
Cryopreservation of Semen in Domestic Animals: A Review of Current Challenges, Applications, and Prospective Strategies

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Cryobiology of Sperm
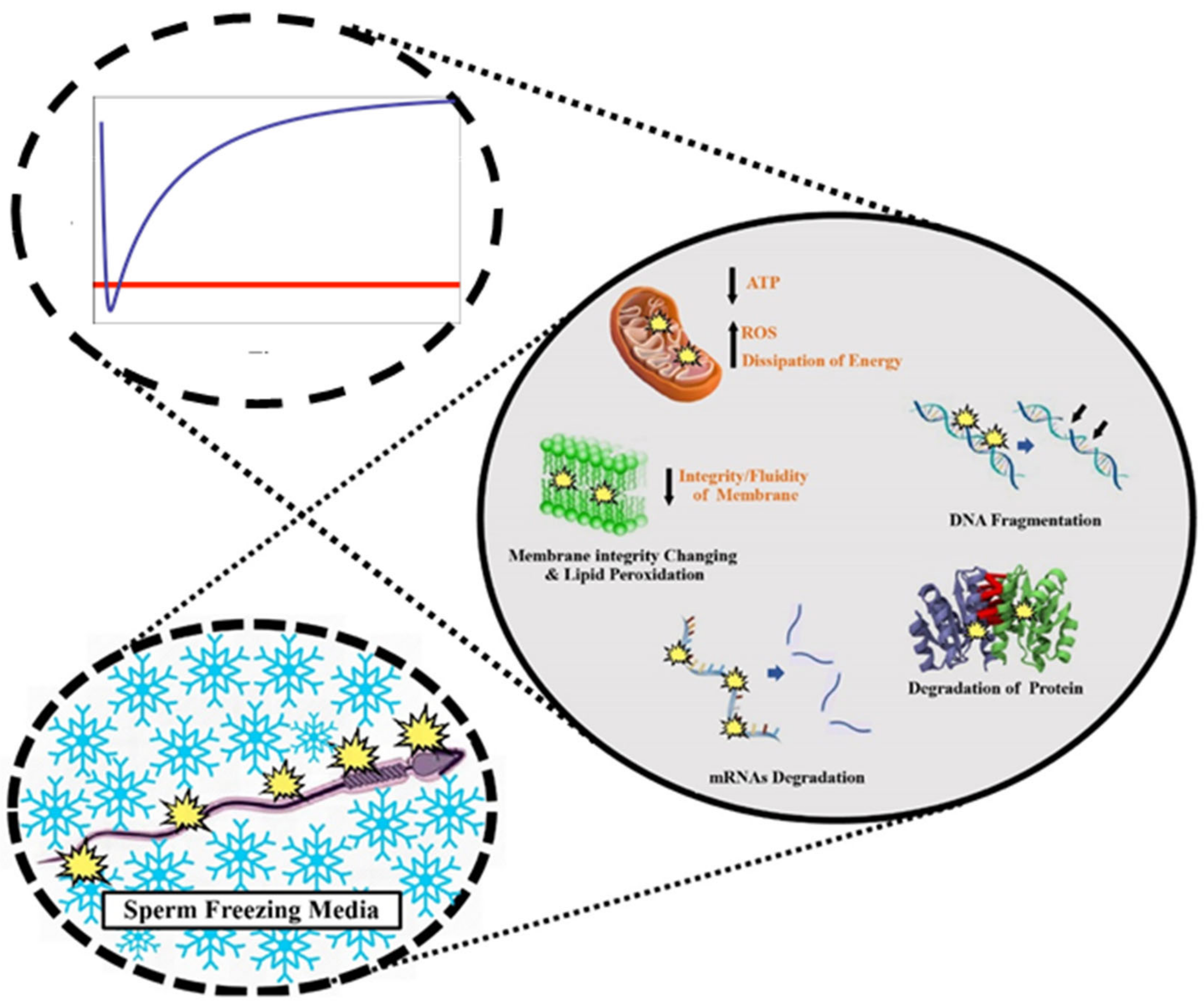
3. Cryopreservation Injuries in Sperm
3.1. Changes in Sperm Plasma Membrane
3.2. Changes in Sperm Plasma Membrane
3.3. Proteome Alterations
3.4. Epigenetic Modifications
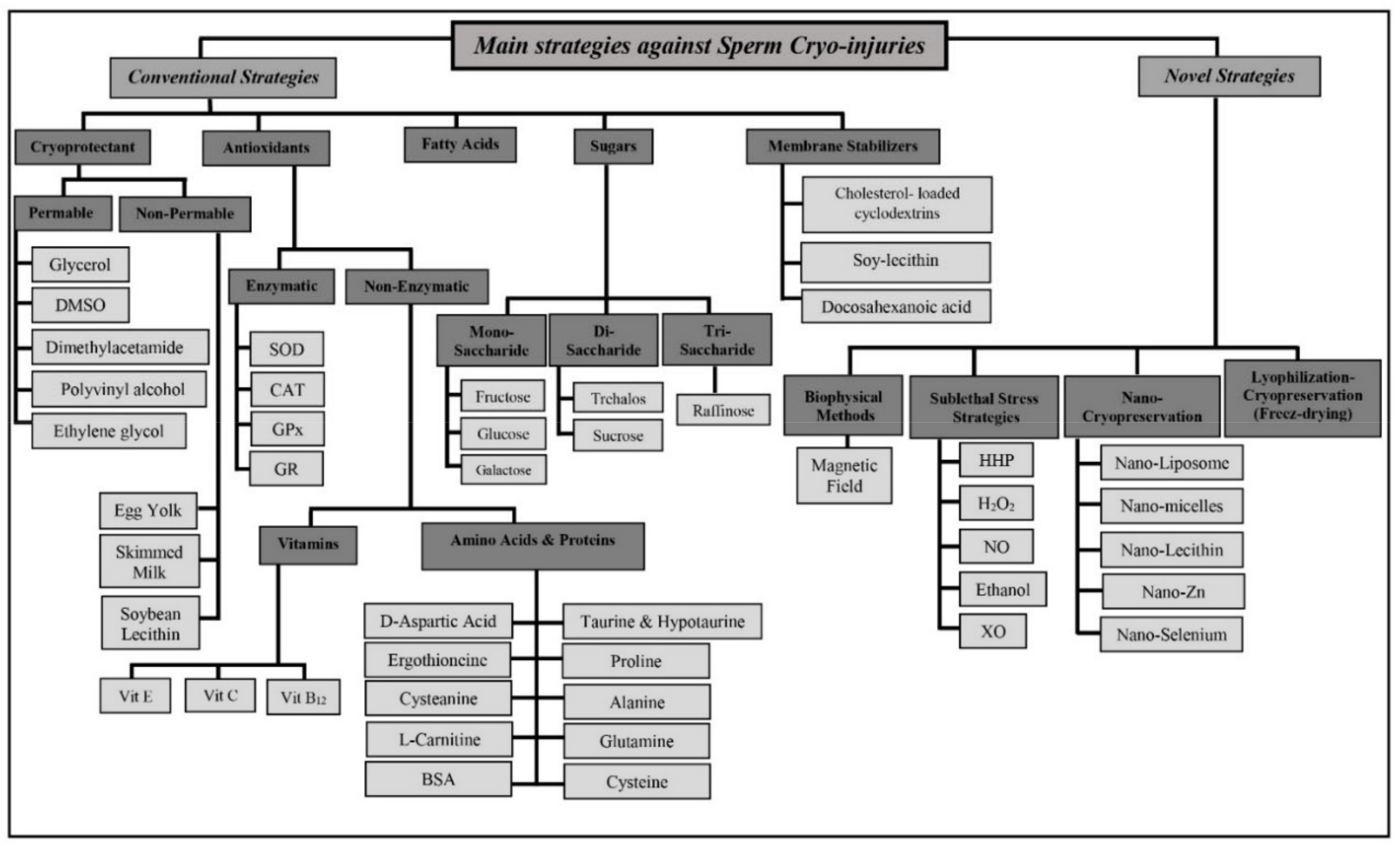
4. Prevention of Sperm Cryoinjury
4.1. Conventional Strategies
4.1.1. Cryoprotectants
4.1.2. Antioxidants
4.1.3. Sugars
4.1.4. Membrane Stabilizers
4.1.5. Using Dietary Additives
4.1.6. Warming/Thawing
4.2. Novel Strategies
4.2.1. Induction of Mild Sublethal Stress before Freezing
4.2.2. Induction of a Magnetic Field before Freezing
4.2.3. Nanoparticle-Enhanced Cryopreservation Media
4.2.4. Sperm Preservation via Freeze-Drying (Lyophilization)
4.2.5. Monolayer Centrifugation
5. Sperm Cryobanking Expansion in Research and Industry
6. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Curry, M.R. Cryopreservation of mammalian semen. Methods Mol. Biol. 2007, 368, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Vishwanath, R. The effect of optimal and suboptimal concentrations of sperm on the fertility of fresh and frozen bovine semen and a theoretical model to explain the fertility differences. Anim. Reprod. Sci. 1995, 39, 1–10. [Google Scholar] [CrossRef]
- Crabo, B. Preservation of boar semen: A worldwide perspective. Reprod. Dom. Anim., Suppl 1990, 1, 3–9. [Google Scholar]
- Maxwell, W.M.C.; Evans, G.; Rhodes, S.L.; Hillard, M.A.; Bindon, B.M. Fetility of superovulated ewes after intrauterine or oviducal insemination with low numbers of fresh or frozen-thawed spermatozoa. Reprod. Fertil. Dev. 1993, 5, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Spallanzani, L. Opuscoli di fisca, Anim. e Veg. Opusculo I I. Oss. e Sperienze Intorno Ai Vermicelli Spermatici Dell’Uomo e Degli Anim. Modena 1776, 15–109. [Google Scholar]
- Ombelet, W.; Van Robays, J. History of human artificial insemination. F V V ObGyn 2010, 1–5. [Google Scholar]
- Foote, R. The history of artificial insemination: Selected notes and notables. J. Anim. Sci. 2010, 80, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Herman, H.A. Improving cattle by the millions. In NAAB and the Development and Worldwide Application of Artificial Insemination; University of Missouri Press: London, UK, 1981. [Google Scholar]
- Sherman, J.K. Research on frozen human semen: Past, present, and future. Fertil. Steril. 1964, 15, 485–500. [Google Scholar] [CrossRef]
- Polge, C.; Smith, A.U.; Parkes, A.S. Revival of spermatozoa after vitrification and dehydration at low temperatures. Nature 1949, 164, 666. [Google Scholar] [CrossRef]
- Stewart, D. Storage of bull spermatozoa at low temperature. Vet. Rec. 1951, 63, 65–66. [Google Scholar]
- Mulder, R.L.; Font-Gonzalez, A.; Green, D.M.; Loeffen, E.A.H.; Jacqueline, M.M.H.; Loonen, J.; Yu, R.C.; Ginsberg, J.P.; Mitchell, R.T.; Byrne, J.; et al. Fertility Preservation in Childhood, Adolescent, and Young Adult Cancer 2 Fertility preservation for male patients with childhood, adolescent, and young adult cancer: Recommendations from the PanCareLIFE Consortium and the International Late Effects of Childhood Cancer Guideline Harmonization Group. Lancet Oncol. 2021, 22, E57–E67. [Google Scholar] [CrossRef]
- Rodriguez-Wallberg, K.A.; Haljestig, J.; Arver, S.; Johansson, A.L.V.; Lundberg, F.E. Sperm quality in transgender women before or after gender affirming hormone therapy-A prospective cohort study. Andrology 2021, 9, 1773–1780. [Google Scholar] [CrossRef]
- Shaffner, C. Longevity of fowl spermatozoa in frozen condition. Science 1942, 96, 337. [Google Scholar] [CrossRef]
- Bunge, R.G.; Sherman, J.K. Fertilizing capacity of frozen human spermatozoa. Nature 1953, 172, 767–768. [Google Scholar] [CrossRef]
- Hess, E.; Teague, H.; Ludwick, T.; Martig, R. Swine can be bred with frozen semen. Ohio Farm. Home Res. 1957, 42, 100. [Google Scholar]
- Barker, C.; Gandier, J. Pregnancy in a mare resulting from frozen epididymal spermatozoa. Can. J. Comp. Med. Vet. Sci. 1957, 21, 47. [Google Scholar]
- Salamon, S.; Lightfoot, R.J. Fertilization and embryonic loss in sheep after insemination with deep frozen semen. Nature 1967, 216, 194–195. [Google Scholar] [CrossRef]
- Watson, P.F. The causes of reduced fertility with cryopreserved semen. Anim. Reprod. Sci. 2000, 60, 481–492. [Google Scholar] [CrossRef]
- Parks, J.E.; Graham, J.K. Effects of cryopreservation procedures on sperm membranes. Theriogenology 1992, 38, 209–222. [Google Scholar] [CrossRef]
- Johnson, L.A.; Weitze, K.F.; Fiser, P.; Maxwell, W.M.C. Storage of boar semen. Anim. Reprod. Sci. 2000, 62, 143–172. [Google Scholar] [CrossRef]
- Mazur, P.; Fuller, B.J. Life in the frozen state. In Principles of Cryobiology; CRC Press: Boca Raton, FL, USA; Academic Press: Boca Raton, FL, USA, 2004; pp. 3–65. [Google Scholar]
- Bucak, N.; Erdoğan, C.; İli, A.; Başpınar, N.; Dursun, Ş.; Güngör, Ş. Effects of cryoprotectants and trehalose on electron microscopic evaluation of cryopreserved semen. J. Reprod. Infertil. 2017, 18, 25–26. [Google Scholar]
- Zhmakin, A.I. Physical aspects of cryobiology. Phys. Uspekhi 2008, 51, 231. [Google Scholar] [CrossRef]
- Pegg, D.E. Principles of Cryopreservation. In Cryopreservation and Freeze—Drying Protocols, 3rd ed.; Wolkers, W.F., Oldenhof, H., Eds.; Methods in Molecular Biology; Humana Press Inc: Totowa, NJ, USA, 2015; Volume 1257, pp. 3–19. [Google Scholar]
- Morris, G.J.; Faszer, K.; Green, J.; Draper, D.; Grout, B.; Fonseca, F. Rapidly cooled horse spermatozoa: Loss of viability is due to osmotic imbalance during thawing, not intracellular ice formation. Theriogenology 2007, 68, 804–812. [Google Scholar] [CrossRef] [PubMed]
- Devireddy, R.; Swanlund, D.; Olin, T.; Vincente, W.; Troedsson, M.; Bischof, J.; Roberts, K. Cryopreservation of equine sperm: Optimal cooling rates in the presence and absence of cryoprotective agents determined using differential scanning calorimetry. Biol. Reprod. 2002, 66, 222–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur, P.; Leibo, S.; Chu, E. A two-factor hypothesis of freezing injury: Evidence from Chinese hamster tissue-culture cells. Exp. Cell Res. 1972, 71, 345–355. [Google Scholar] [CrossRef]
- Toner, M.; Cravalho, E.; Armant, D. Water transport and estimated transmembrane potential during freezing of mouse oocytes. J. Membr. Biol. 1990, 115, 261–272. [Google Scholar] [CrossRef]
- Hammerstedt, R.H.; Graham, J.K.; Nolan, J.P. Cryopreservation of mammalian sperm: What we ask them to survive. J. Androl. 1990, 11, 73–88. [Google Scholar] [CrossRef]
- Oldenhof, H.; Gojowsky, M.; Wang, S.P.; Henke, S.; Yu, C.J.; Rohn, K.; Wolkers, W.F.; Sieme, H. Osmotic Stress and Membrane Phase Changes During Freezing of Stallion Sperm: Mode of Action of Cryoprotective Agents. Biol. Reprod. 2013, 88, 11. [Google Scholar] [CrossRef]
- Holt, W.; North, R. Effects of temperature and restoration of osmotic equilibrium during thawing on the induction of plasma membrane damage in cryopreserved ram spermatozoa. Biol. Reprod. 1994, 51, 414–424. [Google Scholar] [CrossRef] [Green Version]
- Noiles, E.E.; Bailey, J.L.; Storey, B.T. The temperature dependence in the hydraulic conductivity, Lp, of the mouse sperm plasma membrane shows a discontinuity between 4 and 0 °C. Cryobiology 1995, 32, 220–238. [Google Scholar] [CrossRef]
- Lovelock, J. The haemolysis of human red blood-cells by freezing and thawing. Biochim. Biophys. Acta 1953, 10, 414–426. [Google Scholar] [CrossRef]
- Sieme, H.; Oldenhof, H.; Wolkers, W. Sperm membrane behaviour during cooling and cryopreservation. Reprod. Domest. Anim. 2015, 50, 20–26. [Google Scholar] [CrossRef]
- De Leeuw, F.; De Leeuw, A.; Den Daas, J.; Colenbrander, B.; Verkleij, A. Effects of various cryoprotective agents and membrane-stabilizing compounds on bull sperm membrane integrity after cooling and freezing. Cryobiology 1993, 30, 32–44. [Google Scholar] [CrossRef]
- Dufourc, E.J. Sterols and membrane dynamics. J. Chem. Biol. 2008, 1, 63–77. [Google Scholar] [CrossRef] [Green Version]
- Holt, W. Basic aspects of frozen storage of semen. Anim. Reprod. Sci. 2000, 62, 3–22. [Google Scholar] [CrossRef]
- Vadnais, M.L.; Althouse, G.C. Characterization of capacitation, cryoinjury, and the role of seminal plasma in porcine sperm. Theriogenology 2011, 76, 1508–1516. [Google Scholar] [CrossRef]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef]
- Casas, I.; Flores, E. Gene banking: The freezing strategy. In Boar Reproduction; Springer: Berlin/Heidelberg, Germany, 2013; pp. 551–588. [Google Scholar]
- Paoli, D.; Lombardo, F.; Lenzi, A.; Gandini, L. Sperm cryopreservation: Effects on chromatin structure. Genet. Damage Hum. Spermatozoa 2014, 791, 137–150. [Google Scholar] [CrossRef]
- Ward, W.S. Function of sperm chromatin structural elements in fertilization and development. MHR Basic Sci. Reprod. Med. 2009, 16, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Gosálvez, J.; López-Fernández, C.; Fernández, J.L.; Gouraud, A.; Holt, W.V. Relationships between the dynamics of iatrogenic DNA damage and genomic design in mammalian spermatozoa from eleven species. Mol. Reprod. Dev. 2011, 78, 951–961. [Google Scholar] [CrossRef]
- Irvine, D.S.; Twigg, J.P.; Gordon, E.L.; Fulton, N.; Milne, P.A.; Aitken, R.J. DNA integrity in human spermatozoa: Relationships with semen quality. J. Androl. 2000, 21, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Restrepo, G.; Varela, E.; Duque, J.E.; Gomez, J.E.; Rojas, M. Freezing, Vitrification, and Freeze-Drying of Equine Spermatozoa: Impact on Mitochondrial Membrane Potential, Lipid Peroxidation, and DNA Integrity. J. Equine Vet. Sci. 2019, 72, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Kurland, C.G.; Andersson, S.G.E. Origin and evolution of the mitochondrial proteome. Microbiol. Mol. Biol. Rev. 2000, 64, 786–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.X.; Wang, K.; Zhao, X.X.; Zhang, Y.; Ma, Y.J.; Hu, J.J. Differential proteome association study of freeze-thaw damage in ram sperm. Cryobiology 2016, 72, 60–68. [Google Scholar] [CrossRef]
- Hezavehei, M.; Sharafi, M.; Kouchesfahani, H.M.; Henkel, R.; Agarwal, A.; Esmaeili, V.; Shahverdi, A. Sperm cryopreservation: A review on current molecular cryobiology and advanced approaches. Reprod. Biomed. Online 2018, 37, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Bogle, O.A.; Kumar, K.; Attardo-Parrinello, C.; Lewis, S.E.M.; Estanyol, J.M.; Ballesca, J.L.; Oliva, R. Identification of protein changes in human spermatozoa throughout the cryopreservation process. Andrology 2017, 5, 10–22. [Google Scholar] [CrossRef]
- Parrilla, I.; Perez-Patino, C.; Li, J.; Barranco, I.; Padilla, L.; Rodriguez-Martinez, H.; Martinez, E.A.; Roca, J. Boar semen proteomics and sperm preservation. Theriogenology 2019, 137, 23–29. [Google Scholar] [CrossRef]
- Chen, X.L.; Zhu, H.B.; Hu, C.H.; Hao, H.S.; Zhang, J.F.; Li, K.P.; Zhao, X.M.; Qin, T.; Zhao, K.; Zhu, H.S.; et al. Identification of differentially expressed proteins in fresh and frozen-thawed boar spermatozoa by iTRAQ-coupled 2D LC-MS/MS. Reproduction 2014, 147, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, D.B.; Barros, T.B.; van Tilburg, M.F.; Martins, J.A.M.; Moura, A.A.; Moreno, F.B.; Monteiro-Moreira, A.C.; Moreira, R.A.; Toniolli, R. Sperm membrane proteins associated with the boar semen cryopreservation. Anim. Reprod. Sci. 2017, 183, 27–38. [Google Scholar] [CrossRef]
- Nynca, J.; Arnold, G.J.; Frohlich, T.; Ciereszko, A. Cryopreservation-induced alterations in protein composition of rainbow trout semen. Proteomics 2015, 15, 2643–2654. [Google Scholar] [CrossRef]
- Wojtusik, J.; Wang, Y.; Pukazhenthi, B.S. Pretreatment with cholesterol-loaded cyclodextrins prevents loss of motility associated proteins during cryopreservation of addra gazelle (Nanger dama ruficollis) spermatozoa. Cryobiology 2018, 81, 74–80. [Google Scholar] [CrossRef]
- Ryu, D.Y.; Song, W.H.; Pang, W.K.; Yoon, S.J.; Rahman, M.S.; Pang, M.G. Freezability biomarkers in bull epididymal spermatozoa. Sci. Rep. 2019, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Hinsch, K.D.; De Pinto, V.; Aires, V.A.; Schneider, X.; Messina, A.; Hinsch, E. Voltage-dependent anion-selective channels VDAC2 and VDAC3 are abundant proteins in bovine outer dense fibers, a cytoskeletal component of the sperm flagellum. J. Biol. Chem. 2004, 279, 15281–15288. [Google Scholar] [CrossRef] [Green Version]
- Hemachand, T.; Shaha, C. Functional role of sperm surface glutathione S-transferases and extracellular glutathione in the haploid spermatozoa under oxidative stress. FEBS Lett. 2003, 538, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Gaitskell-Phillips, G.; Martin-Cano, F.E.; Ortiz-Rodriguez, J.M.; Silva-Rodriguez, A.; Gil, M.C.; Ortega-Ferrusola, C.; Pena, F.J. Differences in the proteome of stallion spermatozoa explain stallion-to-stallion variability in sperm quality post-thaw. Biol. Reprod. 2021, 104, 1097–1113. [Google Scholar] [CrossRef]
- O’Donnell, L.; O’Bryan, M.K. Microtubules and spermatogenesis. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2014; pp. 45–54. [Google Scholar]
- Cheng, C.Y.; Chen, P.R.; Chen, C.J.; Wang, S.H.; Chen, C.F.; Lee, Y.P.; Huang, S.Y. Differential protein expression in chicken spermatozoa before and after freezing-thawing treatment. Anim. Reprod. Sci. 2015, 152, 99–107. [Google Scholar] [CrossRef]
- Urrego, R.; Rodriguez-Osorio, N.; Niemann, H. Epigenetic disorders and altered gene expression after use of Assisted Reproductive Technologies in domestic cattle. Epigenetics 2014, 9, 803–815. [Google Scholar] [CrossRef]
- Chao, S.B.; Li, J.C.; Jin, X.J.; Tang, H.X.; Wang, G.X.; Gao, G.L. Epigenetic reprogramming of embryos derived from sperm frozen at-20A degrees C. Sci. China-Life Sci. 2012, 55, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Salehi, M.; Mahdavi, A.H.; Sharafi, M.; Shahverdi, A. Cryopreservation of rooster semen: Evidence for the epigenetic modifications of thawed sperm. Theriogenology 2020, 142, 15–25. [Google Scholar] [CrossRef]
- Tunc, O.; Tremellen, K. Oxidative DNA damage impairs global sperm DNA methylation in infertile men. J. Assist. Reprod. Genet. 2009, 26, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Flores, E.; Ramio-Lluch, L.; Bucci, D.; Fernandez-Novell, J.M.; Pena, A.; Rodriguez-Gil, J.E. Freezing-thawing induces alterations in histone H1-DNA binding and the breaking of protein-DNA disulfide bonds in boar sperm. Theriogenology 2011, 76, 1450–1464. [Google Scholar] [CrossRef] [PubMed]
- Oganesyan, N.; Ankoudinova, I.; Kim, S.H.; Kim, R. Effect of osmotic stress and heat shock in recombinant protein overexpression and crystallization. Protein Expr. Purif. 2007, 52, 280–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aurich, C.; Schreiner, B.; Ille, N.; Alvarenga, M.; Scarlet, D. Cytosine methylation of sperm DNA in horse semen after cryopreservation. Theriogenology 2016, 86, 1347–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riesco, M.F.; Robles, V. Cryopreservation Causes Genetic and Epigenetic Changes in Zebrafish Genital Ridges. PLoS ONE 2013, 8, 9. [Google Scholar] [CrossRef]
- Vassena, R.; Boue, S.; Gonzalez-Roca, E.; Aran, B.; Auer, H.; Veiga, A.; Belmonte, J.C.I. Waves of early transcriptional activation and pluripotency program initiation during human preimplantation development. Development 2011, 138, 3699–3709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, G.; Maxwell, W.C. Salamons’ Artificial Insemination of Sheep and Goats; Butterworths: London, UK, 1987. [Google Scholar]
- Lovelock, J.E. The protective action of neutral solutes against haemolysis by freezing and thawing. Biochem. J. 1954, 56, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, M.A.; Papa, F.O.; Landim-Alvarenga, F.C.; Medeiros, A.S.L. Amides as cryoprotectants for freezing stallion semen: A review. Anim. Reprod. Sci. 2005, 89, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Hammerstedt, R.H.; Graham, J.K. Cryopreservation of poultry sperm—The enigma of glycerol. Cryobiology 1992, 29, 26–38. [Google Scholar] [CrossRef]
- Si, W.; Zheng, P.; Li, Y.H.; Dinnyes, A.; Ji, W.Z. Effect of glycerol and dimethyl sulfoxide on cryopreservation of rhesus monkey (Macaca mulatta) sperm. Am. J. Primatol. 2004, 62, 301–306. [Google Scholar] [CrossRef]
- Bucak, M.N.; Keskin, N.; Ili, P.; Bodu, M.; Akalin, P.P.; Ozturk, A.E.; Ozkan, H.; Topraggaleh, T.R.; Sari, F.; Baspinar, N.; et al. Decreasing glycerol content by co-supplementation of trehalose and taxifolin hydrate in ram semen extender: Microscopic, oxidative stress, and gene expression analyses. Cryobiology 2020, 96, 19–29. [Google Scholar] [CrossRef]
- Aisen, E.G.; Alvarez, H.L.; Venturino, A.; Garde, J.J. Effect of trehalose and EDTA on cryoprotective action of ram semen diluents. Theriogenology 2000, 53, 1053–1061. [Google Scholar] [CrossRef]
- Moussa, M.; Martinet, V.; Trimeche, A.; Tainturier, D.; Anton, M. Low density lipoproteins extracted from hen egg yolk by an easy method: Cryoprotective effect on frozen-thawed bull semen. Theriogenology 2002, 57, 1695–1706. [Google Scholar] [CrossRef]
- Emamverdi, M.; Zhandi, M.; Shahneh, A.Z.; Sharafi, M.; Akbari-Sharif, A. Optimization of Ram Semen Cryopreservation Using a Chemically Defined Soybean Lecithin-Based Extender. Reprod. Domest. Anim. 2013, 48, 899–904. [Google Scholar] [CrossRef]
- Forouzanfar, M.; Sharafi, M.; Hosseini, S.M.; Ostadhosseini, S.; Hajian, M.; Hosseini, L.; Abedi, P.; Nili, N.; Rahmani, H.R.; Nasr-Esfahani, M.H. In vitro comparison of egg yolk-based and soybean lecithin-based extenders for cryopreservation of ram semen. Theriogenology 2010, 73, 480–487. [Google Scholar] [CrossRef]
- Salmani, H.; Nabi, M.M.; Vaseghi-Dodaran, H.; Rahman, M.B.; Mohammadi-Sangcheshmeh, A.; Shakeri, M.; Towhidi, A.; Shahneh, A.Z.; Zhandi, M. Effect of glutathione in soybean lecithin-based semen extender on goat semen quality after freeze-thawing. Small Rumin. Res. 2013, 112, 123–127. [Google Scholar] [CrossRef]
- Aires, V.A.; Hinsch, K.D.; Mueller-Schloesser, F.; Bogner, K.; Mueller-Schloesser, S.; Hinsch, E. In vitro and in vivo comparison of egg yolk-based and soybean lecithin-based extenders for cryopreservation of bovine semen. Theriogenology 2003, 60, 269–279. [Google Scholar] [CrossRef]
- Akhter, S.; Ansari, M.S.; Andrabi, S.M.H.; Rakha, B.A.; Ullah, N.; Khalid, M. Soya-lecithin in Extender Improves the Freezability and Fertility of Buffalo (Bubalus bubalis) Bull Spermatozoa. Reprod. Domest. Anim. 2012, 47, 815–819. [Google Scholar] [CrossRef]
- Reed, M.L.; Ezeh, P.C.; Hamic, A.; Thompson, D.J.; Caperton, C.L. Soy lecithin replaces egg yolk for cryopreservation of human sperm without adversely affecting postthaw motility, morphology, sperm DNA integrity, or sperm binding to hyaluronate. Fertil. Steril. 2009, 92, 1787–1790. [Google Scholar] [CrossRef]
- Amidi, F.; Pazhohan, A.; Nashtaei, M.S.; Khodarahmian, M.; Nekoonam, S. The role of antioxidants in sperm freezing: A review. Cell Tissue Bank. 2016, 17, 745–756. [Google Scholar] [CrossRef]
- Aitken, R.J. Free radicals, lipid peroxidation and sperm function. Reprod. Fertil. Dev. 1995, 7, 659–668. [Google Scholar] [CrossRef]
- Maxwell, W.M.C.; Stojanov, T. Liquid storage of ram semen in the absence or presence of some antioxidants. Reprod. Fertil. Dev. 1996, 8, 1013–1020. [Google Scholar] [CrossRef] [PubMed]
- Forouzanfar, M.; Ershad, S.F.; Hosseini, S.M.; Hajian, M.; Ostad-Hosseini, S.; Abid, A.; Tavalaee, M.; Shahverdi, A.; Dizaji, A.V.; Esfahani, M.H.N. Can permeable super oxide dismutase mimetic agents improve the quality of frozen-thawed ram semen? Cryobiology 2013, 66, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.V.; Soares, A.T.; Batista, A.M.; Almeida, F.C.; Nunes, J.F.; Peixoto, C.A.; Guerra, M.M.P. In Vitro and In Vivo Evaluation of Ram Sperm Frozen in Tris Egg-yolk and Supplemented with Superoxide Dismutase and Reduced Glutathione. Reprod. Domest. Anim. 2011, 46, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Roca, J.; Rodriguez, M.J.; Gil, M.A.; Carvajal, G.; Garcia, E.M.; Cuello, C.; Vazquez, J.M.; Martinez, E.A. Survival and in vitro fertility of boar spermatozoa frozen in the presence of superoxide dismutase and/or catalase. J. Androl. 2005, 26, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Santos, M.R.; Dominguez-Rebolledo, A.E.; Esteso, M.C.; Garde, J.J.; Martinez-Pastor, F. Catalase supplementation on thawed bull spermatozoa abolishes the detrimental effect of oxidative stress on motility and DNA integrity. Int. J. Androl. 2009, 32, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Uysal, O.; Bucak, M.N. Effects of oxidized glutathione, bovine serum albumin, cysteine and lycopene on the quality of frozen-thawed ram semen. Acta Vet. BRNO 2007, 76, 383–390. [Google Scholar] [CrossRef]
- Izanloo, H.; Soleimanzadeh, A.; Bucak, M.N.; Imani, M.; Zhandi, M. The effects of glutathione supplementation on post-thawed Turkey semen quality and oxidative stress parameters and fertilization, and hatching potential. Theriogenology 2022, 179, 32–38. [Google Scholar] [CrossRef]
- Pezo, F.; Zambrano, F.; Uribe, P.; Moya, C.; de Andrade, A.F.C.; Risopatron, J.; Yeste, M.; Burgos, R.A.; Sanchez, R. Oxidative and nitrosative stress in frozen-thawed pig spermatozoa. I: Protective effect of melatonin and butylhydroxytoluene on sperm function. Res. Vet. Sci. 2021, 136, 143–150. [Google Scholar] [CrossRef]
- Pool, K.R.; Rickard, J.P.; de Graaf, S.P. Melatonin improves the motility and DNA integrity of frozen-thawed ram spermatozoa likely via suppression of mitochondrial superoxide production. Domest. Anim. Endocrinol. 2021, 74, 8. [Google Scholar] [CrossRef]
- Tanhaei Vash, N.; Nadri, P.; Karimi, A. Synergistic effects of myo-inositol and melatonin on cryopreservation of goat spermatozoa. Reprod. Domest. Anim. 2022, 57, 876–885. [Google Scholar] [CrossRef]
- Divar, M.R.; Azari, M.; Mogheiseh, A.; Ghahramani, S. Supplementation of melatonin to cooling and freezing extenders improves canine spermatozoa quality measures. BMC Vet. Res. 2022, 18, 16. [Google Scholar] [CrossRef]
- Felix, F.; Oliveira, C.C.V.; Cabrita, E. Antioxidants in Fish Sperm and the Potential Role of Melatonin. Antioxidants 2021, 10, 36. [Google Scholar] [CrossRef]
- Su, G.H.; Wu, S.S.; Wu, M.L.; Wang, L.N.; Yang, L.; Du, M.X.; Zhao, X.Y.; Su, X.H.; Liu, X.F.; Bai, C.L.; et al. Melatonin improves the quality of frozen bull semen and influences gene expression related to embryo genome activation. Theriogenology 2021, 176, 54–62. [Google Scholar] [CrossRef]
- Minucci, S.; Venditti, M. New insight on the in vitro effects of melatonin in preserving human sperm quality. Int. J. Mol. Sci. 2022, 23, 5128. [Google Scholar] [CrossRef]
- Hull, M.G.; North, K.; Taylor, H.; Farrow, A.; Ford, W.C.L. Delayed conception and active and passive smoking. Fertil. Steril. 2000, 74, 725–733. [Google Scholar] [CrossRef]
- Cerolini, S.; Maldjian, A.; Surai, P.; Noble, R. Viability, susceptibility to peroxidation and fatty acid composition of boar semen during liquid storage. Anim. Reprod. Sci. 2000, 58, 99–111. [Google Scholar] [CrossRef]
- Dalvit, G.C.; Cetica, P.D.; Beconi, M.T. Effect of alpha-tocopherol and ascorbic acid on bovine in vitro fertilization. Theriogenology 1998, 49, 619–627. [Google Scholar] [CrossRef]
- Khan, R.U.; Rahman, Z.U.; Javed, I.; Muhammad, F. Effect of vitamins, probiotics and protein on semen traits in post-molt male broiler breeders. Anim. Reprod. Sci. 2012, 135, 85–90. [Google Scholar] [CrossRef]
- Brzezinskaslebodzinska, E.; Slebodzinski, A.B.; Pietras, B.; Wieczorek, G. Antioxidant Effect of Vitamin-E and Glutathione on Lipid-Peroxidation In Boar Semen Plasma. Biol. Trace Elem. Res. 1995, 47, 69–74. [Google Scholar] [CrossRef]
- Yousef, M.I. Vitamin E modulates reproductive toxicity of pyrethroid lambda-cyhalothrin in male rabbits. Food Chem. Toxicol. 2010, 48, 1152–1159. [Google Scholar] [CrossRef]
- Masoudi, R.; Sharafi, M.; Shahneh, A.Z.; Towhidi, A.; Kohram, H.; Esmaeili, V.; Shahverdi, A.; Davachi, N.D. Fertility and flow cytometry study of frozen-thawed sperm in cryopreservation medium supplemented with soybean lecithin. Cryobiology 2016, 73, 69–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majid, A.; Qureshi, M.S.; Khan, R.U. In vivo adverse effects of alpha-tocopherol on the semen quality of male bucks. J. Anim. Physiol. Anim. Nutr. 2015, 99, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Sonmez, M.; Turk, G.; Yuce, A. The effect of ascorbic acid supplementation on sperm quality, lipid peroxidation and testosterone levels of male Wistar rats. Theriogenology 2005, 63, 2063–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraga, C.G.; Motchnik, P.A.; Shigenaga, M.K.; Helbock, H.J.; Jacob, R.A.; Ames, B.N. Ascorbic-Acid Protects against Endogenous Oxidative DNA Damage in Human Sperm. Proc. Natl. Acad. Sci. USA 1991, 88, 11003–11006. [Google Scholar] [CrossRef] [Green Version]
- Azawi, O.I.; Hussein, E.K. Effect of vitamins C or E supplementation to Tris diluent on the semen quality of Awassi rams preserved at 5 C. In Veterinary Research Forum; Faculty of Veterinary Medicine, Urmia University: Urmia, Iran, 2013; p. 157. [Google Scholar]
- Sanchez-Partida, L.G.; Setchell, B.P.; Maxwell, W.M.C. Epididymal compounds and antioxidants in diluents for the frozen storage of ram spermatozoa. Reprod. Fertil. Dev. 1997, 9, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Rietjens, I.; Boersma, M.G.; de Haan, L.; Spenkelink, B.; Awad, H.M.; Cnubben, N.H.P.; van Zanden, J.J.; van der Woude, H.; Alink, G.M.; Koeman, J.H. The pro-oxidant chemistry of the natural antioxidants vitamin C, vitamin E, carotenoids and flavonoids. Environ. Toxicol. Pharmacol. 2002, 11, 321–333. [Google Scholar] [CrossRef]
- Bucak, M.N.; Tuncer, P.B.; Sariozkan, S.; Ulutas, P.A. Comparison of the effects of glutamine and an amino acid solution on post-thawed ram sperm parameters, lipid peroxidation and anti-oxidant activities. Small Rumin. Res. 2009, 81, 13–17. [Google Scholar] [CrossRef]
- Sangeeta, S.; Kulkarni, S.; Arangasamy, A.; Selvaraju, S. Effect of amino acids on sperm motility, velocity parameters, plasma membrane integrity and lipid peroxidation levels at cooled and post-thawed ram epididymal semen. Indian J. Anim. Sci. 2015, 85, 729–735. [Google Scholar]
- Fattah, A.; Sharafi, M.; Masoudi, R.; Shahverdi, A.; Esmaeili, V. L-carnitine is a survival factor for chilled storage of rooster semen for a long time. Cryobiology 2017, 74, 13–18. [Google Scholar] [CrossRef]
- Zhandi, M.; Sharafi, M. Negative effect of combined cysteine and glutathione in soy lecithin-based extender on post-thawed ram spermatozoa. Cell Tissue Bank. 2015, 16, 443–448. [Google Scholar] [CrossRef]
- Lewis, S.E.M.; Sterling, E.S.L.; Young, I.S.; Thompson, W. Comparison of individual antioxidants of sperm and seminal plasma in fertile and infertile men. Fertil. Steril. 1997, 67, 142–147. [Google Scholar] [CrossRef]
- Matsuoka, T.; Imai, H.; Kohno, H.; Fukui, Y. Effects of bovine serum albumin and trehalose in semen diluents for improvement of frozen-thawed ram spermatozoa. J. Reprod. Dev. 2006, 52, 675–683. [Google Scholar] [CrossRef] [Green Version]
- Fukui, Y.; Kohno, H.; Togari, T.; Hiwasa, M. Fertility of ewes inseminated intrauterinally with frozen semen using extender containing bovine serum albumin. J. Reprod. Dev. 2007, 53, 959–962. [Google Scholar] [CrossRef] [Green Version]
- Coyan, K.; Bucak, M.N.; Baspinar, N.; Taspinar, M.; Aydos, S. Ergothioneine attenuates the DNA damage of post-thawed Merino ram sperm. Small Rumin. Res. 2012, 106, 165–167. [Google Scholar] [CrossRef]
- Usuga, A.; Tejera, I.; Gomez, J.; Restrepo, O.; Rojano, B.; Restrepo, G. Cryoprotective Effects of Ergothioneine and Isoespintanol on Canine Semen. Animals 2021, 11, 2757. [Google Scholar] [CrossRef]
- Crowe, L.M.; Crowe, J.H.; Int Assoc Biol, S. Stabilization of dry liposomes by carbohydrates. In Developments in Biological Standardization; Warren Grant Magnuson Clinical Center of the National Institutes: Bethesda, MD, USA, 1992; pp. 285–294. [Google Scholar]
- Aboagla, E.M.E.; Terada, T. Trehalose-enhanced fluidity of the goat sperm membrane and its protection during freezing. Biol. Reprod. 2003, 69, 1245–1250. [Google Scholar] [CrossRef]
- Patist, A.; Zoerb, H. Preservation mechanisms of trehalose in food and biosystems. Colloid Surf. B Biointerfaces 2005, 40, 107–113. [Google Scholar] [CrossRef]
- Ahmad, E.; Naseer, Z.; Aksoy, M.; Kucuk, N.; Ucan, U.; Serin, I.; Ceylan, A. Trehalose enhances osmotic tolerance and suppresses lysophosphatidylcholine-induced acrosome reaction in ram spermatozoon. Andrologia 2015, 47, 786–792. [Google Scholar] [CrossRef]
- Jia, B.Y.; Memon, S.; Liang, J.C.; Lv, C.R.; Hong, Q.H.; Wu, G.Q.; Quan, G.B. Trehalose modifies the protein profile of ram spermatozoa during cryopreservation. Theriogenology 2021, 171, 21–29. [Google Scholar] [CrossRef]
- Bucak, M.N.; Keskin, N.; Bodu, M.; Bulbul, B.; Kirbas, M.; Ozturk, A.E.; Frootan, F.; Ili, P.; Ozkan, H.; Baspinar, N.; et al. Combination of trehalose and low boron in presence of decreased glycerol improves post-thawed ram sperm parameters: A model study in boron research. Andrology 2022, 10, 585–594. [Google Scholar] [CrossRef]
- Tada, N.; Sato, M.; Yamanoi, J.; Mizorogi, T.; Kasai, K. Cryopreservation of mouse spermatozoa in the presence of raffinose and glycerol. J. Reprod. Fertil. 1990, 89, 511–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thananurak, P.; Chuaychu-noo, N.; Thelie, A.; Phasuk, Y.; Vongpralub, T.; Blesbois, E. Sucrose increases the quality and fertilizing ability of cryopreserved chicken sperms in contrast to raffinose. Poult. Sci. 2019, 98, 4161–4171. [Google Scholar] [CrossRef] [PubMed]
- De Mercado, E.; Rodríguez, A.; Gómez, E.; Sanz, E. Cryopreservation of Iberian pig spermatozoa. Comparison of different freezing extenders based on post-thaw sperm quality. Anim. Reprod. Sci. 2010, 118, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Pezo, F.; Zambrano, F.; Uribe, P.; Risopatrón, J.; Moya, C.; de Andrade, A.F.C.; Burgos, R.A.; Yeste, M.; Sánchez, R. Oxidative and nitrosative stress in frozen-thawed pig spermatozoa. II: Effect of the addition of saccharides to freezing medium on sperm function. Cryobiology 2020, 97, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.; Cunha, E.; Blume, G.; Malaquias, J.; Báo, S.; Martins, C. Cryopreservation of boar sperm comparing different cryoprotectants associated in media based on powdered coconut water, lactose and trehalose. Cryobiology 2015, 70, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Chanapiwat, P.; Kaeoket, K.; Tummaruk, P. Cryopreservation of boar semen by egg yolk-based extenders containing lactose or fructose is better than sorbitol. J. Vet. Med. Sci. 2012, 74, 351–354. [Google Scholar] [CrossRef] [Green Version]
- Molinia, F.; Evans, G.; Casares, P.Q.; Maxwell, W. Effect of monosaccharides and disaccharides in Tris-based diluents on motility, acrosome integrity and fertility of pellet frozen ram spermatozoa. Anim. Reprod. Sci. 1994, 36, 113–122. [Google Scholar] [CrossRef]
- Bucak, M.N.; Atessahin, A.; Varish, O.; Yuce, A.; Tekin, N.; Akcay, A. The influence of trehalose, taurine, cysteamine and hyaluronan on ram semen—Microscopic and oxidative stress parameters after freeze-thawing process. Theriogenology 2007, 67, 1060–1067. [Google Scholar] [CrossRef]
- Jafaroghli, M.; Khalili, B.; Farshad, A.; Zamiri, M.J. The effect of supplementation of cryopreservation diluents with sugars on the post-thawing fertility of ram semen. Small Rumin. Res. 2011, 96, 58–63. [Google Scholar] [CrossRef]
- Aisen, E.G.; Medina, V.H.; Venturino, A. Cryopreservation and post-thawed fertility of ram semen frozen in different trehalose concentrations. Theriogenology 2002, 57, 1801–1808. [Google Scholar] [CrossRef]
- Najafi, A.; Zhandi, M.; Towhidi, A.; Sharafi, M.; Sharif, A.A.; Motlagh, M.K.; Martinez-Pastor, F. Trehalose and glycerol have a dose-dependent synergistic effect on the post-thawing quality of ram semen cryopreserved in a soybean lecithin-based extender. Cryobiology 2013, 66, 275–282. [Google Scholar] [CrossRef]
- Uysal, O.; Bucak, M.N. The role of different trehalose concentrations and cooling rates in freezing of ram semen. Ank. Üniv. Vet. Fak. Derg. 2009, 56, 99–103. [Google Scholar]
- Ozturk, A.E.; Bodu, M.; Bucak, M.N.; Agir, V.; Ozcan, A.; Keskin, N.; Ili, P.; Topraggaleh, T.R.; Sidal, H.; Baspinar, N.; et al. The synergistic effect of trehalose and low concentrations of cryoprotectants can improve post-thaw ram sperm parameters. Cryobiology 2020, 95, 157–163. [Google Scholar] [CrossRef]
- Bucak, M.N.; Tekin, N. Protective effect of taurine, glutathione and trehalose on the liquid storage of ram semen. Small Rumin. Res. 2007, 73, 103–108. [Google Scholar] [CrossRef]
- Keskin, N.; Erdogan, C.; Bucak, M.N.; Ozturk, A.E.; Bodu, M.; Ili, P.; Baspinar, N.; Dursun, S. Cryopreservation Effects on Ram Sperm Ultrastructure. Biopreserv. Biobank. 2020, 18, 441–448. [Google Scholar] [CrossRef]
- Reddy, N.S.S.; Mohanarao, G.J.; Atreja, S.K. Effects of adding taurine and trehalose to a tris-based egg yolk extender on buffalo (Bubalus bubalis) sperm quality following cryopreservation. Anim. Reprod. Sci. 2010, 119, 183–190. [Google Scholar] [CrossRef]
- Ozturk, C.; Gungor, S.; Ataman, M.B.; Bucak, M.N.; Baspinar, N.; Ili, P.; Inanc, M.E. Effects of Arginine and Trehalose on Post-Thawed Bovine Sperm Quality. Acta Vet. Hung. 2017, 65, 429–439. [Google Scholar] [CrossRef] [Green Version]
- Akhter, S.; Ansari, M.S.; Rakha, B.A.; Andrabi, S.M.H.; Anwar, M.; Ullah, N. Effect of Fructose Addition in Skim Milk Extender on the Quality of Liquid Nili-Ravi Buffalo (Bubalus bubalis) Semen. Pak. J. Zool. 2010, 42, 227–231. [Google Scholar]
- Salamon, S.; Visser, D. Effect of composition of Tris-based diluent and of thawing solution on survival of ram spermatozoa frozen by the pellet method. Aust. J. Biol. Sci. 1972, 25, 605–618. [Google Scholar] [CrossRef] [Green Version]
- De los Reyes, M.; Saenz, L.; Lapierre, L.; Crosby, J.; Barros, C. Evaluation of glucose as a cryoprotectant for boar semen. Vet. Rec. 2002, 151, 477–480. [Google Scholar] [CrossRef]
- Bucak, M.N.; Keskin, N.; Taspinar, M.; Coyan, K.; Baspinar, N.; Cenariu, M.C.; Bilgili, A.; Ozturk, C.; Kursunlu, A.N. Raffinose and hypotaurine improve the post-thawed Merino ram sperm parameters. Cryobiology 2013, 67, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Sztein, J.M.; Noble, K.; Farley, J.S.; Mobraaten, L.E. Comparison of permeating and nonpermeating cryoprotectants for mouse sperm cryopreservation. Cryobiology 2001, 42, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Buyukleblebici, S.; Tuncer, P.B.; Bucak, M.N.; Tasdemir, U.; Eken, A.; Buyukleblebici, O.; Durmaz, E.; Sanozkan, S.; Endirlik, B.U. Comparing ethylene glycol with glycerol and with or without dithiothreitol and sucrose for cryopreservation of bull semen in egg-yolk containing extenders. Cryobiology 2014, 69, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Consuegra, C.; Crespo, F.; Bottrel, M.; Ortiz, I.; Dorado, J.; Diaz-Jimenez, M.; Pereira, B.; Hidalgo, M. Stallion sperm freezing with sucrose extenders: A strategy to avoid permeable cryoprotectants. Anim. Reprod. Sci. 2018, 191, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Salmon, V.M.; Castonguay, F.; Demers-Caron, V.; Leclerc, P.; Bailey, J.L. Cholesterol-loaded cyclodextrin improves ram sperm cryoresistance in skim milk-extender. Anim. Reprod. Sci. 2017, 177, 1–11. [Google Scholar] [CrossRef]
- Yang, S.X.; Adams, G.P.; Zwiefelhofer, E.M.; Rajapaksha, K.; Anzar, M. Cholesterol-cyclodextrin complex as a replacement for egg yolk in bull semen extender: Sperm characteristics post-thawing and in vivo fertility. Anim. Reprod. Sci. 2021, 225, 106691. [Google Scholar] [CrossRef]
- Glazar, A.I.; Mullen, S.F.; Liu, J.; Benson, J.D.; Critser, J.K.; Squires, E.L.; Graham, J.K. Osmotic tolerance limits and membrane permeability characteristics of stallion spermatozoa treated with cholesterol. Cryobiology 2009, 59, 201–206. [Google Scholar] [CrossRef]
- Lone, S.A. Possible mechanisms of cholesterol-loaded cyclodextrin action on, sperm during cryopreservation. Anim. Reprod. Sci. 2018, 192, 1–5. [Google Scholar] [CrossRef]
- Moce, E.; Blanch, E.; Tomas, C.; Graham, J.K. Use of Cholesterol in Sperm Cryopreservation: Present Moment and Perspectives to Future. Reprod. Domest. Anim. 2010, 45, 57–66. [Google Scholar] [CrossRef]
- Baishya, S.K.; Biswas, R.K.; Govindasamy, K.; Deka, B.C.; Sinha, S.; Singh, M. Pre-freezing treatment with butylated hydoxytoluene and cholesterol-loaded methyl-beta-cyclodextrin improves quality of cryopreserved boar semen. Cryoletters 2018, 39, 336–344. [Google Scholar]
- Yadav, H.P.; Kumar, A.; Shah, N.; Chauhan, D.S.; Lone, S.A.; Swain, D.K.; Saxena, A. Effect of cholesterol-loaded cyclodextrin on membrane and acrosome status of hariana bull sperm during cryopreservation. Cryoletters 2018, 39, 386–390. [Google Scholar]
- Khellouf, A.; Benhenia, K.; Fatami, S.; Iguer-Ouada, M. The complementary effect of cholesterol and vitamin E preloaded in cyclodextrins on frozen bovine semen: Motility parameters, membrane integrity and lipid peroxidation. Cryo Lett. 2018, 39, 113–120. [Google Scholar]
- Chuaychu-noo, N.; Thananurak, P.; Chankitisakul, V.; Vongpralub, T. Supplementing rooster sperm with Cholesterol-Loaded-Cyclodextrin improves fertility after cryopreservation. Cryobiology 2017, 74, 8–12. [Google Scholar] [CrossRef]
- Murphy, C.; English, A.M.; Holden, S.A.; Fair, S. Cholesterol-loaded-cyclodextrins improve the post-thaw quality of stallion sperm. Anim. Reprod. Sci. 2014, 145, 123–129. [Google Scholar] [CrossRef]
- Purdy, P.H.; Moce, E.; Stobart, R.; Murdoch, W.J.; Moss, G.E.; Larson, B.; Ramsey, S.; Graham, J.K.; Blackburn, H.D. The fertility of ram sperm held for 24 h at 5 degrees C prior to cryopreservation. Anim. Reprod. Sci. 2010, 118, 231–235. [Google Scholar] [CrossRef]
- Spizziri, B.E.; Fox, M.H.; Bruemmer, J.E.; Squires, E.L.; Graham, J.K. Cholesterol-loaded-cyclodextrins and fertility potential of stallions spermatozoa. Anim. Reprod. Sci. 2010, 118, 255–264. [Google Scholar] [CrossRef]
- Oliveira, R.R.; Rates, D.M.; Pugliesi, G.; Ker, P.G.; Arruda, R.P.; Moraes, E.A.; Carvalho, G.R. Use of Cholesterol-Loaded Cyclodextrin in Donkey Semen Cryopreservation Improves Sperm Viability but Results in Low Fertility in Mares. Reprod. Domest. Anim. 2014, 49, 845–850. [Google Scholar] [CrossRef]
- Zanussi, H.P.; Shariatmadari, F.; Sharafi, M.; Ahmadi, H. Dietary supplementation with flaxseed oil as source of Omega-3 fatty acids improves seminal quality and reproductive performance in aged broiler breeder roosters. Theriogenology 2019, 130, 41–48. [Google Scholar] [CrossRef]
- Brinsko, S.P.; Varner, D.D.; Love, C.C.; Blanchard, T.L.; Day, B.C.; Wilson, M.E. Effect of feeding a DHA-enriched nutriceutical on the quality of fresh, cooled and frozen stallion semen. Theriogenology 2005, 63, 1519–1527. [Google Scholar] [CrossRef]
- Ansari, M.; Zhandi, M.; Kohram, H.; Zaghari, M.; Sadeghi, M.; Sharafi, M. Improvement of post-thawed sperm quality and fertility of Arian rooster by oral administration of D-aspartic acid. Theriogenology 2017, 92, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Díaz, R.; Torres, M.A.; Paz, E.; Quiñones, J.; Bravo, S.; Farías, J.G.; Sepúlveda, N. Dietary inclusion of fish oil changes the semen lipid composition but does not improve the post-thaw semen quality of ram spermatozoa. Anim. Reprod. Sci. 2017, 183, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, V.; Shahverdi, A.H.; Alizadeh, A.R.; Alipour, H.; Chehrazi, M. Saturated, omega-6 and omega-3 dietary fatty acid effects on the characteristics of fresh, frozen-thawed semen and blood parameters in rams. Andrologia 2014, 46, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Masoudi, R.; Sharafi, M.; Shahneh, A.Z.; Towhidi, A.; Kohram, H.; Zhandi, M.; Esmaeili, V.; Shahverdi, A. Effect of dietary fish oil supplementation on ram semen freeze ability and fertility using soybean lecithin- and egg yolk-based extenders. Theriogenology 2016, 86, 1583–1588. [Google Scholar] [CrossRef] [PubMed]
- Hashem, E.Z.; Haddad, R.; Eslami, M. Evaluation of ram semen enrichment with oleic acid on different spermatozoa parameters during low temperature liquid storage. Small Rumin. Res. 2017, 150, 30–39. [Google Scholar] [CrossRef]
- Henry, M.A.; Noiles, E.E.; Gao, D.; Mazur, P.; Critser, J.K. Cryopreservation of human spermatozoa. IV. The effects of cooling rate and warming rate on the maintenance of motility, plasma membrane integrity, and mitochondrial function. Fertil. Steril. 1993, 60, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Seki, S.; Jin, B.; Mazur, P. Extreme rapid warming yields high functional survivals of vitrified 8-cell mouse embryos even when suspended in a half-strength vitrification solution and cooled at moderate rates to −196 °C. Cryobiology 2014, 68, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Almenar, C.; De Mercado, E. Optimization of the Thawing Protocol for Iberian Boar Sperm. Animals 2022, 12, 2600. [Google Scholar] [CrossRef]
- Hernandez, M.; Roca, J.; Gil, M.A.; Vazquez, J.M.; Martinez, E.A. Adjustments on the cryopreservation conditions reduce the incidence of boar ejaculates with poor sperm freezability. Theriogenology 2007, 67, 1436–1445. [Google Scholar] [CrossRef]
- Lyashenko, A. Effect of different thawing procedures on the quality and fertility of the bull spermatozoa. Asian Pac. J. Reprod. 2015, 4, 17–21. [Google Scholar] [CrossRef]
- Correa, J.; Rodriguez, M.; Patterson, D.; Zavos, P. Thawing and processing of cryopreserved bovine spermatozoa at various temperatures and their effects on sperm viability, osmotic shock and sperm membrane functional integrity. Theriogenology 1996, 46, 413–420. [Google Scholar] [CrossRef]
- Huang, S.Y.; Pribenszky, C.; Kuo, Y.H.; Teng, S.H.; Chen, Y.H.; Chung, M.T.; Chiu, Y.F. Hydrostatic pressure pre-treatment affects the protein profile of boar sperm before and after freezing-thawing. Anim. Reprod. Sci. 2009, 112, 136–149. [Google Scholar] [CrossRef]
- Lin, L.; Kragh, P.M.; Purup, S.; Kuwayama, M.; Du, Y.T.; Zhang, X.Q.; Yang, H.M.; Bolund, L.; Callesen, H.; Vajta, G. Osmotic stress induced by sodium chloride, sucrose or trehalose improves cryotolerance and developmental competence of porcine oocytes. Reprod. Fertil. Dev. 2009, 21, 338–344. [Google Scholar] [CrossRef]
- Sharafi, M.; Zhandi, M.; Shahverdi, A.; Shakeri, M. Beneficial effects of nitric oxide induced mild oxidative stress on post-thawed bull semen quality. Int. J. Fertil. Steril. 2015, 9, 230. [Google Scholar] [CrossRef]
- Csermely, P.; Schnaider, T.; Soti, C.; Prohaszka, Z.; Nardai, G. The 90-kDa molecular chaperone family: Structure, function, and clinical applications. A comprehensive review. Pharmacol. Ther. 1998, 79, 129–168. [Google Scholar] [CrossRef]
- Hezavehei, M.; Kouchesfahani, H.M.; Shahverdi, A.; Sharafi, M.; Salekdeh, G.H.; Eftekhari-Yazdi, P. Induction of Sublethal Oxidative Stress on Human Sperm before Cryopreservation: A Time-Dependent Response in Post-Thawed Sperm Parameters. Cell J. 2019, 20, 537–543. [Google Scholar] [CrossRef]
- Safian, F.; Novin, M.G.; Nazarian, H.; Mofarahe, Z.S.; Abdollahifar, M.A.; Jajarmi, V.; Karimi, S.; Kazemi, M.; Chien, S.F.; Bayat, M. Photobiomodulation preconditioned human semen protects sperm cells against detrimental effects of cryopreservation. Cryobiology 2021, 98, 239–244. [Google Scholar] [CrossRef]
- Iaffaldano, N.; Meluzzi, A.; Manchisi, A.; Passarella, S. Improvement of stored turkey semen quality as a result of He-Ne laser irradiation. Anim. Reprod. Sci. 2005, 85, 317–325. [Google Scholar] [CrossRef]
- Fernandes, G.H.C.; de Carvalho, P.d.T.C.; Serra, A.J.; Crespilho, A.M.; Peron, J.P.S.; Rossato, C.; Leal-Junior, E.C.P.; Albertini, R. The Effect of Low-Level Laser Irradiation on Sperm Motility, and Integrity of the Plasma Membrane and Acrosome in Cryopreserved Bovine Sperm. PLoS ONE 2015, 10, e0121487. [Google Scholar] [CrossRef]
- Iaffaldano, N.; Rosato, M.P.; Paventi, G.; Pizzuto, R.; Gambacorta, M.; Manchisi, A.; Passarella, S. The irradiation of rabbit sperm cells with He–Ne laser prevents their in vitro liquid storage dependent damage. Anim. Reprod. Sci. 2010, 119, 123–129. [Google Scholar] [CrossRef]
- Malkin, T.L.; Murray, B.J.; Brukhno, A.V.; Anwar, J.; Salzmann, C.G. Structure of ice crystallized from supercooled water. Proc. Natl. Acad. Sci. USA 2012, 109, 1041–1045. [Google Scholar] [CrossRef] [Green Version]
- Huebinger, J.; Han, H.M.; Hofnagel, O.; Vetter, I.R.; Bastiaens, P.I.H.; Grabenbauer, M. Direct Measurement of Water States in Cryopreserved Cells Reveals Tolerance toward Ice Crystallization. Biophys. J. 2016, 110, 840–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askarianzadeh, Z.; Sharafi, M.; Torshizi, M.A.K. Sperm quality characteristics and fertilization capacity after cryopreservation of rooster semen in extender exposed to a magnetic field. Anim. Reprod. Sci. 2018, 198, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Gholami, D.; Ghaffari, S.M.; Riazi, G.; Fathi, R.; Benson, J.; Shahverdi, A.; Sharafi, M. Electromagnetic field in human sperm cryopreservation improves fertilizing potential of thawed sperm through physicochemical modification of water molecules in freezing medium. PLoS ONE 2019, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- Falchi, L.; Khalil, W.A.; Hassan, M.; Marei, W.F.A. Perspectives of nanotechnology in male fertility and sperm function. Int. J. Vet. Sci. Med. 2018, 6, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teli, M.K.; Mutalik, S.; Rajanikant, G.K. Nanotechnology and Nanomedicine: Going Small Means Aiming Big. Curr. Pharm. Des. 2010, 16, 1882–1892. [Google Scholar] [CrossRef] [PubMed]
- Feugang, J.M.; Rhoads, C.E.; Mustapha, P.A.; Tardif, S.; Parrish, J.J.; Willard, S.T.; Ryan, P.L. Treatment of boar sperm with nanoparticles for improved fertility. Theriogenology 2019, 137, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Nadri, T.; Towhidi, A.; Zeinoaldini, S.; Martinez-Pastor, F.; Mousavi, M.; Noei, R.; Tar, M.; Sangcheshmeh, A.M. Lecithin nanoparticles enhance the cryosurvival of caprine sperm. Theriogenology 2019, 133, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Khalil, W.A.; El-Harairy, M.A.; Zeidan, A.E.B.; Hassan, M.A.E. Impact of selenium nano-particles in semen extender on bull sperm quality after cryopreservation. Theriogenology 2019, 126, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Safa, S.; Moghaddam, G.; Jozani, R.J.; Kia, H.D.; Janmohammadi, H. Effect of vitamin E and selenium nanoparticles on post-thaw variables and oxidative status of rooster semen. Anim. Reprod. Sci. 2016, 174, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Mehdipour, M.; Kia, H.D.; Nazari, M.; Najafi, A. Effect of lecithin nanoliposome or soybean lecithin supplemented by pomegranate extract on post-thaw flow cytometric, microscopic and oxidative parameters in ram semen. Cryobiology 2017, 78, 34–40. [Google Scholar] [CrossRef]
- Anzalone, D.A.; Palazzese, L.; Iuso, D.; Martino, G.; Loi, P. Freeze-dried spermatozoa: An alternative biobanking option for endangered species. Anim. Reprod. Sci. 2018, 190, 85–93. [Google Scholar] [CrossRef]
- Kaneko, T.; Serikawa, T. Successful Long-Term Preservation of Rat Sperm by Freeze-Drying. PLoS ONE 2012, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Saragusty, J.; Loi, P. Exploring dry storage as an alternative biobanking strategy inspired by Nature. Theriogenology 2019, 126, 17–27. [Google Scholar] [CrossRef]
- Oldenhof, H.; Zhang, M.; Narten, K.; Bigalk, J.; Sydykov, B.; Wolkers, W.F.; Sieme, H. Freezing-induced uptake of disaccharides for preservation of chromatin in freeze-dried stallion sperm during accelerated aging. Biol. Reprod. 2017, 97, 892–901. [Google Scholar] [CrossRef] [Green Version]
- Wakayama, T.; Yanagimachi, R. Development of normal mice from oocytes injected with freeze-dried spermatozoa. Nat. Biotechnol. 1998, 16, 639–641. [Google Scholar] [CrossRef]
- Ward, M.A.; Kaneko, T.; Kusakabe, H.; Biggers, J.D.; Whittingham, D.G.; Yanagimachi, R. Long-term preservation of mouse spermatozoa after freeze-drying and freezing without cryoprotection. Biol. Reprod. 2003, 69, 2100–2108. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.L.; Kusakabe, H.; Chang, C.C.; Suzuki, H.; Schmidt, D.W.; Julian, M.; Pfeffer, R.; Bormann, C.L.; Tian, X.C.; Yanagimachi, R.; et al. Freeze-dried sperm fertilization leads to full-term development in rabbits. Biol. Reprod. 2004, 70, 1776–1781. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.H.; Varner, D.D.; Love, C.C.; Hartman, D.L.; Hinrichs, K. Production of live foals via intracytoplasmic injection of lyophilized sperm and sperm extract in the horse. Reproduction 2011, 142, 529–538. [Google Scholar] [CrossRef] [Green Version]
- Lima-Verde, I.; Hurri, E.; Ntallaris, T.; Johannisson, A.; Stålhammar, H.; Morrell, J.M. Sperm Quality in Young Bull Semen Can Be Improved by Single Layer Centrifugation. Animals 2022, 12, 2435. [Google Scholar] [CrossRef] [PubMed]
- Goodla, L.; Morrell, J.M.; Yusnizar, Y.; Stålhammar, H.; Johannisson, A. Quality of bull spermatozoa after preparation by single-layer centrifugation. J. Dairy Sci. 2014, 97, 2204–2212. [Google Scholar] [CrossRef] [Green Version]
- Morrell, J.; Johannisson, A.; Dalin, A.M.; Rodriguez-Martinez, H. Morphology and chromatin integrity of stallion spermatozoa prepared by density gradient and single layer centrifugation through silica colloids. Reprod. Domest. Anim. 2009, 44, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.U.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the Earth’s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Svoradova, A.; Kuzelova, L.; Vasicek, J.; Olexikova, L.; Chrenek, P. Cryopreservation of chicken blastodermal cells and their quality assessment by flow cytometry and transmission electron microscopy. Biotechnol. Prog. 2018, 34, 778–783. [Google Scholar] [CrossRef]
- Joost, S.; Bruford, M.W.; Genomic-Resources, C. Editorial: Advances in Farm Animal Genomic Resources. Front. Genet. 2015, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Canovas, S.; Campos, R.; Aguilar, E.; Cibelli, J.B. Progress towards human primordial germ cell specification in vitro. Mol. Hum. Reprod. 2017, 23, 4–15. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Year | Reference |
|---|---|---|
| Avian | 1942 | [14] |
| Bovine | 1951 | [11] |
| Human | 1953 | [15] |
| Porcine | 1957 | [16] |
| Equine | 1957 | [17] |
| Ovine | 1967 | [18] |
| Sugars | Species | Results | Authors |
|---|---|---|---|
| Trehalose | Ram | Increased motility of frozen–thawed sperm | [135] |
| Improved motility of frozen–thawed sperm | [136] | ||
| Improved viability and membrane integrity of frozen–thawed sperm | [137] | ||
| Improved post-thaw parameters | [126] | ||
| Improved acrosome integrity | [138] | ||
| Improved kinetic parameters, morphology, membrane integrity, and mitochondrial activity | [139] | ||
| Improved post-thaw recovery using 50 and 100 mM trehalose with slow cooling | [140] | ||
| Increased post-thaw parameters using combination of 3% glycerol and 60 mM trehalose | [141] | ||
| Improved survival rate during cold storage using combination of 50 mM taurine and 50 mM trehalose | [142] | ||
| Improved ultrastructural morphology of sperm using combination of 1.5% ethylene glycol and 100 mM trehalose | [143] | ||
| Goat | Increased post-thaw motility and acrosome integrity | [124] | |
| Buffalo | Improved motility, viability, and membrane integrity | [144] | |
| Bull | Improved post-thaw mitochondrial activity and viability | [145] | |
| Fructose | Boar | Fructose-based extender improved post-thaw motility and viability | [134] |
| Bull | Fructose-based extender improved motility, plasma membrane integrity on the 3rd, 5th, and 7th day of storage | [146] | |
| Glucose | Ram | Glucose improved post-thaw parameters | [147] |
| Boar | Glucose increased post-thaw recovery | [148] | |
| Raffinose | Ram | Improved motility, viability, mitochondrial activity of frozen–thawed sperm | [149] |
| Increased viability and motility and decreased acrosome abnormalities | [137] | ||
| Chicken | Reduced fertility performance | [130] | |
| Bull | Improved motility and plasma membrane integrity on 3rd, 5th and 7th day of storage | [146] | |
| Mouse | Raffinose is required for the standard inbred mouse sperm cryopreservation protocol | [150] | |
| Sucrose | Bull | Improved motility, acrosome integrity, and plasma membrane functionality | [151] |
| Stallion | Increased several kinetic parameters using 100 mM sucrose | [152] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharafi, M.; Borghei-Rad, S.M.; Hezavehei, M.; Shahverdi, A.; Benson, J.D. Cryopreservation of Semen in Domestic Animals: A Review of Current Challenges, Applications, and Prospective Strategies. Animals 2022, 12, 3271. https://doi.org/10.3390/ani12233271
Sharafi M, Borghei-Rad SM, Hezavehei M, Shahverdi A, Benson JD. Cryopreservation of Semen in Domestic Animals: A Review of Current Challenges, Applications, and Prospective Strategies. Animals. 2022; 12(23):3271. https://doi.org/10.3390/ani12233271
Chicago/Turabian StyleSharafi, Mohsen, Seyyed Mohsen Borghei-Rad, Maryam Hezavehei, Abdolhossein Shahverdi, and James D. Benson. 2022. "Cryopreservation of Semen in Domestic Animals: A Review of Current Challenges, Applications, and Prospective Strategies" Animals 12, no. 23: 3271. https://doi.org/10.3390/ani12233271