
Transcription Regulator YgeK Affects the Virulence of Avian Pathogenic Escherichia coli

1
Anhui Province Key Laboratory of Veterinary Pathobiology and Disease Control, College of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, China
2
College of Animal Science and Food Engineering, Jinling Institute Technology, Nanjing 211169, China
*
Authors to whom correspondence should be addressed.
Animals 2021, 11(11), 3018; https://doi.org/10.3390/ani11113018
Submission received: 31 May 2021
/
Revised: 14 October 2021
/
Accepted: 15 October 2021
/
Published: 20 October 2021
(This article belongs to the Special Issue Avian Disease Pathogenesis)
Abstract
:Simple Summary
Avian pathogenic Escherichia coli (APEC) is the responsible pathogen for colibacillosis in poultry. Transcriptional regulator YgeK was a transcriptional regulator locating at E. coli type three secretion system 2 (ETT2) in APEC. However, the role of YgeK in APEC has not been reported. In this study, we found that the inactivation of YgeK in APEC decreased the flagellar formation ability, bacterial motility ability, serum sensitivity, adhesion ability, and virulence. Results suggested that the transcriptional regulator YgeK plays a crucial role in APEC virulence.
Abstract
Avian pathogenic Escherichia coli (APEC) is the responsible pathogen for colibacillosis in poultry, and is a potential gene source for human extraintestinal pathogenic Escherichia coli. Escherichia coli type III secretion system 2 (ETT2) is widely distributed in human and animal ExPEC isolates, and is crucial for the virulence of ExPEC. Transcriptional regulator YgeK, located in the ETT2 gene cluster, was identified as an important regulator of gene expression in enterohemorrhagic E. coli (EHEC). However, the role of YgeK in APEC has not been reported. In this study, we performed amino acid alignment analysis of YgeK among different E. coli strains and generated ygeK mutant strain AE81ΔygeK from clinical APEC strain AE81. Flagellar formation, bacterial motility, serum sensitivity, adhesion, and virulence were all significantly reduced following the inactivation of YgeK in APEC. Then, we performed transcriptome sequencing to analyze the functional pathways involved in the biological processes. Results suggested that ETT2 transcriptional regulator YgeK plays a crucial role in APEC virulence. These findings thus contribute to our understanding of the function of the ETT2 cluster, and clarify the pathogenic mechanism of APEC.
1. Introduction
Extraintestinal pathogenic Escherichia coli (ExPEC) is an important zoonotic pathogen that can cause serious extraintestinal infections in humans and animals [1]. ExPEC can be divided into avian pathogenic E. coli (APEC), uropathogenic E. coli (UPEC) and new meningitis E. coli (NMEC) according to the affected host and organs [2,3,4,5]. APEC is the leading cause of avian colibacillosis, and one of the major causes of morbidity, mortality, and economic loss in the global poultry industry [6]. Known virulence factors of APEC are adhesin, toxin, iron-acquisition factor, lipopolysaccharide, and invasion factors ibeA, ibeB, and ibeC [7,8,9]. Several studies showed that APEC and human ExPEC share some virulence genes, and suggest that APEC may act as a virulence gene reservoir for ExPEC, presenting a threat to public health [3,10,11]. However, the pathogenic mechanism of APEC is still unknown, and difficulties still exist in the prevention and treatment of avian colibacillosis. Therefore, it is necessary to study the pathogenic mechanism of APEC to promote to development of effective vaccines or look for new potential drug targets to control APEC.
Protein secretion systems play important roles in prokaryotes, including in the defense against biological and chemical agents, and facilitating disease [12]. The Type III secretion system (T3SS) is the most common secretion system, and it is used to subvert eukaryotic signaling pathways in Gram-negative bacteria, such as Yersinia species, Salmonella enteria serovars, Shigella species, and Escherichia species [13,14]. E. coli type III secretion system 2 (ETT2) was first identified in enterohaemorrhagic E. coli O157: H7. The prevalence of the ETT2 cluster in APEC was higher than that in human ExPEC (UPEC and NMEC) [15,16,17]. The intact ETT2 locus is 29.9 kb with 35 genes, including yqe (ecs3703–3706), yge (ecs3707–3712), epr (ecs3716–3719), etr (ecs3720), epa (ecs3721–3726), and eiv (ecs3727–3737) [18]; however, not all 35 genes exist in clinical E. coli isolates. Though the presence of ETT2 is often accompanied by extensive deletions and insertions in clinical E. coli isolates, some studies showed that the ETT2 cluster or the genes within the cluster were vital for virulence in ExPEC strains, such as the ETT2 cluster in UPEC, ETT2 ATPase EivC in APEC, and ETT2 transcriptional regulator YqeI in APEC [19,20,21].
YgeK, which encodes a transcription regulator, is located in the ETT2 cluster. It was identified as an important regulator of gene expression in EHEC [22]. However, the function of transcriptional regulator YgeK in APEC remains unknown. In this study, we evaluated the role of YgeK in a clinical APEC isolate with an intact ETT2 cluster. First, we carried out a BLAST search of the YgeK amino acid sequence in the clinical APEC isolate AE81, E. coli K12, and E. coli O157: H7. We then constructed ygeK mutant strain AE81ΔygeK and determined the impact of inactivation of YgeK on APEC characteristics. We also analyzed the affected pathways by transcriptional regulator YgeK by transcriptome sequencing. We confirmed that transcription regulator YgeK played an important role in APEC, thus providing new clues for future research into the pathogenic mechanism of APEC.
2. Results
2.1. Amino Acid Sequence of YgeK in AE81 Differed from That in E. coli K12
A schematic diagram of the ETT2 cluster is shown in Figure 1a, according to the AE81 sequence. Full-length YgeK in AE81 was composed by 210 amino acids. The amino acid sequence was the same in E. coli O157: H7 except for the 66th amino acid, which was serine (S) in AE81 and alanine (A) in E. coli O157: H7. However, the sequence in E. coli K12 was missing 189 bp (63 amino acids) at the 5′ end compared with the sequences in AE81 and E. coli O157: H7 (Figure 1b).
2.2. Inactivation of YgeK Influenced Morphological Structure and Motility of AE81
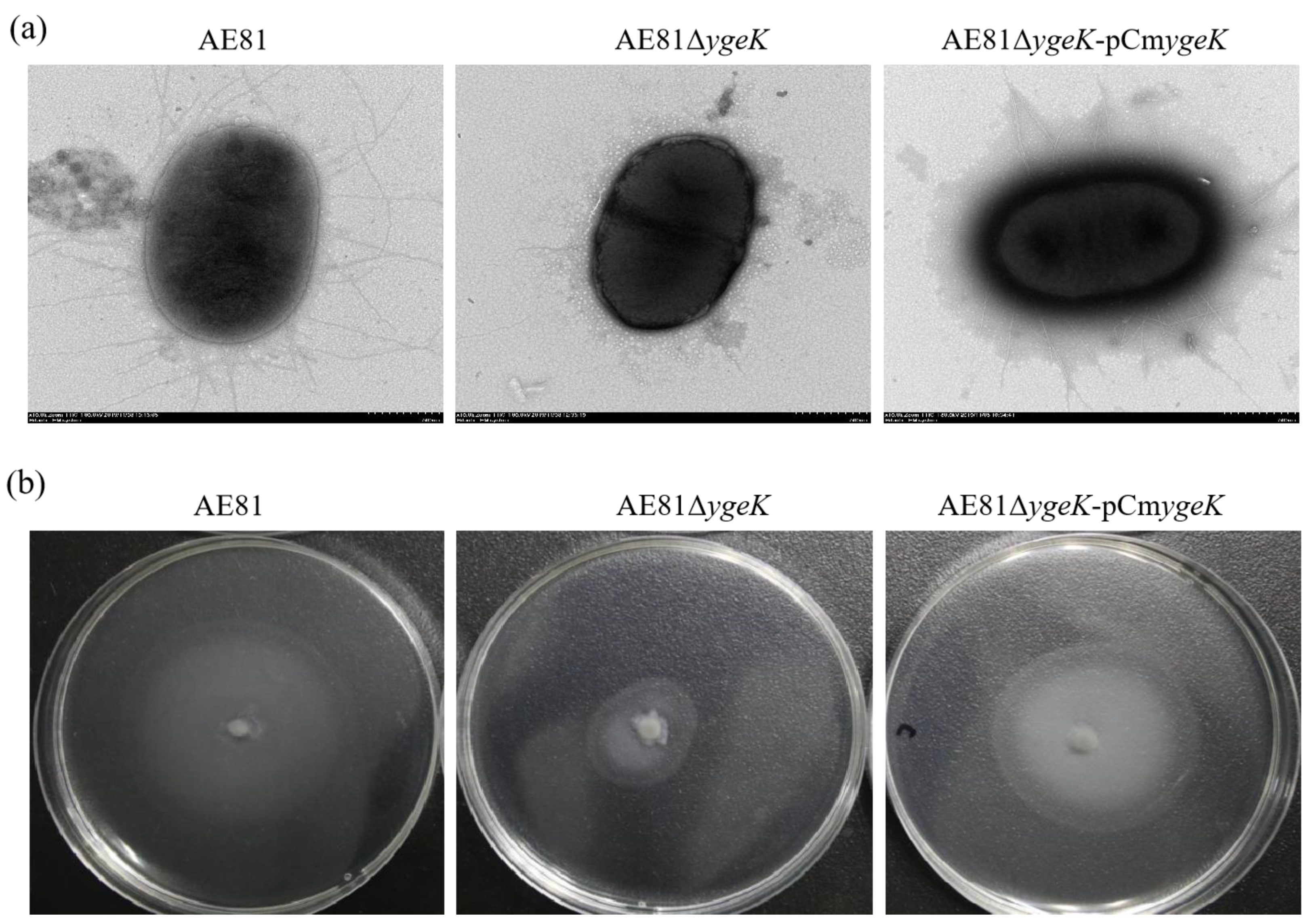
Mutant strain AE81ΔygeK and complemented strain AE81ΔygeK-pCmygeK were successfully constructed, and AE81ΔygeK also showed a normal growth rate lysogeny broth (Figure S1). However, there were notable differences in morphological structure between AE81 and AE81ΔygeK. Under transmission electron microscopy, there were long curved flagella covering the surface in AE81. In contrast, flagellar production was impaired in mutant strain AE81ΔygeK, and only one flagellum was observed on the surface of AE81ΔygeK. Flagellar formation was partially recovered in the complemented strain, and a few broken flagella appeared on the surface of AE81ΔygeK-pCmygeK (Figure 2a). In addition, after incubation on semisolid LB agar plates for 8 h at 37 °C, the swarming circle of AE81ΔygeK was much smaller than that of AE81 and AE81ΔygeK-pCmygeK, but the wild-type phenotype was restored in AE81ΔygeK-pCmygeK, indicating that transcription regulator YgeK upregulates the motility of AE81 (Figure 2b).
2.3. YgeK Played an Important Role in AE81 Serum Resistance
AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK showed similar survival abilities in PBS and 10% SPF chicken serum. After incubation with 20%, 30%, 40%, and 50% serum, the survival capacity of AE81ΔygeK was significantly lower than that of AE81 (** p < 0.01, *** p < 0.001). The bactericidal survival activity of AE81ΔygeK-pCmygeK was intermediate between that of AE81 and AE81ΔygeK-pCmygeK (Figure 3), showing that YgeK was crucial for serum resistance in AE81. Results of plasmid control are shown in Figure S2.
2.4. YgeK Upregulated APEC Capacity to Adhere to DF-1 Cells
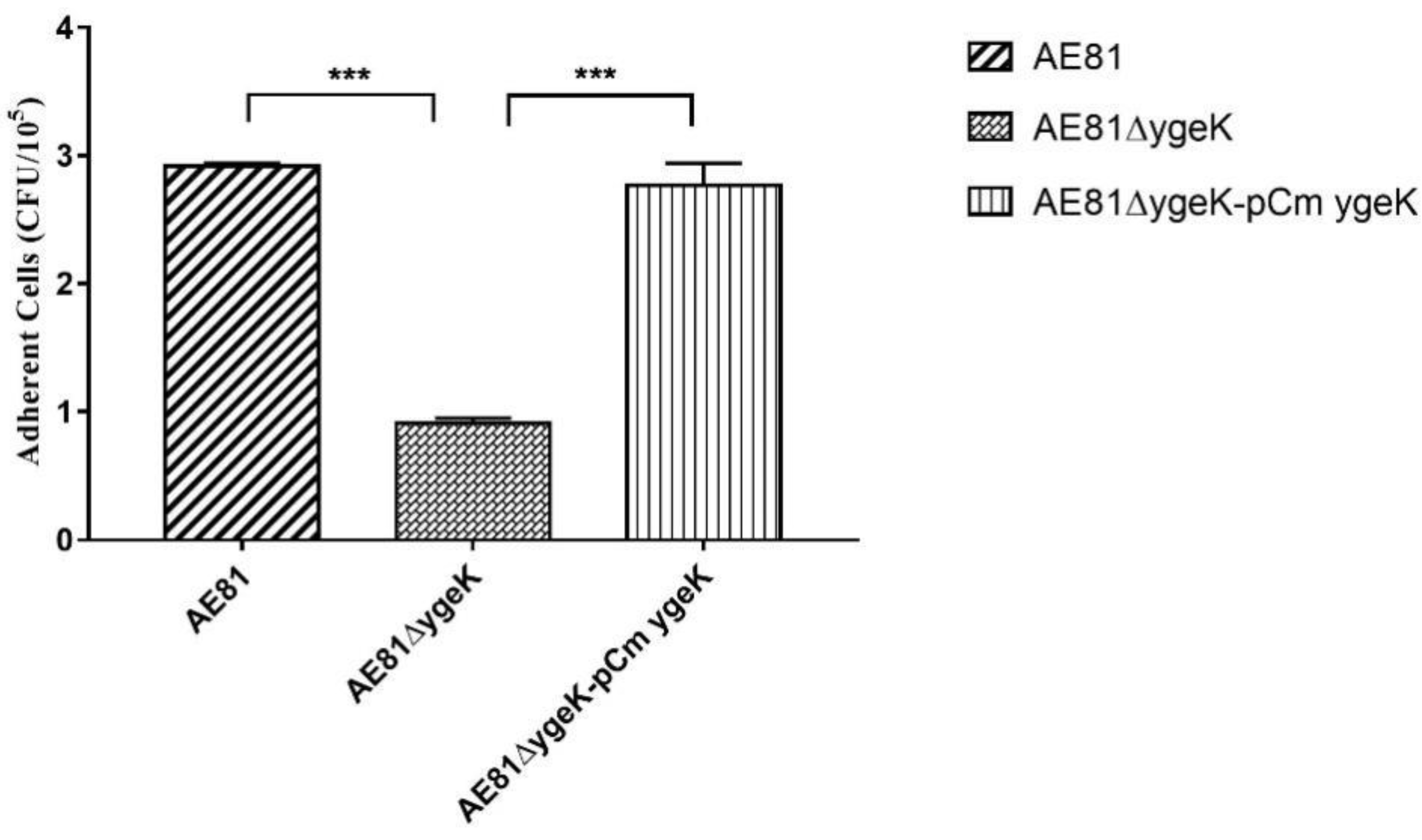
We investigated the role of YgeK in bacterial adhesion by infecting DF-1 chicken fibroblasts with AE81, AE81ΔygeK, or AE81ΔygeK-pCmygeK. AE81ΔygeK showed decreased adhesion to DF-1 cells compared with AE81 and AE81ΔygeK-pCmygeK (Figure 4). Results of plasmid control are shown in Figure S3. These results suggested that YgeK upregulated the capacity of APEC to adhere to DF-1 cells.
2.5. Transcriptional Profiling of AE81 and AE81ΔygeK
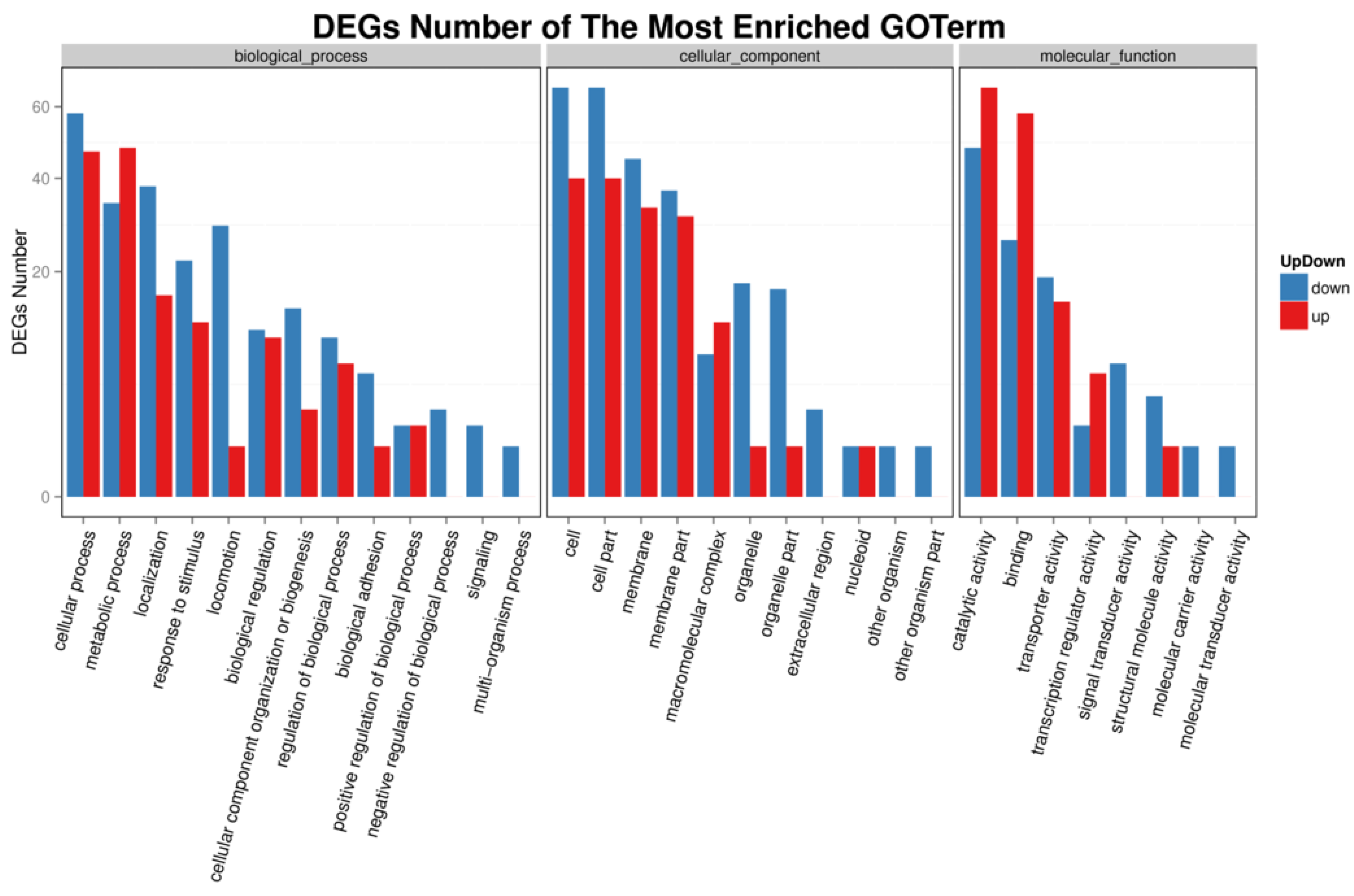
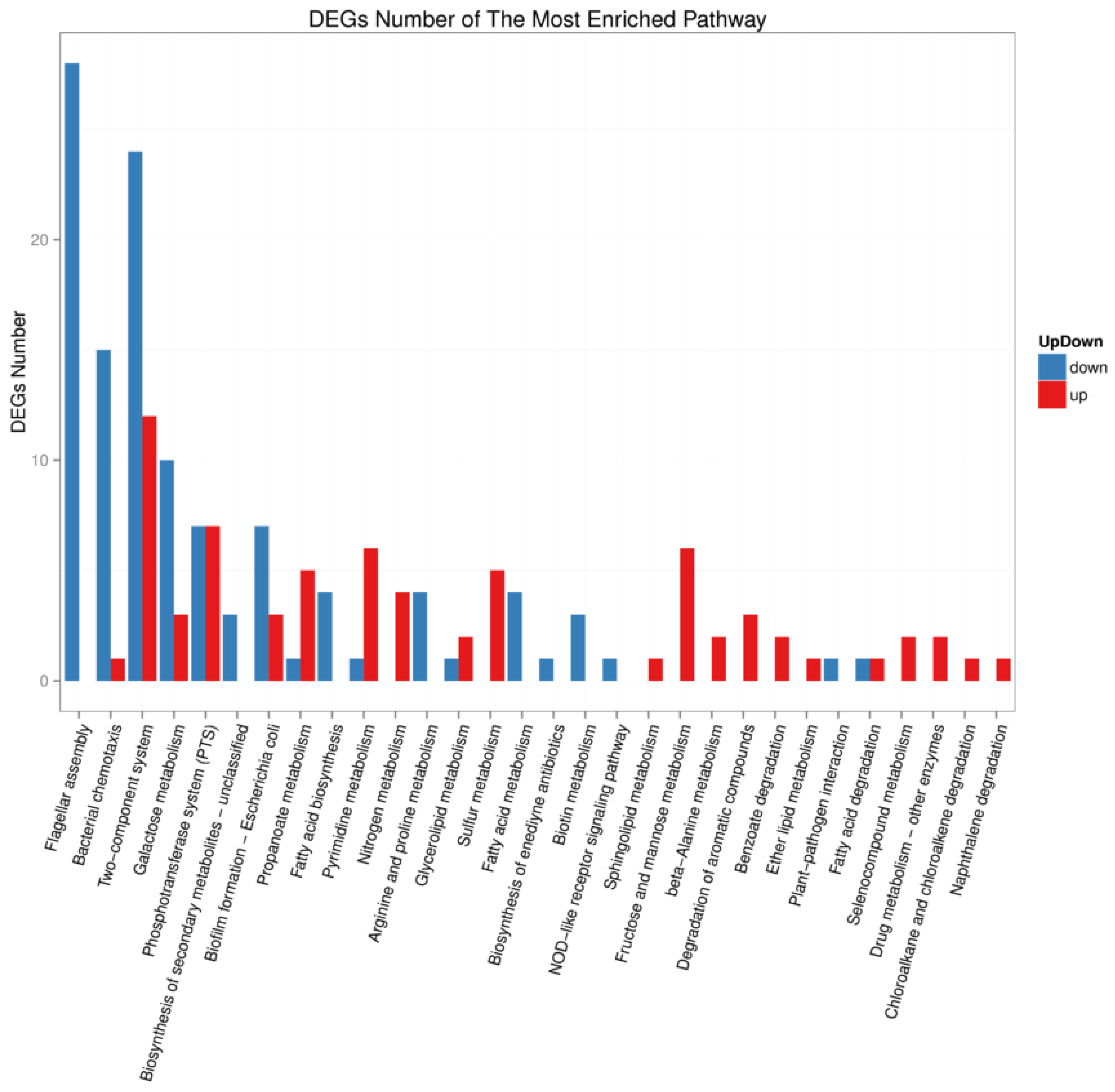
We investigated the functional pathways affected by transcriptional regulator YgeK by transcriptome sequencing. A total of 4028 genes were quantified, and a total of 275 gene transcripts were defined as differentially expressed genes (DEGs) after deleting YgeK, including 137 upregulated and 138 downregulated DEGs (p ≤ 0.05, fold change ≥2.0) (Table S1). The most enriched Gene Ontology (GO) terms of DEGs were cellular process, metabolic process, and localization in biological process, cell, cell part and membrane in cellular component, catalytic activity, binding, and transporter activity in molecular function (Figure 5). The most enriched DEGs pathways included flagellar assembly, bacterial chemotaxis, two-component system, and biological adhesion (Figure 6).
2.6. YgeK Upregulated Flagellar and Virulence-Associated Genes
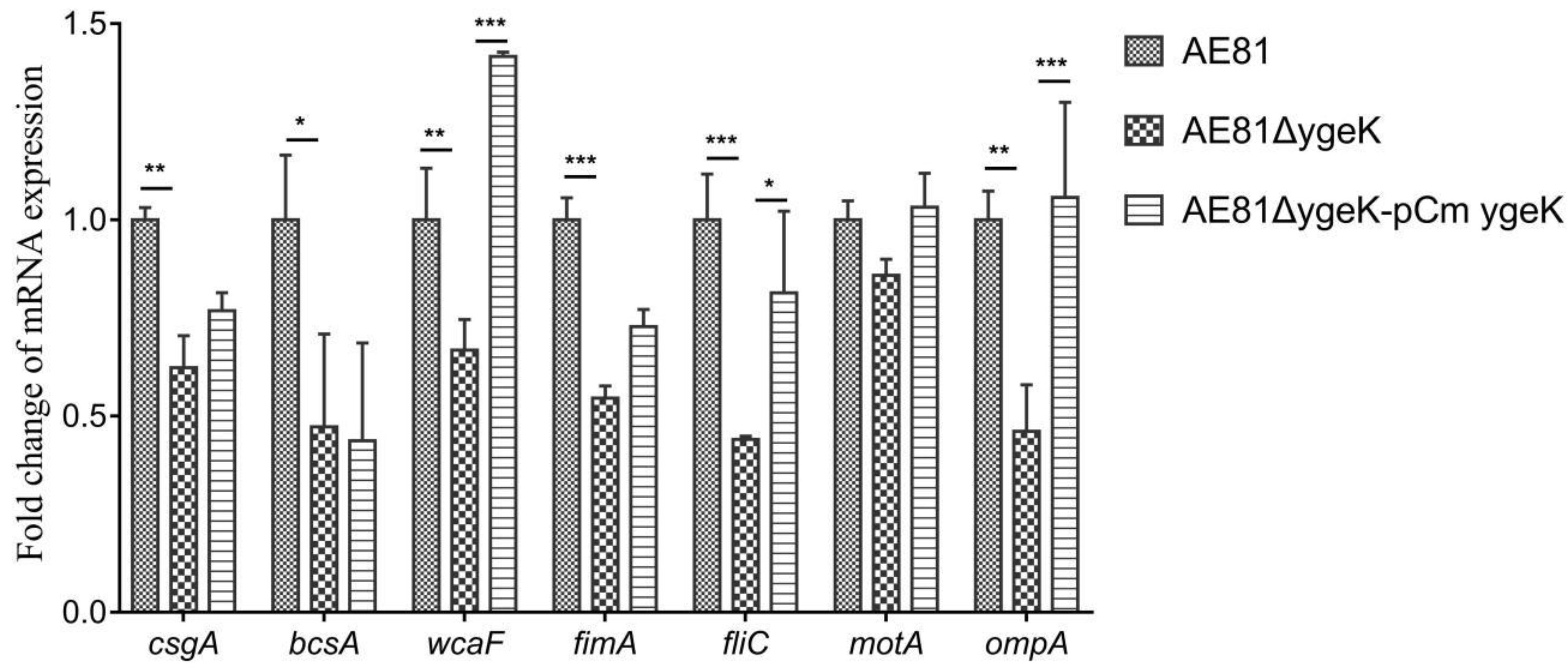
We determined the transcript levels of biofilm-associated genes csgA, bcsA, and wcaF, flagellar genes fliC and motA, fimbriae gene fimA, and virulence gene ompA in AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK by qRT-PCR. There were no changes in the transcript levels of motA in AE81ΔygeK. The transcript levels of csgA, bcsA, wcaF, fliC, fimA, and ompA were significantly decreased in AE81ΔygeK compared with AE81, and were restored in AE81ΔygeK-pCmygeK (Figure 7). These results confirmed that YgeK significantly upregulated the transcription of several flagellar and virulence-associated genes.
3. Discussion
Due to its presence in nonpathogenic E. coli K12 and pathogenic E. coli, whether ETT2 cluster can be used as a marker for detecting bacterial virulence is still inconclusive. However, E. coli K-12 did not possess an intact ETT2 cluster, and an intact ETT2 cluster or the genes encoded by the cluster affect the pathogenicity of ExPEC [19,20,23]. Transcriptional regulator YgeK is considered to be a pseudogene in E. coli K12, but it affects virulence in E. coli O157: H7. In this study, we analyzed the amino acid sequence of YgeK in different E. coli strains and found that the amino acid sequence of YgeK in AE81 was the same as that in E. coli O157: H7 except for one base difference, but both were significantly different from the sequence in E. coli K12. Each protein has a characteristic and unique amino acid sequence. The unique sequence or order of amino acids dictates the 3D conformation that the folded protein has, and this conformation determines the function of the protein. Thus, the similarity of the amino acid sequence of AE81 and E. coli O157: H7 provides the possibility of similar functions.
As expected, the inactivation of transcriptional regulator YgeK in AE81 led to a reduction in many bacterial functions, including flagellar formation, motility, and bactericidal activity, and adhesion ability. Flagella are more locomotive organelles in E. coli, and critical for biofilm formation and bacterial virulence [24,25]. Several studies revealed that transcriptional regulators YqeI and ATPase EivC both influence APEC’s flagella and motility [20,21], and this study supplements another transcriptional regulator in the ETT2 cluster that has an effect on APEC’s flagella and motility. Serum resistance is a prerequisite for septicemia, which is a typical symptom of avian colibacillosis [26]. The ETT2 cluster or the genes located at the ETT2 cluster are involved in serum survival of septicemia E. coli, NMEC, APEC and UPEC [19,20,27,28]. Deletion of YgeK attenuated serum resistance, supporting a vital role of the ETT2 cluster in serum resistance. In addition, adhesion to the host cells is an initial and important step in APEC pathogenesis. Meningitis-causing E. coli strain K1 with mutant ETT2 exhibited defects in invasion and intracellular survival compared with the parental strain. Yao et al. (2009) indicated that ETT2 was necessary for the pathogenic interaction between the E. coli K1 strain and host cells [28]. In this study, the inactivation of YgeK reduced the adhesion of APEC to DF-1 cells.
Since transcription regulator YgeK significantly influences APEC characteristics, we detected the effect of YgeK on the transcriptional level of other genes using transcriptome sequencing. Transcriptional analysis revealed that the top three most enriched pathways were flagellar assembly, bacterial chemotaxis, and two-component system. The expression of differently expressed flagellar genes was all down, which explained the differences in flagellar formation and motility between AE81 and AE81ΔygeK. The motility and transcriptional level of flagellar genes in UPEC changed after deleting either the intact ETT2 or just part of this cluster [19].
A variety of transcription units in APEC have been reported, and an intricate regulatory network connecting various APEC virulence factors exists in APEC. These reported genes include global transcriptional regulators and two-component systems. They often affect APEC motility, biofilm formation, or other characteristics. For example, YjjQ, a global transcriptional regulator in APEC, affects APEC’s virulence through iron uptake [29]; McbR, acts as a transcriptional regulator involved in biofilm formation and the H2O2 stress response [30]; KdpDE, BarA-UvrY, and PhoPQ, as two-component systems in APEC, are all involved in APEC’s virulence [31,32,33]. There is a connection between different regulatory units. For example, phosphorylated CpxR directly regulates the expression of a Type VI secretion system (T6SS2) and promotes the pathogenicity of APEC by binding to the hcp2B promoter region of T6SS2 [34]. The functional description of transcriptional regulator YgeK contributes to the analysis of APEC pathogenicity.
4. Materials and Methods
4.1. Amino Acid Analysis of YgeK among Different E. coli Strains
The sequence of the ETT2 cluster and YgeK in AE81 was obtained from whole-genome sequencing [21]. The whole-genome sequencing library was prepared by Novogene Co., Ltd. Through the quality control of the Novogene pipeline, raw data weree filtered to obtain the clean data. SPAdes 3.10.2 and Velvet 1.2.10 software was used to splice the preprocessed sequence fragments and assemble them into a contig containing the information of the entire bacterial genome. The YgeK sequences of E. coli K12 and E. coli O157: H7 were downloaded from National Centre for Biotechnology Information (NCBI). Homology including the deleted sequence was calculated. The multiple sequence alignment among amino acid sequences was estimated with Molecular Evolutionary Genetics Analysis Version 7.0 (Mega 7.0).
4.2. Construction and Verification of the ygeK Mutant Strain
The mutant strain was constructed using the lambda Red recombinase system [35]. According to the sequence of YgeK in AE81, two long homology arm primers were designed. The chloramphenicol resistant fragment was amplified with the pKD3 plasmid as the template, gel-recovered, and transformed into AE81 cells with pKD46 plasmids using Gene Pulser Xcell with 200 Ω and 2.5 kV (Bio Rad, Hercules, CA, USA). Mutant strain AE81ΔygeK was identified by PCR and sequencing. The pCP20 plasmid was used to cure pKD3, and the mutant strain was named AE81ΔygeK. Strains and the plasmids are listed in Table 1, and primers are listed in Table S2.
4.3. Construction and Verification of Complemented Strain
The complemented fragments were amplified using the AE81 genome as a template with primers C-BamHI-ygeK-f and C-HindIII-ygeK-r. The fragments and plasmid pSTV28 DNA were digested with BamHI and HindIII, recovered by glue, and ligated with T4 ligase. Complemented strain AE81ΔygeK-pCmygeK was generated by cloning the target genes into plasmid pSTV28 and transforming the resulting plasmid into AE81ΔygeK cells. Strains were grown in lysogeny broth (LB) medium (tryptone, 1.0 g; NaCl, 1.0 g; yeast extract, 0.5 g with 100 mL ultrapure water; when necessary, medium was supplemented with 1.5 g agar) with chloramphenicol (Cm, 30 µg/mL) at 37 °C with biochemical incubator (Boxun, Shanghai, China).
4.4. Bacterial Growth Curves
The overnight cultures of AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK were diluted to an optical density at 600 nm (OD600) of approximately 0.03 in LB medium. Bacteria were cultured at 37 °C with 150 rpm and each hour to monitor optimal density using a UV/Vis spectrophotometer (DU730, Beckman Coulter, Miami, FL, USA).
4.5. Micromorphology Observation by Transmission Electron Microscopy
To observe the micromorphology of AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK, they were stationary cultured overnight at 37 °C in LB and then washed with phosphate buffer saline (PBS) three times. The bacteria were placed on a 200 mesh formvar-coated copper microscopy grid, incubated at room temperature, stained with 2% aqueous uranyl acetate for 30 s, and dried. The micromorphology of AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK was observed using transmission electron microscopy (HitachiHT-7700, Tokyo, Japan).
4.6. Motility Assay
The motility ability of AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK was determined with semisolid medium (1.0 g tryptone, 1.0 g NaCl, 0.5 g yeast extract, 0.25 g agar with 100 mL ultrapure water). Stationary bacterial cultures were diluted 1:100 into a fresh LB medium and then cultured overnight. After culturing overnight, bacteria were washed with PBS three times and concentrated (OD600 = 2.0). The concentrated bacteria were dripped on a 0.25% semisolid medium and incubated for 8 h at 37 °C. Motility was assessed by examining the radius of the bacterial migration of the plate [21].
4.7. Serum Bactericidal Assay
Overnight cultures were diluted 1:25 into LB medium and cultured to the logarithmic phase. Bacteria were incubated with 10%, 20%, 30%, 40%, and 50% diluted specific-pathogen-free (SPF) chicken serum (Gibco, Grand Island, NY, USA) or heat-inactivated serum at 37 °C [19]. Growth was determined using FilterMax F3 Multi-Mode Microplate Reader (Molecular Devices, CA, USA), and optical density at 600 nm (OD600) was measured each hour.
4.8. Bacterial Adhesion Assay
Chicken embryo fibroblast DF-1 cell monolayers were washed with Dulbecco’s modified Eagle’s medium (DMEM) without fetal bovine serum, and infected with bacteria at a multiplicity of infection (MOI) of 100 for 2 h at 37 °C under 5% CO2. After washing with PBS, cells were lysed with 0.5% Triton X-100, and bacteria were counted by plating on LB agar plates. Assay was performed three times.
4.9. RNA Extraction and Library Preparation for Transcriptome Sequencing
Total RNA was extracted from AE81 and AE81ΔygeK, and RNA was purified and fragmented. Total RNA was examined by a NanoDrop nd-2000 spectrophotometer and Agilent Bioanalyzer 2100 (Agilent Technologies, Santa Clara, CA, USA). Sequencing libraries were generated following the manufacturer’s recommendations. Library quality was assessed, and the constructed library was sequenced using the Illumina HiSeq sequencing platform.
4.10. Differential Expression Analysis
Reads mapped to each gene were counted using HTSeq. The reads per kilobase of exon model per million mapped reads (FPKM) of each gene were calculated on the basis of gene length and the number of mapped reads. Resulting p values were adjusted using the Benjamini and Hochberg approach for controlling the false discovery rate; a p-adj < 0.05 was set as the threshold for a significant difference. GO enrichment analysis of different expressed genes (DEGs) was implemented with the GO seq R package. The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway was analyzed in KEGG mapper [36,37]. Transcriptome sequencing data were deposited into the NCBI Gene Expression database with SRA accession number SRR9835574.
4.11. Quantitative Real-Time PCR (qRT-PCR)
The expression of several DEGs was investigated with StepOnePlus™ Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA, USA). Total RNA was isolated from bacterial cultures using Total RNA Extractor (Trizol) (Sangon Biotech). Contaminated DNA was removed from the samples with RNase-free DNase I (TaKaRa, Dalian, China). cDNA synthesis was performed using the PrimeScript RT reagent kit (TaKaRa) according to the manufacturer’s protocol. qRT-PCR was performed using SYBR Premix Ex TaqTM (TaKaRa) with specific primers. Relative gene expression was normalized to the expression of housekeeping gene 16 S via the 2−ΔΔCt method (where Ct = cycle threshold). The assay was repeated three times. Primers used in this assay are listed in Table S2.
4.12. Statistical Analysis
All data were analyzed using the statistical software SPSS (v19.0) using one-way ANOVA. A paired t-test was used for statistical comparisons between groups. The level of statistical significance was set at a p value of 0.05.
5. Conclusions
This study first proved that transcriptional regulator YgeK located at the ETT2 cluster variously and robustly impacts APEC characteristics, including flagellar formation ability, motility, serum sensitivity, and adhesion ability. YgeK significantly upregulated the transcription of several flagellar and virulence-associated genes. Potential pathways explained for phenotypic differences further clarify APEC’s pathogenicity mechanism and the search for new prevention strategies. We clarified the role of the transcriptional regulator YgeK in APEC, and elucidated its regulatory network using transcription sequencing. These results define the critical role of YgeK in APEC’s pathogenicity, providing reference for research on ETT2 cluster function, and guiding the search for new drug targets and vaccines.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/ani11113018/s1. Table S1: Gene Information, Table S2: Primers used in this study. Figure S1: normal growth rate lysogeny broth. Figure S2: Results of plasmid control. Figure S3: Results of plasmid control.
Author Contributions
Conceptualization: K.Q. and M.X.; data curation, J.T., D.F. and M.X.; formal analysis, J.T., D.F. and M.X.; funding acquisition, K.Q.; methodology, M.X., D.F. and Y.G.; project administration, Y.S. and X.S.; resources, Y.S.; software, D.F. and Y.G.; validation, J.T.; writing—original draft, J.T. and M.X.; writing—review and editing, X.S., J.T. and K.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (31772707) and the University Synergy Innovation Program of Anhui Province (GXXT-2019-035).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Transcriptome sequencing data were deposited into the NCBI Gene Expression database with SRA accession number SRR9835574.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Johnson, J.R.; Russo, T.A. Molecular Epidemiology of Extraintestinal Pathogenic (Uropathogenic) Escherichia coli. Int. J. Med. Microbiol. 2005, 295, 383–404. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Nolan, L.K. Characterizing the APEC pathotype. Vet. Res. 2005, 36, 241–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Fakhr, M.K.; Nolan, L.K. Comparison of Escherichia coli isolates implicated in human urinary tract infection and avian colibacillosis. Microbiology 2005, 151, 2097–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonacorsi, S.; Bingen, E. Molecular epidemiology of Escherichia coli causing neonatal meningitis. Int. J. Med. Microbiol. 2005, 295, 373–381. [Google Scholar] [CrossRef]
- Dho-Moulin, M.; Fairbrother, J.M. Avian pathogenic Escherichia coli (APEC). Vet. Res. 1999, 30, 299–316. [Google Scholar]
- Ewers, C.; Antao, E.M.; Diehl, I.; Philipp, H.C.; Wieler, L.H. Intestine and environment of the chicken as reservoirs for extraintestinal pathogenic Escherichia coli strains with zoonotic potential. Appl. Environ. Microbiol. 2009, 75, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Ewers, C.; Janssen, T.; Kiessling, S.; Philipp, H.C.; Wieler, L.H. Molecular epidemiology of avian pathogenic Escherichia coli (APEC) isolated from colisepticemia in poultry. Vet. Microbiol. 2004, 104, 91–101. [Google Scholar] [CrossRef]
- Mokady, D.; Gophna, U.; Ron, E.Z. Virulence factors of septicemic Escherichia coli strains. Int. J. Med. Microbiol. 2005, 295, 455–462. [Google Scholar] [CrossRef]
- Manges, A.R. Escherichia coli and urinary tract infections: The role of poultry-meat. Clin. Microbiol. Infect. 2016, 22, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Ewers, C.; Li, G.; Wilking, H.; Kiessling, S.; Alt, K.; Antao, E.M.; Laturnus, C.; Diehl, I.; Glodde, S.; Homeier, T.; et al. Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related are they? Int. J. Med. Microbiol. 2007, 297, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Green, E.R.; Mecsas, J. Bacterial Secretion Systems: An Overview. Microbiol. Spectr. 2016, 4, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Burkinshaw, B.J.; Strynadka, N.C. Assembly and structure of the T3SS. Biochim. Biophys. Acta 2014, 1843, 1649–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nat. Rev. Microbiol. 2017, 15, 323–337. [Google Scholar] [CrossRef]
- Hayashi, T.; Makino, K.; Ohnishi, M.; Kurokawa, K.; Ishii, K.; Yokoyama, K.; Han, C.G.; Ohtsubo, E.; Nakayama, K.; Murata, T.; et al. Complete genome sequence of enterohemorrhagic Escherichia coli O157:H7 and genomic comparison with a laboratory strain K-12. DNA Res. 2001, 8, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Hartleib, S.; Prager, R.; Hedenstrom, I.; Lofdahl, S.; Tschape, H. Prevalence of the new, SPI1-like, pathogenicity island ETT2 among Escherichia coli. Int. J. Med. Microbiol. 2003, 292, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, X.; Xu, X.; Zhao, Y.; Yang, D.; Han, X.; Tian, M.; Ding, C.; Peng, D.; Yu, S. Escherichia coli type III secretion system 2 (ETT2) is widely distributed in avian pathogenic Escherichia coli isolates from Eastern China. Epidemiol. Infect. 2016, 144, 2824–2830. [Google Scholar] [CrossRef] [Green Version]
- Makino, S.; Tobe, T.; Asakura, H.; Watarai, M.; Ikeda, T.; Takeshi, K.; Sasakawa, C. Distribution of the secondary type III secretion system locus found in enterohemorrhagic Escherichia coli O157:H7 isolates among Shiga toxin-producing E. coli strains. J. Clin. Microbiol. 2003, 41, 2341–2347. [Google Scholar] [CrossRef] [Green Version]
- Shulman, A.; Yair, Y.; Biran, D.; Sura, T.; Otto, A.; Gophna, U.; Becher, D.; Hecker, M.; Ron, E.Z. The Escherichia coli Type III Secretion System 2 Has a Global Effect on Cell Surface. mBio 2018, 9, e01070-18. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liu, X.; Xu, X.; Yang, D.; Wang, D.; Han, X.; Shi, Y.; Tian, M.; Ding, C.; Peng, D. Escherichia coli Type III Secretion System 2 ATPase EivC Is Involved in the Motility and Virulence of Avian Pathogenic Escherichia coli. Front. Microbiol. 2016, 7, 1387. [Google Scholar] [CrossRef] [Green Version]
- Xue, M.; Xiao, Y.; Fu, D.; Raheem, M.A.; Shao, Y.; Song, X.; Tu, J.; Xue, T.; Qi, K. Transcriptional Regulator YqeI, Locating at ETT2 Locus, Affects the Pathogenicity of Avian Pathogenic Escherichia coli. Animals 2020, 10, 1658. [Google Scholar] [CrossRef]
- Luzader, D.H.; Willsey, G.G.; Wargo, M.J.; Kendall, M.M. The Type Three Secretion System 2-Encoded Regulator EtrB Modulates Enterohemorrhagic Escherichia coli Virulence Gene Expression. Infect. Immun. 2016, 84, 2555–2565. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.P.; Chaudhuri, R.R.; Fivian, A.; Bailey, C.M.; Antonio, M.; Barnes, W.M.; Pallen, M.J. The ETT2 gene cluster, encoding a second type III secretion system from Escherichia coli, is present in the majority of strains but has undergone widespread mutational attrition. J. Bacteriol. 2004, 186, 3547–3560. [Google Scholar] [CrossRef] [Green Version]
- Wood, T.K. Precedence for the structural role of flagella in biofilms. mBio 2013, 4, e00225-13. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Yang, Y.; Chen, P.; Hu, H.; Hardwidge, P.R.; Zhu, G. More than a locomotive organelle: Flagella in Escherichia coli. Appl. Microbiol. Biotechnol. 2015, 99, 8883–8890. [Google Scholar] [CrossRef]
- Ewers, C.; Janssen, T.; Wieler, L.H. Avian pathogenic Escherichia coli (APEC). Berliner Münchener Tierärztliche Wochenschrift 2003, 116, 381–395. [Google Scholar]
- Ideses, D.; Gophna, U.; Paitan, Y.; Chaudhuri, R.R.; Pallen, M.J.; Ron, E.Z. A degenerate type III secretion system from septicemic Escherichia coli contributes to pathogenesis. J. Bacteriol. 2005, 187, 8164–8171. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Xie, Y.; Perace, D.; Lu, J.; Tao, J.; Guo, X.; Kim, K.S. The type III secretion system is involved in the invasion and intracellular survival of Escherichia coli K1 in human brain microvascular endothelial cells. FEMS Microbiol. Lett. 2009, 300, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Ewers, C.; Laturnus, C.; Diehl, I.; Alt, K.; Dai, J.; Antao, E.M.; Schnetz, K.; Wieler, L.H. Characterization of a yjjQ mutant of avian pathogenic Escherichia coli (APEC). Microbiology 2008, 154, 1082–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Li, W.; Qi, K.; Wang, S.; Chen, X.; Ni, J.; Deng, R.; Shang, F.; Xue, Y. McbR is involved in biofilm formation and H2O2 stress response in avian pathogenic Escherichia coli X40. Poult. Sci. 2019, 98, 4094–4103. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Raheem, M.A.; Gu, Y.; Lu, H.; Song, X.; Tu, J.; Xue, T.; Qi, K. The KdpD/KdpE two-component system contributes to the motility and virulence of avian pathogenic Escherichia coli. Res. Vet. Sci. 2020, 131, 24–30. [Google Scholar] [CrossRef]
- Herren, C.D.; Mitra, A.; Palaniyandi, S.K.; Coleman, A.; Elankumaran, S.; Mukhopadhyay, S. The BarA-UvrY two-component system regulates virulence in avian pathogenic Escherichia coli O78:K80:H9. Infect. Immun. 2006, 74, 4900–4909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, J.; Huang, B.; Zhang, Y.; Zhang, Y.; Xue, T.; Li, S.; Qi, K. Modulation of virulence genes by the two-component system PhoP-PhoQ in avian pathogenic Escherichia coli. Pol. J. Vet. Sci. 2016, 19, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Yi, Z.; Wang, D.; Xin, S.; Zhou, D.; Li, T.; Tian, M.; Qi, J.; Ding, C.; Wang, S.; Yu, S. The CpxR regulates type VI secretion system 2 expression and facilitates the interbacterial competition activity and virulence of avian pathogenic Escherichia coli. Vet. Res. 2019, 50, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
(a) ETT2 cluster containing 35 genes from ECs3703 to ECs3737 in AE81; (b) alignment analysis of YgeK amino acids in E. coli K12, AE81, and E. coli_O157: H7. Black filled area has the same amino acid, and gray area has different amino acids.
Figure 1.
(a) ETT2 cluster containing 35 genes from ECs3703 to ECs3737 in AE81; (b) alignment analysis of YgeK amino acids in E. coli K12, AE81, and E. coli_O157: H7. Black filled area has the same amino acid, and gray area has different amino acids.

Figure 2.
(a) Bacterial micromorphology of AE81, AE81ΔygeK, and AE81AE81ΔygeK-pCmygeK observed by transmission electron microscopy (×10,000). (b) Motilities of AE81, AE81ΔygeK, and AE81AE81ΔygeK-pCmygeK on semisolid medium.
Figure 2.
(a) Bacterial micromorphology of AE81, AE81ΔygeK, and AE81AE81ΔygeK-pCmygeK observed by transmission electron microscopy (×10,000). (b) Motilities of AE81, AE81ΔygeK, and AE81AE81ΔygeK-pCmygeK on semisolid medium.

Figure 3.
Bacterial resistance of AE81, AE81ΔygeK, and AE81AE81ΔygeK-pCmygeK to SPF chicken serum. SPF chicken serum was added at concentrations of 10%, 20%, 30%, 40%, 50%, and heat-inactivated (HI). (a) SPF chicken serum was added at concentrations of 10%; (b) SPF chicken serum was added at concentrations of 20%; (c) SPF chicken serum was added at concentrations of 30%; (d) SPF chicken serum was added at concentrations of 40%; (e) SPF chicken serum was added at concentrations of 50%; (f) SPF chicken serum was heat-inactivated. Growth was determined using a BioTek Eon plate reader. Turbidity at 600 nm was measured each hour (** p < 0.01, *** p < 0.001, ns: no significance).
Figure 3.
Bacterial resistance of AE81, AE81ΔygeK, and AE81AE81ΔygeK-pCmygeK to SPF chicken serum. SPF chicken serum was added at concentrations of 10%, 20%, 30%, 40%, 50%, and heat-inactivated (HI). (a) SPF chicken serum was added at concentrations of 10%; (b) SPF chicken serum was added at concentrations of 20%; (c) SPF chicken serum was added at concentrations of 30%; (d) SPF chicken serum was added at concentrations of 40%; (e) SPF chicken serum was added at concentrations of 50%; (f) SPF chicken serum was heat-inactivated. Growth was determined using a BioTek Eon plate reader. Turbidity at 600 nm was measured each hour (** p < 0.01, *** p < 0.001, ns: no significance).

Figure 4.
Bacterial adhesion assay with avian DF-1. Inactivation of YgeK decreased adhesion to avian DF-1 cells compared with wild-type AE81 and complemented AE81ΔygeK-pCmygeK strains. Values are the average of three independent experiments. Error bars indicate standard deviation. *** p < 0.01.
Figure 4.
Bacterial adhesion assay with avian DF-1. Inactivation of YgeK decreased adhesion to avian DF-1 cells compared with wild-type AE81 and complemented AE81ΔygeK-pCmygeK strains. Values are the average of three independent experiments. Error bars indicate standard deviation. *** p < 0.01.

Figure 5.
DEG GO classification analysis of wild-type AE81 and mutant AE81ΔygeK strains. DEGs were mainly enriched in biological process, cell composition, and molecular function. Downregulated genes indicated in blue bars; upregulated genes indicated in red bars.
Figure 5.
DEG GO classification analysis of wild-type AE81 and mutant AE81ΔygeK strains. DEGs were mainly enriched in biological process, cell composition, and molecular function. Downregulated genes indicated in blue bars; upregulated genes indicated in red bars.

Figure 6.
KEGG pathway classification analysis of wild-type AE81 and mutant AE81ΔygeK strains. Downregulated genes indicated in blue, and upregulated genes in red.
Figure 6.
KEGG pathway classification analysis of wild-type AE81 and mutant AE81ΔygeK strains. Downregulated genes indicated in blue, and upregulated genes in red.

Figure 7.
Comparative gene transcription levels (cDNA abundance). Transcript levels of biofilm genes csgA, bcsA, and wcaF, flagellar genes fliC and motA, fimbrial gene fimA, and virulence gene ompA in AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK detected by real-time PCR. Results shown as expression ratios relative to expression in wild-type strain AE81. Error bars indicate standard deviation. Statistical significance assessed using two-way ANOVA (* p < 0.05; ** p < 0.01; *** p < 0.001).
Figure 7.
Comparative gene transcription levels (cDNA abundance). Transcript levels of biofilm genes csgA, bcsA, and wcaF, flagellar genes fliC and motA, fimbrial gene fimA, and virulence gene ompA in AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK detected by real-time PCR. Results shown as expression ratios relative to expression in wild-type strain AE81. Error bars indicate standard deviation. Statistical significance assessed using two-way ANOVA (* p < 0.05; ** p < 0.01; *** p < 0.001).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Strains and plasmid used in this study.
| Strains or Plasmid | Genotype or Description | Source |
|---|---|---|
| Strains | ||
| AE81 | APEC clinical strain, isolated from lung | Laboratory stock |
| AE81ΔygeK | AE81 ygeK deletion mutant | This study |
| AE81ΔygeK-pCmygeK | AE81ΔygeK with the plasmid pCmygeK, Cmr 1 | This study |
| Plasmid | ||
| pCmygeK | pSTV28 with ygeK gene, Cmr 1 | This study |
1 Cmr, chloramphenicol-resistant.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tu, J.; Fu, D.; Gu, Y.; Shao, Y.; Song, X.; Xue, M.; Qi, K. Transcription Regulator YgeK Affects the Virulence of Avian Pathogenic Escherichia coli. Animals 2021, 11, 3018. https://doi.org/10.3390/ani11113018
AMA Style
Tu J, Fu D, Gu Y, Shao Y, Song X, Xue M, Qi K. Transcription Regulator YgeK Affects the Virulence of Avian Pathogenic Escherichia coli. Animals. 2021; 11(11):3018. https://doi.org/10.3390/ani11113018
Chicago/Turabian StyleTu, Jian, Dandan Fu, Yi Gu, Ying Shao, Xiangjun Song, Mei Xue, and Kezong Qi. 2021. "Transcription Regulator YgeK Affects the Virulence of Avian Pathogenic Escherichia coli" Animals 11, no. 11: 3018. https://doi.org/10.3390/ani11113018
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.