

Neurophysiology of Milk Ejection and Prestimulation in Dairy Buffaloes

 ,
,  ,
,  ,
,  , ,
, ,  , , , ,
, , , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
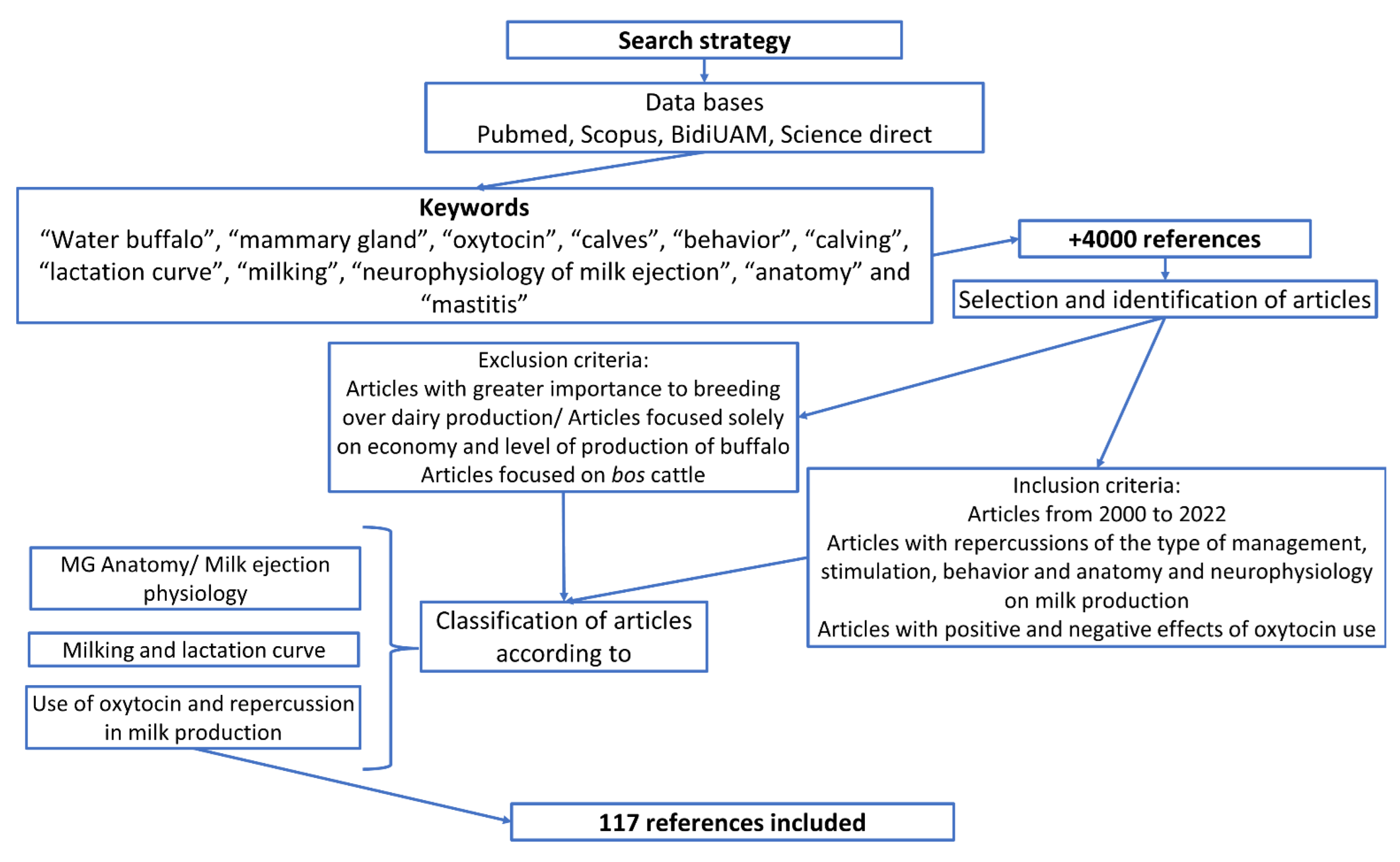
2. Materials and Methods
3. The Mammary Gland: Pros and Contras Advantages and Disadvantages for Milk Ejection
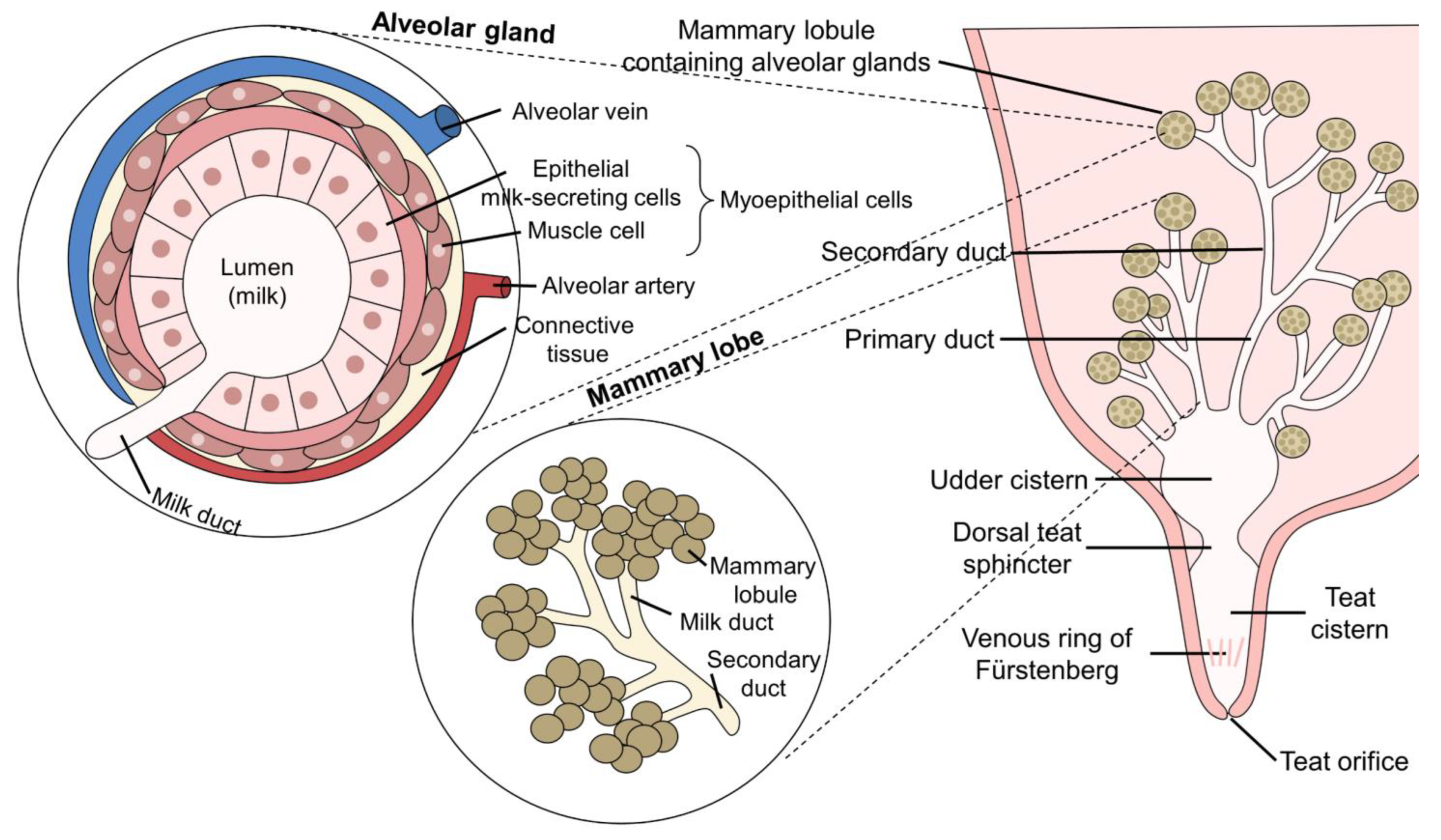
3.1. Influence of Anatomical Characteristics on Milking Capacity and the Lactation Curve
3.2. Frequency of Mastitis in Dairy Buffaloes and Its Relation to the Anatomy of the Mammary Gland
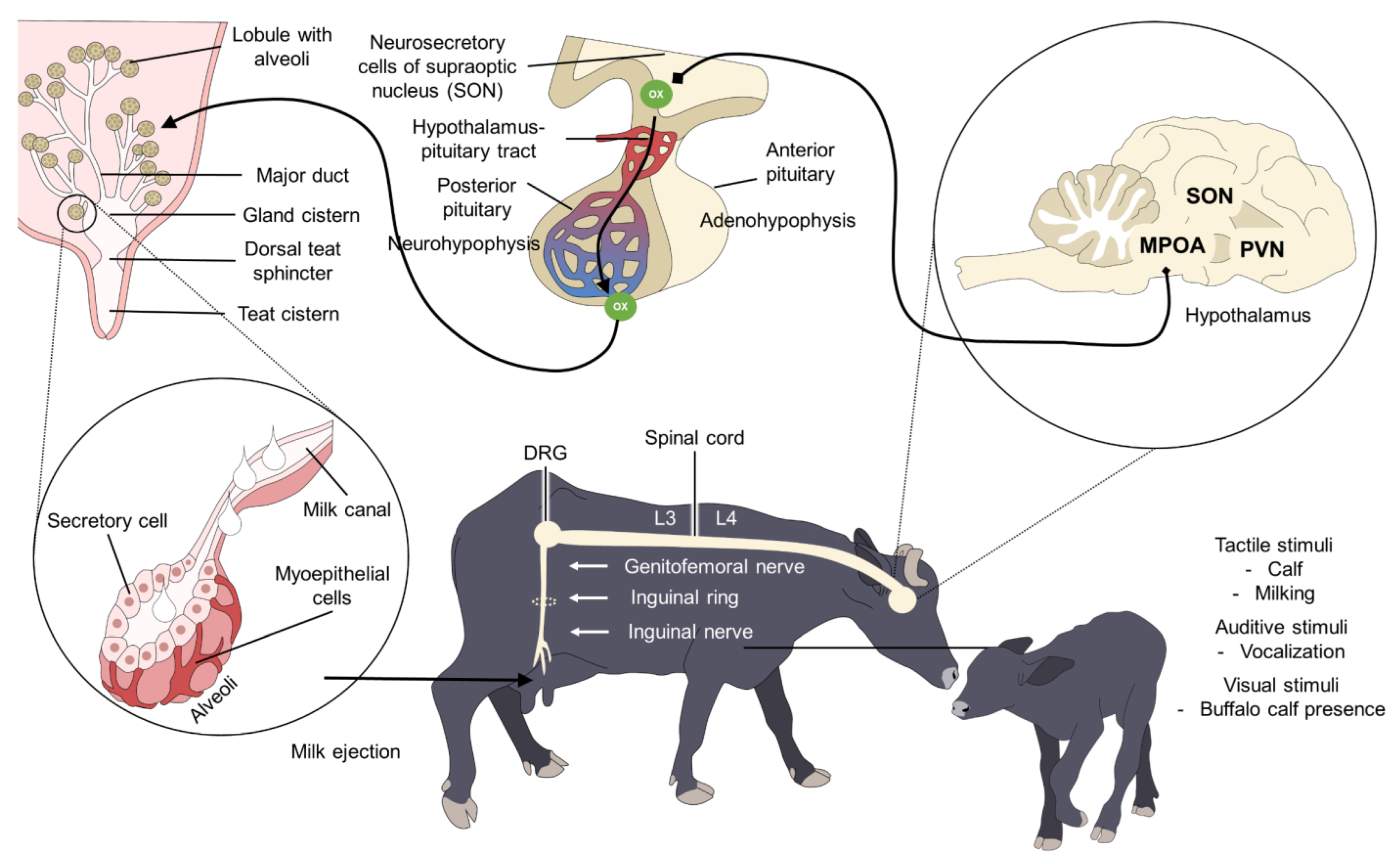
4. Neurophysiology of Milk Ejection
Alterations of Milk Ejection
5. Factors That Condition Affecting Milking Capacity in Dairy Buffalo
5.1. Presence of the Calf during Milking in Buffalo Cows
5.2. Feeding during Milking
5.3. Manual and Auditory Stimulation (Music)
5.4. Mechanical and Manual Milking
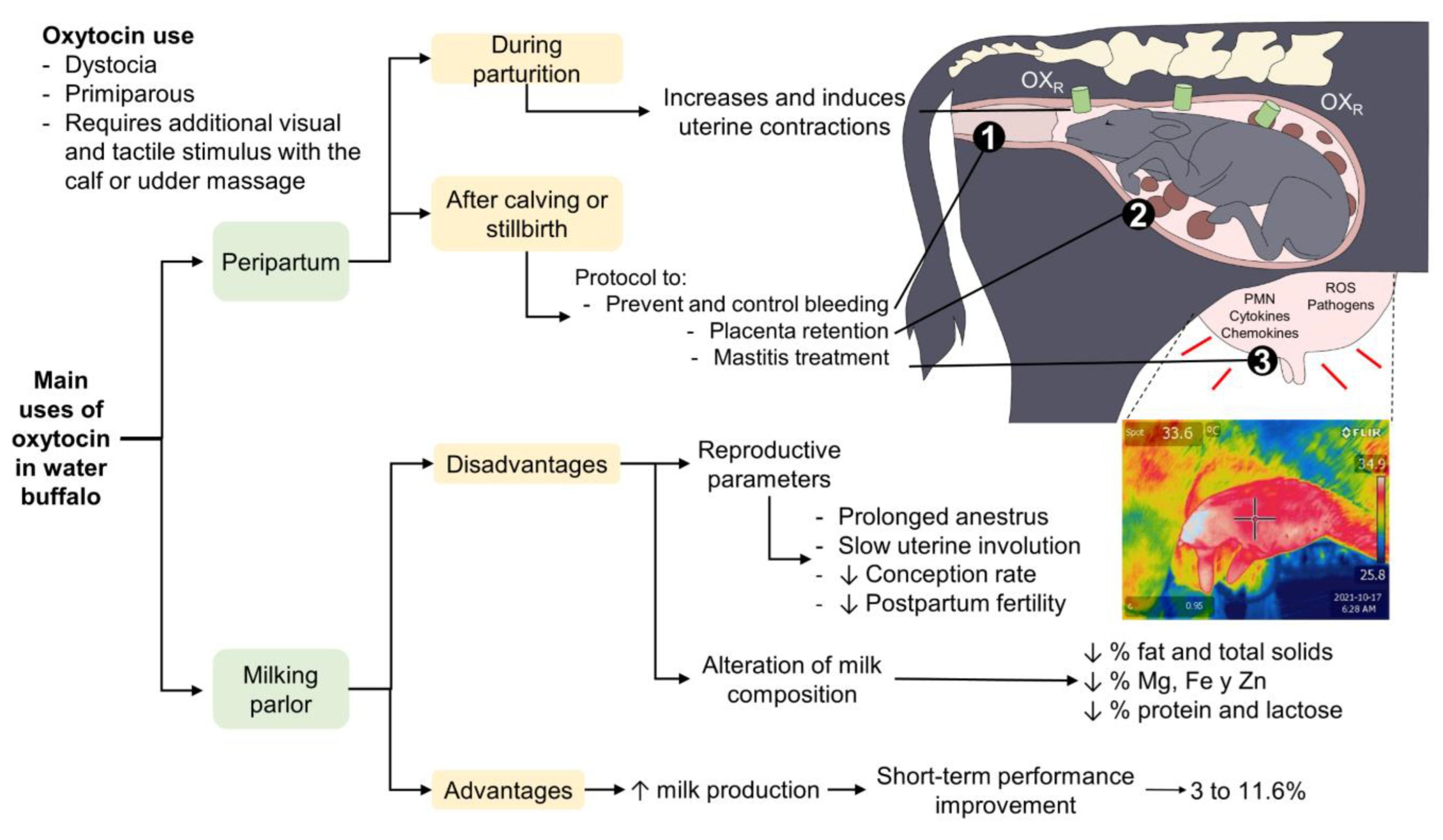
6. Use and Efficacy of Oxytocin in Milk Ejection
6.1. Effect on the Synthesis and Production of Milk
6.2. Effect on Nutritional Characteristics of the Milk and Somatic Cells
6.3. Other Effects
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barboza-Jiménez, G. Bondades ecológicas del búfalo de agua: Camino hacia la certificación. Tecnol. Marcha 2011, 24, 82–88. [Google Scholar]
- Bertoni, A.; Álvarez-Macías, A.; Mota-Rojas, D.; Álvarez-Macias, A.; Mota-Rojas, D. Productive performance of buffaloes and their development options in tropical regions. Soc. Rural Prod. Med. Amb. 2019, 19, 59–80. [Google Scholar]
- Ghezzi, M.D.; Napolitano, F.; Mota-Rojas, D.; Martínez, G.M.; de Rosa, G.; Álvarez-Macías, A.; Braghieri, A.; Bertoni, A.; Gómez-Prado, J.; Torres-Bernal, F.E.I.; et al. La búfala de agua en la producción de leche: Una visión internacional. In El Búfalo de Agua en las Américas, 3rd ed.; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; BM Editores: Mexico City, Mexico, 2020; pp. 257–328. Available online: https://lifescienceglobal.com/journals/journal-of-buffalo-science/97-abstract/jbs/4550-el-bufalo-de-agua-en-latinoamerica-hallazgos-recientes (accessed on 15 August 2022).
- Mota-Rojas, D.; Napolitano, F.; Braghieri, A.; de Rosa, G.; Álvarez-Macías, A.; Mora-Medina, P.; de Varona, E.; Bertoni, A.; Berdugo-Gutiérrez, J.; Masucci, F. Comportamiento y bienestar de la búfala lechera. In El Búfalo de Agua en Latinoamérica, Hallazgos Recientes, 3rd ed.; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; BM Editores: Mexico City, Mexico, 2020; pp. 95–130. Available online: https://lifescienceglobal.com/journals/journal-of-buffalo-science/97-abstract/jbs/4550-el-bufalo-de-agua-en-latinoamerica-hallazgos-recientes (accessed on 15 August 2022).
- Rodríguez-González, D.; Hamad, A.M.; Orihuela, A.; Bertoni, A.; Morales-Canela, D.A.; Álvarez-Macías, A.; José-Pérez, N.; Domínguez-Oliva, A.; Mota-Rojas, D. Handling and physiological aspects of the dual-purpose water buffalo production system in the exican humid tropics. Animals 2022, 12, 608. [Google Scholar] [CrossRef] [PubMed]
- FAO. The Future of Food and Agriculture Trends and Challenges; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018; pp. 1–52. [Google Scholar]
- OECD-FAO. Agricultural Outlook 2022-2031; OECD Publishing: Paris, France, 2022; p. 363. [Google Scholar]
- Crudeli, G.A.; Konrad, J.L.; Patiño, E.M. Reproducción en Búfalas; Mongolia Ediciones: Buenos Aires, Argentina, 2016; Volume 1, pp. 1–4. [Google Scholar]
- Napolitano, F.; Mota-Rojas, D.; Guerrero-Legarreta, I.; Orihuela, A. El Búfalo de Agua en Latinoamérica, Hallazgos Recientes, 3rd ed.; BM Editores: Mexico City, Mexico, 2020; p. 1544. Available online: https://lifescienceglobal.com/journals/journal-of-buffalo-science/97-abstract/jbs/4550-el-bufalo-de-agua-en-latinoamerica-hallazgos-recientes (accessed on 15 August 2022).
- Boselli, C.; Borghese, A.; Della Marta, U.; Leto, A.; Filippetti, F. Study of milkability in Mediterranean Italian Buffalo cow raised in central Italy. Rev. CES Med. Vet. Zootec. 2016, 11, 155. [Google Scholar]
- Mota-Rojas, D.; de Rosa, G.; Mora-Medina, P.; Braghieri, A.; Guerrero-Legarreta, I.; Napolitano, F. Dairy buffalo behaviour and welfare from calving to milking. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2019, 14, 1–9. [Google Scholar] [CrossRef]
- Boselli, C.; de Marchi, M.; Costa, A.; Borghese, A. Study of milkability and its relation with milk yield and somatic cell in mediterranean italian water buffalo. Front. Vet. Sci. 2020, 7, 432. [Google Scholar] [CrossRef]
- Cavallina, R.; Roncoroni, C.; Campagna, M.C.; Minero, M.; Canali, E. Buffalo behavioural response to machine milking in early lactation. Ital. J. Anim. Sci. 2008, 7, 287–295. [Google Scholar] [CrossRef]
- Faraz, A.; Tauqir, N.A.; Waheed, A.; Hameed, A. Effect of exogenous oxytocin administration on the performance of lactating Nili Ravi Buffalo. Irian J. Appl. Anim. Sci. 2021, 11, 517–525. [Google Scholar]
- Faraz, A.; Waheed, A.; Nazir, M.M.; Hameed, A.; Tauqir, N.A.; Mirza, R.H.; Ishaq, H.M.; Bilal, R.M. Impact of oxytocin administration on milk quality, reproductive performance and residual effects in dairy animals—A review. Punjab Univ. J. Zool. 2020, 35, 61–67. [Google Scholar] [CrossRef]
- Truchet, S.; Honvo-Houéto, E. Physiology of milk secretion. Best Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 367–384. [Google Scholar] [CrossRef]
- Colleluori, G.; Perugini, J.; Barbatelli, G.; Cinti, S. Mammary gland adipocytes in lactation cycle, obesity and breast cancer. Rev. Endocr. Metab. Disord. 2021, 22, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Stelwagen, K.; Carpenter, E.; Haigh, B.; Hodgkinson, A.; Wheeler, T.T. Immune components of bovine colostrum and milk. J. Anim. Sci. 2009, 87, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coroian, A.; Erler, S.; Matea, C.T.; Mireșan, V.; Răducu, C.; Bele, C.; Coroian, C.O. Seasonal changes of buffalo colostrum: Physicochemical parameters, fatty acids and cholesterol variation. Chem. Cent. J. 2013, 7, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrath, B.A.; Fox, P.F.; McSweeney, P.L.H.; Kelly, A.L. Composition and properties of bovine colostrum: A review. Dairy Sci. Technol. 2016, 96, 133–158. [Google Scholar] [CrossRef] [Green Version]
- Ashmawy, N. Chemical composition, hormonal levels and immunoglobin G concentration in colostrums, milk and blood plasma of Egyptian buffaloes following calving. Int. J. Adv. Res. 2015, 3, 471–478. [Google Scholar]
- Furman-Fratczak, K.; Rzasa, A.; Stefaniak, T. The influence of colostral immunoglobulin concentration in heifer calves’ serum on their health and growth. J. Dairy Sci. 2011, 94, 5536–5543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGee, M.; Earley, B. Review: Passive immunity in beef-suckler calves. Animal 2019, 13, 810–825. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.W.; Marnet, P.-G.; Yart, L.; Haenlein, G.F.W. Mammary secretion and lactation. In Milk and Dairy Products in Human Nutrition; Park, Y.W., Haenlein, G.F.W., Eds.; John Wiley & Sons: Oxford, MA, USA, 2013; pp. 31–45. [Google Scholar]
- Tătaru, M.; Stan, F.; Martonoș, C.O.; Gal, A.; Mârza, S.M.; Purdoiu, R.C.; Lăcătuș, R.; Damian, A.; Șonea, C.; Mireșan, V.; et al. Morphology of the mammary gland in romanian buffalo. Anat. Histol. Embryol. 2022, 51, 250–258. [Google Scholar] [CrossRef]
- Rodrigues, A.F.; Corrêa, E.S. Aspectos relacionados à búfala leiteira. Rev. Bras. Reprod. Anim. 2005, 29, 111–117. [Google Scholar]
- Olmos-Hernández, S.A.; Ghezzi, M.D.; Napolitano, F.; Cuibus, A.; Álvarez-Macías, A.; Braghieri, A.; Mota-Rojas, D. Anatomofisiología de la glándula mamaria: Neuroendocrinología de la eyección láctea en la búfala de agua. In El Búfalo de Agua en Latinoamérica, Hallazgos Recientes, 3rd ed.; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; BM Editores: Mexico City, Mexico, 2020; pp. 720–771. Available online: https://lifescienceglobal.com/journals/journal-of-buffalo-science/97-abstract/jbs/4550-el-bufalo-de-agua-en-latinoamerica-hallazgos-recientes (accessed on 15 August 2022).
- Borghese, A.; Boselli, C.; Rosati, R. Lactation curve and milk flow. Buffalo Bull. 2013, 32, 334–350. [Google Scholar]
- Bertoni, A.; Napolitano, F.; Mota-Rojas, D.; Strappini, A.C.; Sabia, E.; Álvarez-Macías, A.; Braghieri, A.; Mora-Medina, P.; Di Francia, A.; Morales-Canela, A.; et al. Comparative study between water buffaloes and conventional cattle: Zootechnical and morphophysiological aspects. In El Búfalo de Agua en las Américas: Comportamiento y Productividad, 4th ed.; Napolitano, F., Mota-Rojas, D., Orihuela, A., Braghieri, A., Hufana-Duran, D., Strappini, A.C., Pereira, A.M.F., Ghezzi, M.D., Guerrero-Legarreta, I., Martínez-Burnes, J., Eds.; B.M. Editores: Mexico City, Mexico, 2022; pp. 43–68. Available online: https://www.researchgate.net/profile/Fabio-Napolitano-2 (accessed on 15 August 2022).
- El-Khodery, S.A.; Osman, S.A. Cryptosporidiosis in buffalo calves (Bubalus bubalis): Prevalence and potential risk factors. Trop. Anim. Health Prod. 2008, 40, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, Y.; Ponce, P.; Capdevila, J. Efecto de la estimulación con bucerro, oxitocina y manual sobre los indicarores de ordeño en búfalas. Rev. Salud Anim. 2011, 33, 90–96. [Google Scholar]
- Guler, O.; Yanar, M.; Aydin, R.; Bayram, B.; Dogru, U.; Kopuzlu, S. Genetic and environmental parameters of milkability traits in Holstein Friesian cows. CAB Rev. 2009, 8, 143–147. [Google Scholar]
- Bertoni, A.; Martínez, G.M.; Napolitano, F.; Álvarez-Macías, A.; Ghezzi, M.D.; Mora-Medina, P.; Braghieri, A.; Guerrero-Legarreta, I.; Cruz-Monterrosa, R.; Daniel, M.-R.; et al. Ventajas y desventajas de los sistemas de ordeño manual y mecánico: Productividad, bienestar animal y rentabilidad. In El Búfalo de Agua en Latinoamérica, Hallazgos Recientes, 3rd ed.; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; BM Editores: Mexico City, Mexico, 2020; pp. 772–807. [Google Scholar]
- Bruckmaier, R.M.; Hilger, M. Milk ejection in dairy cows at different degrees of udder filling. J. Dairy Res. 2001, 68, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.S.; Svennersten-Sjaunja, K.; Bhosrekar, M.R.; Bruckmaier, R.M. Mammary cisternal size, cisternal milk and milk ejection in Murrah buffaloes. J. Dairy Res. 2004, 71, 162–168. [Google Scholar] [CrossRef]
- Borghese, A.; Rasmussen, M.; Thomas, C.S. Milking management of dairy buffalo. Ital. J. Anim. Sci. 2007, 6, 39–50. [Google Scholar] [CrossRef]
- Costa, A.; de Marchi, M.; Visentin, G.; Campagna, M.C.; Borghese, A.; Boselli, C. The effect of pre-milking stimulation on teat morphological parameters and milk traits in the Italian Water Buffalo. Front. Vet. Sci. 2020, 7, 572422. [Google Scholar] [CrossRef]
- Patel, H.A.; Srivastava, A.K.; Chauhan, H.D.; Patel, J.B. Parity effect on milk let-down time in Mehsana buffaloes. Adv. Anim. Vet. Sci. 2017, 5, 234–238. [Google Scholar] [CrossRef]
- Ambord, S.; Thomas, C.S.; Borghese, A.; Mazzi, M.; Boselli, C.; Bruckmaier, R.M. Teat anatomy, vacuum to open the teat canal, and fractionized milk composition in Italian buffaloes. CAB Direct 2009, 64, 351–353. [Google Scholar]
- Boselli, C.; Rosati, R.; Giangolini, G.; Arcuri, S.; Fagiolo, A.; Ballico, S.; Borghese, A. Milk Flow Measurements in Buffalo Herds. In Proceedings of the 7th World Buffalo Congress, Manila, Philippines, 4–19 October 2004. [Google Scholar]
- Gutiérrez-Valencia, A.; Hurtado-Lugo, N.; Cerón-Muñoz, M. Estimativas de factores de corrección para duración de la lactancia, edad y época de parto en búfalas de la Costa Atlántica Colombiana. Livest. Res. Rural Dev. 2006, 18, 19–23. [Google Scholar]
- Vázquez-Luna, D.; Lara-Rodríguez, D.A.; Ácar-Martínes, N.B. Búfalo de agua Bubalus bubalis parámetros zootécnicos en el sur de Veracruz, México. Available online: https://bmeditores.mx/ganaderia/bufalo-de-agua-bubalus-bubalis-parametros-zootecnicos-en-el-sur-de-veracruz-mexico/ (accessed on 15 August 2022).
- Napolitano, F.; Mota-Rojas, D.; Strappini, A.C.; Braghieri, A.; de Rosa, G.; Álvarez-Macías, A.; Ghezzi, M.D.; Mora-Medina, P.; de Varona, E.; Bertoni, A.; et al. Dairy buffalo behaviour and welfare. In El Búfalo de Agua en las Américas: Comportamiento y Productividad, 4th ed.; Napolitano, F., Mota-Rojas, D., Orihuela, A., Braghieri, A., Hufana-Duran, D., Strappini, A.C., Pereira, A.M.F., Ghezzi, M.D., Guerrero-Legarreta, I., Martínez- Burnes, J., Eds.; B.M. Editores: Mexico City, Mexico, 2022; pp. 111–132. [Google Scholar]
- Rodríguez-González, D.; Mota-Rojas, D.; Cruz-Monterrosa, R.G.; Guerrero-Legarreta, I. Neurofisiología en la Eyección Láctea en la Búfala de Agua: Perspectivas y Factores que Afectan su Productividad. Available online: https://www.ganaderia.com/destacado/neurofisiologia-en-la-eyeccion-lactea-en-la-bufala-de-agua-perspectivas-y-factores-que-afectan-su-productividad (accessed on 15 August 2022).
- Caria, M.; Boselli, C.; Murgia, L.; Rosati, R.; Pazzona, A. Effect of vacuum level on milk flow traits in Mediterranean Italian buffalo cow. Ital. J. Anim. Sci. 2012, 11, e25. [Google Scholar] [CrossRef]
- Tonhati, H.; Seno, L.O.; Otaviano, A.R. Uso de polinômios segmentados para o estudo da curva de lactação em Búfalos Murrah e seus mestiços en sistema de criação extensivo, no Estado de São Paulo Use of segmented polynomials to study the lactation curve of Murrah and their crossed breeds of buf. Arch. Latinoam. Prod. Anim. 2003, 13, 19–23. [Google Scholar]
- Urbańska, D.; Puchała, R.; Jarczak, J.; Czopowicz, M.; Kaba, J.; Horbańczuk, K.; Bagnicka, E. Does small ruminant lentivirus infection in goats predispose to bacterial infection of the mammary gland? A preliminary study. Animals 2021, 11, 1851. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, G.; Grasso, F.; Braghieri, A.; Bilancione, A.; Di Francia, A.; Napolitano, F. Behavior and milk production of buffalo cows as affected by housing system. J. Dairy Sci. 2009, 92, 907–912. [Google Scholar] [CrossRef] [Green Version]
- Burvenich, C.; van Merris, V.; Mehrzad, J.; Diez-Fraile, A.; Duchateau, L. Severity of E. coli mastitis is mainly determined by cow factors. Vet. Res. 2003, 34, 521–564. [Google Scholar] [CrossRef] [Green Version]
- Seegers, H.; Fourichon, C.; Beaudeau, F. Production effects related to mastitis and mastitis economics in dairy cattle herds. Vet. Res. 2003, 34, 475–491. [Google Scholar] [CrossRef] [Green Version]
- Sharun, K.; Dhama, K.; Tiwari, R.; Gugjoo, M.B.; Iqbal Yatoo, M.; Patel, S.K.; Pathak, M.; Karthik, K.; Khurana, S.K.; Singh, R.; et al. Advances in therapeutic and managemental approaches of bovine mastitis: A comprehensive review. Vet. Q. 2021, 41, 107–136. [Google Scholar] [CrossRef]
- Guimarães, J.L.B.; Brito, M.A.V.P.; Lange, C.C.; Silva, M.R.; Ribeiro, J.B.; Mendonça, L.C.; Mendonça, J.F.M.; Souza, G.N. Estimate of the economic impact of mastitis: A case study in a Holstein dairy herd under tropical conditions. Prev. Vet. Med. 2017, 142, 46–50. [Google Scholar] [CrossRef]
- Romano, E.; Roma, R.; Tidona, F.; Giraffa, G.; Bragaglio, A. Dairy farms and life cycle assessment (LCA): The allocation criterion useful to estimate undesirable products. Sustainability 2021, 13, 4354. [Google Scholar] [CrossRef]
- Fareed, S.K.; Memon, K.H.; Kachiwal, A.B.; Azhar, S.; Brula, M.I.; Mehmood-ul-Hasan; Ali, M.; Khan, T.A. Prevalence and economic losses of reproductive disorders and mastitis in buffaloes at Karachi, Pakistan. Indian J. Anim. Res. 2016, 51, 1130–1133. [Google Scholar] [CrossRef] [Green Version]
- Lakshmi Kavitha, K.; Rajesh, K.; Satheesh, K.S.K.; Syama Sundar, N. Buffalo mastitis—Risk factors. Buffalo Bull. 2009, 28, 134–137. [Google Scholar]
- Sharma, A.; Sindhu, N. Occurrence of clinical and subclinical mastitis in buffaloes in the State of Haryana (India). Ital. J. Anim. Sci. 2007, 6, 965–967. [Google Scholar] [CrossRef]
- Elhaig, M.M.; Selim, A. Molecular and bacteriological investigation of subclinical mastitis caused by Staphylococcus aureus and Streptococcus agalactiae in domestic bovids from Ismailia, Egypt. Trop. Anim. Health Prod. 2015, 47, 271–276. [Google Scholar] [CrossRef]
- De Boyer des Roches, A.; Faure, M.; Lussert, A.; Herry, V.; Rainard, P.; Durand, D.; Foucras, G. Behavioral and patho-physiological response as possible signs of pain in dairy cows during Escherichia coli mastitis: A pilot study. J. Dairy Sci. 2017, 100, 8385–8397. [Google Scholar] [CrossRef] [Green Version]
- Rainard, P.; Riollet, C. Innate immunity of the bovine mammary gland. Vet. Res. 2006, 37, 369–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guccione, J. Mastitis in Mediterranean Buffaloes. J. Dairy Vet. Sci. 2017, 2, 555596. [Google Scholar] [CrossRef]
- Fagiolo, A.; Lai, O. Mastitis in buffalo. Ital. J. Anim. Sci. 2007, 6, 200–206. [Google Scholar] [CrossRef]
- Bertoni, A.; Álvarez-Macías, A.; Dávalos-Flores, J.L.; Mota-Rojas, D. Productividad de los búfalos de agua y sus expectativas de desarrollo en zonas tropicales. In El Búfalo de Agua en las Américas, 3rd ed.; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; BM Editores: Mexico City, Mexico, 2020; pp. 167–207. Available online: https://lifescienceglobal.com/journals/journal-of-buffalo-science/97-abstract/jbs/4550-el-bufalo-de-agua-en-latinoamerica-hallazgos-recientes (accessed on 15 August 2022).
- Bidarimath, M.; Aggarwal, A. Studies on cisternal and alveolar fractions & its composition and mammary health of Murrah buffaloes administered oxytocin. Trop. Anim. Health Prod. 2007, 39, 433–438. [Google Scholar] [CrossRef]
- Dûidić, A. Studies on Milk Ejection and Milk Removal during Machine Milking in Different Species. Ph.D. Thesis, Fakultät Wissenschaftszentrum Weihenstephan, Munich, Germany, 2004. [Google Scholar]
- Cosenza, G.; Pauciullo, A.; Mancusi, A.; Nicodemo, D.; Di Palo, R.; Zicarelli, L.; Di Berardino, D.; Ramunno, L. Mediterranean river buffalooxytocin-neurophysin I (OXT) gene: Structure, promoter analysis and allele detection. Ital. J. Anim. Sci. 2007, 6, 303–306. [Google Scholar] [CrossRef]
- Akhtar, M.S.; Lodhi, L.A.; Farooq, A.A.; Ayaz, M.M.; Maqbool Hussain, M.; Chaudhary, H.L.Z.I. Effect of Oxytocin administration before milking on milk production, somatic cells count and fat contents in milk of Nili-Ravi Buffaloes. Pak. Vet. J. 2012, 32, 435–437. [Google Scholar] [CrossRef] [Green Version]
- Bruckmaier, R.M. Normal and disturbed milk ejection in dairy cows. Domest. Anim. Endocrinol. 2005, 29, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Crowley, W.R.; Armstrong, W.E. Neurochemical regulation of oxytocin secretion in lactation. Endocr. Rev. 1992, 13, 33–65. [Google Scholar] [CrossRef] [PubMed]
- Kamikawa, A.; Seko, J. Physiological and pharmacological evaluation of oxytocin-induced milk ejection in mice. Exp. Anim. 2020, 69, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Tallo-Parra, O.; Carbajal, A.; Monclús, L.; Manteca, X.; Lopez-Bejar, M. Hair cortisol and progesterone detection in dairy cattle: Interrelation with physiological status and milk production. Domest. Anim. Endocrinol. 2018, 64, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Stelwagen, K.; van Espen, D.; Verkerk, G.; McFadden, H.; Farr, V. Elevated plasma cortisol reduces permeability of mammary tight junctions in the lactating bovine mammary epithelium. J. Endocrinol. 1998, 159, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Mayer, H.K.; Lefcourt, A.M. Failure of cortisol injected prior to milking to inhibit milk ejection in dairy cattle. J. Dairy Res. 1987, 54, 173–177. [Google Scholar] [CrossRef]
- Bruckmaier, R.M.; Wellnitz, O. Induction of milk ejection and milk removal in different production systems. J. Anim. Sci. 2008, 86, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Tancin, V.; Bruckmaier, R.M. Factors affecting milk ejection and removal during milking and suckling of dairy cows. Vet. Med. 2001, 46, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.S. Milking Management of Dairy Buffaloes. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2004. [Google Scholar]
- Caria, M.; Murgia, L.; Pazzona, A. Effects of the working vacuum level on mechanical milking of buffalo. J. Dairy Sci. 2011, 94, 1755–1761. [Google Scholar] [CrossRef] [Green Version]
- Bava, L.; Sandrucci, A.; Tamburini, A.; Zucali, M. Milk flow traits of buffalo cows in intensive farming system. Ital. J. Anim. Sci. 2007, 6, 500–502. [Google Scholar] [CrossRef]
- Belo, C.J.; Bruckmaier, R.M. Suitability of low-dosage oxytocin treatment to induce milk ejection in dairy cows. J. Dairy Sci. 2010, 93, 63–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boselli, C.; Campagna, M.C.; Amatiste, S.; Rosati, R.; Borghese, A. Pre-stimulation effects on teat anatomy and milk flow curves in mediterranean Italian buffalo cows. J. Anim. Vet. Adv. 2014, 13, 912–916. [Google Scholar] [CrossRef]
- Wellnitz, O.; Bruckmaier, R.M. Central and peripheral inhibition of milk ejection. Livest. Prod. Sci. 2001, 70, 135–140. [Google Scholar] [CrossRef]
- Erskine, R.J.; Norby, B.; Neuder, L.M.; Thomson, R.S. Decreased milk yield is associated with delayed milk ejection. J. Dairy Sci. 2019, 102, 6477–6484. [Google Scholar] [CrossRef]
- Moore-Foster, R.; Norby, B.; Schewe, R.L.; Thomson, R.; Bartlett, P.C.; Erskine, R.J. Herd-level variables associated with delayed milk ejection in Michigan dairy herds. J. Dairy Sci. 2019, 102, 696–705. [Google Scholar] [CrossRef] [Green Version]
- Lollivier, V.; Guinard-Flament, J.; Ollivier-Bousquet, M.; Marnet, P.-G. Oxytocin and milk removal: Two important sources of variation in milk production and milk quality during and between milkings. Reprod. Nutr. Dev. 2002, 42, 173–186. [Google Scholar] [CrossRef]
- Watters, R.D.; Bruckmaier, R.M.; Crawford, H.M.; Schuring, N.; Schukken, Y.H.; Galton, D.M. The effect of manual and mechanical stimulation on oxytocin release and milking characteristics in Holstein cows milked 3 times daily. J. Dairy Sci. 2015, 98, 1721–1729. [Google Scholar] [CrossRef] [Green Version]
- Weiss, D.; Bruckmaier, R.M. Optimization of individual prestimulation in dairy cows. J. Dairy Sci. 2005, 88, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Mora-Medina, P.; Napolitano, F.; Mota-Rojas, D.; Berdugo-Gutiérrez, J.; Ruiz-Buitrago, J.; Guerrero-Legarreta, I. Imprinting, sucking and allosucking behaviors in buffalo calves. J. Buffalo Sci. 2018, 7, 49–57. [Google Scholar] [CrossRef]
- Singh, P.; Kamboj, M.L.; Chandra, S.; Singh, R.K. Effect of claf suckling dummy calf used and weaning on milk ejection stimuli and milk yield of Murrah buffaloes (Bubalus bubalis). J. Pharmacog. Phytochim. 2017, 6, 1012–1015. [Google Scholar]
- Oliveira, A.D.F.M.; Quirino, C.R.; Bastos, R. Effect of nursing behaviour, sex of the calf, and parity order on milk production of buffaloes. Rev. Colomb. Cienc. Pecu. 2017, 30, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.S.; Bruckmaier, R.M.; Östensson, K.; Svennersten-Sjaunja, K. Effect of different milking routines on milking-related release of the hormones oxytocin, prolactin and cortisol, and on milk yield and milking performance in Murrah buffaloes. J. Dairy Res. 2005, 72, 10–18. [Google Scholar] [CrossRef]
- Ambord, S.; Stoffel, M.H.; Bruckmaier, R.M. Teat anatomy affects requirements for udder preparation in Mediterranean buffaloes. J. Dairy Res. 2010, 77, 468–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tangorra, F.M.; Leonardi, S.; Bronzo, V.; Rota, N.; Moroni, P. Pre-milking mechanical teat stimulation and milking performance of dairy buffaloes in early lactation. J. Agric. Eng. 2017, 48, 53–55. [Google Scholar] [CrossRef]
- Arnold, N.A.; Ng, K.T.; Jongman, E.C.; Hemsworth, P.H. The behavioural and physiological responses of dairy heifers to tape-recorded milking facility noise with and without a pre-treatment adaptation phase. Appl. Anim. Behav. Sci. 2007, 106, 13–25. [Google Scholar] [CrossRef]
- Uetake, K.; Hurnik, J.F.; Johnson, L. Effect of music on voluntary approach of dairy cows to an automatic milking system. Appl. Anim. Behav. Sci. 1997, 53, 175–182. [Google Scholar] [CrossRef]
- Ciborowska, P.; Michalczuk, M.; Bień, D. The effect of music on livestock: Cattle, poultry and pigs. Animals 2021, 11, 3572. [Google Scholar] [CrossRef]
- Polikarpus, A.; Grasso, F.; Pacelli, C.; Napolitano, F.; de Rosa, G. Milking behaviour of buffalo cows: Entrance order and side preference in the milking parlour. J. Dairy Res. 2014, 81, 24–29. [Google Scholar] [CrossRef]
- Sannino, M.; Faugno, S.; Crimaldi, M.; Di Francia, A.; Ardito, L.; Serrapica, F.; Masucci, F. Effects of an automatic milking system on milk yield and quality of Mediterranean buffaloes. J. Dairy Sci. 2018, 101, 8308–8312. [Google Scholar] [CrossRef]
- Singh, M.; Prakash, B.S.; Mallick, S. Hormone release, milk production and composition in Murrah buffaloes milked by hand and machine. Indian J. Anim. Res. 2014, 48, 444. [Google Scholar] [CrossRef]
- Polikarpus, A.; Napolitano, F.; Grasso, F.; Di Palo, R.; Zicarelli, F.; Arney, D.; de Rosa, G. Effect of pre-partum habituation to milking routine on behaviour and lactation performance of buffalo heifers. Appl. Anim. Behav. Sci. 2014, 161, 1–6. [Google Scholar] [CrossRef]
- Ni, Y.; Chen, Q.; Cai, J.; Xiao, L.; Zhang, J. Three lactation-related hormones: Regulation of hypothalamus-pituitary axis and function on lactation. Mol. Cell. Endocrinol. 2021, 520, 111084. [Google Scholar] [CrossRef] [PubMed]
- Nwosu, E.U.; James, I.J.; Olowofeso, O.; Williams, T.J.; Bemji, M.N. Effects of oxytocin administration on milk yield and milk composition of West African Dwarf does. Small Rumin. Res. 2019, 181, 45–50. [Google Scholar] [CrossRef]
- Weiss, D.; Dzidic, A.; Bruckmaier, R.M. Effect of stimulation intensity on oxytocin release before, during and after machine milking. J. Dairy Res. 2003, 70, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Nostrand, S.D.; Galton, D.M.; Erb, H.N.; Bauman, D.E. Effects of daily exogenous oxytocin on lactation milk yield and composition. J. Dairy Sci. 1991, 74, 2119–2127. [Google Scholar] [CrossRef]
- Ballou, L.U.; Bleck, J.L.; Bleck, G.T.; Bremel, R.D. The effects of daily oxytocin injections before and after milking on milk production, milk plasmin, and milk composition. J. Dairy Sci. 1993, 76, 1544–1549. [Google Scholar] [CrossRef]
- Singh, M.; Aggarwal, A. Effect of oxytocin administration on certain mineralis in milk of buffaloes (Bubalus bubalis). Asian-Australas. J. Anim. Sci. 2001, 11, 1523–1526. [Google Scholar] [CrossRef]
- Lollivier, V.; Marnet, P.G. Galactopoietic effect of milking in lactating Holstein cows: Role of physiological doses of oxytocin. Livest. Prod. Sci. 2005, 95, 131–142. [Google Scholar] [CrossRef]
- Rushen, J.; de Passillé, A.M. The importance of good stockmanship and its benefits to animals. In Improving Animal Welfare: A Practical Approach; Grandin, T., Ed.; CAB International: Wallingford, UK, 2010; pp. 50–63. [Google Scholar]
- Sitkowska, B. Effect of the cow age group and lactation stage on the count of somatic cells in cow milk. J. Cent. Eur. Agric. 2008, 9, 57–61. [Google Scholar]
- Bencini, R.; Martin, G.; Purvis, I.; Hartmann, P. Use of oxytocin to measure milk output in Merino ewes and its effect on fat content. Aust. J. Exp. Agric. 1992, 32, 601. [Google Scholar] [CrossRef]
- Kiran, U. Effect of Intramuscular Injection of Oxytocin on Milk Composition. Ph.D. Thesis, University of Agriculture, Faisalabad, Pakistan, 2001. [Google Scholar]
- Dang, A.K.; Anand, S.K. Effect of milking systems on the milk somatic cell counts and composition. Livest. Res. Rural Dev. 2007, 19, 771. [Google Scholar]
- Mustafa, M.Y.; Saleem, K.; Munir, R.; Butt, T.M. Effect of oxytocin on the productive and reproductive performance of buffalo and cattle in Sheikhupura-Pakistan (A field study). Livest. Res. Rural Dev. 2008, 20, 45. [Google Scholar]
- Qureshi, M.S.; Ahmad, N. Interaction of calf suckling, use of oxytocin and milk yield with reproductive performance of dairy buffaloes. Anim. Reprod. Sci. 2008, 106, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M. Oxytocin; Effects on milk production. Pure Appl. Biol. 2021, 10, 318–324. [Google Scholar] [CrossRef]
- Bertoni, A.; Napolitano, F.; Mota-Rojas, D.; Sabia, E.; Álvarez-Macías, A.; Mora-Medina, P.; Morales-Canela, A.; Berdugo-Gutiérrez, J.; Guerrero- Legarreta, I. Similarities and differences between river buffaloes and cattle: Health, physiological, behavioral and productivity aspects. J. Buffalo Sci. 2020, 9, 92–109. [Google Scholar] [CrossRef]
- Murtaza, S.; Sattar, A.; Ahmad, N.; Jamil Ahmad, M.; Akhtar, S.; Ahmad, E.; Ahmad, T.; Omer, T. Effect of exogenous administration of oxytocin on postpartum follicular dynamics, oestrous rate and ovulation in Nili-Ravi buffaloes. Reprod. Domest. Anim. 2021, 56, 1369–1376. [Google Scholar] [CrossRef]
- Napolitano, F.; Mota-Rojas, D.; Alvarez, A.; Braghieri, A. Factores productivos y su incidencia en el bienestar de la búfala lechera en sistemas de producción extensivos e intensivos. Soc. Rur. Prod. Med. Amb. 2020, 20, 155–173. [Google Scholar]
- Ozenc, E.; Bozkurt, M.F.; Yazici, E.; Seker, E.; Bayraktaroglu, A.G.; Ozcinar, U.; Dogan, N. Teat characteristics in relation to animal temperament during milking in buffaloes, and comparison of buffalo and cow teat morphology. Reprod. Domest. Anim. 2020, 55, 559–566. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Napolitano, F.; Braghieri, A.; Bragaglio, A.; Rodríguez-González, D.; Mora-Medina, P.; Ghezzi, M.D.; Álvarez-Macías, A.; Lendez, P.A.; Sabia, E.; Domínguez-Oliva, A.; et al. Neurophysiology of Milk Ejection and Prestimulation in Dairy Buffaloes. Animals 2022, 12, 2649. https://doi.org/10.3390/ani12192649
Napolitano F, Braghieri A, Bragaglio A, Rodríguez-González D, Mora-Medina P, Ghezzi MD, Álvarez-Macías A, Lendez PA, Sabia E, Domínguez-Oliva A, et al. Neurophysiology of Milk Ejection and Prestimulation in Dairy Buffaloes. Animals. 2022; 12(19):2649. https://doi.org/10.3390/ani12192649
Chicago/Turabian StyleNapolitano, Fabio, Ada Braghieri, Andrea Bragaglio, Daniela Rodríguez-González, Patricia Mora-Medina, Marcelo Daniel Ghezzi, Adolfo Álvarez-Macías, Pamela Anahí Lendez, Emilio Sabia, Adriana Domínguez-Oliva, and et al. 2022. "Neurophysiology of Milk Ejection and Prestimulation in Dairy Buffaloes" Animals 12, no. 19: 2649. https://doi.org/10.3390/ani12192649