
Oxidized β-Carotene Is a Novel Phytochemical Immune Modulator That Supports Animal Health and Performance for Antibiotic-Free Production


{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Antimicrobial Alternatives for Enhancement of Health and Performance
1.1. Classes of Alternatives
1.2. Immune Modulators
1.3. Phytochemicals
2. Carotenoids. Sources of Phytochemicals with a Unique Role in Health Promotion
3. Oxidized β-Carotene (OxBC). A Naturally Occurring Phytochemical Immunomodulator
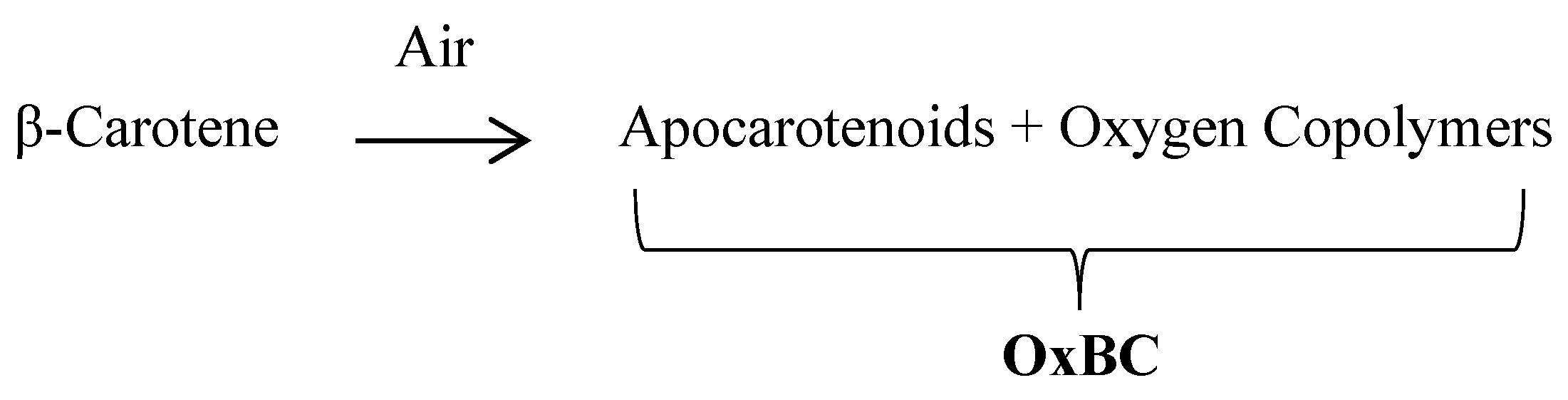
3.1. What Is OxBC? Oxygen Copolymer and Apocarotenoid Compounds
3.2. OxBC’s Dual Immunological Function
3.3. Livestock Applications. Health and Performance and Metaphylaxis of Sub-Clinical Conditions
3.4. OxBC Safety
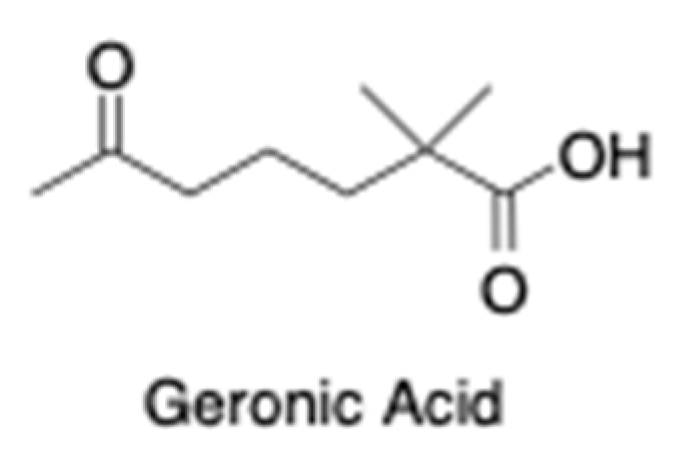
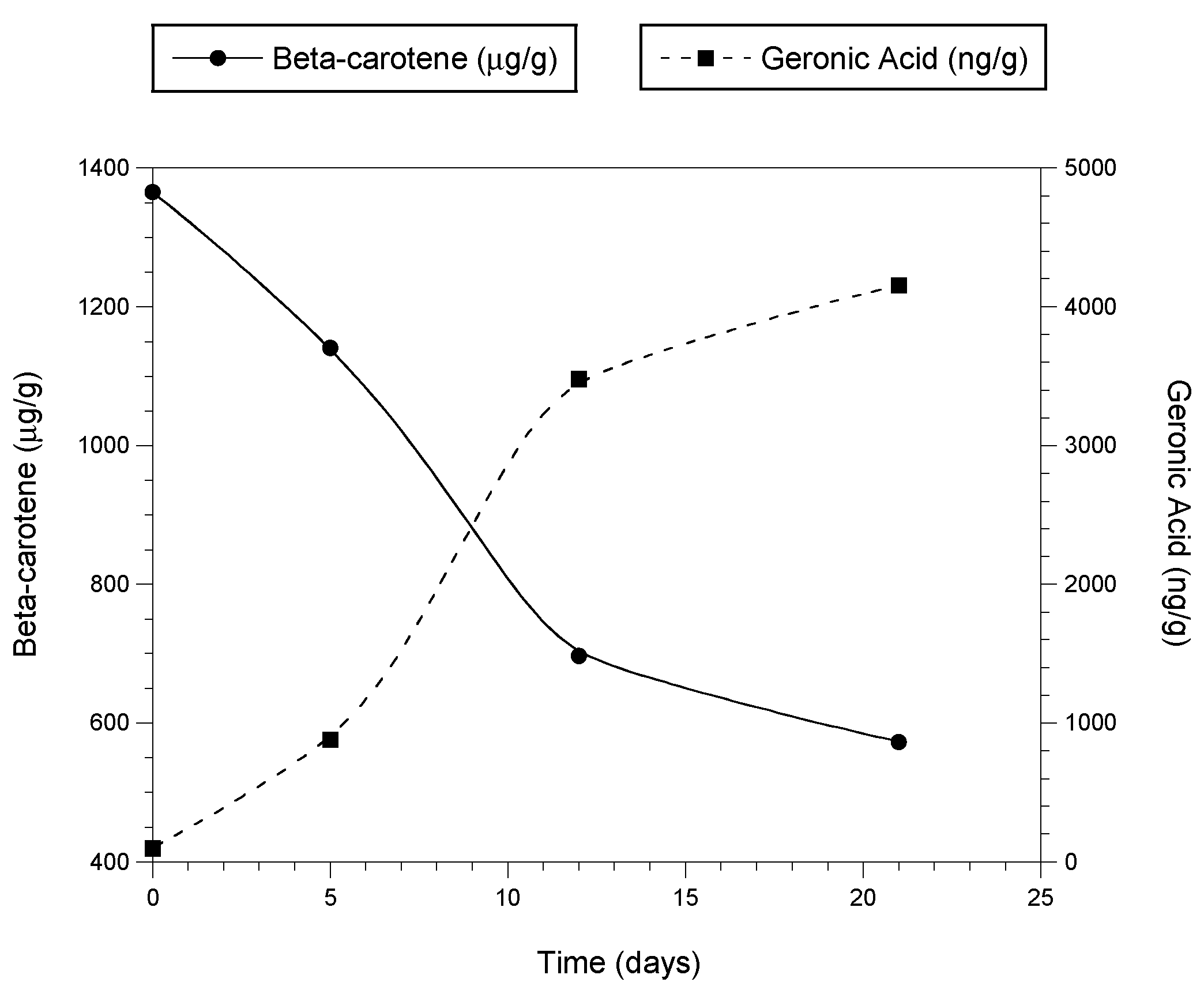
3.5. OxBC Stability
4. OxBC as an Effective Alternative to Antimicrobials in Livestock Species
4.1. Poultry
4.2. Swine
4.3. Cattle
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, Y.; Espinosa, C.D.; Abelilla, J.J.; Casas, G.A.; Lagos, L.V.; Lee, S.A.; Kwon, W.B.; Mathai, J.K.; Navarro, D.M.D.L.; Jaworski, N.W.; et al. Non-antibiotic feed additives in diets for pigs: A review. Anim. Nutr. 2018, 4, 113–125. [Google Scholar] [CrossRef]
- Tellez-Isaias, G.; Latorre, J.D. Editorial: Alternatives to Antimicrobial Growth Promoters and Their Impact in Gut Microbiota, Health and Disease: Volume II. Front. Vet. Sci. 2022, 9, 857583. [Google Scholar] [CrossRef]
- Tellez, G.; Latorre, J.D. Editorial: Alternatives to Antimicrobial Growth Promoters and Their Impact in Gut Microbiota, Health and Disease. Front. Vet. Sci. 2017, 4, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef] [Green Version]
- Lillehoj, H.; Liu, Y.; Calsamiglia, S.; Fernandez-Miyakawa, M.E.; Chi, F.; Cravens, R.L.; Oh, S.; Gay, C.G. Phytochemicals as antibiotic alternatives to promote growth and enhance host health. Vet. Res. 2018, 49, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niewold, T.A. The nonantibiotic anti-inflammatory effect of antimicrobial growth promoters, the real mode of action? A hypothesis. Poult. Sci. 2007, 86, 605–609. [Google Scholar] [CrossRef]
- Cheng, G.; Hao, H.; Xie, S.; Wang, X.; Dai, M.; Huang, L.; Yuan, Z.Y. Antibiotic alternatives: The substitution of antibiotics in animal husbandry? Front. Microbiol. 2014, 5, 217. [Google Scholar] [CrossRef] [Green Version]
- Burton, G.W.; Daroszewski, J.; Nickerson, J.G.; Johnston, J.B.; Mogg, T.J.; Nikiforov, G.B. ß-Carotene autoxidation: Oxygen copolymerization, non-vitamin A products and immunological activity. Can. J. Chem. 2014, 92, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Johnston, J.B.; Nickerson, J.G.; Daroszewski, J.; Mogg, T.J.; Burton, G.W. Biologically active polymers from spontaneous carotenoid oxidation. A new frontier in carotenoid activity. PLoS ONE 2014, 9, e111346. [Google Scholar] [CrossRef]
- Britton, G. Functions of carotenoid metabolites and breakdown products. In Carotenoids: Natural Functions; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 2008; Volume 4, pp. 309–324. [Google Scholar]
- Wang, X.-D. Biological activities of carotenoid metabolites. In Carotenoids: Nutrition and Health; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 2009; Volume 5, pp. 383–408. [Google Scholar]
- Kaulmann, A.; Bohn, T. Carotenoids, inflammation, and oxidative stress--implications of cellular signaling pathways and relation to chronic disease prevention. Nutr. Res. 2014, 34, 907–929. [Google Scholar] [CrossRef]
- Sharoni, Y.; Linnewiel-Hermoni, K.; Khanin, M.; Salman, H.; Veprik, A.; Danilenko, M.; Levy, J. Carotenoids and apocarotenoids in cellular signaling related to cancer: A review. Mol. Nutr. Food Res. 2012, 56, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Kulczyński, B.; Gramza-Michałowska, A.; Kobus-Cisowska, J.; Kmiecik, D. The role of carotenoids in the prevention and treatment of cardiovascular disease—Current state of knowledge. J. Funct. Foods 2017, 38, 45–65. [Google Scholar] [CrossRef]
- Akhtar, S.; Ahmed, A.; Randhawa, M.A.; Atukorala, S.; Arlappa, N.; Ismail, T.; Ali, Z. Prevalence of vitamin A deficiency in South Asia: Causes, outcomes, and possible remedies. J. Health Popul. Nutr. 2013, 31, 413–423. [Google Scholar] [CrossRef] [Green Version]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Editors’ assessment. In Carotenoids. Nutrition and Health; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 2009; Volume 5, pp. 409–422. [Google Scholar]
- Britton, G. Functions of intact carotenoids. In Carotenoids: Natural Functions; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 2008; Volume 4, pp. 189–212. [Google Scholar]
- Chung, R.W.S.; Leanderson, P.; Lundberg, A.K.; Jonasson, L. Lutein exerts anti-inflammatory effects in patients with coronary artery disease. Atherosclerosis 2017, 262, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chew, B.; Park, J. The Immune System. In Carotenoids: Nutrition and Health; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 2009; Volume 5, pp. 363–382. [Google Scholar]
- Blount, J.D.; Metcalfe, N.B.; Birkhead, T.R.; Surai, P.F. Carotenoid modulation of immune function and sexual attractiveness in zebra finches. Science 2003, 300, 125–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGraw, K.J.; Ardia, D.R. Carotenoids, Immunocompetence, and the Information Content of Sexual Colors: An Experimental Test. Am. Nat. 2003, 162, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Chew, B.P.; Park, J.S. Carotenoid action on the immune response. J. Nutr. 2004, 134, 257S–261S. [Google Scholar] [CrossRef] [Green Version]
- Leclaire, S.; Bourret, V.; Blanchard, P.; de Franceschi, C.; Merkling, T.; Hatch, S.A.; Danchin, É. Carotenoids increase immunity and sex specifically affect color and redox homeostasis in a monochromatic seabird. Behav. Ecol. Sociobiol. 2015, 69, 1097–1111. [Google Scholar] [CrossRef]
- Lopez-Rull, I.; Hornero-Mendez, D.; Frias, O.; Blanco, G. Age-Related Relationships between Innate Immunity and Plasma Carotenoids in an Obligate Avian Scavenger. PLoS ONE 2015, 10, e0141759. [Google Scholar] [CrossRef] [PubMed]
- Koutsos, E.A.; Calvert, C.C.; Klasing, K.C. The effect of an acute phase response on tissue carotenoid levels of growing chickens (Gallus gallus domesticus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2003, 135, 635–646. [Google Scholar] [CrossRef]
- Koutsos, E.A.; Garcia Lopez, J.C.; Klasing, K.C. Carotenoids from in ovo or dietary sources blunt systemic indices of the inflammatory response in growing chicks (Gallus gallus domesticus). J. Nutr. 2006, 136, 1027–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepys, M.B.; Baltz, M.L.; Tennent, G.A.; Kent, J.; Ousey, J.; Rossdale, P.D. Serum amyloid A protein (SAA) in horses: Objective measurement of the acute phase response. Equine Vet. J. 1989, 21, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Ceron, J.J.; Eckersall, P.D.; Martynez-Subiela, S. Acute phase proteins in dogs and cats: Current knowledge and future perspectives. Vet. Clin. Pathol. 2005, 34, 85–99. [Google Scholar] [CrossRef]
- Caliendo, V.; McKinney, P.; Bailey, T.; Kinne, J.; Wernery, U. Serum amyloid A as an indicator of health status in falcons. J. Avian Med. Surg. 2013, 27, 83–89. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific Opinion on the safety and efficacy of beta-carotene as a feed additive for all animal species and categories. EFSA J. 2012, 10, 2737. [Google Scholar] [CrossRef]
- Miller, A.A.; Mayo, F.R. Oxidation of unsaturated compounds. I. The oxidation of styrene. J. Am. Chem. Soc. 1956, 78, 1017–1023. [Google Scholar] [CrossRef]
- Handelman, G.J.; van Kuijk, F.J.G.M.; Chatterjee, A.; Krinsky, N.I. Characterization of products formed during the autoxidation of ß-carotene. Free Rad. Biol. Med. 1991, 10, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Mordi, R.C.; Walton, J.C.; Burton, G.W.; Hughes, L.; Ingold, K.U.; Lindsay, D.A.; Moffatt, D.J. Oxidative degradation of ß-carotene and ß-apo-8’-carotenal. Tetrahedron 1993, 49, 911–928. [Google Scholar] [CrossRef]
- Crouzet, J.; Kanasawud, P.; Sakho, M. Thermal Generation of Carotenoid-Derived Compounds. In Carotenoid-Derived Aroma Compounds; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2001; Volume 802, pp. 115–129. [Google Scholar]
- Burton, G.W.; Daroszewski, J.; Mogg, T.J.; Nikiforov, G.B.; Nickerson, J.G. Discovery and characterization of carotenoid-oxygen copolymers in fruits and vegetables with potential health benefits. J. Agric. Food Chem. 2016, 64, 3767–3777. [Google Scholar] [CrossRef] [Green Version]
- Schaub, P.; Wust, F.; Koschmieder, J.; Yu, Q.; Virk, P.; Tohme, J.; Beyer, P. Nonenzymatic beta-carotene degradation in provitamin A-biofortified crop plants. J. Agric. Food Chem. 2017, 65, 6588–6598. [Google Scholar] [CrossRef]
- Mogg, T.J.; Burton, G.W. The β-carotene–oxygen copolymer: Its relationship to apocarotenoids and β-carotene function. Can. J. Chem. 2021, 99, 751–762. [Google Scholar] [CrossRef]
- Burton, G.W.; Daroszewski, J.; Mogg, T.J.; Nikiforov, G.B.; Nickerson, J.G.; Groome, C.L. Plant or Microorganism-Derived Carotenoid-Oxygen Copolymer Compositions, Methods of Identifying, Quantifying and Producing Same and Uses Thereof. World Intellectual Property Organization. WO 2017/143460, 31 August 2017. [Google Scholar]
- Winterhalter, P.; Rouseff, R.L. (Eds.) Carotenoid-Derived Aroma Compounds; American Chemical Society: Washington, DC, USA, 2001; Volume 802. [Google Scholar]
- Melendez-Martinez, A.J. An Overview of Carotenoids, Apocarotenoids, and Vitamin A in Agro-Food, Nutrition, Health, and Disease. Mol. Nutr. Food Res. 2019, 63, e1801045. [Google Scholar] [CrossRef] [Green Version]
- U.S. Food & Drug Administration. Substances Added to Food. Available online: https://www.cfsanappsexternal.fda.gov/scripts/fdcc/index.cfm?set=FoodSubstances (accessed on 15 October 2022).
- Duquette, S.C.; Fischer, C.D.; Feener, T.D.; Muench, G.P.; Morck, D.W.; Barreda, D.R.; Nickerson, J.G.; Buret, A.G. Anti-inflammatory benefits of retinoids and carotenoid derivatives: Retinoic acid and fully oxidized β-carotene induce caspase-3-dependent apoptosis and promote efferocytosis of bovine neutrophils. Am. J. Vet. Res. 2014, 75, 1064–1075. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, J.; Zhang, Y.; Lv, J.; Qiao, H.; Tian, M.; Cheng, L.; Chen, F.; Zhang, S.; Guan, W. Effects of maternal supplementation with fully oxidised β-carotene on the reproductive performance and immune response of sows, as well as the growth performance of nursing piglets. Brit. J. Nutr. 2021, 125, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Aloum, L.; Alefishat, E.; Adem, A.; Petroianu, G. Ionone Is More than a Violet’s Fragrance: A Review. Molecules 2020, 25, 5822. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.W.; Mogg, T.J.; Stupak, J.; Stark, F.C.; Twine, S.M.; Li, J. Safety and uptake of fully oxidized beta-carotene. Food Chem. Toxicol. 2022, 168, 113387. [Google Scholar] [CrossRef] [PubMed]
- Hurnik, D.; Daroszewski, J.; Burton, G.W. Determination of the effect of a fully oxidized β-carotene dietary supplement on the immune system and growth performance of weaned pigs. In Proceedings of the American Association of Swine Veterinarians 42nd Annual Meeting, Phoenix, Arizona, 5–8 March 2011; pp. 277–279. [Google Scholar]
- Kinh, L.V.; Riley, W.W.; Nickerson, J.G.; Huyen, L.T.T.; Burton, G.W. Effect of oxidized β-carotene on swine growth performance under commercial production conditions in Vietnam. Animals 2022, 12, 3200. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Oh, J.Y.; Cha, S.Y.; Kim, W.I.; Cho, H.S.; Jang, H.K. Efficacy of polymers from spontaneous carotenoid oxidation in reducing necrotic enteritis in broilers. Poult. Sci. 2018, 97, 3058–3062. [Google Scholar] [CrossRef]
- McDougall, S. Evaluation of fully oxidised β-carotene as a feed ingredient to reduce bacterial infection and somatic cell counts in pasture-fed cows with subclinical mastitis. N.Z. Vet. J. 2021, 69, 285–293. [Google Scholar] [CrossRef]
- Khadem, A.; Soler, L.; Everaert, N.; Niewold, T.A. Growth promotion in broilers by both oxytetracycline and Macleaya cordata extract is based on their anti-inflammatory properties. Br. J. Nutr. 2014, 112, 1110–1118. [Google Scholar] [CrossRef]
- Niewold, T.A. Organic more healthy? Green shoots in a scientific semi-desert. Br. J. Nutr. 2010, 103, 627–628. [Google Scholar] [CrossRef] [Green Version]
- Soler, L.; Miller, I.; Hummel, K.; Razzazi-Fazeli, E.; Jessen, F.; Escribano, D.; Niewold, T. Growth promotion in pigs by oxytetracycline coincides with down regulation of serum inflammatory parameters and of hibernation-associated protein HP-27. Electrophoresis 2016, 37, 1277–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ATBC Cancer Prevention Study Group. The effect of vitamin E and beta carotene on the incidence of lung cancer and other cancers in male smokers. N. Engl. J. Med. 1994, 330, 1029–1035. [Google Scholar] [CrossRef]
- Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Meyskens, F.L., Jr.; Omenn, G.S.; Valanis, B.; Williams, J.H., Jr. The beta-carotene and retinol efficacy trial: Incidence of lung cancer and cardiovascular disease mortality during 6-year follow-up after stopping beta-carotene and retinol supplements. J. Natl. Cancer Inst. 2004, 96, 1743–1750. [Google Scholar] [CrossRef] [Green Version]
- Omenn, G.S.; Goodman, G.; Thornquist, M.; Grizzle, J.; Rosenstock, L.; Barnhart, S.; Balmes, J.; Cherniack, M.G.; Cullen, M.R.; Glass, A.; et al. The beta-carotene and retinol efficacy trial (CARET) for chemoprevention of lung cancer in high risk populations: Smokers and asbestos-exposed workers. Cancer Res. 1994, 54, 2038s–2043s. [Google Scholar]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L., Jr.; Valanis, B.; Williams, J.H., Jr.; et al. Risk factors for lung cancer and for intervention effects in CARET, the Beta-Carotene and Retinol Efficacy Trial. J. Natl. Cancer Inst. 1996, 88, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L.; Valanis, B.; Williams, J.H.; et al. Effects of a Combination of Beta Carotene and Vitamin A on Lung Cancer and Cardiovascular Disease. New Engl. J. Med. 1996, 334, 1150–1159. [Google Scholar] [CrossRef] [Green Version]
- Virtamo, J.; Taylor, P.R.; Kontto, J.; Mannisto, S.; Utriainen, M.; Weinstein, S.J.; Huttunen, J.; Albanes, D. Effects of alpha-tocopherol and beta-carotene supplementation on cancer incidence and mortality: 18-year postintervention follow-up of the Alpha-tocopherol, Beta-carotene Cancer Prevention Study. Int. J. Cancer 2014, 135, 178–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Food Safety Authority. Scientific Opinion on the re-evaluation of mixed carotenes (E 160a (i)) and beta-carotene (E 160a (ii)) as a food additive. EFSA Panel on Food Additives and Nutrient Sources added to Food. EFSA J. 2012, 10, 2593. [Google Scholar] [CrossRef]
- Burton, G.W.; Mogg, T.J.; Riley, W.W.; Nickerson, J.G. beta-Carotene oxidation products—Function and safety. Food Chem. Toxicol. 2021, 152, 112207. [Google Scholar] [CrossRef]
- Kogut, M.H. Impact of nutrition on the innate immune response to infection in poultry. J. Appl. Poult. Res. 2009, 18, 111–124. [Google Scholar] [CrossRef]
- Diaz-Sanchez, S.; Moscoso, S.; Solίs de los Santos, F.; Andino, A.; Hanning, I. Antibiotic use in poultry: A driving force for organic poultry production. Food Prot. Trends 2015, 35, 440–447. [Google Scholar]
- Riley, W.W.; Nickerson, J.G.; Burton, G.W. Effect of oxidized β-carotene on the growth and feed efficiency of broilers. Poult. Sci. 2021, 100, 101088. [Google Scholar] [CrossRef] [PubMed]
- Butaye, P.; Devriese, L.A.; Haesebrouck, F. Antimicrobial growth promoters used in animal feed: Effects of less well known antibiotics on gram-positive bacteria. Clin. Microbiol. Rev. 2003, 16, 175–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, A.; Phillips, G.J. Differential Effects of Bacitracin Methylene Disalicylate (BMD) on the Distal Colon and Cecal Microbiota of Young Broiler Chickens. Front. Vet. Sci. 2019, 6, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet. Scand. 2017, 59, 31. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Liu, Y.; Dong, W.; Zhu, G.Q.; Wu, S.; Bao, W. CD14 in the TLRs signaling pathway is associated with the resistance to E. coli F18 in Chinese domestic weaned piglets. Sci. Rep.-UK 2016, 6, 24611. [Google Scholar] [CrossRef] [Green Version]
- Patience, J.F.; Rossoni-Serao, M.C.; Gutierrez, N.A. A review of feed efficiency in swine: Biology and application. J. Anim. Sci. Biotechnol. 2015, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Das, U.N. Metabolic syndrome is a low-grade systemic inflammatory condition. Expert Rev. Endocrinol. Metab. 2010, 5, 577–592. [Google Scholar] [CrossRef]
- Halasa, T.; Huijps, K.; Osteras, O.; Hogeveen, H. Economic effects of bovine mastitis and mastitis management: A review. Vet. Q. 2007, 29, 18–31. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riley, W.W.; Nickerson, J.G.; Mogg, T.J.; Burton, G.W. Oxidized β-Carotene Is a Novel Phytochemical Immune Modulator That Supports Animal Health and Performance for Antibiotic-Free Production. Animals 2023, 13, 289. https://doi.org/10.3390/ani13020289
Riley WW, Nickerson JG, Mogg TJ, Burton GW. Oxidized β-Carotene Is a Novel Phytochemical Immune Modulator That Supports Animal Health and Performance for Antibiotic-Free Production. Animals. 2023; 13(2):289. https://doi.org/10.3390/ani13020289
Chicago/Turabian StyleRiley, William W., James G. Nickerson, Trevor J. Mogg, and Graham W. Burton. 2023. "Oxidized β-Carotene Is a Novel Phytochemical Immune Modulator That Supports Animal Health and Performance for Antibiotic-Free Production" Animals 13, no. 2: 289. https://doi.org/10.3390/ani13020289