S.; Orvieto, E.; Patruno, M.; Zappulli, V. Epithelial-to-Mesenchymal Transition and Phenotypic Marker Evaluation in Human, Canine, and Feline Mammary Gland Tumors. Animals 2023, 13, 878. https://doi.org/10.3390/ani13050878
Sammarco A, Gomiero C, Beffagna G, Cavicchioli L, Ferro S, Michieletto S, Orvieto E, Patruno M, Zappulli V. Epithelial-to-Mesenchymal Transition and Phenotypic Marker Evaluation in Human, Canine, and Feline Mammary Gland Tumors. Animals. 2023; 13(5):878. https://doi.org/10.3390/ani13050878
Chicago/Turabian StyleSammarco, Alessandro, Chiara Gomiero, Giorgia Beffagna, Laura Cavicchioli, Silvia Ferro, Silvia Michieletto, Enrico Orvieto, Marco Patruno, and Valentina Zappulli. 2023. "Epithelial-to-Mesenchymal Transition and Phenotypic Marker Evaluation in Human, Canine, and Feline Mammary Gland Tumors" Animals 13, no. 5: 878. https://doi.org/10.3390/ani13050878