
Fresh Food Consumption Increases Microbiome Diversity and Promotes Changes in Bacteria Composition on the Skin of Pet Dogs Compared to Dry Foods

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Dog Recruitment and General Study Guidelines
2.2. Feeding Protocol and Diet Nutrient and Ingredient Compositions
2.3. Skin Microbiome Sample Collection
2.4. DNA Extraction and Analysis
2.5. Data Processing and Taxonomic Composition
2.6. Statistical Methods
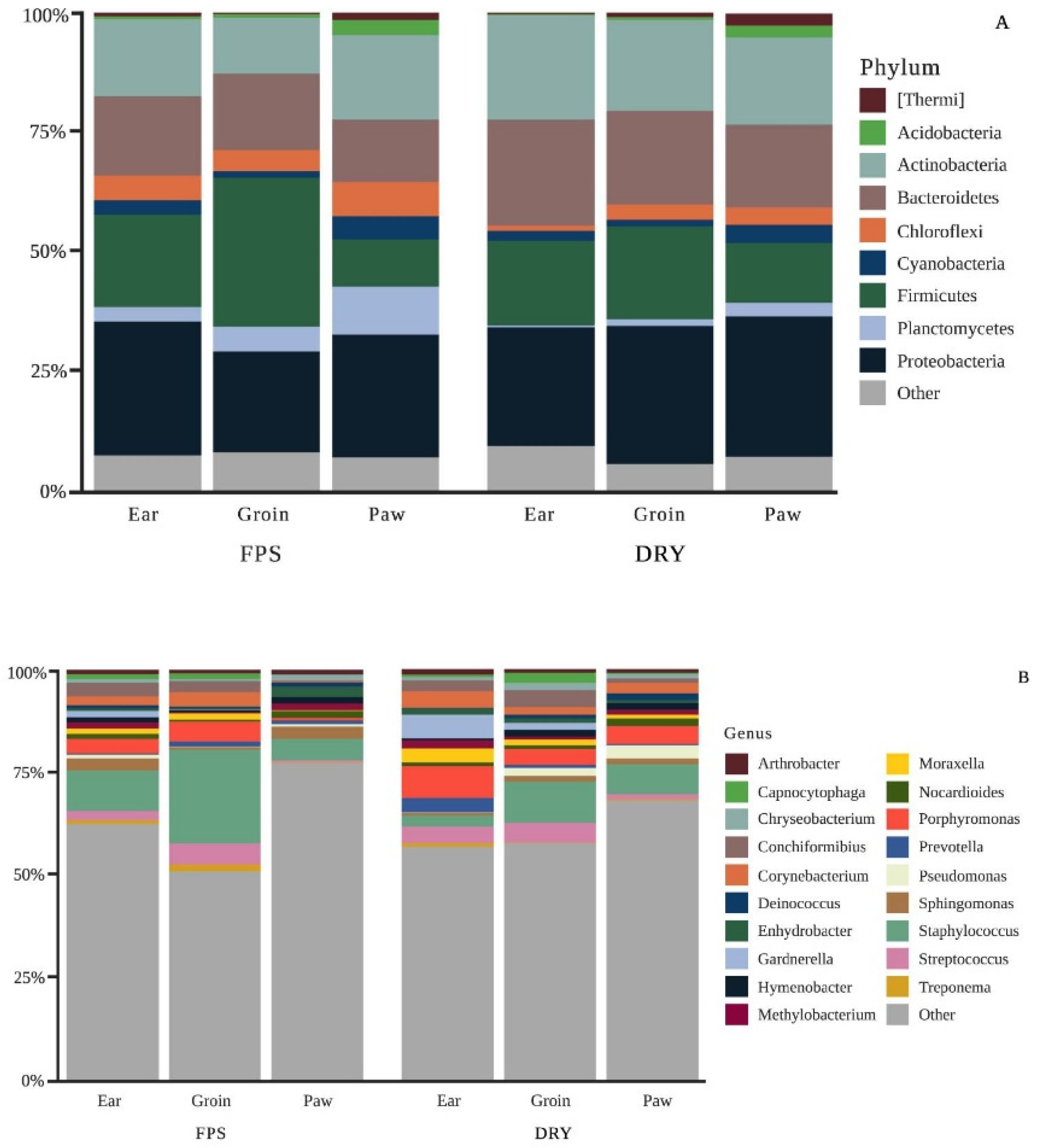
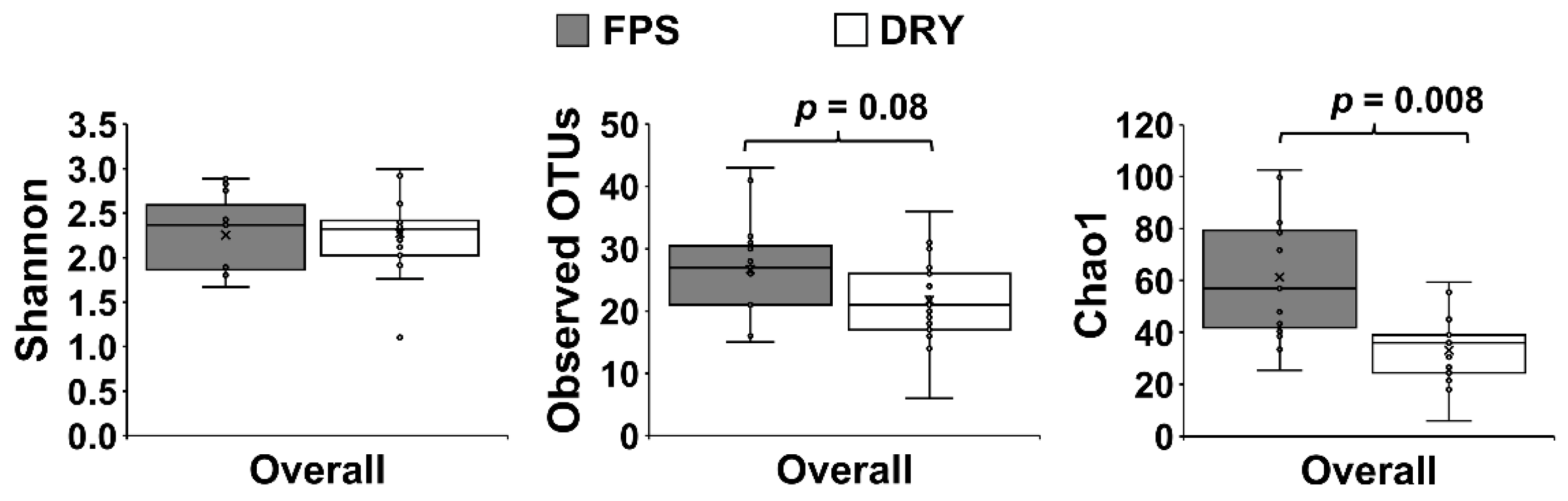
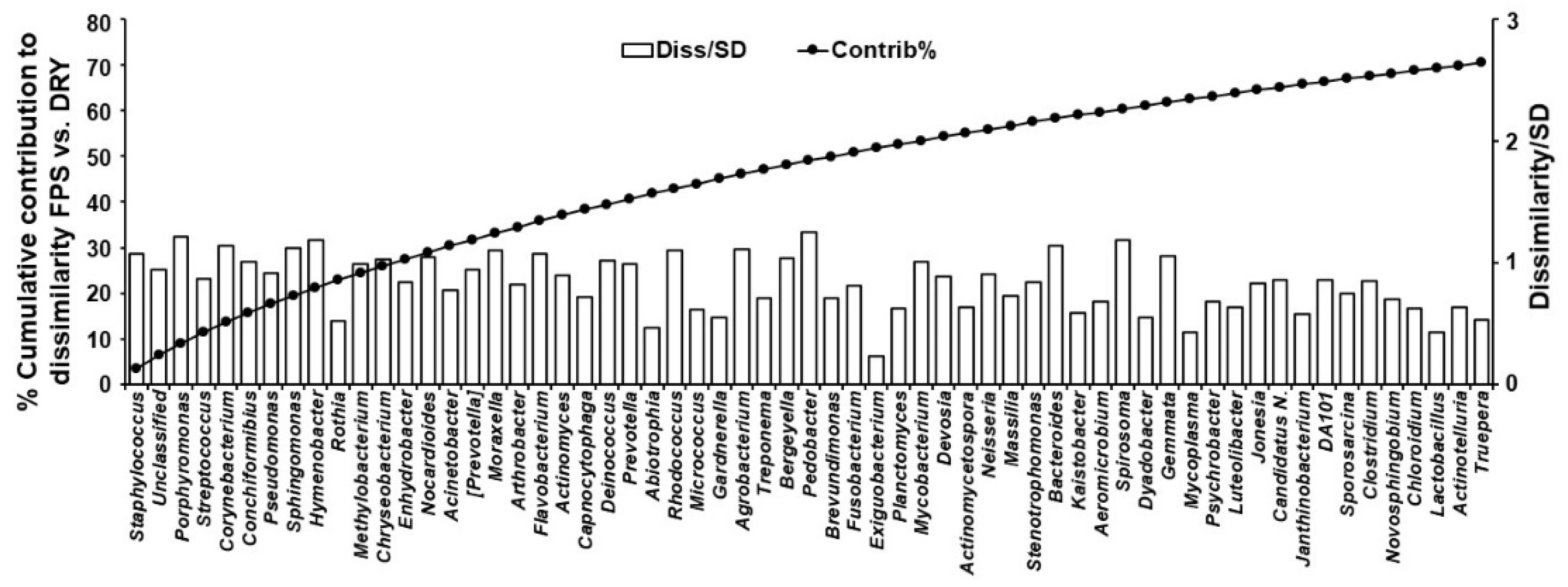
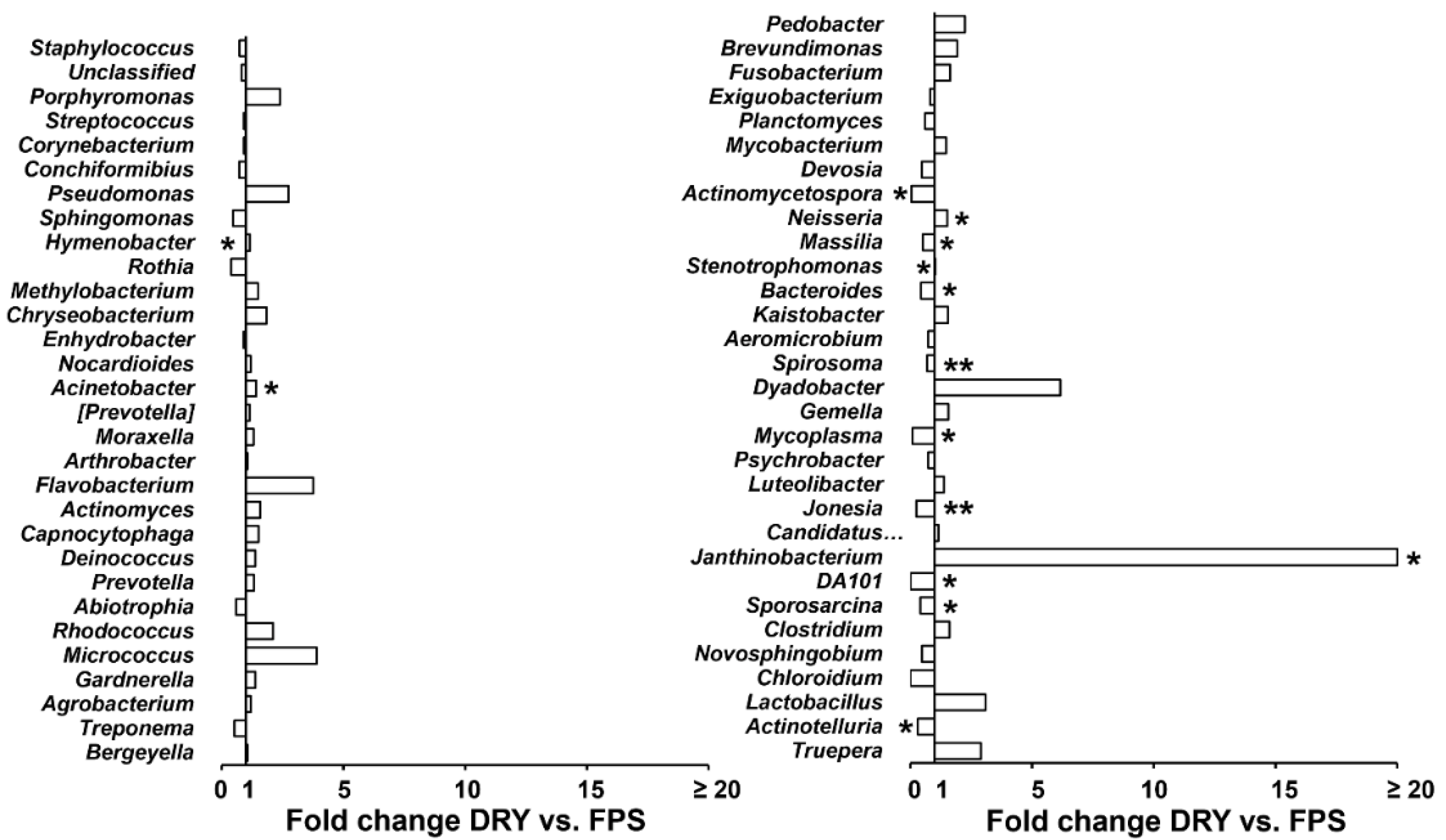
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ihrke, P.J. An overview of bacterial skin disease in the dog. Br. Vet. J. 1987, 143, 112–118. [Google Scholar] [CrossRef]
- Stewart, L.J. Newly Reported Skin Disease Syndromes in the Dog. Vet. Clin. N. Am. Small Anim. Pract. 1990, 20, 1603–1613. [Google Scholar] [CrossRef]
- Gross, T.L.; Ihrke, P.J.; Walder, E.J.; Affolter, V.K. Skin Diseases of the Dog and Cat; Blackwell Science: Oxford, UK, 2005. [Google Scholar] [CrossRef]
- Hoffmann, A.R.; Patterson, A.P.; Diesel, A.; Lawhon, S.D.; Ly, H.J.; Stephenson, C.E.; Mansell, J.; Steiner, J.M.; Dowd, S.E.; Olivry, T.; et al. The Skin Microbiome in Healthy and Allergic Dogs. PLoS ONE 2014, 9, e83197. [Google Scholar] [CrossRef]
- Bradley, C.W.; Morris, D.O.; Rankin, S.C.; Cain, C.L.; Misic, A.M.; Houser, T.; Mauldin, E.A.; Grice, E.A. Longitudinal Evaluation of the Skin Microbiome and Association with Microenvironment and Treatment in Canine Atopic Dermatitis. J. Investig. Dermatol. 2016, 136, 1182–1190. [Google Scholar] [CrossRef] [Green Version]
- Pierezan, F.; Olivry, T.; Paps, J.S.; Lawhon, S.D.; Wu, J.; Steiner, J.M.; Suchodolski, J.S.; Hoffmann, A.R. The skin microbiome in allergen-induced canine atopic dermatitis. Vet. Dermatol. 2016, 27, 332. [Google Scholar] [CrossRef]
- Pet Insurer Nationwide Reveals Annual Cost of Common Health Conditions. Available online: https://www.veterinarypracticenews.com/10-top-pet-health-conditions-cost-96-treat-2017/ (accessed on 15 March 2018).
- Chermprapai, S.; Ederveen, T.H.A.; Broere, F.; Broens, E.M.; Schlotter, Y.M.; van Schalkwijk, S.; Boekhorst, J.; van Hijum, S.A.F.T.; Rutten, V.P.M.G. The bacterial and fungal microbiome of the skin of healthy dogs and dogs with atopic dermatitis and the impact of topical antimicrobial therapy, an exploratory study. Vet. Microbiol. 2019, 229, 90–99. [Google Scholar] [CrossRef]
- DeCandia, A.L.; Leverett, K.N.; von Holdt, B.M. Of microbes and mange: Consistent changes in the skin microbiome of three canid species infected with Sarcoptes scabiei mites. Parasites Vectors 2019, 12, 488. [Google Scholar] [CrossRef]
- Marsh, K.A.; Ruedisueli, F.L.; Coe, S.L.; Watson, T.G.D. Effects of zinc and linoleic acid supplementation on the skin and coat quality of dogs receiving a complete and balanced diet. Vet. Dermatol. 2000, 11, 277–284. [Google Scholar] [CrossRef]
- Kirby, N.A.; Hester, S.L.; Bauer, J.E. Dietary fats and the skin and coat of dogs. J. Am. Vet. Med. Assoc. 2007, 230, 1641–1644. [Google Scholar] [CrossRef]
- Kirby, N.A.; Hester, S.L.; Rees, C.A.; Kennis, R.A.; Zoran, D.L.; Bauer, J.E. Skin surface lipids and skin and hair coat condition in dogs fed increased total fat diets containing polyunsaturated fatty acids. J. Anim. Physiol. Anim. Nutr. 2009, 93, 505–511. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Bosko, C.A. Skin Barrier Insights: From Bricks and Mortar to Molecules and Microbes. J. Drugs Dermatol. 2019, 18, s63–s67. [Google Scholar]
- Logas, D.; Kunkle, G.A. Double-blinded Crossover Study with Marine Oil Supplementation Containing High-dose eicosapentaenoic Acid for the Treatment of Canine Pruritic Skin Disease. Vet. Dermatol. 1994, 5, 99–104. [Google Scholar] [CrossRef]
- Nesbitt, G.H.; Freeman, L.M.; Hannah, S.S. Effect of n-3 fatty acid ratio and dose on clinical manifestations, plasma fatty acids and inflammatory mediators in dogs with pruritus. Vet. Dermatol. 2003, 14, 67–74. [Google Scholar] [CrossRef]
- Schmidt, M.; Unterer, S.; Suchodolski, J.S.; Honneffer, J.B.; Guard, B.C.; Lidbury, J.A.; Steiner, J.M.; Fritz, J.; Kolle, P. The fecal microbiome and metabolome differs between dogs fed Bones and Raw Food (BARF) diets and dogs fed commercial diets. PLoS ONE 2018, 13, e0201279. [Google Scholar] [CrossRef] [Green Version]
- Ephraim, E.; Cochrane, C.-Y.; Jewell, D.E. Varying Protein Levels Influence Metabolomics and the Gut Microbiome in Healthy Adult Dogs. Toxins 2020, 12, 517. [Google Scholar] [CrossRef]
- Wernimont, S.M.; Radosevich, J.; Jackson, M.I.; Ephraim, E.; Badri, D.V.; MacLeay, J.M.; Jewell, D.E.; Suchodolski, J.S. The Effects of Nutrition on the Gastrointestinal Microbiome of Cats and Dogs: Impact on Health and Disease. Front. Microbiol. 2020, 11, 1266. [Google Scholar] [CrossRef]
- Skowron, K.; Bauza-Kaszewska, J.; Kraszewska, Z.; Wiktorczyk-Kapischke, N.; Grudlewska-Buda, K.; Kwiecińska-Piróg, J.; Walecka-Zacharska, E.; Radtka, L.; Gospodarek-Komkowska, E. Human Skin Microbiome: Impact of Intrinsic and Extrinsic Factors on Skin Microbiota. Microorganisms 2021, 9, 543. [Google Scholar] [CrossRef]
- Watson, T.D.G. Diet and Skin Disease in Dogs and Cats. J. Nutr. 1998, 128, 2783S–2789S. [Google Scholar] [CrossRef] [Green Version]
- Association of American Feed Control Officials. 2021 Official Publication; AAFCO: Champaign, IL, USA, 2021. [Google Scholar]
- Oba, P.M.; Utterback, P.L.; Parsons, C.M.; de Godoy, M.R.C.; Swanson, K.S. Chemical composition, true nutrient digestibility, and true metabolizable energy of chicken-based ingredients differing by processing method using the precision-fed cecectomized rooster assay1. J. Anim. Sci. 2019, 97, 998–1009. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Manjarin, R.; Maj, M.A.; La Frano, M.R.; Glanz, H. %polynova_2way: A SAS macro for implementation of mixed models for metabolomics data. PLoS ONE 2020, 15, e0244013. [Google Scholar] [CrossRef]
- Isaiah, A.; Hoffmann, A.R.; Kelley, R.; Mundell, P.; Steiner, J.M.; Suchodolski, J.S. Characterization of the nasal and oral microbiota of detection dogs. PLoS ONE 2017, 12, e0184899. [Google Scholar] [CrossRef]
- Elias, P.M. Epidermal lipids, barrier function, and desquamation. J. Investig. Dermatol. 1983, 80, 44s–49s. [Google Scholar] [CrossRef]
- Cusco, A.; Belanger, J.M.; Gershony, L.; Islas-Trejo, A.; Levy, K.; Medrano, J.F.; Sanchez, A.; Oberbauer, A.M.; Francino, O. Individual signatures and environmental factors shape skin microbiota in healthy dogs. Microbiome 2017, 5, 139. [Google Scholar] [CrossRef] [Green Version]
- Prescott, S.L.; Larcombe, D.-L.; Logan, A.C.; West, C.; Burks, W.; Caraballo, L.; Levin, M.; van Etten, E.; Horwitz, P.; Kozyrskyj, A.; et al. The skin microbiome: Impact of modern environments on skin ecology, barrier integrity, and systemic immune programming. World Allergy Organ. J. 2017, 10, 29. [Google Scholar] [CrossRef]
- Fazakerley, J.; Nuttall, T.; Sales, D.; Schmidt, V.; Carter, S.D.; Hart, C.A.; McEwan, N.A. Staphylococcal colonization of mucosal and lesional skin sites in atopic and healthy dogs. Vet. Dermatol. 2009, 20, 179–184. [Google Scholar] [CrossRef]
- Iwase, T.; Uehara, Y.; Shinji, H.; Tajima, A.; Seo, H.; Takada, K.; Agata, T.; Mizunoe, Y. Staphylococcus epidermitis esp inhibits Staphylococcus aureus biofilm formation and nasal colonization. Nature 2010, 465, 346–349. [Google Scholar] [CrossRef]
- Lunjani, N.; Hlela, C.; O’Mahony, L. Microbiome and skin biology. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 328–333. [Google Scholar] [CrossRef]
- Lowe, J.A.; Wiseman, J.; Cole, D.J.A. Zinc source influences zinc retention in hair and hair growth in the dog. J. Nutr. 1994, 124, 2575S–2576S. [Google Scholar] [CrossRef]
- Trevisan, L.; Fischer, M.M.; Rodenbusch, C.R.; Labres, R.V.; Kessler, A.M. Effects of diets containing organic and inorganic zinc sources on hair characteristics, zinc concentration in blood and hair, and the immune response of dogs. Acta Sci. Vet. 2013, 41, 1154–1161. [Google Scholar]
- Swanson, K.S.; Grieshop, C.M.; Clapper, G.M.; Shields, R.G.; Belay, T.; Merchen, N.R.; Fahey, G.C., Jr. Fruit and vegetable fiber fermentation by gut microflora from canines. J. Anim. Sci. 2001, 79, 919–926. [Google Scholar] [CrossRef] [Green Version]
- Middelbos, I.S.; Vester Boler, B.M.; Qu, A.; White, B.A.; Swanson, K.S.; Fahey, G.C., Jr. Phylogenetic Characterization of Fecal Microbial Communities of Dogs Fed Diets with or without Supplemental Dietary Fiber Using 454 Pyrosequencing. PLoS ONE 2010, 5, e9768. [Google Scholar] [CrossRef] [Green Version]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.-J. Review article: The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef]
- Anderson, R.S. Water balance in the dog and cat. J. Small Anim. Pract. 1982, 23, 588–598. [Google Scholar] [CrossRef]
- Ramsay, D.J.; Thrasher, T.N. Regulation of fluid intake in dogs following water deprivation. Brain Res. Bull. 1991, 27, 495–499. [Google Scholar] [CrossRef]
- Mac-Mary, S.; Creidi, P.; Marsaut, D.; Courderot-Masuyer, C.; Cochet, V.; Gharbi, T.; Guidicelli-Arranz, D.; Tondu, F.; Humbert, P. Assessment of effects of an additional dietary natural mineral water uptake on skin hydration in healthy subjects by dynamic barrier function measurements and clinic scoring. Ski. Res. Technol. 2006, 12, 199–205. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mitra, R.; Maitra, A.; Gupta, S.; Kumaran, S.; Chakrabortty, A.; Majumder, P.P. Sebum and Hydration Levels in Specific Regions of Human Face Significantly Predict the Nature and Diversity of Facial Skin Microbiome. Sci. Rep. 2016, 6, 36062. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dog ID | Breed | Sex | Age (Years) |
|---|---|---|---|
| D010 | Mixed | Male | 1 |
| D011 | Goldendoodle | Male | 3 |
| D012 | Australian Cattle | Male | 8 |
| D013 | Mixed | Female | 2 |
| D014 | Mixed | Male | 0.8 |
| D015 | Mixed | Male | 5 |
| D016 | American Bully | Male | 5 |
| D017 | American Bully | Female | 4 |
| Nutrient | DRY 1 | DRY 2 | DRY 3 | DRY 4 | DRY 5 | DRY Average | FPS |
|---|---|---|---|---|---|---|---|
| Moisture | 7.9 | 9.9 | 7.6 | 10.4 | 6.4 | 8.5 | 76.3 |
| Crude Protein | 37.2 | 33.8 | 40 | 26.8 | 28.4 | 33.0 | 43.9 |
| Fat | 20.5 | 11.8 | 19.7 | 15.4 | 13.2 | 16.1 | 21.4 |
| Ash | 10.1 | 7.8 | 9.6 | 7.2 | 5.7 | 8.1 | 7.9 |
| Total Dietary Fiber | 12.2 | 10.9 | 12.0 | 8.0 | 14.2 | 11.5 | 11.8 |
| Insoluble Fiber | 9.8 | 8.9 | 9.4 | 6.6 | 12.1 | 9.4 | 6.3 |
| Soluble Fiber | 2.4 | 2.0 | 2.6 | 1.5 | 2.1 | 2.1 | 5.5 |
| Nitrogen-free Extract 1 | 19.4 | 35.8 | 19.8 | 42.5 | 38.5 | 31.2 | 17.7 |
| Metabolizable Energy 2, Dry Basis (Wet Basis) | 3724 (3429) | 3439 (3099) | 3768 (3481) | 3735 (3346) | 3464 (3242) | 3616 (3308) | 3975 (942) |
| Zinc | 265 | 289 | 232 | 208 | 222 | 243 | 215 |
| Nutrient | DRY 1 | DRY 2 | DRY 3 | DRY 4 | DRY 5 | DRY Average | FPS |
|---|---|---|---|---|---|---|---|
| Saturated fats | 6.58 | 2.71 | 5.78 | 6.63 | 3.29 | 5.00 | 6.82 |
| Monosaturated fats | 8.41 | 4.64 | 8.49 | 5.84 | 4.72 | 6.42 | 8.85 |
| Polyunsaturated fats | 3.94 | 3.18 | 3.87 | 1.28 | 4.22 | 3.30 | 5.21 |
| ω-6 fatty acids | 3.65 | 2.50 | 3.20 | 1.18 | 3.35 | 2.78 | 4.78 |
| ω-3 fatty acids | 0.26 | 0.67 | 0.63 | 0.10 | 0.85 | 0.50 | 0.39 |
| ω-6: ω-3 ratio 1 | 14.04 | 3.73 | 5.08 | 11.80 | 3.94 | 5.56 | 12.26 |
| Linoleic acid | 3.51 | 2.38 | 3.06 | 1.14 | 3.26 | 2.67 | 4.32 |
| α-Linolenic acid | 0.23 | 0.42 | 0.44 | 0.09 | 0.82 | 0.40 | 0.24 |
| Arachidonic acid | 0.08 | 0.06 | 0.08 | 0.02 | 0.04 | 0.06 | 0.35 |
| Eicosapentaenoic acid | nd 2 | 0.07 | 0.09 | nd | nd | 0.03 1 | 0.03 |
| Docosahexaenoic acid | nd | 0.14 | 0.08 | nd | nd | 0.04 1 | 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leverett, K.; Manjarín, R.; Laird, E.; Valtierra, D.; Santiago-Rodriguez, T.M.; Donadelli, R.; Perez-Camargo, G. Fresh Food Consumption Increases Microbiome Diversity and Promotes Changes in Bacteria Composition on the Skin of Pet Dogs Compared to Dry Foods. Animals 2022, 12, 1881. https://doi.org/10.3390/ani12151881
Leverett K, Manjarín R, Laird E, Valtierra D, Santiago-Rodriguez TM, Donadelli R, Perez-Camargo G. Fresh Food Consumption Increases Microbiome Diversity and Promotes Changes in Bacteria Composition on the Skin of Pet Dogs Compared to Dry Foods. Animals. 2022; 12(15):1881. https://doi.org/10.3390/ani12151881
Chicago/Turabian StyleLeverett, Kennedy, Rodrigo Manjarín, Erica Laird, Diana Valtierra, Tasha M. Santiago-Rodriguez, Renan Donadelli, and Gerardo Perez-Camargo. 2022. "Fresh Food Consumption Increases Microbiome Diversity and Promotes Changes in Bacteria Composition on the Skin of Pet Dogs Compared to Dry Foods" Animals 12, no. 15: 1881. https://doi.org/10.3390/ani12151881