
Saliva and Blood Cortisol Measurement in Bottlenose Dolphins (Tursiops truncatus): Methodology, Application, and Limitations

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
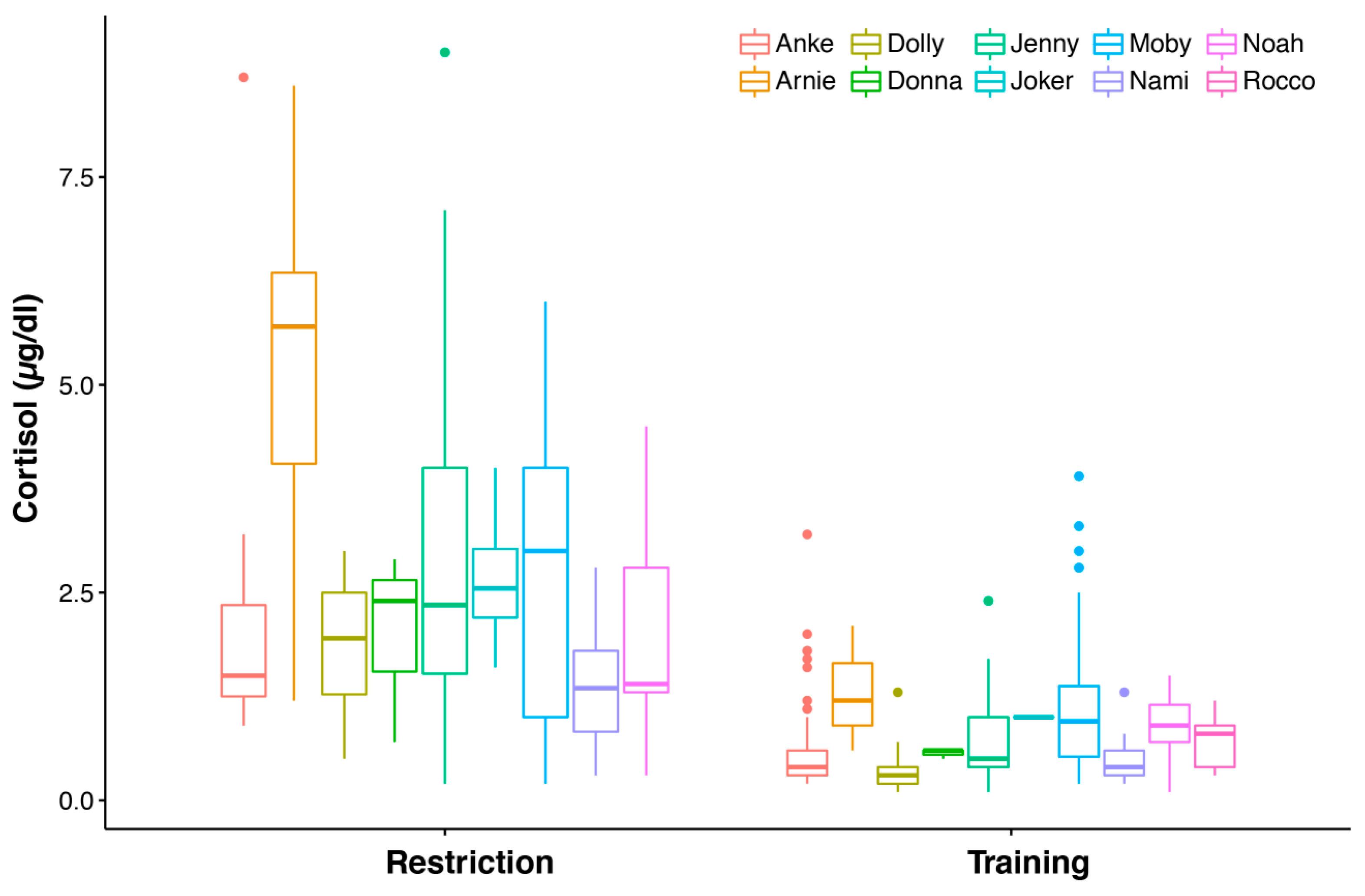
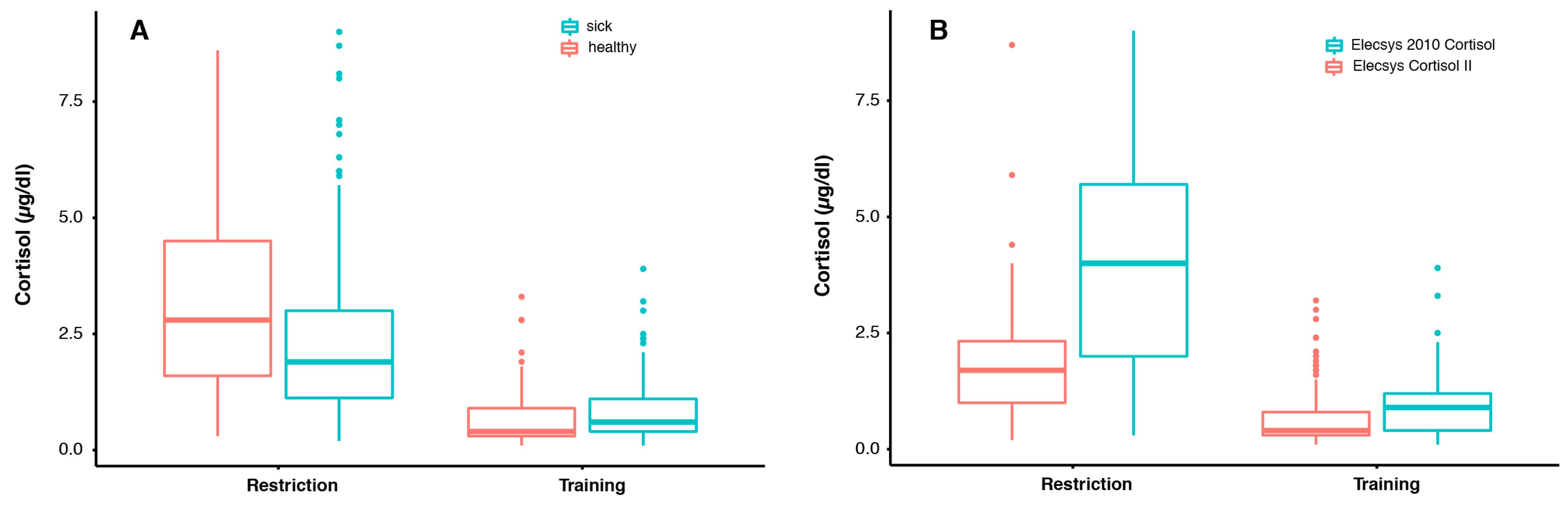
3.1. Blood Cortisol Values under Two Different Conditions
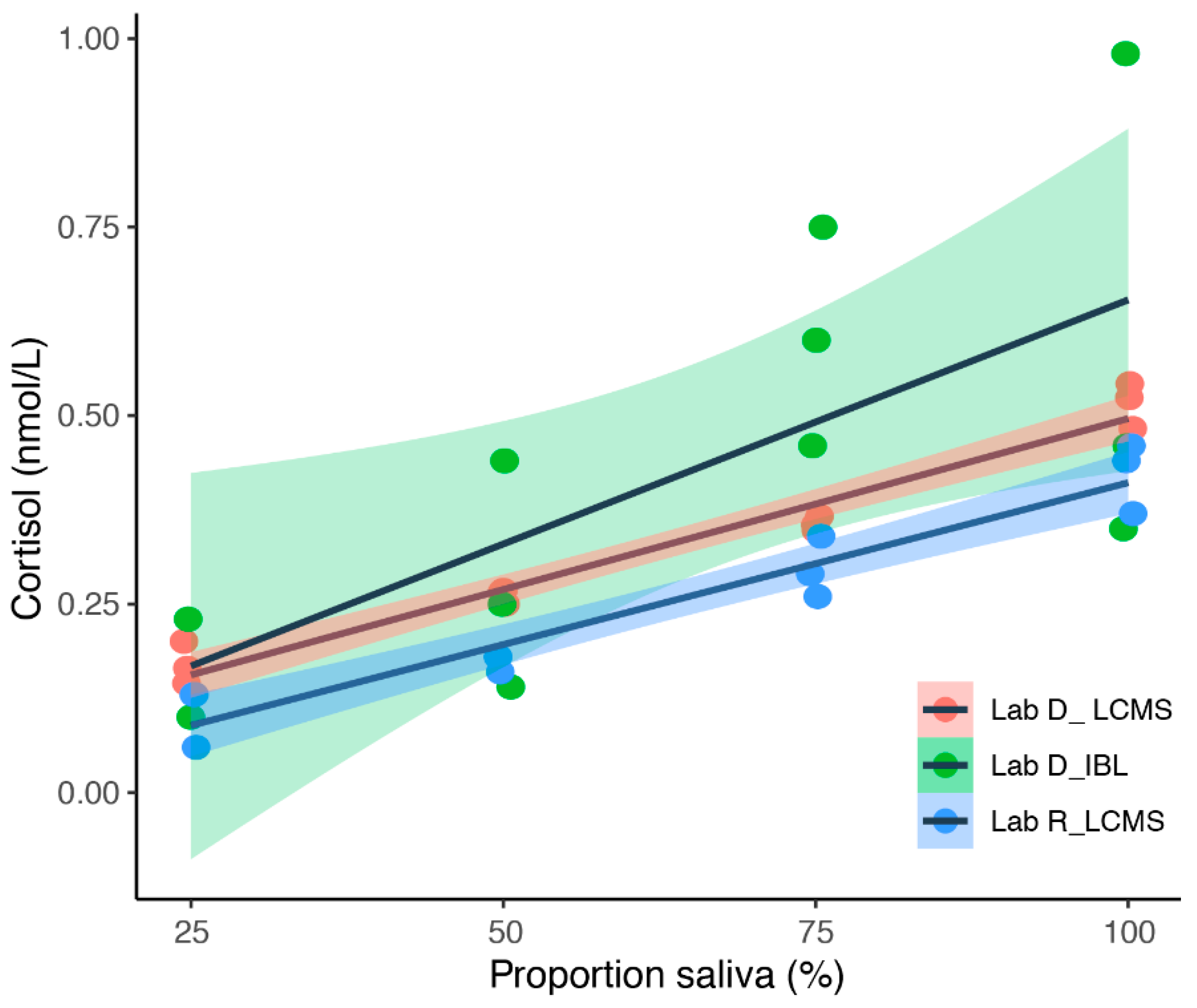
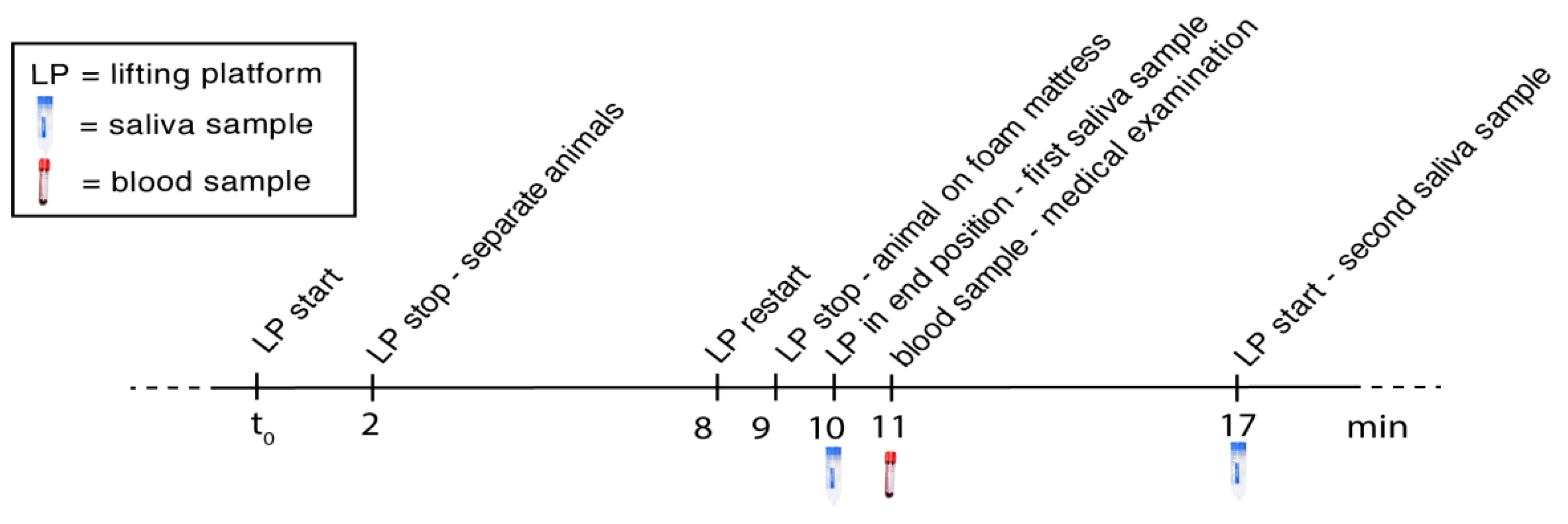
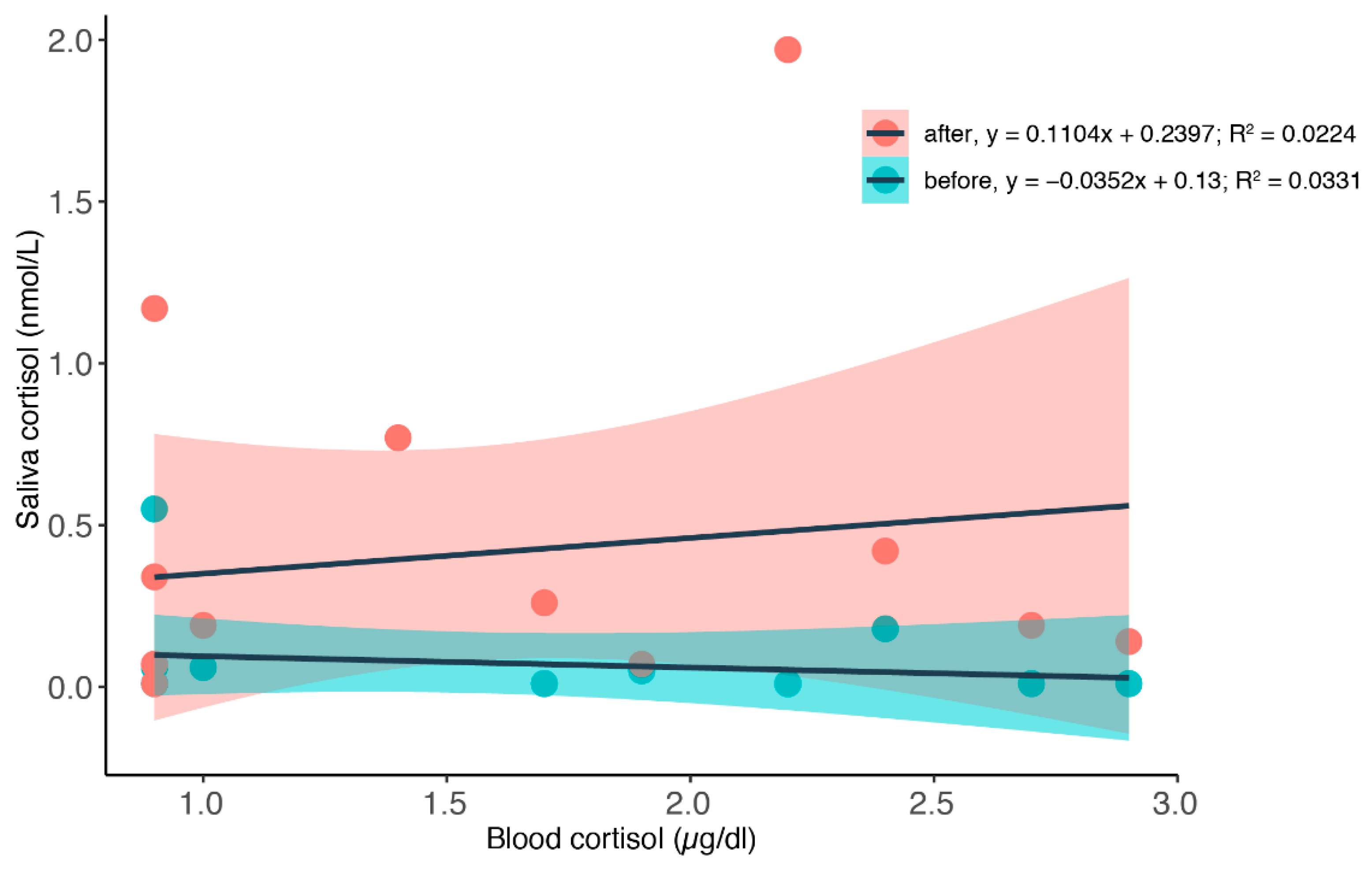
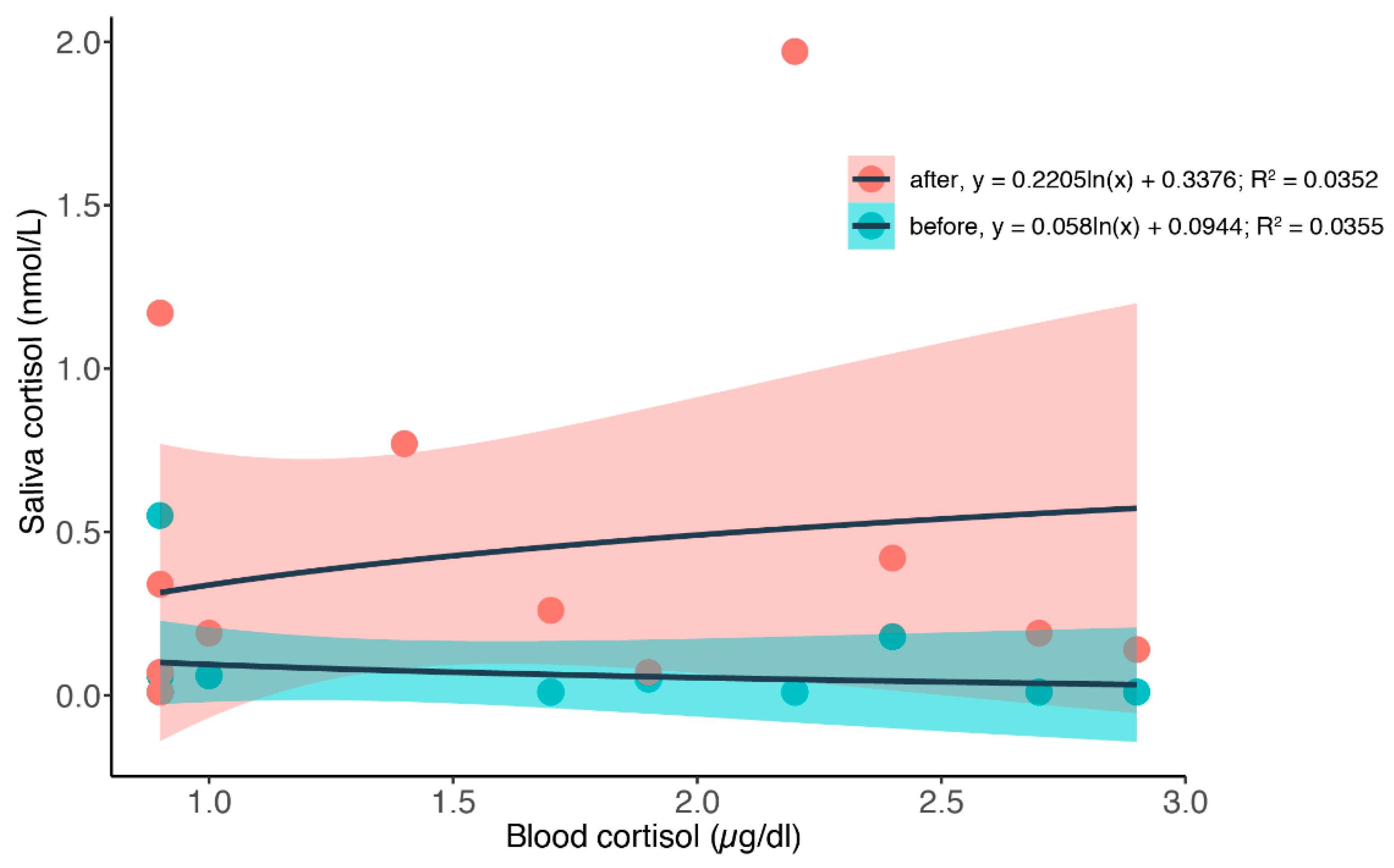
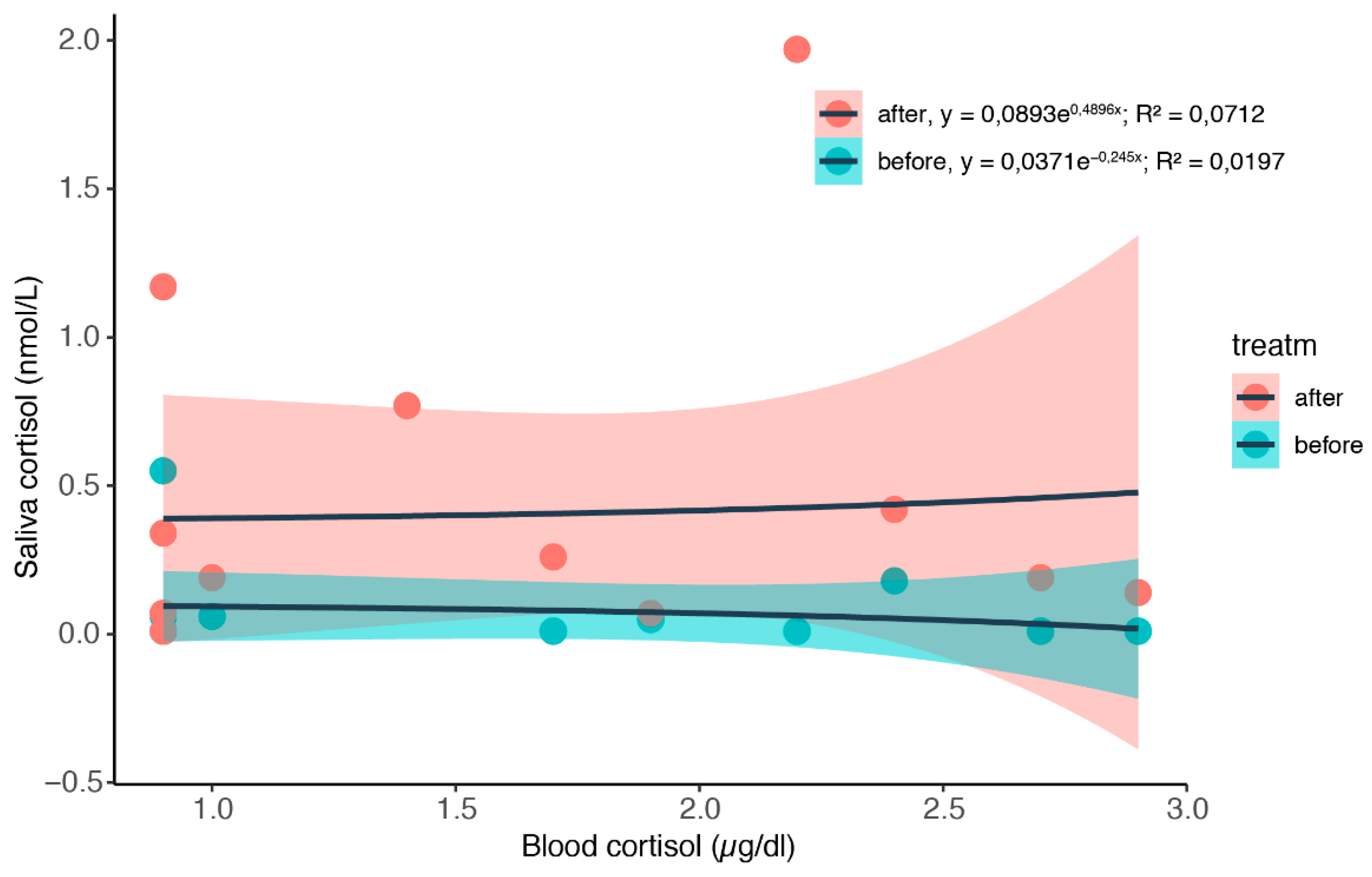
3.2. Simultaneous Blood and Saliva Sampling
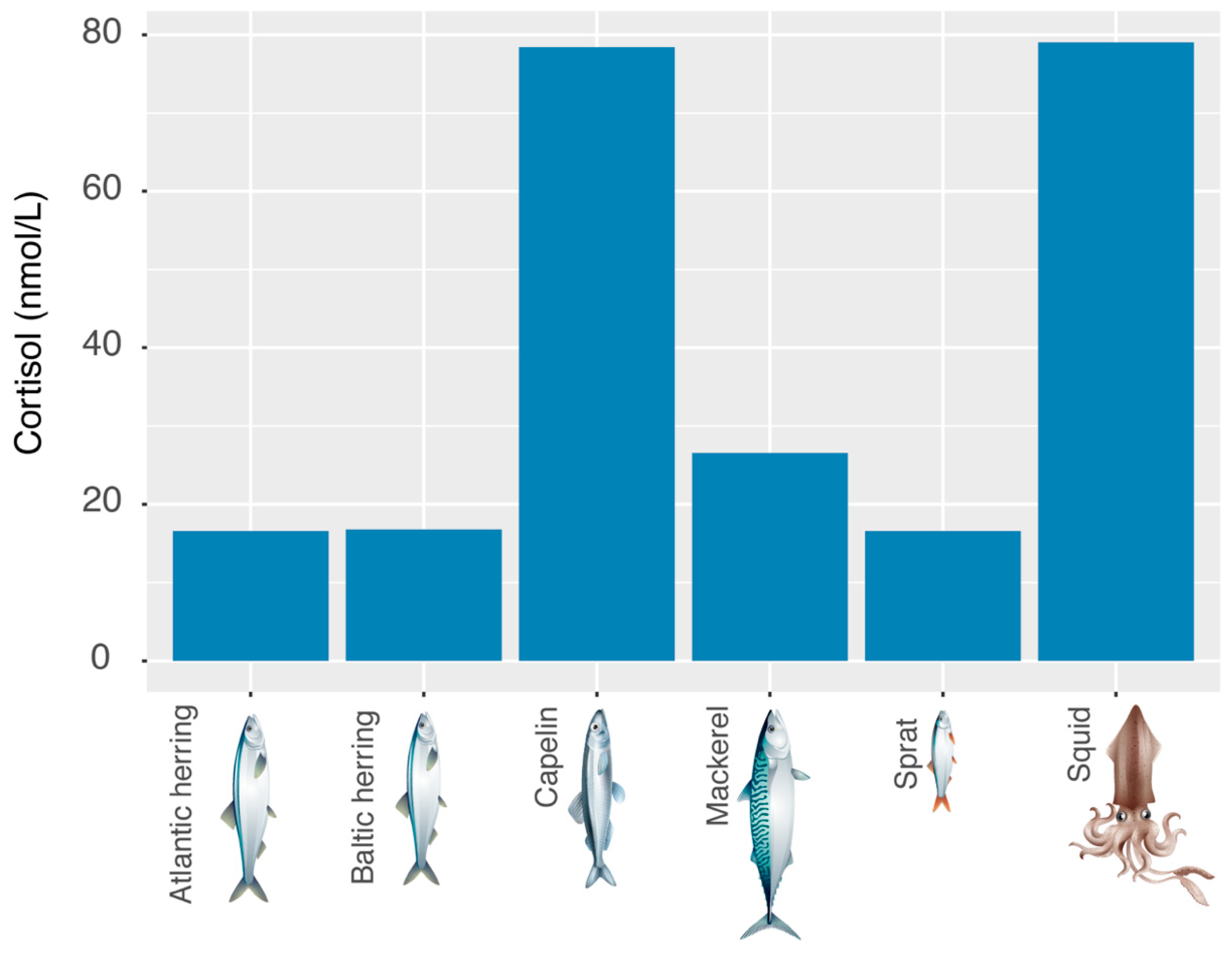
3.3. Cortisol Contamination of Fodder Fish
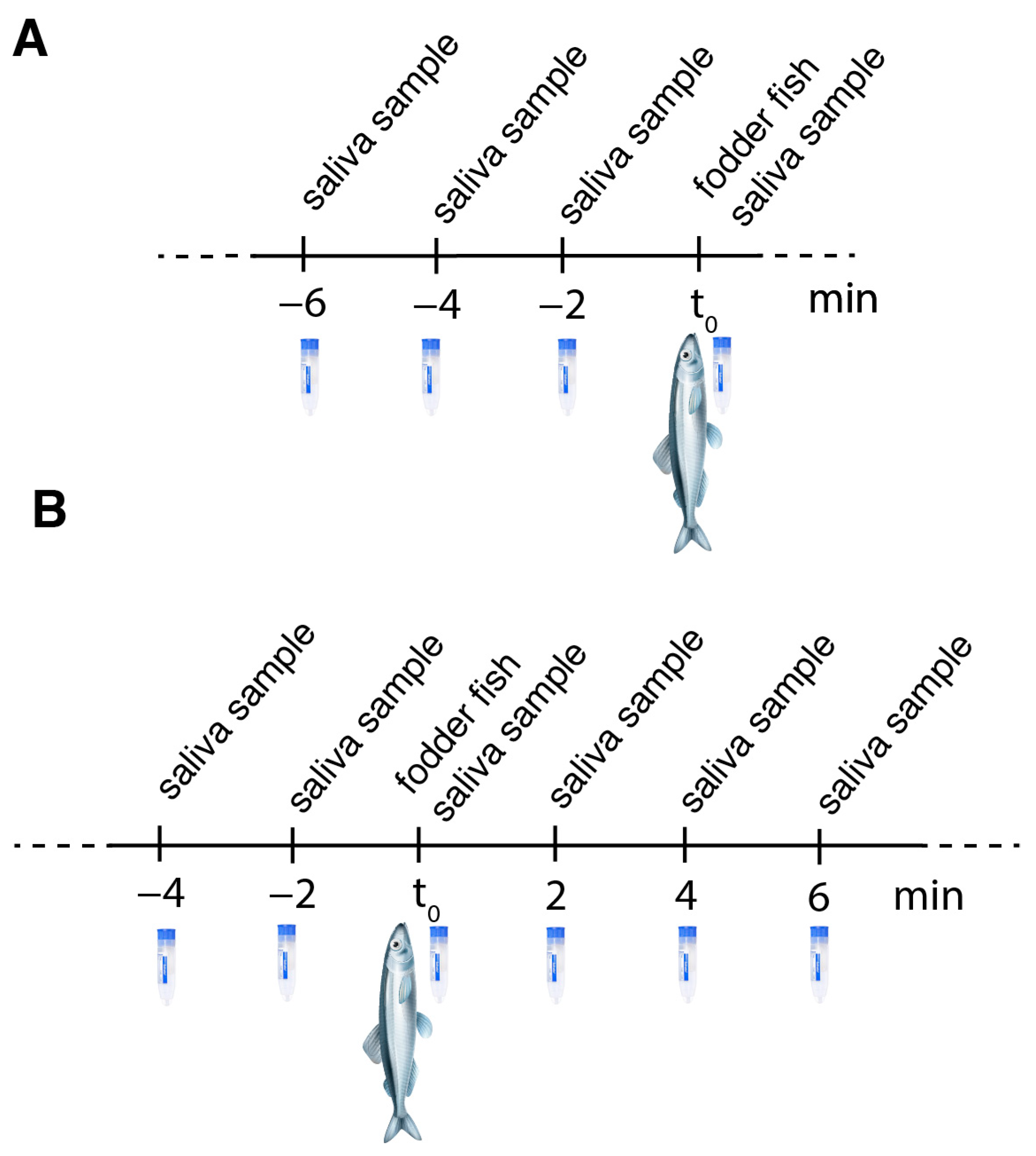
3.4. Feeding Experiment I
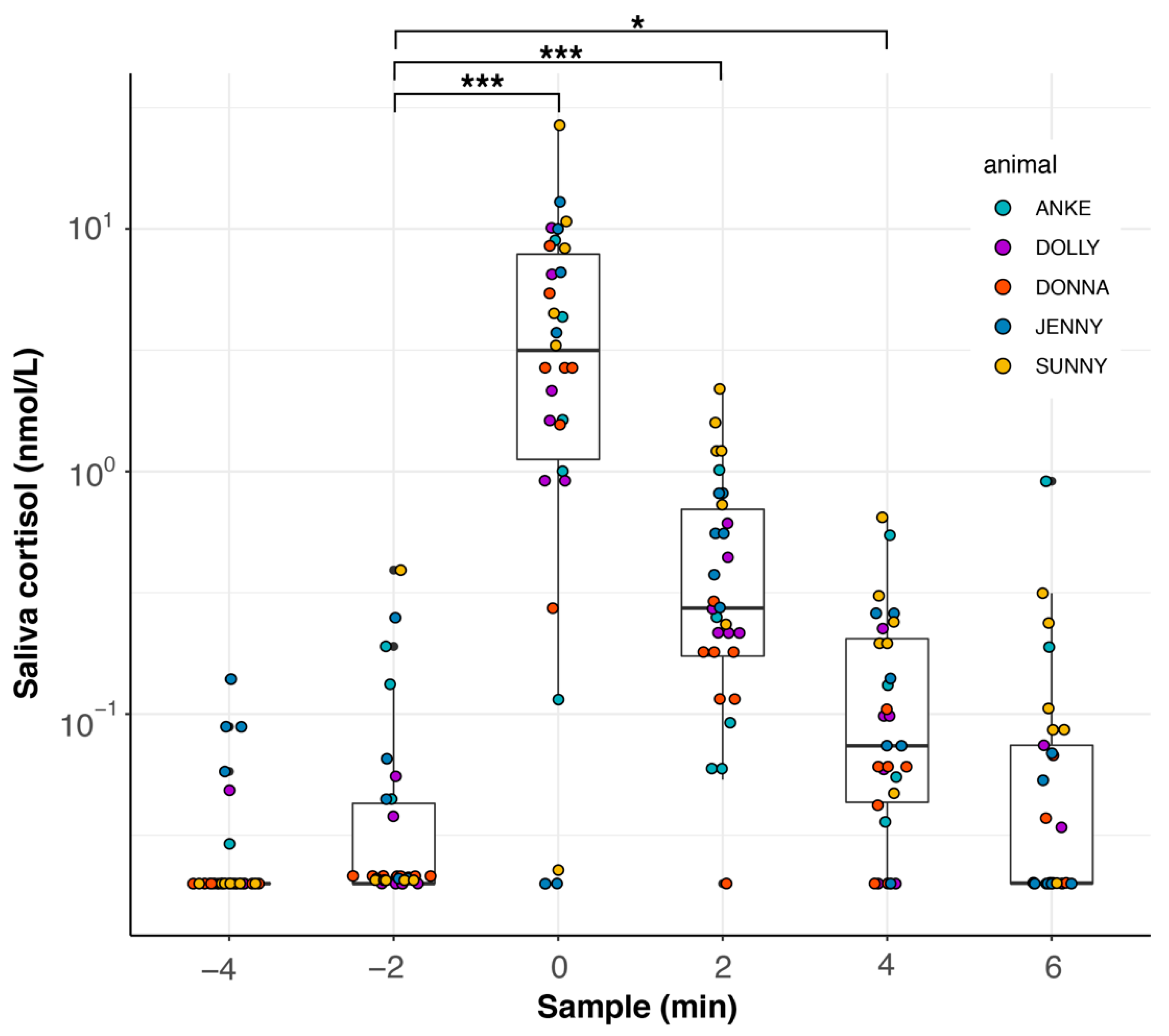
3.5. Feeding Experiment II
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Do not feed the dolphin before sampling.
- Wear gloves.
- Remove the cotton swab from the plastic tube.
- Collect saliva from the side of the tongue for 5 s.
- Put the swab back into the tube.
- Record the date, time of the collection, and name of the animal on the plastic tube.
- If more than one saliva sample is taken, reward the animal with ice cubes or gelatin (only if necessary). If available, take notes on the documentation sheet.
- Send tubes to the veterinarians.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LMM-Model | Observation Periods | Estimate | Std. Error | Df | T-Value | p-Value |
|---|---|---|---|---|---|---|
| Blood cortisol | Intercept | 0.38350 | 0.14475 | 16.75493 | 2.649 | 0.017 * |
| REML | Treatment training | −1.08618 | 0.07620 | 464.74523 | −14.253 | <2 × 10−16 *** |
| AIC: 1008.779 | Elecsys Cortisol III | 0.45200 | 0.07766 | 461.23621 | 5.820 | 1.1 × 10−8 *** |
| Health status sick | 0.04378 | 0.06810 | 463.24528 | 0.643 | 0.521 |

| Contrast | Estimate | SE | df | T Ratio | p-Value |
|---|---|---|---|---|---|
| −6 effect vs. −4 effect | −0.595 | 0.287 | 122 | −2.074 | 0.1675 |
| −6 effect vs. −2 effect | −0.937 | 0.294 | 122 | −3.185 | 0.0098 |
| −6 effect vs. 0 effect | −2.305 | 0.291 | 122 | −7.911 | <0.0001 |
| −4 effect vs. −2 effect | −0.342 | 0.292 | 122 | −1.173 | 0.6451 |
| −4 effect vs. 0 effect | −1.71 | 0.289 | 122 | −5.916 | <0.0001 |
| −2 effect vs. 0 effect | −1.368 | 0.297 | 122 | −4.614 | 0.0001 |
| Contrast | Estimate | SE | df | T Ratio | p-Value |
|---|---|---|---|---|---|
| −4 effect vs. −2 effect | −0.242 | 0.289 | 179 | −0.838 | 0.9601 |
| −4 effect vs. 0 effect | −4.339 | 0.289 | 179 | −15.012 | <0.0001 |
| −4 effect vs. 2 effect | −2.492 | 0.289 | 179 | −8.623 | <0.0001 |
| −4 effect vs. 4 effect | −1.19 | 0.289 | 179 | −4.119 | 0.0008 |
| −4 effect vs. 6 effect | −0.51 | 0.292 | 179 | −1.749 | 0.5012 |
| −2 effect vs. 0 effect | −4.097 | 0.289 | 179 | −14.174 | <0.0001 |
| −2 effect vs. 2 effect | −2.25 | 0.289 | 179 | −7.785 | <0.0001 |
| −2 effect vs. 4 effect | −0.948 | 0.289 | 179 | −3.281 | 0.0155 |
| −2 effect vs. 6 effect | −0.268 | 0.292 | 179 | −0.919 | 0.9412 |
| 0 effect vs. 2 effect | 1.847 | 0.289 | 179 | 6.389 | <0.0001 |
| 0 effect vs. 4 effect | 3.148 | 0.289 | 179 | 10.894 | <0.0001 |
| 0 effect vs. 6 effect | 3.829 | 0.292 | 179 | 13.131 | <0.0001 |
| 2 effect vs. 4 effect | 1.302 | 0.289 | 179 | 4.504 | 0.0002 |
| 2 effect vs. 6 effect | 1.982 | 0.292 | 179 | 6.798 | <0.0001 |
| 4 effect vs. 6 effect | 0.68 | 0.292 | 179 | 2.333 | 0.1863 |


References
- Clegg, I.; Van Elk, C.; Delfour, F. Applying welfare science to bottlenose dolphins (Tursiops truncatus). Anim. Welf. 2017, 26, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Von Fersen, L.; Encke, D.; Hüttner, T.; Baumgartner, K. Establishment and implementation of an animal welfare decision tree to evaluate the welfare of zoo animals. Aquat. Mamm. 2018, 44, 211–220. [Google Scholar] [CrossRef]
- Rees, P.A. Studying Captive Animals: A Workbook of Methods in Behaviour, Welfare and Ecology; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Romano, M.C.; Zulema Rodas, A.; Valdez, R.A.; Elizabeth Hernandez, S.; Galindo, F.; Canales, D.; Maria Brousset, D. Stress in wildlife species: Noninvasive monitoring of glucocorticoids. Neuroimmunomodulation 2010, 17, 209–212. [Google Scholar] [CrossRef]
- Berger, A. Activity patterns, chronobiology and the assessment of stress and welfare in zoo and wild animals. Int. Zoo Yearb. 2011, 45, 80–90. [Google Scholar] [CrossRef]
- Webster, J. Assessment of animal welfare: The five freedoms. In Animal Welfare: A Cool Eye Towards Eden; Blackwell Science: Oxford, UK, 1994; pp. 10–14. [Google Scholar]
- Mellor, D.J. Updating animal welfare thinking: Moving beyond the “five freedoms” towards “a life worth living”. Animals 2016, 6, 21. [Google Scholar] [CrossRef]
- Clegg, I.L.K.; Borger-Turner, J.L.; Eskelinen, H.C. C-well: The development of a welfare assessment index for captive bottlenose dolphins (Tursiops truncatus). Anim. Welf. 2015, 24, 267–282. [Google Scholar] [CrossRef]
- Thompson, L.A.; Spoon, T.R.; Goertz, C.E.; Hobbs, R.C.; Romano, T.A. Blow collection as a non-invasive method for measuring cortisol in the beluga (Delphinapterus leucas). PLoS ONE 2014, 9, e114062. [Google Scholar]
- American Humane. American Humane Association: Humane Conservation Program. Available online: http://humaneconservation.org/app/uploads/2016/05/Certification-Standards-Details-Humane-Conservation.pdf (accessed on 17 December 2021).
- Reeder, D.M.; Kramer, K.M. Stress in free-ranging mammals: Integrating physiology, ecology and natural history. J. Mammal. 2005, 86, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Carrasco, G.A.; Van de Kar, L.D. Neuroendocrine pharmacology of stress. Eur. J. Pharmacol. 2003, 463, 235–272. [Google Scholar] [CrossRef]
- Voigt, K. Endokrines System, 4th ed.; Georg Thieme Verlag: Stuttgart, Germany, 2003. [Google Scholar]
- Mormède, P.; Andanson, S.; Auperin, B.; Beerda, B.; Guemene, D.; Malmkvist, J.; Manteca, X.; Manteuffel, G.; Prunet, P.; van Reenen, C.G.; et al. Exploration of the hypothalamic-pituitary-adrenal function as a tool to evaluate animal welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef]
- Palazzolo, D.L.; Quadri, S.K. The effects of aging on the circadian rhythm of serum cortisol in the dog. Exp. Gerontol. 1987, 22, 379–387. [Google Scholar] [CrossRef]
- Irvine, C.H.G.; Alexander, S.L. Factors affecting the circadian rhythm in plasma cortisol concentrations in the horse. Domest. Anim. Endocrinol. 1994, 11, 227–238. [Google Scholar] [CrossRef]
- De Jong, I.C.; Prelle, I.T.; van de Burgwal, J.A.; Lambooij, E.; Korte, S.M.; Blokhuis, H.J.; Koolhaas, J.M. Effects of environmental enrichment on behavioral responses to novelty, learning, and memory, and the circadian rhythm in cortisol in growing pigs. Physiol. Amp. Behav. 2000, 68, 571–578. [Google Scholar] [CrossRef]
- Van Eekelen, A.P.J.; Kerhof, G.A.; Amsterdam, J.G.C. Circadian variation in cortisol reactivity to an acute stressor. Chronobiol. Int. J. Biol. Med. Rhythm. Res. 2003, 20, 863. [Google Scholar] [CrossRef] [PubMed]
- Sage, D.; Ganem, J.; Guillaumond, F.; Laforge-Anglade, G.; François-Bellan, A.; Bosler, O.; Becquet, D. Influence of the corticosterone rhythm on photic entrainment of locomotor activity in rats. J. Biol. Rhythm. 2004, 19, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, L.A.W.; Pévet, P.; Saboureau, M.; Sicard, B.; Nesme, B.; Claustrat, B.; Buijs, R.M.; Kalsbeek, A. Temporal organization of the 24-h corticosterone rhythm in the diurnal murid rodent arvicanthis ansorgei thomas 1910. Brain Res. 2004, 995, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Assenza, A.; Fazio, F.; Marceno, G.; Piccione, G.; Caola, G. Daily rhythms of serum and salivary parameters in goats. Aust. Vet. J. 2009, 87, 397–401. [Google Scholar] [CrossRef]
- Refinetti, R. Ciracadian Physiology, 2nd ed.; Taylor & Francis: Boca Raton, FL, USA, 2006; pp. 105–216. [Google Scholar]
- Amaral, R.S. Use of alternative matrices to monitor steroid hormones in aquatic mammals: A review. Aquat. Mamm. 2010, 36, 162–171. [Google Scholar] [CrossRef]
- Säkkinen, H.; Tornbeg, J.; Goddard, P.J.; Eloranta, E.; Ropstad, E.; Saarela, S. The effect of blood sampling method on indicators of physiological stress in reindeer (Rangifer tarandus tarandus). Domest. Anim. Endocrinol. 2004, 26, 87–98. [Google Scholar] [CrossRef]
- Higham, J.P.; Vitale, A.B.; Rivera, A.M.; Ayala, J.E.; Maestripieri, D. Measuring salivary analytes from free-ranging monkeys. Physiol. Amp. Behav. 2010, 101, 601–607. [Google Scholar] [CrossRef] [Green Version]
- Erhard, M. Physiologie und Tierschutz. In Physiologie der Haustiere; von Engelhardt, W., Ed.; Enke: Stuttgart, Germany, 2010; p. 712. [Google Scholar]
- Romano, T.A.; Keogh, M.J.; Kelly, C.; Feng, P.; Berk, L.; Schlundt, C.E.; Carder, D.A.; Finneran, J.J. Anthropogenic sound and marine mammal health: Measures of the nervous and immune systems before and after intense sound exposure. Can. J. Fish. Aquat. Sci. 2004, 61, 1124–1134. [Google Scholar] [CrossRef]
- Galligan, T.M.; Schwacke, L.H.; Houser, D.S.; Wells, R.S.; Rowles, T.; Boggs, A.S.P. Characterization of circulating steroid hormone profiles in the bottlenose dolphin (Tursiops truncatus) by liquid chromatography-tandem mass spectrometry (lc-ms/ms). Gen. Comp. Endocrinol. 2018, 263, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Tobayama, T.; Katsumata, E.; Uchida, S.; Ueda, K.; Yoshioka, M.; Aida, K. Secretory patterns of cortisol in indo-pacific bottlenose dolphins and killer whales. Fish. Sci. 2002, 68, 451–452. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Tobayama, T.; Katsumata, E.; Yoshioka, M.; Aida, K. Serum cortisol levels in captive killer whale and bottlenose dolphin. Fish. Sci. 1998, 64, 643–647. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Uchida, S.; Ueda, K.; Tobayama, T.; Katsumata, E.; Yoshioka, M.; Aida, K. Diurnal and annual changes in serum cortisol concentrations in indo-pacific bottlenose dolphins Tursiops aduncus and killer whales orcinus orca. Gen. Comp. Endocrinol. 2003, 132, 427–433. [Google Scholar] [CrossRef]
- Bertelsmann, H. Haltungsbedingungen, Soziale Beziehungen und Streß Bei Großen Tümmlern (Tursiops truncatus) in Delphinarien. Ph.D. Thesis, Universität Bayreuth, Bayreuth, Germany, 1999. [Google Scholar]
- Creel, S.; Fox, J.E.; Hardy, A.; Sands, J.; Garrott, B.; Peterson, R.O. Snowmobile activity and glucocorticoid stress responses in wolves and elk actividad de vehículos para nieve y respuestas de stress glucocorticoide en lobos y alces. Conserv. Biol. 2002, 16, 809–814. [Google Scholar] [CrossRef]
- Creel, S. Dominance, aggression and glucocorticoid levels in social carnivores. J. Mammal. 2005, 96, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Palme, R.; Rettenbacher, S.; Touma, C.; El-Bahr, S.M.; Mostl, E. Stress hormones in mammals and birds—comparative aspects regarding metabolism, excretion, and noninvasive measurement in fecal samples. In Trends in Comparative Endocrinology and Neurobiology; Vaudry, H., Roubos, E., Schoofs, L., Fiik, G., Larhammar, D., Eds.; New York Acad Sciences: New York, NY, USA, 2005; Volume 1040, pp. 162–171. [Google Scholar]
- Howell-Stephens, J.A.; Brown, J.S.; Bernier, D.; Mulkerin, D.; Santymire, R.M. Characterizing adrenocortical activity in zoo-housed southern three-banded armadillos (Tolypeutes matacus). Gen. Comp. Endocrinol. 2012, 178, 64–74. [Google Scholar] [CrossRef]
- Touma, C.; Palme, R. Measuring fecal glucocorticoid metabolites in mammals and birds: The importance of validation. Ann. N. Y. Acad. Sci. 2005, 1046, 54–74. [Google Scholar] [CrossRef] [PubMed]
- Hein, A.; Palme, R.; Baumgartner, K.; von Fersen, L.; Woelfing, B.; Greenwood, A.D.; Bechshoft, T.; Siebert, U. Faecal glucocorticoid metabolites as a measure of adrenocortical activity in polar bears (Ursus maritimus). Conserv. Physiol. 2020, 8, coaa012. [Google Scholar] [CrossRef]
- Kalliokoski, O.; Timm, J.A.; Ibsen, I.B.; Hau, J.; Frederiksen, A.M.B.; Bertelsen, M.F. Fecal glucocorticoid response to environmental stressors in green iguanas (Iguana iguana). Gen. Comp. Endocrinol. 2012, 177, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Stalder, T.; Kirschbaum, C. Analysis of cortisol in hair—state of the art and future directions. Brain Behav. Immun. 2012, 26, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Cattet, M.; Macbeth, B.J.; Janz, D.M.; Zedrosser, A.; Swenson, J.E.; Dumond, M.; Stenhouse, G.B. Quantifying long-term stress in brown bears with the hair cortisol concentration: A biomarker that may be confounded by rapid changes in response to capture and handling. Conserv. Physiol. 2014, 2, cou026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastromonaco, G.F.; Gunn, K.; McCurdy-Adams, H.; Edwards, D.B.; Schulte-Hostedde, A.I. Validation and use of hair cortisol as a measure of chronic stress in eastern chipmunks (Tamias striatus). Conserv. Physiol. 2014, 2, cou005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakotoniaina, J.H.; Kappeler, P.M.; Kaesler, E.; Hämäläinen, A.M.; Kirschbaum, C.; Kraus, C. Hair cortisol concentrations correlate negatively with survival in a wild primate population. BMC Ecol. 2017, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Reese, L.; Baumgartner, K.; von Fersen, L.; Merle, R.; Ladwig-Wiegard, M.; Will, H.; Haase, G.; Tallo-Parra, O.; Carbajal, A.; Lopez-Bejar, M. Feather corticosterone measurements of greater flamingos living under different forms of flight restraint. Animals 2020, 10, 605. [Google Scholar] [CrossRef] [Green Version]
- Jenni-Eiermann, S.; Helfenstein, F.; Vallat, A.; Glauser, G.; Jenni, L. Corticosterone: Effects on feather quality and deposition into feathers. Methods Ecol. Evol. 2015, 6, 237–246. [Google Scholar] [CrossRef]
- Biancani, B.; Da Dalt, L.; Lacave, G.; Romagnoli, S.; Gabai, G. Measuring fecal progestogens as a tool to monitor reproductive activity in captive female bottlenose dolphins (Tursiops truncatus). Theriogenology 2009, 72, 1282–1292. [Google Scholar] [CrossRef]
- Mercera, K. Le Dosage du Cortisol Fécal: Faisabilité, Intérêts et Limites Dans L’étude du Bien-être du Grand Dauphin (Tursiops truncatus). Ph.D. Thesis, École nationale vétérinaire d’Alfort, Alfort, Sweden, 2019. [Google Scholar]
- Biancani, B.; Dalt, L.D.; Gallina, G.; Capolongo, F.; Gabai, G. Fecal cortisol radioimmunoassay to monitor adrenal gland activity in the bottlenose dolphin (Tursiops truncatus) under human care. Mar. Mammal Sci. 2017, 33, 1014–1034. [Google Scholar] [CrossRef]
- Miller, L.J.; Lauderdale, L.K.; Walsh, M.T.; Bryant, J.L.; Mitchell, K.A.; Granger, D.A.; Mellen, J.D. Reference intervals and values for fecal cortisol, aldosterone, and the ratio of cortisol to dehydroepiandrosterone metabolites in four species of cetaceans. PLoS ONE 2021, 16, e0250331. [Google Scholar]
- Muraco, H.; Cheng, A.; Ravida, N.; Arn, D.; Hudson, J.; Durrant, B. A new approach to detection of luteinizing hormone in a bottlenose dolphin (Tursiops truncatus). Aquat. Mamm. 2009, 35, 386. [Google Scholar] [CrossRef]
- Lettrichová, M.I. Saliva as Diagnostic Fluid: Its Applicability to Noninvasive Evaluation of Oxidative Stress Status and Ovulation Detection. Ph.D. Thesis, Comenuis University, Bratislava, Slovakia, 2015. [Google Scholar]
- Humphrey, S.P.; Williamson, R.T. A review of saliva: Normal composition, flow, and function. J. Prosthet. Dent. 2001, 85, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Veerman, E.; Van den Keybus, P.; Vissink, A.; Amerongen, A.N. Human glandular salivas: Their separate collection and analysis. Eur. J. Oral Sci. 1996, 104, 346–352. [Google Scholar] [CrossRef]
- Vincent, I.C.; Michell, A.R. Comparison of cortisol concentrations in saliva and plasma of dogs. Res. Vet. Sci. 1992, 53, 342–345. [Google Scholar] [CrossRef]
- Beerda, B.; Schilder, M.; Janssen, N.; Mol, J.A. The use of saliva cortisol, urinary cortisol, and catecholamine measurements for a noninvasive assessment of stress responses in dogs. Horm. Behav. 1996, 30, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Pedernera-Romano, C.; Valdez, R.A.; Singh, S.; Chiappa, X.; Romano, M.C.; Galindo, F. Salivary cortisol in captive dolphins (Tursiops truncatus): A non-invasive technique. Anim. Welf. 2006, 15, 359–362. [Google Scholar]
- Cook, N.J. Review: Minimally invasive sampling media and the measurement of corticosteroids as biomarkers of stress in animals. Can. J. Anim. Sci. 2012, 92, 227–259. [Google Scholar] [CrossRef]
- Ugaz, C.; Valdez, R.A.; Romano, M.C.; Galindo, F. Behavior and salivary cortisol of captive dolphins (Tursiops truncatus) kept in open and closed facilities. J. Vet. Behav. Clin. Appl. Res. 2013, 8, 285–290. [Google Scholar] [CrossRef]
- Monreal-Pawlowsky, T.; Carbajal, A.; Tallo-Parra, O.; Sabés-Alsina, M.; Monclús, L.; Almunia, J.; Fernández-Bellon, H.; Lopez-Bejar, M. Daily salivary cortisol levels in response to stress factors in captive common bottlenose dolphins (Tursiops truncatus): A potential welfare indicator. Vet. Rec. 2017, 180, 593. [Google Scholar] [CrossRef] [PubMed]
- Bonga, S.E.W. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Campbell, M.D.; Patino, R.; Tolan, J.; Strauss, R.; Diamond, S.L. Sublethal effects of catch-and-release fishing: Measuring capture stress, fish impairment, and predation risk using a condition index. ICES J. Mar. Sci. 2009, 67, 513–521. [Google Scholar] [CrossRef]
- Sadoul, B.; Geffroy, B. Measuring cortisol, the major stress hormone in fishes. J. Fish Biol. 2019, 94, 540–555. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Stalder, T.; Kirschbaum, C. Quantitative analysis of estradiol and six other steroid hormones in human saliva using a high throughput liquid chromatography–tandem mass spectrometry assay. Talanta 2015, 143, 353–358. [Google Scholar] [CrossRef]
- Pankhurst, N.; Sharples, D. Effects of capture and confinement on plasma cortisol concentrations in the snapper, Pagrus auratus. Mar. Freshw. Res. 1992, 43, 345–355. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Reidarson, T.H.; McBain, J.F. Hematologic, biochemical, and endocrine effects of dexamethasone on bottlenose dolphins (Tursiops truncatus). J. Zoo Wildl. Med. 1999, 30, 310–312. [Google Scholar]
- Waples, K.A.; Gales, N.J. Evaluating and minimising social stress in the care of captive bottlenose dolphins (Tursiops aduncus). Zoo Biol. 2002, 21, 5–26. [Google Scholar] [CrossRef]
- Judd, H.L.; Ridgway, S. Twenty-four hour patterns of circulating androgens and cortisol in male dolphins. In Breeding Dolphin: Present Status, Suggestion for the Future; National Technical Information Service: Springfield, VA, USA, 1977; pp. 269–277. [Google Scholar]
- Pfefferle, D.; Plümer, S.; Burchardt, L.; Treue, S.; Gail, A. Assessment of stress responses in rhesus macaques (Macaca mulatta) to daily routine procedures in system neuroscience based on salivary cortisol concentrations. PLoS ONE 2018, 13, e0190190. [Google Scholar] [CrossRef] [Green Version]
- Behringer, V.; Borchers, C.; Deschner, T.; Möstl, E.; Selzer, D.; Hohmann, G. Measurements of salivary alpha amylase and salivary cortisol in hominoid primates reveal within-species consistency and between-species differences. PLoS ONE 2013, 8, e60773. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, B.P.; Knackfuss, F.B.; Labarthe, N.; Mendes-de-Almeida, F. Effect of a synthetic analogue of the feline facial pheromone on salivary cortisol levels in the domestic cat. Pesqui. Vet. Bras. 2017, 37, 287–290. [Google Scholar]
- Menargues, A.; Urios, V.; Mauri, M. Welfare assessment of captive asian elephants (Elephas maximus) and indian rhinoceros (Rhinoceros unicornis) using salivary cortisol measurement. Anim. Welf. 2008, 17, 305–312. [Google Scholar]
- Peeters, M.; Sulon, J.; Beckers, J.F.; Ledoux, D.; Vandenheede, M. Comparison between blood serum and salivary cortisol concentrations in horses using an adrenocorticotropic hormone challenge. Equine Vet. J. 2011, 43, 487–493. [Google Scholar] [CrossRef]
- Huettner, T.; Dollhaeupl, S.; Simon, R.; Baumgartner, K.; von Fersen, L. Activity budget comparisons using long-term observations of a group of bottlenose dolphins (Tursiops truncatus) under human care: Implications for animal welfare. Animals 2021, 11, 2107. [Google Scholar] [CrossRef]









| Name | Sex | Born | Birth Year | Blood Cortisol | Blood vs. Saliva Cortisol | Feeding Experiment I | Feeding Experiment II |
|---|---|---|---|---|---|---|---|
| Anke | female | wild | ca. 1983 | × | × | × | - |
| Arnie | male | captive | 2000 | × | - | - | - |
| Jenny | female | wild | ca. 1987 | × | × | × | - |
| Moby | male | wild | ca. 1960 | × | × | - | - |
| Noah | male | captive | 2000 | × | - | - | - |
| Rocco | male | captive | 2005 | × | - | - | - |
| Nami | female | captive | 2014 | × | × | - | - |
| Dolly | female | captive | 2007 | × | - | × | × |
| Donna | female | captive | 2007 | × | - | × | × |
| Sunny | female | captive | 1999 | - | - | × | × |
| Joker | male | captive | 1991 | × | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rickert, D.; Simon, R.; von Fersen, L.; Baumgartner, K.; Bertsch, T.; Kirschbaum, C.; Erhard, M. Saliva and Blood Cortisol Measurement in Bottlenose Dolphins (Tursiops truncatus): Methodology, Application, and Limitations. Animals 2022, 12, 22. https://doi.org/10.3390/ani12010022
Rickert D, Simon R, von Fersen L, Baumgartner K, Bertsch T, Kirschbaum C, Erhard M. Saliva and Blood Cortisol Measurement in Bottlenose Dolphins (Tursiops truncatus): Methodology, Application, and Limitations. Animals. 2022; 12(1):22. https://doi.org/10.3390/ani12010022
Chicago/Turabian StyleRickert, Daniela, Ralph Simon, Lorenzo von Fersen, Katrin Baumgartner, Thomas Bertsch, Clemens Kirschbaum, and Michael Erhard. 2022. "Saliva and Blood Cortisol Measurement in Bottlenose Dolphins (Tursiops truncatus): Methodology, Application, and Limitations" Animals 12, no. 1: 22. https://doi.org/10.3390/ani12010022