Effect of Flaxseed Supplementation on Milk and Plasma Fatty Acid Composition and Plasma Parameters of Holstein Dairy Cows

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cows and Experimental Design
2.2. Sampling, Measurements, and Analysis
2.3. Statistical Analysis
3. Results
3.1. Feed Fatty Acid Composition
3.2. Feed Intake, Milk Yield, and Milk Composition
3.3. Plasma Fatty Acid Composition
3.4. Milk Fatty Acid Composition
3.5. Antioxidant Indexes in Plasma and Milk
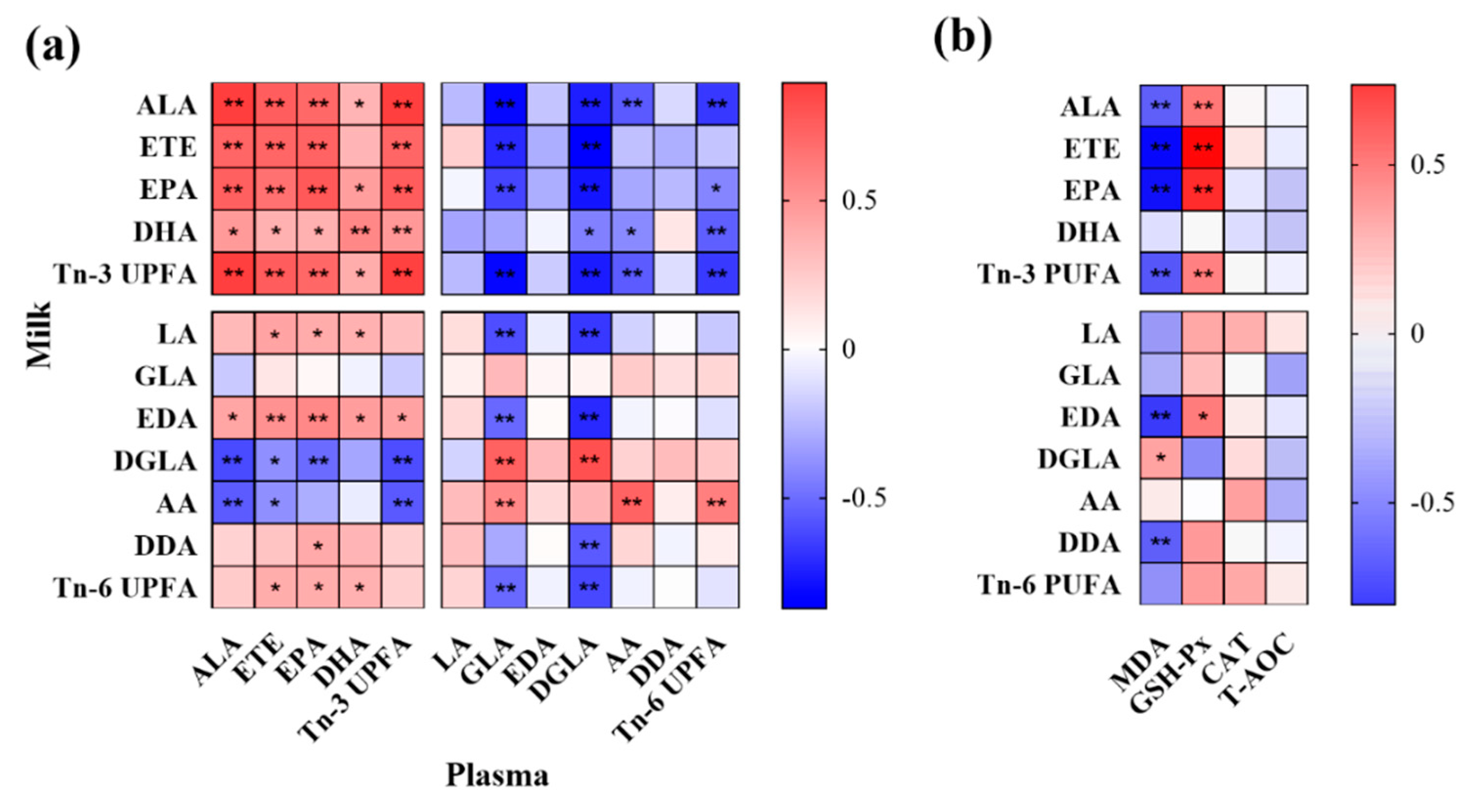
3.6. Correlation Analysis
3.7. Plasma Parameters
4. Discussion
4.1. Feed Intake and Milk Yield
4.2. Milk Composition
4.3. Plasma Fatty Acid Composition
4.4. Milk Fatty Acid Composition
4.5. Antioxidant Indexes in Plasma and Milk
4.6. Plasma Parameters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kabeya, N.; Fonseca, M.M.; Ferrier, D.E.K.; Navarro, J.C.; Bay, L.K.; Francis, D.S.; Tocher, D.R.; Castro, L.F.C.; Monroig, Ó. Genes for de novo biosynthesis of omega-3 polyunsaturated fatty acids are widespread in animals. Sci. Adv. 2018, 4, r6849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, R.K.; Keum, Y. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance-A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Fofana, B.; Cloutier, S.; Kirby, C.W.; McCallum, J.; Duguid, S.A. A well balanced omega-6/omega-3 ratio in developing flax bolls after heating and its implications for use as a fresh vegetable by humans. Food Res. Int. 2011, 44, 2459–2464. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 6, 674–688. [Google Scholar] [CrossRef]
- Simopoulos, A. An Increase in the Omega-6/Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Brzozowska, A.M.; Micek, P.; Kuczynska, B.; Lukaszewicz, M.; Oprzadek, J.M. Corrigendum to: The use of energy-protein supplement increases performance of high-yielding dairy cows and improves health-promoting properties of milk. Anim. Prod. Sci. 2018, 58, 1766. [Google Scholar] [CrossRef]
- Marino, R.; della Malva, A.; Caroprese, M.; de Palo, P.; Santillo, A.; Sevi, A.; Albenzio, M. Effects of whole linseed supplementation and treatment duration on fatty acid profile and endogenous bioactive compounds of beef muscle. Animal 2019, 13, 444–452. [Google Scholar] [CrossRef]
- Meignan, T.; Lechartier, C.; Chesneau, G.; Bareille, N. Effects of feeding extruded linseed on production performance and milk fatty acid profile in dairy cows: A meta-analysis. J. Dairy Sci. 2017, 100, 4394–4408. [Google Scholar] [CrossRef] [Green Version]
- Moallem, U.; Vyas, D.; Teter, B.B.; Delmonte, P.; Zachut, M.; Erdman, R.A. Transfer rate of α-linolenic acid from abomasally infused flaxseed oil into milk fat and the effects on milk fatty acid composition in dairy cows. J. Dairy Sci. 2012, 95, 5276–5284. [Google Scholar] [CrossRef] [Green Version]
- Pi, Y.; Ma, L.; Wang, H.R.; Wang, J.Q.; Xu, J.C.; Bu, D.P. Rubber seed oil and flaxseed oil supplementation on serum fatty acid profile, oxidation stability of serum and milk, and immune function of dairy cows. Asian Australas. J. Anim. 2019, 32, 1363–1372. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, A.F.; McKinnon, J.J.; Christensen, D.A.; He, T. Effects of micronization of flaxseed on nutrient disappearance in the gastrointestinal tract of steers. Anim. Feed Sci. Tech. 2002, 95, 123–132. [Google Scholar] [CrossRef]
- Petit, H.V. Digestion, milk production, milk composition, and blood composition of dairy cows fed formaldehyde treated flaxseed or sunflower seed. J. Dairy Sci. 2003, 86, 2637–2646. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Lopez, E.; Moats, J.; Aluthge, N.D.; Ramirez Ramirez, H.A.; Christensen, D.A.; Mutsvangwa, T.; Penner, G.B.; Fernando, S.C. Effect of partially replacing a barley-based concentrate with flaxseed-based products on the rumen bacterial population of lactating Holstein dairy cows. J. Appl. Poult. Res. 2018, 124, 42–57. [Google Scholar] [CrossRef] [Green Version]
- Isenberg, B.J.; Soder, K.J.; Pereira, A.B.; Standish, R.; Brito, A.F. Production, milk fatty acid profile, and nutrient utilization in grazing dairy cows supplemented with ground flaxseed. J. Dairy Sci. 2019, 102, 1294–1311. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, A.; Yuan, K.; Armendariz, C.K.; Hihland, G.; Bello, N.M.; Winowiski, T.; Drouillard, J.S.; Titgemeyer, E.C.; Bradford, B.J. Effects of urea formaldehyde condensation polymer treatment of flaxseed on ruminal digestion and lactation in dairy cows. J. Dairy Sci. 2013, 96, 3907–3915. [Google Scholar] [CrossRef]
- Ariza, J.M.; Meignan, T.; Madouasse, A.; Beaudeau, F.; Bareille, N. Effects on milk quantity and composition associated with extruded linseed supplementation to dairy cow diets. Sci. Rep. 2019, 9, 17563. [Google Scholar] [CrossRef]
- Sterk, A.R.; Vlaeminck, B.; Van Vuuren, A.M.; Hendriks, W.H.; Dijkstra, J. Effects of feeding different linseed sources on omasal fatty acid flows and fatty acid profiles of plasma and milk fat in lactating dairy cows. J. Dairy Sci. 2012, 95, 3149–3165. [Google Scholar] [CrossRef]
- Gómez-Cortés, P.; Juárez, M.; de la Fuente, M.A. Milk fatty acids and potential health benefits: An updated vision. Trends Food Sci. Tech. 2018, 81, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Leduc, M.; Létourneau-Montminy, M.P.; Gervais, R.; Chouinard, P.Y. Effect of dietary flax seed and oil on milk yield, gross composition, and fatty acid profile in dairy cows: A meta-analysis and meta-regression. J. Dairy Sci. 2017, 100, 8906–8927. [Google Scholar] [CrossRef]
- Giannuzzi, D.; Toscano, A.; Pegolo, S.; Gallo, L.; Tagliapietra, F.; Mele, M.; Minuti, A.; Trevisi, E.; Ajmone Marsan, P.; Schiavon, S.; et al. Associations between Milk Fatty Acid Profile and Body Condition Score, Ultrasound Hepatic Measurements and Blood Metabolites in Holstein Cows. Animals 2022, 12, 1202. [Google Scholar] [CrossRef]
- Lock, A.L.; Bauman, D.E. Modifying milk fat composition of dairy cows to enhance fatty acids beneficial to human health. Lipids 2004, 39, 1197–1206. [Google Scholar] [CrossRef] [Green Version]
- Mele, M.; Macciotta, N.P.P.; Cecchinato, A.; Conte, G.; Schiavon, S.; Bittante, G. Multivariate factor analysis of detailed milk fatty acid profile: Effects of dairy system, feeding, herd, parity, and stage of lactation. J. Dairy Sci. 2016, 99, 9820–9833. [Google Scholar] [CrossRef] [Green Version]
- Danesh, M.M.; Jafari, J.R.; Danesh, M.S. Milk production, milk fatty acid composition, and blood biochemical parameters of holstein dairy cows fed whole or ground flaxseed instead of extruded soybeans in the first half of lactation. Iran. J. Vet. Res. 2012, 3, 203–209. [Google Scholar]
- Huang, G.X.; Guo, L.Y.; Chang, X.F.; Liu, K.Z.; Tang, W.H.; Zheng, N.; Zhao, S.G.; Zhang, Y.D.; Wang, J.Q. Effect of Whole or Ground Flaxseed Supplementation on Fatty Acid Profile, Fermentation, and Bacterial Composition in Rumen of Dairy Cows. Front. Microbiol. 2021, 12, 760528. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis, 17th ed.; AOAC International: Arlington, VA, USA, 2000. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Sukhija, P.S.; Palmquist, D.L. Rapid method for determination of total fatty acid content and composition of feedstuffs and feces. J. Agr. Food Chem. 1988, 36, 1202–1206. [Google Scholar] [CrossRef]
- Tyrrell, H.F.; Reid, J.T. Prediction of the energy value of cow’s milk. J. Dairy Sci. 1965, 48, 1215–1223. [Google Scholar] [CrossRef]
- Bu, D.P.; Wang, J.Q.; Dhiman, T.R.; Liu, S.J. Effectiveness of Oils Rich in Linoleic and Linolenic Acids to Enhance Conjugated Linoleic Acid in Milk from Dairy Cows. J. Dairy Sci. 2007, 90, 998–1007. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.E.; Chen, M.Q.; Luo, R.B.; Huang, G.X.; Wu, X.F.; Zheng, N.; Zhang, Y.D.; Wang, J.Q. Fatty acid profiles of milk from Holstein cows, Jersey cows, buffalos, yaks, humans, goats, camels, and donkeys based on gas chromatography–mass spectrometry. J. Dairy Sci. 2022, 105, 1687–1700. [Google Scholar] [CrossRef]
- Allred, S.L.; Dhiman, T.R.; Brennand, C.P.; Khanal, R.C.; Mcmahon, D.J.; Luchini, N.D. Milk and Cheese from Cows Fed Calcium Salts of Palm and Fish Oil Alone or in Combination with Soybean Products. J. Dairy Sci. 2006, 89, 234–248. [Google Scholar] [CrossRef] [Green Version]
- National Research Council (NRC). Nutrient Requirements of Dairy Cattle, 7th rev. ed.; The National Academies Press: Washington, DC, USA, 2001.
- Resende, T.L.; Kraft, J.; Soder, K.J.; Pereira, A.B.D.; Brito, A.F. Incremental amounts of ground flaxseed decrease milk yield but increase n-3 fatty acids and conjugated linoleic acids in dairy cows fed high-forage diets. J. Dairy Sci. 2015, 98, 4785–4799. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Rouel, J.; Jouany, J.P.; Doreau, M.; Chilliard, Y. Methane output and diet digestibility in response to feeding dairy cows crude linseed, extruded linseed, or linseed oil. J. Anim. Sci. 2008, 86, 2642–2650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do Prado, R.M.; Palin, M.F.; Do Prado, I.N.; Dos Santos, G.T.; Benchaar, C.; Petit, H.V. Milk yield, milk composition, and hepatic lipid metabolism in transition dairy cows fed flaxseed or linola. J. Dairy Sci. 2016, 99, 8831–8846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurtaud, C.; Dutreuil, M.; Coppa, M.; Agabriel, C.; Martin, B. Characterization of milk from feeding systems based on herbage or corn silage with or without flaxseed and authentication through fatty acid profile. Dairy Sci. Technol. 2014, 94, 103–123. [Google Scholar] [CrossRef]
- Caroprese, M.; Marzano, A.; Marino, R.; Gliatta, G.; Muscio, A.; Sevi, A. Flaxseed supplementation improves fatty acid profile of cow milk. J. Dairy Sci. 2010, 93, 2580–2588. [Google Scholar] [CrossRef]
- Caroprese, M.; Mancino, R.; Ciliberti, M.G.; Luccia, A.D.; Gatta, B.L.; Albenzio, M. Fatty acid profile and coagulating ability of milk from Jersey and Friesian cows fed whole flaxseed. J. Dairy Res. 2017, 84, 14–22. [Google Scholar] [CrossRef]
- Lerch, S.; Ferlay, A.; Graulet, B.; Cirie, C.; Verdier-Metz, I.; Montel, M.C.; Chilliard, Y.; Martin, B. Extruded linseeds, vitamin E and plant extracts in corn silage-based diets of dairy cows: Effects on sensory properties of raw milk and uncooked pressed cheese. Int. Dairy J. 2015, 51, 65–74. [Google Scholar] [CrossRef]
- Graulet, B.; Cirié, C.; Martin, B. Contrasted effects of dietary extruded linseed supplementation on carotenoid and liposoluble vitamin status in lactating Holstein or Montbéliarde cows fed hay or corn silage. J. Dairy Sci. 2019, 102, 6210–6225. [Google Scholar] [CrossRef]
- Petit, H.V.; Germiquet, C.; Lebel, D. Effect of Feeding Whole, Unprocessed Sunflower Seeds and Flaxseed on Milk Production, Milk Composition, and Prostaglandin Secretion in Dairy Cows. J. Dairy Sci. 2004, 87, 3889–3898. [Google Scholar] [CrossRef] [Green Version]
- Stergiadis, S.; Leifert, C.; Seal, C.J.; Eyre, M.D.; Steinshamn, H.; Butler, G. Improving the fatty acid profile of winter milk from housed cows with contrasting feeding regimes by oilseed supplementation. Food Chem. 2014, 20, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, T.C.; McGuire, M.A. Major advances in nutrition: Impact on milk composition. J. Dairy Sci. 2006, 89, 1302–1310. [Google Scholar] [CrossRef]
- Oba, M.; Thangavelu, G.; Dehghan-Banadaky, M.; Ambrose, D.J. Unprocessed whole flaxseed is as effective as dry-rolled flaxseed at increasing α-linolenic acid concentration in milk of dairy cows. Livest. Sci. 2009, 122, 73–76. [Google Scholar] [CrossRef]
- Bashar, M.A.; Jumat, S. Toxicity Study of Malaysian Rubber (Hevea brasiliensis) Seed Oil as Rats and Shrimps Tests. Asian J. Biochem. 2009, 5, 33–39. [Google Scholar]
- He, M.L.; Chung, Y.H.; McAllister, T.A.; Beauchemin, K.A.; Mir, P.S.; Aalhus, J.L.; Dugan, M.E.R. Inclusion of Flaxseed in Hay- and Barley Silage Diets Increases Alpha-Linolenic Acid in Cow Plasma Independent of Forage Type. Lipids 2011, 46, 577–585. [Google Scholar] [CrossRef]
- Palmquist, D.L.; Jenkins, T.C. Fat in Lactation Rations: Review. J. Dairy Sci. 1980, 63, 1–14. [Google Scholar] [CrossRef]
- Plourde, M.; Cunnane, S.C. Extremely limited synthesis of long chain polyunsaturates in adults: Implications for their dietary essentiality and use as supplements. Appl. Physiol. Nutr. Metab. 2007, 32, 629–639. [Google Scholar] [CrossRef] [Green Version]
- Brossillon, V.; Reis, S.F.; Moura, D.C.; Galvão, J.G.B.; Oliveira, A.S.; Côrtes, C.; Brito, A.F. Production, milk and plasma fatty acid profile, and nutrient utilization in Jersey cows fed flaxseed oil and corn grain with different particle size. J. Dairy Sci. 2018, 101, 2127–2143. [Google Scholar] [CrossRef]
- Correddu, F.; Gaspa, G.; Pulina, G.; Nudda, A. Grape seed and linseed, alone and in combination, enhance unsaturated fatty acids in the milk of Sarda dairy sheep. J. Dairy Sci. 2016, 99, 1725–1735. [Google Scholar] [CrossRef]
- Ward, A.T.; Wittenberg, K.M.; Przybylski, R. Bovine milk fatty acid profiles produced by feeding diets containing solin, flax and canola. J. Dairy Sci. 2002, 85, 1191–1196. [Google Scholar] [CrossRef]
- Akraim, F.; Nicot, M.C.; Juaneda, P.; Enjalbert, F. Conjugated linolenic acid (CLnA), conjugated linoleic acid (CLA) and other biohydrogenation intermediates in plasma and milk fat of cows fed raw or extruded linseed. Animal 2007, 1, 835–843. [Google Scholar] [CrossRef] [Green Version]
- Moallem, U.; Lehrer, H.; Livshits, L.; Zachut, M. The effects of omega-3 α-linolenic acid from flaxseed oil supplemented to high-yielding dairy cows on production, health, and fertility. Livest. Sci. 2020, 242, 104302. [Google Scholar] [CrossRef]
- Lourenco, M.; Ramos-Morales, E.; Wallace, R.J. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Animal 2010, 4, 1008–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santillo, A.; Caroprese, M.; Marino, R.; D’Angelo, F.; Sevi, A.; Albenzio, M. Fatty acid profile of milk and Cacioricotta cheese from Italian Simmental cows as affected by dietary flaxseed supplementation. J. Dairy Sci. 2016, 99, 2545–2551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.X.; Zhang, Y.D.; Xu, Q.B.; Zheng, N.; Zhao, S.G.; Liu, K.J.; Qu, X.Y.; Jing, Y.; Wang, J.Q. DHA content in milk and biohydrogenation pathway in rumen: A review. PeerJ 2020, 8, e10230. [Google Scholar] [CrossRef]
- Sprecher, H. Metabolism of highly unsaturated n-3 and n-6 fatty acids. BBA-Mol. Cell Biol. Lipids 2000, 1486, 219–231. [Google Scholar] [CrossRef]
- Fleming, J.A.; Kris-Etherton, P.M. The evidence for α-linolenic acid and cardiovascular disease benefits: Comparisons with eicosapentaenoic acid and docosahexaenoic acid. Adv. Nutr. 2014, 6, 863S–876S. [Google Scholar] [CrossRef] [Green Version]
- Prisacaru, A.E. Effect of antioxidants on polyunsaturated fatty acids-review. Acta Sci. Pol. Technol. Aliment. 2016, 2, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Mavrommatis, A.; Chronopoulou, E.G.; Sotirakoglou, K.; Labrou, N.E.; Zervas, G.P.; Tsiplakou, E. The impact of the dietary supplementation level with schizochytrium sp, on the oxidative capacity of both goats’ organism and milk. Livest. Sci. 2018, 218, 37–43. [Google Scholar] [CrossRef]
- Prescott, S.L.; Dunstan, J.A. Prenatal fatty acid status and immune development: The pathways and the evidence. Lipids 2007, 42, 801–810. [Google Scholar] [CrossRef]
- Jorjong, S.; van Knegsel, A.T.; Verwaeren, J.; Lahoz, M.V.; Bruckmaier, R.M.; De Baets, B.; Kemp, B.; Fievez, V. Milk fatty acids as possible biomarkers to early diagnose elevated concentrations of blood plasma nonesterified fatty acids in dairy cows. J. Dairy Sci. 2014, 97, 7054–7064. [Google Scholar] [CrossRef] [Green Version]
- Jorjong, S.; van Knegsel, A.T.; Verwaeren, J.; Bruckmaier, R.M.; De Baets, B.; Kemp, B.; Fievez, V. Milk fatty acids as possible biomarkers to diagnose hyperketonemia in early lactation. J. Dairy Sci. 2015, 98, 5211–5221. [Google Scholar] [CrossRef] [Green Version]
- Ference, B.A.; Ginsberg, H.N.; Graham, I.; Ray, K.K.; Packard, C.J.; Bruckert, E.; Hegele, R.A.; Krauss, R.M.; Raal, F.J.; Schunkert, H.; et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease. 1. Evidence from genetic, epidemiologic, and clinical studies. A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2017, 32, 2459–2472. [Google Scholar] [CrossRef] [Green Version]
- Komprda, T.; Rozíková, V.; Zamazalová, N.; Škultéty, O.; Vícenová, M.; Trčková, M.; Faldyna, M. Effect of dietary fish oil on fatty acid deposition and expression of cholesterol homeostasis controlling genes in the liver and plasma lipid profile: Comparison of two animal models. J. Anim. Physiol. Anim. Nutr. 2016, 101, 1093–1102. [Google Scholar] [CrossRef]
- Pi, Y.; Gao, S.T.; Ma, L.; Zhu, Y.X.; Wang, J.Q.; Zhang, J.M.; Xu, J.C.; Bu, D.P. Effectiveness of rubber seed oil and flaxseed oil to enhance the α-linolenic acid content in milk from dairy cows. J. Dairy Sci. 2016, 99, 5719–5730. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Liu, P.; Wu, C.; Wang, T.; Liu, G.; Cao, H.; Zhang, C.; Hu, G.; Guo, X. Effects of fatty liver hemorrhagic syndrome on the AMP-activated protein kinase signaling pathway in laying hens. Poult. Sci. 2019, 98, 2201–2210. [Google Scholar] [CrossRef]
- Wu, X.L.; Zou, X.Y.; Zhang, M.; Hu, H.Q.; Jiang, S. Osteocalcin prevents insulin resistance, hepatic inflammation, and activates autophagy associated with high-fat diet-induced fatty liver hemorrhagic syndrome in aged laying hens. Poult. Sci. 2020, 100, 73–83. [Google Scholar] [CrossRef]
- Xia, S.L.; Li, X.F.; Abasubong, K.P.; Xu, C.; Shi, H.J.; Liu, W.B.; Zhang, D.D. Effects of dietary glucose and starch levels on the growth, apparent digestibility, and skin-associated mucosal non-specific immune parameters in juvenile blunt snout bream (Megalobrama amblycephala). Fish Shellfish Immunol. 2018, 79, 193–201. [Google Scholar] [CrossRef]
- Jha, A.K.; Pal, A.K.; Sahu, N.P.; Kumar, S.; Mukherjee, S.C. Haemato-immunological responses to dietary yeast RNA, ω-3 fatty acid and β-carotene in Catla catla juveniles. Fish Shellfish Immunol. 2007, 23, 917–927. [Google Scholar] [CrossRef]


{kind=link}
| Item | Treatments | ||
|---|---|---|---|
| CK | WF | GF | |
| Total FA, % diet DM | 4.59 | 8.52 | 8.50 |
| FA 1, % of total FA reported | |||
| C14:0 | 2.04 | 1.43 | 1.46 |
| C16:0 | 76.05 | 67.02 | 68.04 |
| C16:1 cis-9 | 0.03 | 0.03 | 0.04 |
| C18:0 | 0.03 | 0.04 | 0.05 |
| C18:1 cis-9 | 0.77 | 0.94 | 1.01 |
| C18:2 cis-9,cis-12 (LA) | 18.86 | 9.84 | 9.49 |
| C18:3 cis-9,cis-12,cis-15 (ALA) | 2.10 | 20.60 | 19.80 |
| C20:0 | 0.04 | 0.03 | 0.04 |
| C20:1 cis-11 | 0.01 | 0.01 | 0.01 |
| C22:0 | 0.04 | 0.03 | 0.04 |
| C24:0 | 0.03 | 0.02 | 0.03 |
| Item | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| CK | WF | GF | |||
| DMI, kg/d | 22.47 | 21.89 | 22.36 | 0.330 | 0.756 |
| Milk yield, kg/d | 36.05 | 36.39 | 38.09 | 0.486 | 0.122 |
| 4% FCM 1, kg/d | 36.78 | 37.05 | 37.96 | 0.537 | 0.658 |
| ECM 2, kg/d | 39.14 | 39.96 | 40.71 | 0.551 | 0.524 |
| Efficiency, ECM/DMI | 1.61 | 1.68 | 1.73 | 0.026 | 0.187 |
| Milk composition, % | |||||
| Milk fat | 4.15 | 4.12 | 3.95 | 0.082 | 0.583 |
| Milk Protein | 3.10 b | 3.31 a | 3.14 a,b | 0.035 | 0.037 |
| Milk lactose | 4.84 | 4.78 | 4.81 | 0.020 | 0.566 |
| Nonfat milk solids | 8.43 | 8.65 | 8.53 | 0.038 | 0.060 |
| Total milk solids | 12.58 | 12.77 | 12.48 | 0.092 | 0.451 |
| Milk production, kg/d | |||||
| Milk fat yield | 1.49 | 1.50 | 1.51 | 0.030 | 0.971 |
| Milk protein yield | 1.12 | 1.20 | 1.20 | 0.017 | 0.059 |
| Milk lactose yield | 1.74 | 1.74 | 1.84 | 0.025 | 0.168 |
| Nonfat milk solids yield | 3.04 | 3.15 | 3.27 | 0.043 | 0.087 |
| Total milk solids yield | 4.53 | 4.65 | 1.78 | 0.059 | 0.238 |
| Fatty Acid, % of Total | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| CK | WF | GF | |||
| C4:0 | 0.051 | 0.051 | 0.048 | 0.002 | 0.697 |
| C6:0 | 0.021 | 0.023 | 0.022 | 0.001 | 0.722 |
| C8:0 | 0.019 | 0.02 | 0.02 | 0.001 | 0.737 |
| C10:0 | 0.035 | 0.041 | 0.039 | 0.002 | 0.439 |
| C12:0 | 0.084 | 0.087 | 0.082 | 0.003 | 0.813 |
| C14:0 | 0.56 a,b | 0.63 a | 0.52 b | 0.020 | 0.064 |
| C14:1 cis-9 | 0.054 | 0.061 | 0.052 | 0.002 | 0.189 |
| C16:0 | 10.16 a | 9.25b | 9.86 a | 0.136 | 0.015 |
| C16:1 trans-7 | 0.43 a | 0.36b | 0.27c | 0.014 | <0.001 |
| C16:1 cis-9 | 0.66 | 0.76 | 0.65 | 0.023 | 0.116 |
| C18:0 | 12.33 a | 10.90 b | 12.17 a | 0.234 | 0.017 |
| C18:1 cis-6 | 0.053 a | 0.048 a,b | 0.045 b | 0.001 | 0.037 |
| C18:1 cis-9 | 4.7 | 4.88 | 4.48 | 0.106 | 0.318 |
| C18:1 trans-9 | 0.36 | 0.34 | 0.27 | 0.019 | 0.105 |
| C18:1 trans-6 + trans-11 | 0.62 b | 0.59 b | 0.77 a | 0.022 | <0.001 |
| other C18:1 1 | 0.83 b | 0.81 b | 1.68 a | 0.077 | <0.001 |
| C18:2 cis-9,cis-12 (LA) | 50.41 a,b | 51.82 a | 49.14 b | 0.360 | 0.006 |
| C18:2 trans-9,trans-12 | 0.10 b | 0.09 b | 0.11 a | 0.002 | <0.001 |
| C18:2 cis-9,trans-11 (CLA) | 0.20 | 0.20 | 0.21 | 0.005 | 0.483 |
| C18:2 trans-10,cis-12 (CLA) | 0.09 | 0.08 | 0.08 | 0.002 | 0.415 |
| C18:3 cis-6,cis-9,cis-12 (GLA) | 1.16 a | 0.87 b | 0.54 c | 0.058 | <0.001 |
| C18:3 cis-9,cis-12,cis-15 (ALA) | 2.40 c | 5.30 b | 6.59 a | 0.341 | <0.001 |
| C20:0 | 0.12 | 0.10 | 0.10 | 0.002 | 0.080 |
| C20:1 cis-9 | 0.05 a | 0.04 b | 0.04 b | 0.001 | 0.006 |
| C20:1 cis-11 | 0.13 a | 0.09 b | 0.10 b | 0.005 | <0.001 |
| C20:2 cis-11,cis-14 (EDA) | 0.099 a | 0.086 b | 0.0910 a,b | 0.002 | 0.017 |
| C20:3 cis-8,cis-11,cis-14 (DGLA) | 2.51 a | 1.61 b | 1.47 b | 0.103 | <0.001 |
| C20:3 cis-11,cis-14,cis-17 (ETE) | 0.00 b | 0.05 a | 0.05 a | 0.004 | <0.001 |
| C20:4 cis-5,cis-8,cis-11,cis-14 (AA) | 1.79 a | 1.71 a | 1.43 b | 0.058 | 0.022 |
| C20:5 cis-5,cis-8,cis-11,cis-14,cis-17 (EPA) | 0.31 b | 0.50 a | 0.56 a | 0.027 | <0.001 |
| C22:0 | 0.20 a | 0.17 b | 0.19 a | 0.004 | 0.007 |
| C22:1 cis-13 | 6.49 | 5.75 | 5.51 | 0.200 | 0.112 |
| C22:2 cis-13,cis-16 (DDA) | 0.20 a | 0.18 b | 0.19 a,b | 0.005 | 0.078 |
| C22:6 cis-4,cis-7,cis-10,cis-13,cis-16,cis-19 (DHA) | 0.06 | 0.06 | 0.07 | 0.002 | 0.219 |
| C24:0 | 0.25 | 0.23 | 0.25 | 0.005 | 0.135 |
| C24:1cis-15 | 0.19 a | 0.16 b | 0.17 a,b | 0.004 | 0.075 |
| Total n-6 PUFA 2 | 56.17 a | 56.27 a | 52.87 b | 0.409 | <0.001 |
| Total n-3 PUFA 3 | 2.76 c | 5.90 b | 7.26 a | 0.366 | <0.001 |
| n-6/n-3 4 | 20.39 a | 9.67 b | 7.35 c | 1.075 | <0.001 |
| SFA | 23.83 a | 21.49 b | 23.29 a | 0.271 | <0.001 |
| MUFA | 14.70 | 14.23 | 13.96 | 0.234 | 0.440 |
| PUFA | 59.32 b | 62.54 a | 60.53 b | 0.380 | 0.001 |
| Fatty Acid, % of Total | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| CK | WF | GF | |||
| C4:0 | 3.23 a | 2.84 a,b | 2.67 b | 0.095 | 0.042 |
| C6:0 | 2.40 a | 2.00 b | 1.91 b | 0.069 | 0.005 |
| C8:0 | 1.50 a | 1.23 b | 1.20 b | 0.045 | 0.007 |
| C10:0 | 3.08 a | 2.52 b | 2.47 b | 0.095 | 0.009 |
| C10:1 cis-9 | 0.65 a | 0.53 b | 0.45 b | 0.024 | 0.001 |
| C12:0 | 3.18 a | 2.58 b | 2.69 b | 0.091 | 0.012 |
| C12:1 cis-9 | 0.09 a | 0.08 b | 0.07 b | 0.003 | 0.005 |
| C14:0 | 9.85 | 9.00 | 9.42 | 0.168 | 0.119 |
| C14:1 cis-9 | 0.89 a | 0.74 b | 0.72 b | 0.030 | 0.042 |
| C16:0 | 35.07 a | 31.15 b | 30.55 b | 0.468 | <0.001 |
| C16:1 trans-7 | 0.13 b | 0.15 a | 0.12 b | 0.003 | 0.010 |
| C16:1 cis-9 | 1.36 | 1.40 | 1.37 | 0.042 | 0.919 |
| C18:0 | 10.15 b | 11.58 a | 11.27 a | 0.221 | 0.016 |
| C18:1 cis-6 | 0.05 b | 0.07 a | 0.06 a,b | 0.003 | 0.070 |
| C18:1 cis-9 | 18.36 b | 22.42 a | 19.67 b | 0.506 | 0.001 |
| C18:1 trans-9 | 0.26 c | 0.32 b | 0.55 a | 0.024 | <0.001 |
| C18:1 trans-6 + trans-11 | 1.19 c | 1.53 b | 2.28 a | 0.101 | <0.001 |
| Other C18:1 1 | 1.49 c | 2.21 b | 4.07 a | 0.206 | <0.001 |
| C18:2 cis-9,cis-12 (LA) | 2.62 b | 2.80 a,b | 3.02 a | 0.060 | 0.020 |
| C18:2 trans-9,trans-12 | 0.12 c | 0.18 b | 0.26 a | 0.011 | <0.001 |
| C18:2 cis-9,trans-11 (CLA) | 0.46 c | 0.62 b | 0.83 a | 0.036 | <0.001 |
| C18:2 trans-10,cis-12 (CLA) | 0.04 | 0.05 | 0.05 | 0.002 | 0.142 |
| C18:3 cis-6,cis-9,cis-12 (GLA) | 0.05 | 0.05 | 0.05 | 0.001 | 0.518 |
| C18:3 cis-9,cis-12,cis-15 (ALA) | 0.26 c | 0.58 b | 0.91 a | 0.052 | <0.001 |
| C20:0 | 0.10 b | 0.12 a | 0.11 a,b | 0.002 | 0.007 |
| C20:1 cis-9 | 0.08 b | 0.09 a | 0.08 b | 0.002 | 0.019 |
| C20:1 cis-11 | 0.033 b | 0.041 a | 0.042 a | 0.001 | 0.006 |
| C20:2 cis-11,cis-14 (EDA) | 0.035 b | 0.043 a | 0.044 a | 0.001 | 0.014 |
| C20:3 cis-8,cis-11,cis-14 (DGLA) | 0.012 a | 0.010 a,b | 0.009 b | 0.004 | 0.005 |
| C20:3 cis-11,cis-14,cis-17 (ETE) | 0.00 b | 0.03 a | 0.03 a | 0.002 | <0.001 |
| C20:4 cis-5,cis-8,cis-11,cis-14 (AA) | 0.14 a | 0.13 a | 0.11 b | 0.003 | <0.001 |
| C20:5 cis-5,cis-8,cis-11,cis-14,cis-17 (EPA) | 0.03 b | 0.05 a | 0.06 a | 0.002 | <0.001 |
| C22:0 | 0.05 | 0.06 | 0.06 | 0.002 | 0.164 |
| C22:1 cis-13 | 0.08 b | 0.09 a,b | 0.10 a | 0.005 | 0.094 |
| C22:2 cis-13,cis-16 (DDA) | 0.03 | 0.03 | 0.03 | 0.001 | 0.278 |
| C22:6 cis-4,cis-7,cis-10,cis-13,cis-16,cis-19 (DHA) | 0.007 b | 0.009 a,b | 0.017 a | 0.002 | 0.037 |
| C24:0 | 0.05 | 0.06 | 0.06 | 0.002 | 0.383 |
| C24:1cis-15 | 0.02 | 0.02 | 0.02 | 0.001 | 0.357 |
| Total n-6 PUFA 2 | 2.98 b | 3.16 a | 3.34 a | 0.059 | 0.042 |
| Total n-3 PUFA 3 | 0.30 c | 0.67 b | 1.01 a | 0.057 | <0.001 |
| n-6/n-3 4 | 10.07 a | 4.77 b | 3.30 c | 0.545 | <0.001 |
| SFA | 68.66 a | 63.08 b | 62.45 b | 0.685 | <0.001 |
| MUFA | 23.31 b | 27.61 a | 25.66 a | 0.540 | 0.002 |
| PUFA | 3.90 c | 4.67 b | 5.49 a | 0.141 | <0.001 |
| D9- desaturase indices | |||||
| C10:1 | 17.40 a | 17.44 a | 15.15 b | 0.376 | 0.012 |
| C12:1 | 2.74 | 2.84 | 2.55 | 0.059 | 0.131 |
| C14:1 | 8.35 | 7.61 | 7.10 | 0.297 | 0.230 |
| C16:1 | 3.72 | 4.28 | 4.32 | 0.137 | 0.131 |
| C18:1 | 64.34 | 65.86 | 63.51 | 0.552 | 0.218 |
| C20:1 | 44.28 | 43.53 | 42.33 | 0.459 | 0.223 |
| Items | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| CK | WF | GF | |||
| Milk ALA yield, g/d | 3.75 c | 8.71 b | 13.51 a | 0.791 | <0.001 |
| Milk ETE yield, g/d | 0.00 b | 0.38 a | 0.38 a | 0.035 | <0.001 |
| Milk EPA yield, g/d | 0.49 b | 0.77 a | 0.82 a | 0.029 | <0.001 |
| Milk DHA yield, g/d | 0.09 b | 0.14 b | 0.25 a | 0.025 | 0.021 |
| n-3 PUFA 1 yield, g/d | 4.34 c | 10.00 b | 14.96 a | 0.855 | <0.001 |
| Milk ALA/ALA intake, % | 17.30 a | 2.29 c | 3.61 b | 1.277 | <0.001 |
| Item | Antioxidant Indexes | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|---|
| CK | WF | GF | ||||
| Plasma | GSH-Px (ng/L) | 40.99 | 42.95 | 42.99 | 0.606 | 0.823 |
| MDA (nmol/mL) | 16.76 | 16.94 | 17.27 | 1.467 | 0.946 | |
| CAT (ng/L) | 35.63 b | 47.25 a | 48.08 a | 1.607 | 0.000 | |
| T-AOC (nmol/mL) | 1.49 a | 0.99 b | 1.28 a | 0.064 | 0.002 | |
| Milk | GSH-Px (ng/L) | 273.31 c | 501.27 a | 433.47 b | 23.340 | 0.000 |
| MDA (nmol/mL) | 6.51 a | 2.19 c | 3.81 b | 0.417 | 0.000 | |
| CAT (ng/L) | 5.82 | 7.03 | 5.09 | 0.633 | 0.465 | |
| T-AOC (nmol/mL) | 0.27 | 0.25 | 0.24 | 0.016 | 0.788 | |
| Item | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| CK | WF | GF | |||
| TG, mmol/L | 1.17 | 1.00 | 1.07 | 0.071 | 0.615 |
| LDLC, mmol/L | 1.77 | 1.39 | 2.09 | 0.186 | 0.317 |
| VLDLC, mmol/L | 0.53 | 0.53 | 0.48 | 0.027 | 0.677 |
| HDLC, mmol/L | 1.43 | 1.48 | 1.30 | 0.053 | 0.379 |
| TC, mmol/L | 4.29 | 4.14 | 4.64 | 0.144 | 0.359 |
| AST, U/L | 17.81 | 16.13 | 14.4 | 1.688 | 0.726 |
| ALT, U/L | 20.08 | 20.74 | 25.87 | 1.979 | 0.440 |
| ALP, U/L | 97.03 | 109.96 | 86.57 | 5.889 | 0.310 |
| TP, g/L | 71.05 | 73.79 | 75.10 | 1.368 | 0.726 |
| ALB, g/L | 46.31 | 44.58 | 41.34 | 1.135 | 0.196 |
| GLB, g/L | 24.74 | 29.21 | 30.76 | 1.492 | 0.238 |
| BUN, mmol/L | 5.33 | 6.07 | 5.55 | 0.247 | 0.471 |
| GLU, mmol/L | 1.35 | 0.99 | 1.07 | 0.113 | 0.413 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, G.; Wang, J.; Liu, K.; Wang, F.; Zheng, N.; Zhao, S.; Qu, X.; Yu, J.; Zhang, Y.; Wang, J. Effect of Flaxseed Supplementation on Milk and Plasma Fatty Acid Composition and Plasma Parameters of Holstein Dairy Cows. Animals 2022, 12, 1898. https://doi.org/10.3390/ani12151898
Huang G, Wang J, Liu K, Wang F, Zheng N, Zhao S, Qu X, Yu J, Zhang Y, Wang J. Effect of Flaxseed Supplementation on Milk and Plasma Fatty Acid Composition and Plasma Parameters of Holstein Dairy Cows. Animals. 2022; 12(15):1898. https://doi.org/10.3390/ani12151898
Chicago/Turabian StyleHuang, Guoxin, Jie Wang, Kaizhen Liu, Fengen Wang, Nan Zheng, Shengguo Zhao, Xueyin Qu, Jing Yu, Yangdong Zhang, and Jiaqi Wang. 2022. "Effect of Flaxseed Supplementation on Milk and Plasma Fatty Acid Composition and Plasma Parameters of Holstein Dairy Cows" Animals 12, no. 15: 1898. https://doi.org/10.3390/ani12151898