
Comparisons of Corn Stover Silages after Fresh- or Ripe-Corn Harvested: Effects on Digestibility and Rumen Fermentation in Growing Beef Cattle
 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Corn Stover Silages
2.2. In Vitro Incubation
2.3. Animal Trial
2.3.1. Cattle and Diets
2.3.2. Nutrient Digestibility
2.3.3. Rumen Sampling
2.4. Sample Analysis
2.4.1. Chemical Composition
2.4.2. Fermentation Parameters
2.4.3. DNA Extraction and Microbial Analysis
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ranum, P.; Peña-Rosas, J.P.; Garcia-Casal, M.N. Global maize production, utilization, and consumption. Ann. N. Y. Acad. Sci. 2014, 1312, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Ketthaisong, D.; Suriharn, B.; Tangwongchai, R.; Lertrat, K. Changes in physicochemical properties of waxy corn starches at different stages of harvesting. Carbohydr. Polym. 2013, 98, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Yangcheng, H.; Jiang, H.; Blanco, M.; Jane, J.L. Characterization of normal and waxy corn starch for bioethanol production. J. Agric. Food Chem. 2013, 61, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; Guo, X.; Tang, Y.; Chen, L.; Xie, F. Development changes in multi-scale structure and functional properties of waxy corn starch at different stages of kernel growth. Int. J. Biol. Macromol. 2021, 191, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Sousa, D.O.; Velasquez, A.V.; Oliveira, C.A.; Souza, J.M.; Nadeau, E.; Silva, L.F.P. Effect of sugarcane genotype and maturity stage at harvest on feed intake and ruminal parameters of growing steers. Anim. Feed Sci. Tech. 2019, 256, 114258. [Google Scholar] [CrossRef]
- Pang, D.; Yan, T.; Krizsan, S. Effect of strategy for harvesting regrowth grass silage on performance in dairy cows. J. Dairy Sci. 2021, 104, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Garcia, S.C.; Fulkerson, W.J.; Barchia, I.M. Degradation kinetics of leaves, petioles and stems of forage rape (Brassica napus) as affected by maturity. Anim. Feed Sci. Tech. 2011, 168, 165–178. [Google Scholar] [CrossRef]
- Nazli, M.H.; Halim, R.A.; Abdullah, A.M.; Hussin, G.; Samsudin, A.A. Potential of four corn varieties at different harvest stages for silage production in Malaysia. Asian-Australas. J. Anim. Sci. 2019, 32, 224–232. [Google Scholar] [CrossRef]
- Ma, Y.L.; Khan, M.Z.; Liu, Y.F.; Xiao, J.X.; Chen, X.; Ji, S.K.; Cao, Z.J.; Li, S.L. Analysis of nutrient composition, rumen degradation characteristics, and feeding value of Chinese rye grass, barley grass, and naked oat straw. Animals 2021, 11, 2486. [Google Scholar] [CrossRef]
- Moraïs, S.; Mizrahi, I. Islands in the stream: From individual to communal fiber degradation in the rumen ecosystem. FEMS Microbiol. Rev. 2019, 43, 362–379. [Google Scholar] [CrossRef]
- Borreani, G.; Tabacco, E.; Schmidt, R.J.; Holmes, B.J.; Muck, R.E. Silage review: Factors affecting dry matter and quality losses in silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menke, K.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider, W. The estimation of the digestibility and metabolizable energy content of ruminant feedingstuffs from the gas production when they are incubated with rumen liquor in vitro. J. Agric. Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, R.; Yang, S.; Deng, J.P.; Tang, S.X.; Tan, Z.L. Effects of three methane mitigation agents on parameters of kinetics of total and hydrogen gas production, ruminal fermentation and hydrogen balance using in vitro technique. Anim. Sci. J. 2016, 87, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.F.; Zhang, Z.Y. Animal Nutrition Parameters and Feeding Standard; China Agricultural Press: Beijing, China, 1998. [Google Scholar]
- Feng, Y.L. Nutritional Requirements and Feeding Standard for Beef Cattle; China Agricultural University Press: Beijing, China, 2000. [Google Scholar]
- Chen, H.; Wang, C.; Huasai, S.; Chen, A. Effects of dietary forage to concentrate ratio on nutrient digestibility, ruminal fermentation and rumen bacterial composition in Angus cows. Sci. Rep. 2021, 11, 17023. [Google Scholar] [CrossRef]
- Wang, R.; Wang, M.; Lin, B.; Ma, Z.Y.; Ungerfeld, E.M.; Wu, T.T.; Wen, J.N.; Zhang, X.M.; Deng, J.P.; Tan, Z.L. Association of fibre degradation with ruminal dissolved hydrogen in growing beef bulls fed with two types of forages. Br. J. Nutr. 2021, 125, 601–610. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Kartchner, R.J.; Theurer, B. Comparison of hydrolysis methods used in feed, digesta, and fecal starch analysis. J. Agric. Food Chem. 1981, 29, 8–11. [Google Scholar] [CrossRef]
- Wang, M.; Wang, R.; Janssen, P.H.; Zhang, X.M.; Sun, X.Z.; Pacheco, D.; Tan, Z.L. Sampling procedure for the measurement of dissolved hydrogen and volatile fatty acids in the rumen of dairy cows. J. Anim. Sci. 2016, 94, 1159–1169. [Google Scholar] [CrossRef] [Green Version]
- Shahinian, A.H.; Reinhold, J.G. Application of the phenol-hypochlorite reaction to measurement of ammonia concentrations in Kjeldahl digests of serum and various tissues. Clin. Chem. 1971, 17, 1077–1080. [Google Scholar] [CrossRef]
- Wang, M.; Ungerfeld, E.M.; Wang, R.; Zhou, C.S.; Basang, Z.Z.; Ao, S.M.; Tan, Z.L. Supersaturation of dissolved hydrogen and methane in rumen of Tibetan sheep. Front. Microbiol. 2016, 7, 850. [Google Scholar] [CrossRef]
- Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples. Biotechniques 2004, 36, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Wang, P.; He, Z.; Tang, S.; Zhou, C.; Han, X.; Wang, M.; Wu, D.; Kang, J.; Tan, Z. In vitro evaluation on neutral detergent fiber and cellulose digestion by post-ruminal microorganisms in goats. J. Sci. Food Agric. 2014, 94, 1745–1752. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Penner, G.B.; Li, M.; Oba, M.; Guan, L.L. Changes in bacterial diversity associated with epithelial tissue in the beef cow rumen during the transition to a high-grain diet. Appl. Environ. Microbiol. 2011, 77, 5770–5781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, K.R.; Yeoman, C.J.; Kent, A.; Righini, N.; Carbonero, F.; Estrada, A.; Gaskins, H.R.; Stumpf, R.M.; Yildirim, S.; Torralba, M.; et al. Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes. ISME J. 2013, 7, 1344–1353. [Google Scholar] [CrossRef]
- Sylvester, J.T.; Karnati, S.K.; Yu, Z.; Morrison, M.; Firkins, J.L. Development of an assay to quantify rumen ciliate protozoal biomass in cows using real-time PCR. J. Nutr. 2004, 134, 3378–3384. [Google Scholar] [CrossRef] [Green Version]
- Hook, S.E.; Wright, A.D.; McBride, B.W. Methanogens: Methane producers of the rumen and mitigation strategies. Archaea 2010, 2010, 945785. [Google Scholar] [CrossRef] [Green Version]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef]
- Koike, S.; Kobayashi, Y. Development and use of competitive PCR assays for the rumen cellulolytic bacteria: Fibrobacter succinogenes, Ruminococcus albus and Ruminococcus flavefaciens. FEMS Microbiol. Lett. 2001, 204, 361–366. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, R.; Wang, M.; Zhang, X.; Mao, H.; Tan, Z. Short communication: Variability in fermentation end-products and methanogen communities in different rumen sites of dairy cows. J. Dairy Sci. 2018, 101, 5153–5158. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Eco. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarepoua, E.; Tangwongchai, R.; Suriharn, B.; Lertrat, K. Influence of variety and harvest maturity on phytochemical content in corn silk. Food Chem. 2015, 169, 424–429. [Google Scholar] [CrossRef]
- Khalilian, M.E.; Habibi, D.; Golzardi, F.; Aghayari, F.; Khazaei, A. Effect of maturity stage on yield, morphological characteristics, and feed value of sorghum [Sorghum bicolor (L.) Moench] cultivars. Cereal Res. Commun. 2022, in press. [Google Scholar] [CrossRef]
- Cabezas-Garcia, E.H.; Krizsan, S.J.; Shingfield, K.J.; Huhtanen, P. Effects of replacement of late-harvested grass silage and barley with early-harvested silage on milk production and methane emissions. J. Dairy Sci. 2017, 100, 5228–5240. [Google Scholar] [CrossRef] [Green Version]
- Mc Geough, E.J.; O’Kiely, P.; Foley, P.A.; Hart, K.J.; Boland, T.M.; Kenny, D.A. Methane emissions, feed intake, and performance of finishing beef cattle offered maize silages harvested at 4 different stages of maturity. J. Anim. Sci. 2010, 88, 1479–1491. [Google Scholar] [CrossRef] [Green Version]
- Galyean, M.L.; Owens, F.N. Effects of diet composition and level of feed intake on site and extent of digestion in ruminants. In Physiological Aspects of Digestion and Metabolism in Ruminants; Tsuda, T., Sasaki, Y., Kawashima, R., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 483–514. [Google Scholar]
- Grimaud, P.; Richard, D.; Vergeron, M.P.; Guilleret, J.R.; Doreau, M. Effect of drastic undernutrition on digestion in Zebu cattle receiving a diet based on rice straw. J. Dairy Sci. 1999, 82, 974–981. [Google Scholar] [CrossRef]
- Colucci, P.E.; Macleod, G.K.; Grovum, W.L.; Cahill, L.W.; McMillan, I. Comparative digestion in sheep and cattle fed different forage to concentrate ratios at high and low intakes. J. Dairy Sci. 1989, 72, 1774–1785. [Google Scholar] [CrossRef]
- Zhang, X.M.; Wang, M.; Yu, Q.; Ma, Z.Y.; Beauchemin, K.A.; Wang, R.; Wen, J.N.; Lukuyu, B.A.; Tan, Z.L. Liquid hot water treatment of rice straw enhances anaerobic degradation and inhibits methane production during in vitro ruminal fermentation. J. Dairy Sci. 2020, 103, 4252–4261. [Google Scholar] [CrossRef]
- Lage, C.F.A.; Coelho, S.G.; Diniz Neto, H.C.; Malacco, V.M.R.; Rodrigues, J.P.P.; Sacramento, J.P.; Teixeira, V.A.; Machado, F.S.; Pereira, L.G.R.; Tomich, T.R.; et al. Relationship between feed efficiency indexes and thermography, blood, and ruminal parameters in pre-weaning dairy heifers. PLoS ONE 2020, 15, e0236118. [Google Scholar] [CrossRef] [PubMed]
- McGovern, E.; Kenny, D.A.; McCabe, M.S.; Fitzsimons, C.; McGee, M.; Kelly, A.K.; Waters, S.M. 16S rRNA sequencing reveals relationship between potent cellulolytic genera and feed efficiency in the rumen of bulls. Front. Microbiol. 2018, 9, 1842. [Google Scholar] [CrossRef] [PubMed]
- Au Robinson, P.H.; Tamminga, S.; Vanvuuren, A.M. Influence of declining level of feed-intake and varying the proportion of starch in the concentrate on rumen ingesta quantity, composition and kinetics of ingesta turnover in dairy-cows. Livest. Prod. Sci. 1989, 17, 37–62. [Google Scholar] [CrossRef]
- Wang, M.; Wang, R.; Xie, T.Y.; Janssen, P.H.; Sun, X.Z.; Beauchemin, K.A.; Tan, Z.L.; Gao, M. Shifts in rumen fermentation and microbiota are associated with dissolved ruminal hydrogen concentrations in lactating dairy cows fed different types of carbohydrates. J. Nutr. 2016, 146, 1714–1721. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.; Yang, C. Ruminal methane production: Associated microorganisms and the potential of applying hydrogen-utilizing bacteria for mitigation. Sci. Total Environ. 2019, 654, 1270–1283. [Google Scholar] [CrossRef] [PubMed]
- Clauss, M.; Dittmann, M.T.; Vendl, C.; Hagen, K.B.; Frei, S.; Ortmann, S.; Müller, D.W.H.; Hammer, S.; Munn, A.J.; Schwarm, A.; et al. Review: Comparative methane production in mammalian herbivores. Animal 2020, 14, s113–s123. [Google Scholar] [CrossRef] [Green Version]
- Huws, S.A.; Creevey, C.J.; Oyama, L.B.; Mizrahi, I.; Denman, S.E.; Popova, M.; Muñoz-Tamayo, R.; Forano, E.; Waters, S.M.; Hess, M.; et al. Addressing global ruminant agricultural challenges through understanding the rumen microbiome: Past, present, and future. Front. Microbiol. 2018, 9, 2161. [Google Scholar] [CrossRef]
- Gruninger, R.J.; Nguyen, T.T.M.; Reid, I.D.; Yanke, J.L.; Wang, P.; Abbott, D.W.; Tsang, P.; McAllister, T. Application of transcriptomics to compare the carbohydrate active enzymes that are expressed by diverse genera of anaerobic fungi to degrade plant cell wall carbohydrates. Front. Microbiol. 2018, 9, 1581. [Google Scholar] [CrossRef] [Green Version]
- Qi, M.; Wang, P.; O’Toole, N.; Barboza, P.S.; Ungerfeld, E.; Leigh, M.B.; Selinger, L.B.; Butler, G.; Tsang, A.; McAllister, T.A.; et al. Snapshot of the eukaryotic gene expression in muskoxen rumen—A metatranscriptomic approach. PLoS ONE 2011, 6, e20521. [Google Scholar] [CrossRef] [Green Version]
- Raut, M.P.; Couto, N.; Karunakaran, E.; Biggs, C.A.; Wright, P.C. Deciphering the unique cellulose degradation mechanism of the ruminal bacterium Fibrobacter succinogenes S85. Sci. Rep. 2019, 9, 16542. [Google Scholar] [CrossRef]
- Huang, G.; Guo, L.; Chang, X.; Liu, K.; Tang, W.; Zheng, N.; Zhao, S.; Zhang, Y.; Wang, J. Effect of whole or ground flaxseed supplementation on fatty acid profile, fermentation, and bacterial composition in rumen of dairy cows. Front. Microbiol. 2021, 12, 760528. [Google Scholar] [CrossRef] [PubMed]


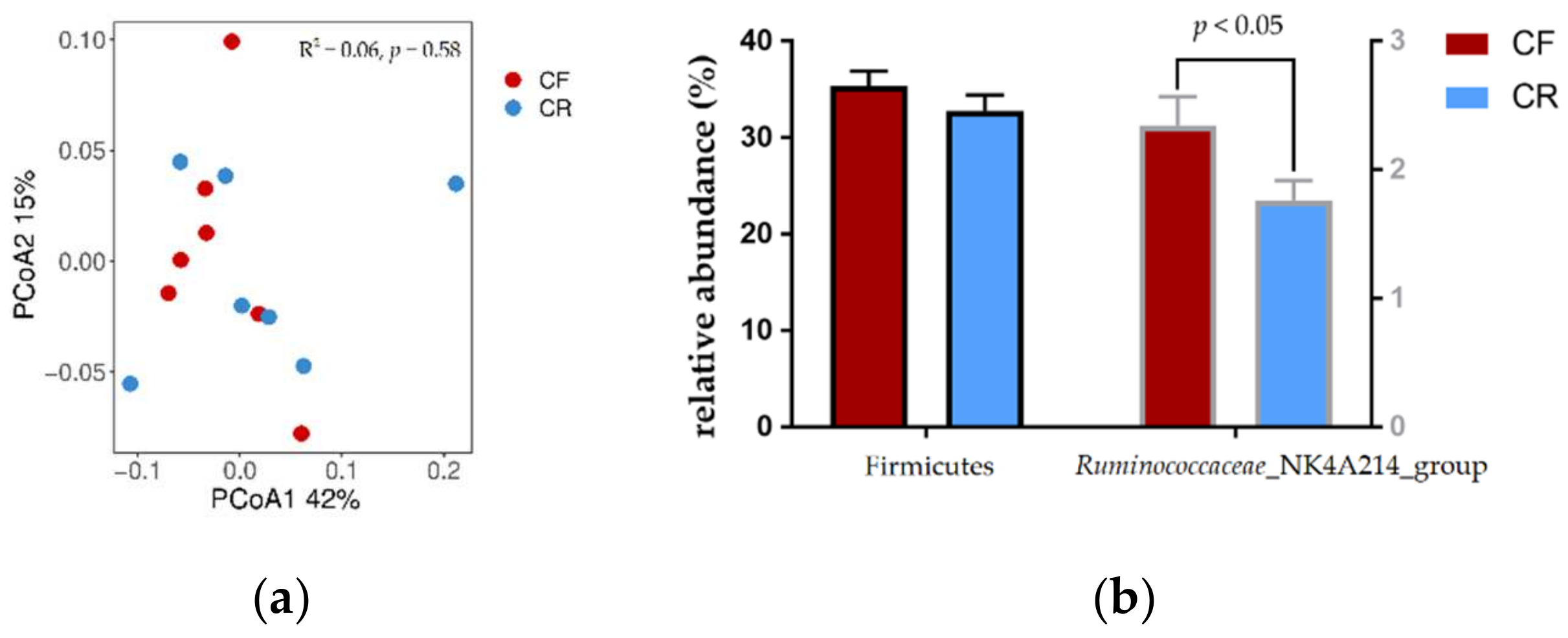{kind=link}
| Items | Silages | |
|---|---|---|
| CF | CR | |
| DM 1 (g/kg) | 232 | 249 |
| Substrate on a DM basis (g/kg DM) | ||
| OM 1 | 928 | 926 |
| CP 1 | 60 | 59 |
| NDF 1 | 539 | 615 |
| ADF 1 | 287 | 366 |
| Starch | 78 | 78 |
| GE 1 (MJ/kg DM) | 17.5 | 16.7 |
| Items | Diets | |
|---|---|---|
| CF | CR | |
| Dietary ingredient (g/kg DM 1) | ||
| CF silage | 530 | 265 |
| CR silage | 0 | 265 |
| Maize bran | 304 | 304 |
| Wheat bran | 56 | 56 |
| Soybean meal | 85 | 85 |
| Limestone powder | 5 | 5 |
| Saccharomyces cerevisiae | 10 | 10 |
| NaCl | 5 | 5 |
| Premix 2 | 5 | 5 |
| Chemical composition (g/kg DM) | ||
| OM 1 | 922 | 921 |
| CP 1 | 125 | 118 |
| NDF 1 | 375 | 400 |
| ADF 1 | 161 | 183 |
| Starch | 210 | 210 |
| NEm 3 (MJ/kg DM) | 6.78 | 6.53 |
| NEf 3 (MJ/kg DM) | 4.23 | 3.98 |
| Microbial Groups | Sequence 1 (5′→3′) | Reference |
|---|---|---|
| Bacteria | F: ACTCCTACGGGAGGCAGCA R: GGACTACHVGGGTWTCTAAT | [26] |
| Protozoa | F: GCTTTCGWTGGTAGTGTATT R: CTTGCCCTCYAATCGTWCT | [27] |
| Methanogens | F: GGATTAGATACCCSGGTAGT R: GTTGARTCCAATTAAACCGCA | [28] |
| Fungi | F: GAGGAAGTAAAAGTCGTAACAAGGTTTC R: CAAATTCACAAAGGGTAGGATGATT | [29] |
| Selected groups of bacteria | ||
| Ruminococcus albus | F: CCCTAAAAGCAGTCTTAGTTCG R: CCTCCTTGCGGTTAGAACA | [30] |
| Ruminococcus flavefaciens | F: GAACGGAGATAATTTGAGTTTACTTAGG R: CGGTCTCTGTATGTTATGAGGTATTACC | [29] |
| Fibrobacter succinogenes | F: GTTCGGAATTACTGGGCGTAAA R: CGCCTGCCCCTGAACTATC | [29] |
| Items | Silages | SEM | p-Value | |
|---|---|---|---|---|
| CF | CR | |||
| Substrate degradation (g/kg) | ||||
| DM 1 | 575 | 585 | 10.7 | 0.64 |
| NDF 1 | 535 | 558 | 10.0 | 0.12 |
| ADF 1 | 503 | 511 | 5.3 | 0.39 |
| pH | 6.49 | 6.61 | 0.016 | 0.003 |
| Total VFAs 1 (mM) | 82.3 | 87.7 | 3.80 | 0.02 |
| Molar percentage of individual VFA (mol/100 mol) | ||||
| Acetate | 63.4 | 64.1 | 1.38 | 0.60 |
| Propionate | 22.4 | 21.1 | 0.29 | 0.046 |
| Butyrate | 9.0 | 10.5 | 0.88 | 0.21 |
| Valerate | 1.2 | 1.2 | 0.18 | 0.84 |
| Isobutyrate | 1.2 | 1.2 | 0.12 | 0.91 |
| Isovalerate | 2.0 | 1.9 | 0.27 | 0.75 |
| Acetate to propionate ratio | 2.83 | 3.04 | 0.03 | 0.003 |
| Items | Diets | SEM | p-Value | |
|---|---|---|---|---|
| CF | CR | |||
| Intake (kg/d) | ||||
| DM 1 | 7.47 | 7.97 | 0.210 | 0.04 |
| OM 1 | 6.90 | 7.34 | 0.194 | 0.04 |
| CP 1 | 0.92 | 0.94 | 0.026 | 0.43 |
| NDF 1 | 2.93 | 3.14 | 0.083 | 0.02 |
| ADF 1 | 1.28 | 1.42 | 0.036 | 0.002 |
| Starch | 1.47 | 1.70 | 0.043 | <0.001 |
| Total-tract apparent digestibility (g/kg) | ||||
| DM | 670 | 636 | 11.9 | 0.02 |
| OM | 695 | 664 | 10.9 | 0.02 |
| CP | 645 | 621 | 16.1 | 0.16 |
| NDF | 582 | 535 | 12.8 | 0.01 |
| ADF | 520 | 468 | 33.0 | 0.15 |
| Starch | 877 | 874 | 4.76 | 0.59 |
| ADG 1 (kg/d) | 1.19 | 1.15 | 0.093 | 0.37 |
| Items | Diets | Sampling Time (h) | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| CF | CR | 0 | 2.5 | Diet | Time | Diet × Time | ||
| Dissolved gases | ||||||||
| Hydrogen (μM) | 0.50 | 0.84 | 0.88 | 0.46 | 0.132 | 0.21 | 0.12 | 0.43 |
| Methane (mM) | 0.71 | 0.77 | 0.80 | 0.68 | 0.026 | 0.25 | 0.03 | 0.19 |
| pH | 6.89 | 6.82 | 6.93 | 6.78 | 0.013 | 0.02 | <0.001 | 0.008 |
| Ammonia-N (mM) | 7.51 | 6.87 | 3.98 | 10.4 | 0.426 | 0.46 | <0.001 | 0.04 |
| Total VFA 1 (mM) | 67.0 | 62.2 | 56.6 | 72.6 | 2.52 | 0.14 | <0.001 | 0.70 |
| Molar percentage of individual VFA (mol/100 mol) | ||||||||
| Acetate | 65.4 | 64.9 | 67.6 | 62.7 | 0.296 | 0.48 | <0.001 | 0.87 |
| Propionate | 19.5 | 19.3 | 17.4 | 21.4 | 0.286 | 0.65 | <0.001 | 0.49 |
| Butyrate | 10.7 | 11.3 | 10.7 | 11.4 | 0.330 | 0.18 | 0.12 | 0.20 |
| Valerate | 1.3 | 1.3 | 1.0 | 1.6 | 0.052 | 0.61 | <0.001 | 0.92 |
| Isobutyrate | 1.3 | 1.3 | 1.4 | 1.2 | 0.015 | 0.03 | <0.001 | 0.17 |
| Isovalerate | 1.8 | 1.8 | 1.9 | 1.8 | 0.039 | 0.69 | 0.10 | 0.45 |
| Acetate to propionate ratio | 3.42 | 3.45 | 3.91 | 2.95 | 0.184 | 0.80 | <0.001 | 0.68 |
| Estimated net hydrogen production relative to the amount of total VFA produced (mol/mol) | ||||||||
| 1.32 | 1.33 | 1.39 | 1.26 | 0.013 | 0.57 | <0.001 | 0.34 | |
| Items | Diets | SEM | p-Value | |
|---|---|---|---|---|
| CF | CR | |||
| Bacteria | 12.1 | 12.0 | 0.030 | 0.17 |
| Protozoa | 10.7 | 10.4 | 0.042 | 0.01 |
| Methanogens | 8.73 | 8.57 | 0.043 | 0.09 |
| Fungi | 7.95 | 7.43 | 0.073 | 0.01 |
| Selected groups of bacteria | ||||
| Ruminococcus albus | 8.94 | 8.72 | 0.060 | 0.10 |
| Ruminococcus flavefaciens | 8.69 | 8.54 | 0.053 | 0.18 |
| Fibrobacter succinogenes | 9.77 | 9.58 | 0.027 | 0.004 |
| Items | Diets | SEM | p-Value | |
|---|---|---|---|---|
| CF | CR | |||
| Alpha diversity indexes | ||||
| Chao1 | 1956 | 2083 | 67.4 | 0.36 |
| Species | 1384 | 1460 | 40.8 | 0.37 |
| Shannon | 8.64 | 8.64 | 0.101 | 0.97 |
| Coverage | 0.966 | 0.963 | 0.001 | 0.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Wang, R.; Wu, T.; Yang, Y.; He, Z.; Ma, Z.; Tan, Z.; Lin, B.; Wang, M. Comparisons of Corn Stover Silages after Fresh- or Ripe-Corn Harvested: Effects on Digestibility and Rumen Fermentation in Growing Beef Cattle. Animals 2022, 12, 1248. https://doi.org/10.3390/ani12101248
Zhang M, Wang R, Wu T, Yang Y, He Z, Ma Z, Tan Z, Lin B, Wang M. Comparisons of Corn Stover Silages after Fresh- or Ripe-Corn Harvested: Effects on Digestibility and Rumen Fermentation in Growing Beef Cattle. Animals. 2022; 12(10):1248. https://doi.org/10.3390/ani12101248
Chicago/Turabian StyleZhang, Min, Rong Wang, Tingting Wu, Yingbai Yang, Zhixiong He, Zhiyuan Ma, Zhiliang Tan, Bo Lin, and Min Wang. 2022. "Comparisons of Corn Stover Silages after Fresh- or Ripe-Corn Harvested: Effects on Digestibility and Rumen Fermentation in Growing Beef Cattle" Animals 12, no. 10: 1248. https://doi.org/10.3390/ani12101248