
In-Feed Supplementation of Resin Acid-Enriched Composition Modulates Gut Microbiota, Improves Growth Performance, and Reduces Post-Weaning Diarrhea and Gut Inflammation in Piglets

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
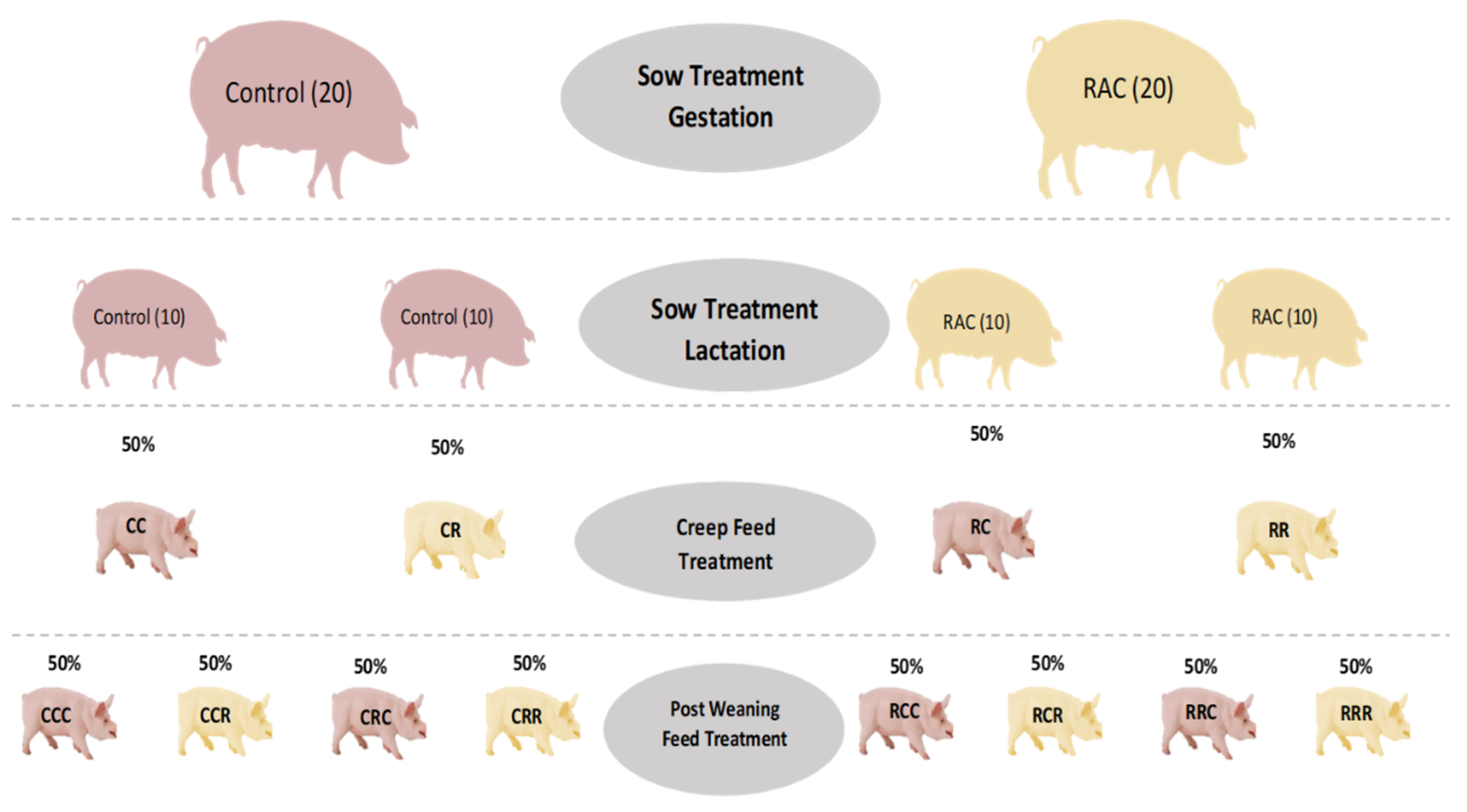
2.1. Animals and Experimental Design
2.1.1. Sow Selection and Feeding
2.1.2. Pre-Weaning/Early Piglet Feeding
2.1.3. Post-Weaning Piglets Feeding
2.2. Parameters and Measurements
2.2.1. Sample Collection
Piglet Sample Collection
Sow Sample Collection
2.3. Laboratory Analysis
2.3.1. Pig Myeloperoxidase ELISA
2.3.2. Gut Microbiota Sequencing
2.4. Statistical Analysis
3. Results
3.1. Live Birth and Total Birth
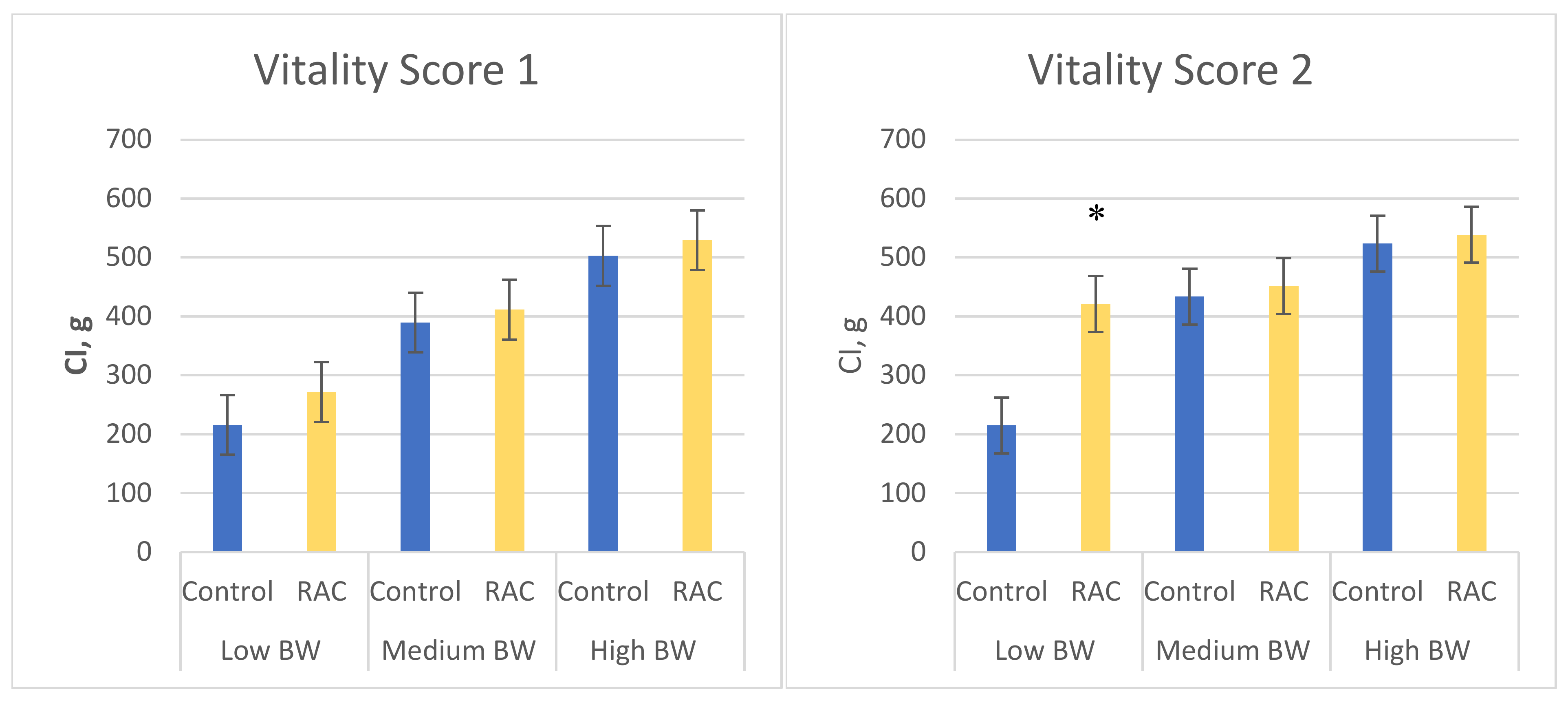
3.2. Piglet Vitality
3.3. Colostrum Intake
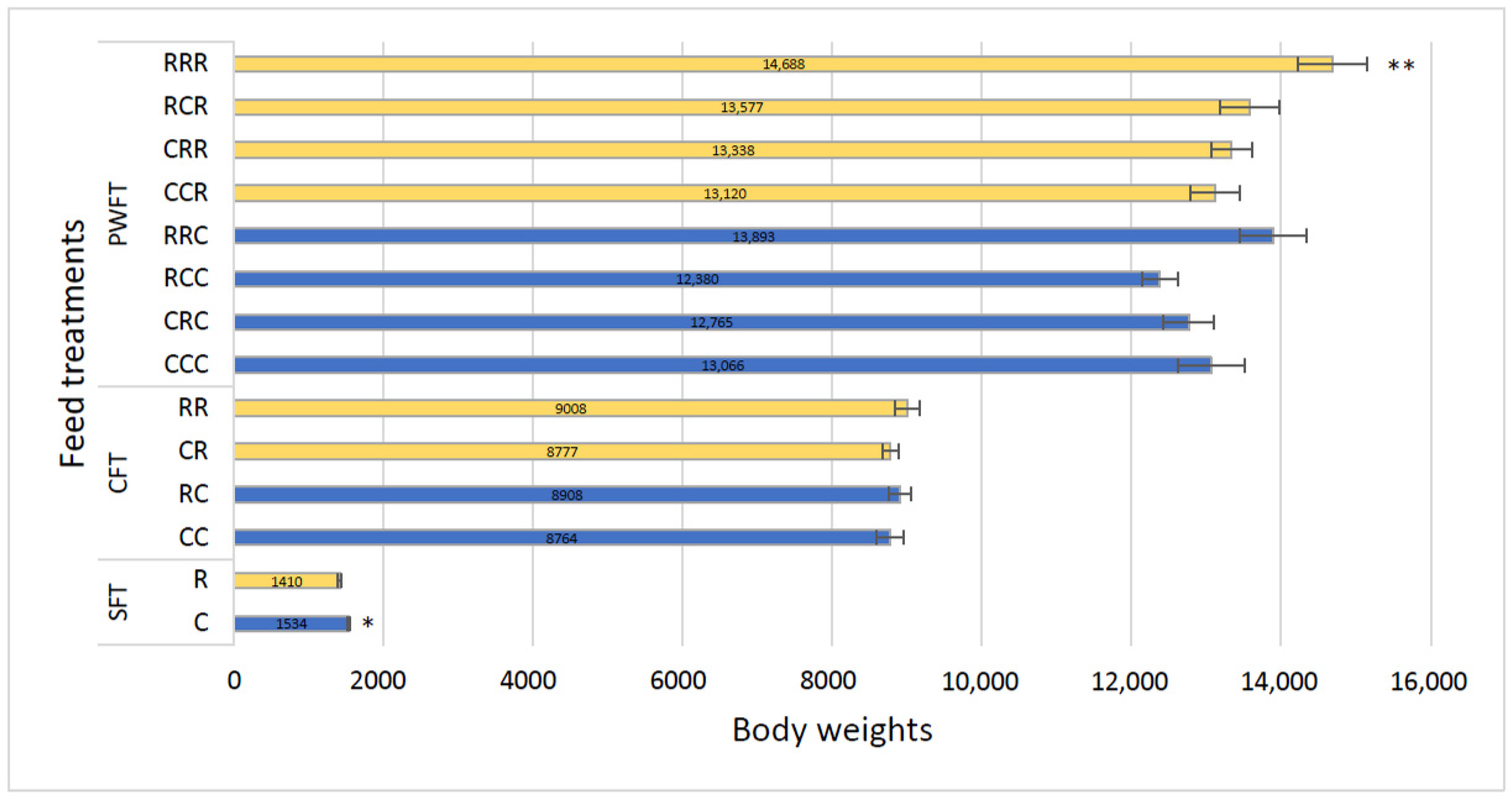
3.4. Piglet Body Weight and Average Daily Growth (ADG)
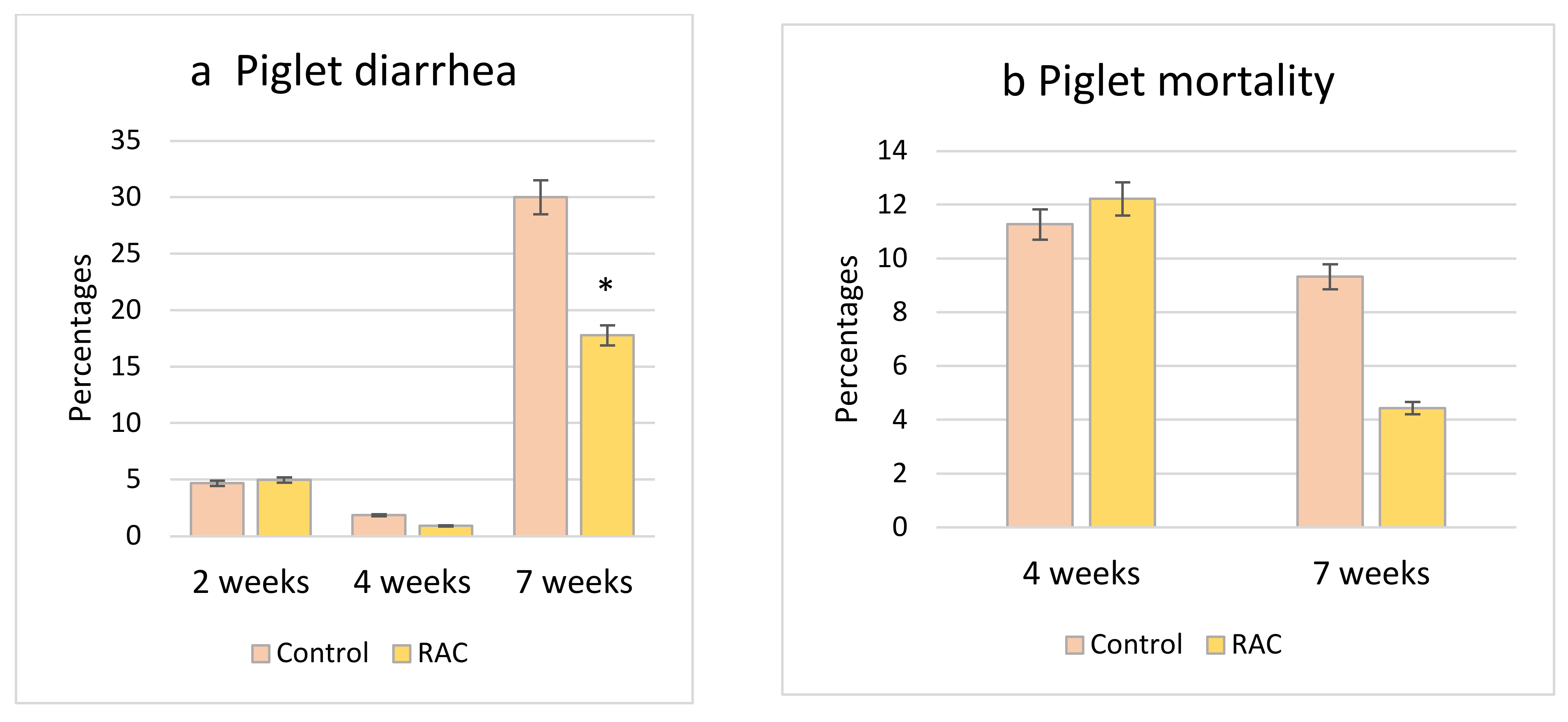
3.5. Piglet Diarrhea Incidence and Mortality Rate
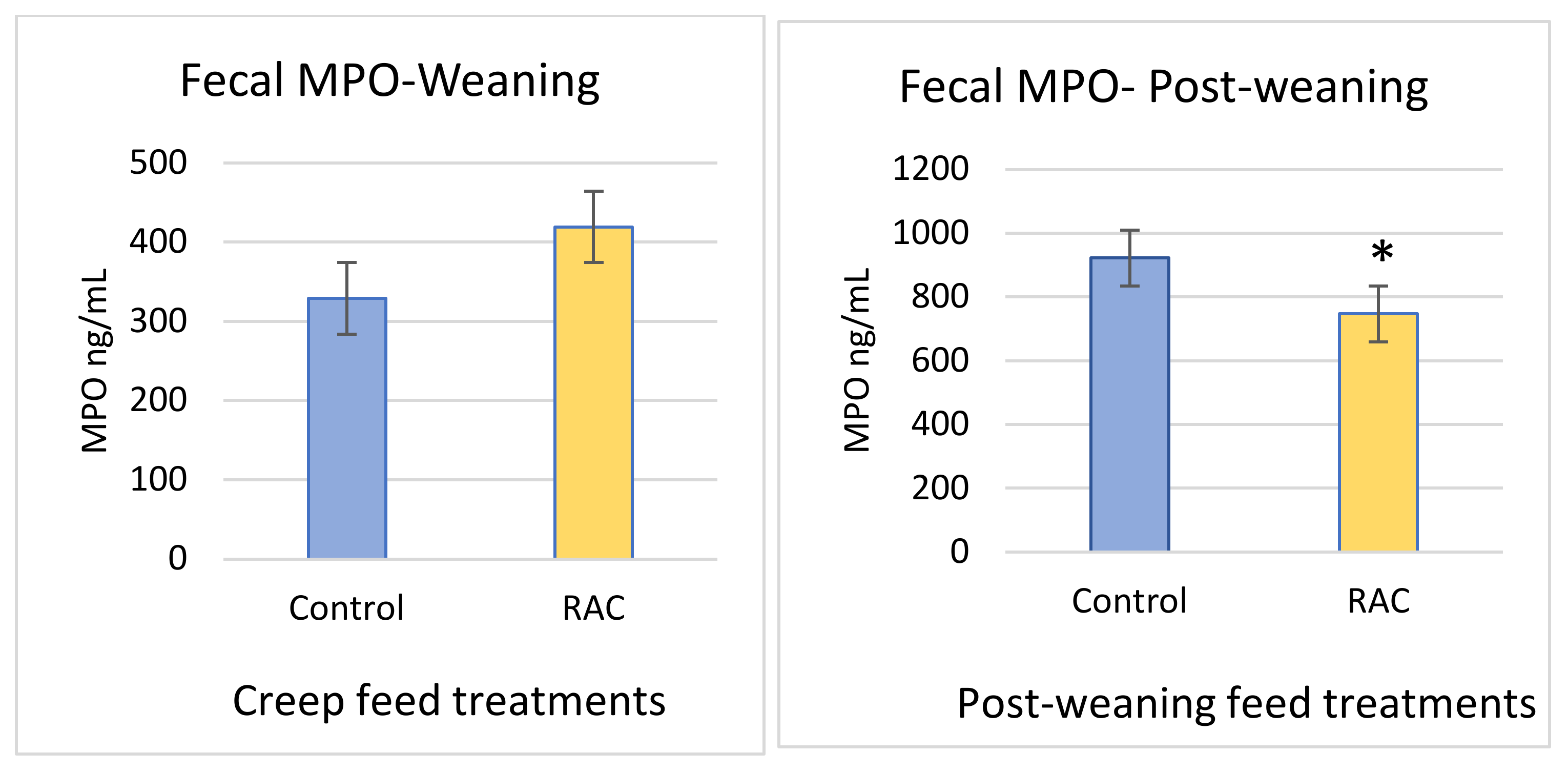
3.6. Fecal Myeloperoxidase
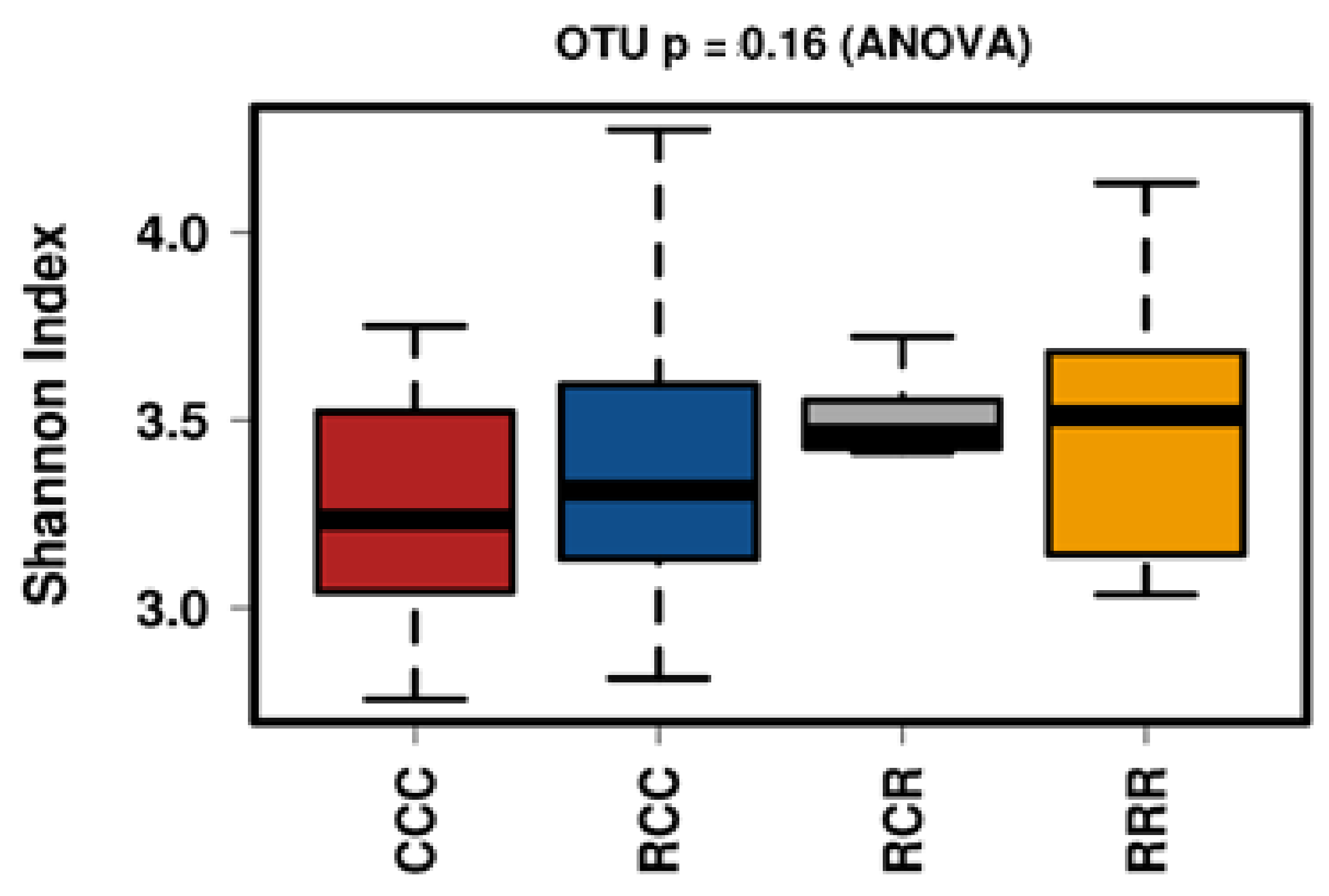
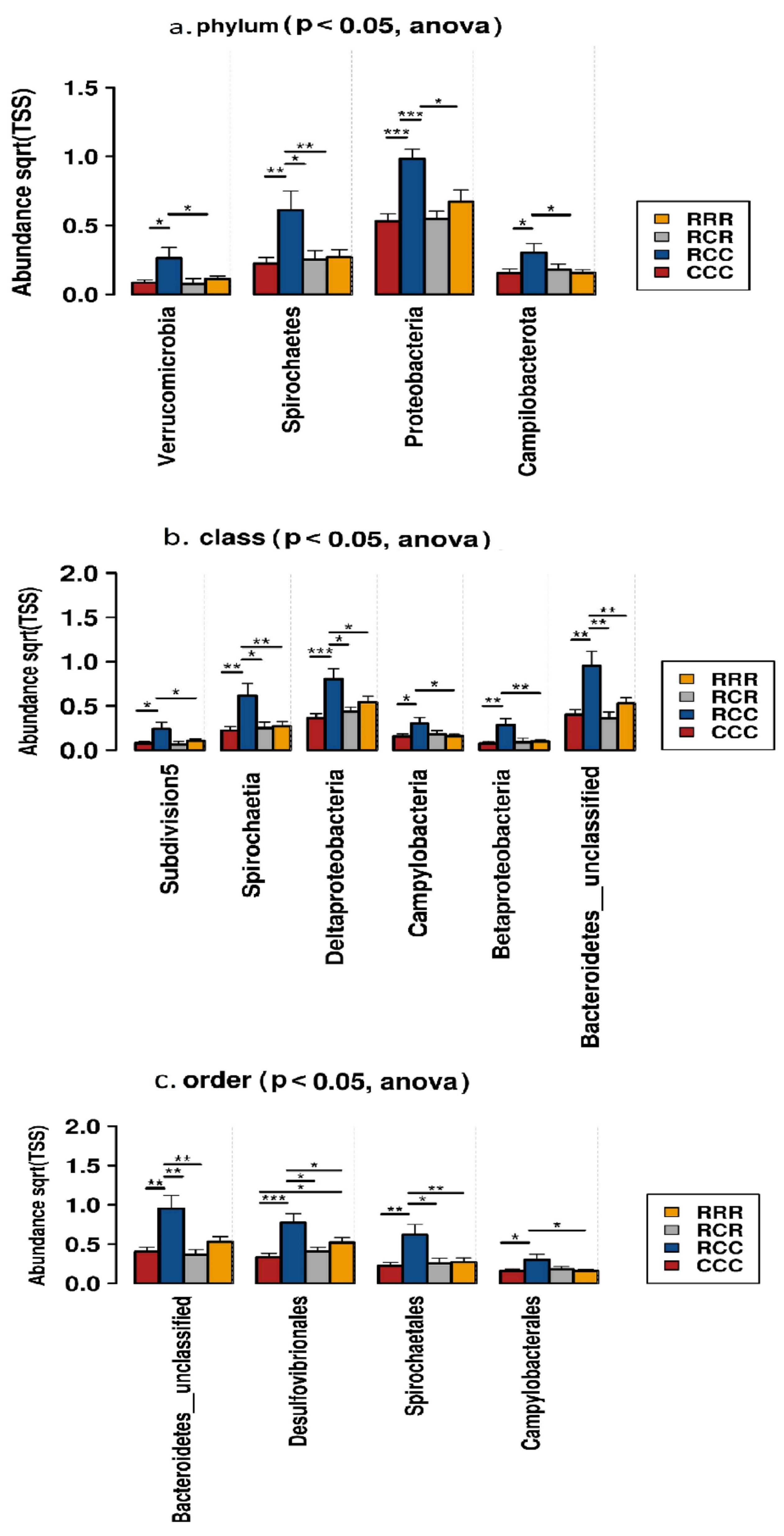
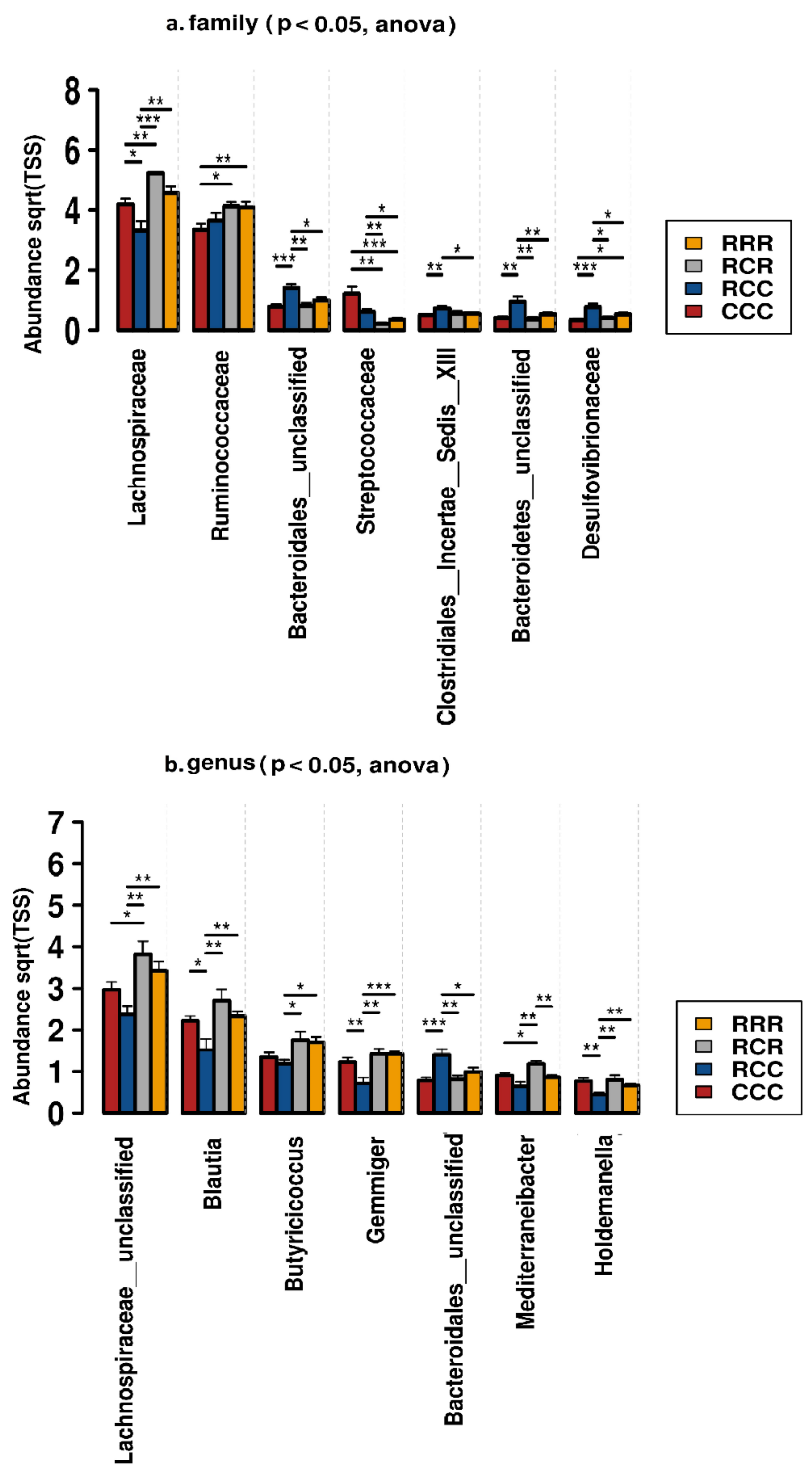
3.7. Microbial Composition and Effects on Performance Parameters
3.8. Association of Microbial Genera with Piglet ADG, and Fecal MPO
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laine, T.M.; Lyytikäinen, T.; Yliaho, M.; Anttila, M. Risk factors for post-weaning diarrhoea on piglet producing farms in Finland. Acta Vet. Scand. 2008, 50, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, J.M.; Opapeju, F.; Pluske, J.; Kim, J.; Hampson, D.; Nyachoti, C.M. Gastrointestinal health and function in weaned pigs: A review of feeding strategies to control post-weaning diarrhoea without using in-feed antimicrobial compounds. J. Anim. Physiol. Anim. Nutr. 2013, 97, 207–237. [Google Scholar] [CrossRef] [PubMed]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut microbiota dysbiosis in postweaning piglets: Understanding the keys to health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- McCracken, B.A.; Spurlock, M.E.; Roos, M.A.; Zuckermann, F.A.; Gaskins, H.R. Weaning anorexia may contribute to local inflammation in the piglet small intestine. J. Nutr. 1999, 129, 613–619. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Duarte, M.E.; Loftus, A.S.; Kim, S.W. Intestinal health of pigs upon weaning: Challenges and nutritional intervention. Front. Vet. Sci. 2021, 8, 628258. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, J.M.; Nadeau, É.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17. [Google Scholar] [CrossRef] [Green Version]
- Vondruskova, H.; Slamova, R.; Trckova, M.; Zraly, Z.; Pavlik, I. Alternatives to antibiotic growth promoters in prevention of diarrhoea in weaned piglets: A review. Vet. Med. 2010, 55, 199–224. [Google Scholar] [CrossRef] [Green Version]
- Long, L.; Chen, J.; Zhang, Y.; Liang, X.; Ni, H.; Zhang, B.; Yin, Y. Comparison of porous and nano zinc oxide for replacing high-dose dietary regular zinc oxide in weaning piglets. PLoS ONE 2017, 12, e0182550. [Google Scholar]
- Adewole, D.; Kim, I.; Nyachoti, C. Gut health of pigs: Challenge models and response criteria with a critical analysis of the effectiveness of selected feed additives—A review. Asian Australas. J. Anim. Sci. 2016, 29, 909. [Google Scholar] [CrossRef] [Green Version]
- Luise, D.; Lauridsen, C.; Bosi, P.; Trevisi, P. Methodology and application of Escherichia coli F4 and F18 encoding infection models in post-weaning pigs. J. Anim. Sci. Biotechnol. 2019, 10, 53. [Google Scholar] [CrossRef]
- Sun, Y.; Kim, S.W. Intestinal challenge with enterotoxigenic Escherichia coli in pigs, and nutritional intervention to prevent postweaning diarrhea. Anim. Nutr. 2017, 3, 322–330. [Google Scholar] [CrossRef]
- Lai, C.; Yin, J.; Li, D.; Zhao, L.; Chen, X. Effects of dietary conjugated linoleic acid supplementation on the performance and immunological responses of weaned pigs after an Escherichia coli lipopolysaccharide challenge. J. Anim. Vet. Adv. 2005, 2, 299–305. [Google Scholar]
- Thacker, P.A. Alternatives to antibiotics as growth promoters for use in swine production: A review. J. Anim. Sci. Biotechnol. 2013, 4, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, S.; Saha, S.; Junnikkala, S.; Orro, T.; Peltoniemi, O.; Oliviero, C. Late gestation diet supplementation of resin acid-enriched composition increases sow colostrum immunoglobulin G content, piglet colostrum intake and improve sow gut microbiota. Animal 2019, 13, 1599–1606. [Google Scholar]
- Kettunen, H.; Vuorenmaa, J.; Rinttilä, T.; Grönberg, H.; Valkonen, E.; Apajalahti, J. Natural resin acid–enriched composition as a modulator of intestinal microbiota and performance enhancer in broiler chicken. J. Appl. Anim. Nutr. 2015, 3, 1–9. [Google Scholar] [CrossRef]
- Roy, K.; Lyhs, U.; Vuorenmaa, J.; Pedersen, K. In vitro inhib ition studies of natural resin acids to Clostridium perfringens, Staphylococcus aureus and Escherichia coli O149. J. Appl. Anim. Nutr. 2018, 6, 1–5. [Google Scholar]
- PináChen, Y. Bio-inspired resin acid-derived materials as anti-bacterial resistance agents with unexpected activities. Chem. Sci. 2014, 5, 2011–2016. [Google Scholar]
- Sipponen, A.; Laitinen, K. Antimicrobial properties of natural coniferous rosin in the European Pharmacopoeia challenge test. Apmis 2011, 119, 720–724. [Google Scholar]
- Wang, J.; Chen, Y.P.; Yao, K.; Wilbon, P.A.; Zhang, W.; Ren, L.; Zhou, J.; Nagarkatti, M.; Wang, C.; Chu, F. Robust antimicrobial compounds and polymers derived from natural resin acids. Chem. Commun. 2012, 48, 916–918. [Google Scholar] [CrossRef]
- Fonseca, T.; Gigante, B.; Marques, M.M.; Gilchrist, T.L.; De Clercq, E. Synthesis and antiviral evaluation of benzimidazoles, quinoxalines and indoles from dehydroabietic acid. Bioorg. Med. Chem. 2004, 12, 103–112. [Google Scholar] [CrossRef]
- Smith, E.; Williamson, E.; Zloh, M.; Gibbons, S. Isopimaric acid from Pinus nigra shows activity against multidrug-resistant and EMRSA strains of Staphylococcus aureus. Phytother. Res. 2005, 19, 538–542. [Google Scholar] [CrossRef]
- Bontempo, V.; Sciannimanico, D.; Pastorelli, G.; Rossi, R.; Rosi, F.; Corino, C. Dietary conjugated linoleic acid positively affects immunologic variables in lactating sows and piglets. J. Nutr. 2004, 134, 817–824. [Google Scholar] [CrossRef]
- Yao, W.; Li, J.; Wang, J.; Zhou, W.; Wang, Q.; Zhu, R.; Wang, F.; Thacker, P. Effects of dietary ratio of n-6 to n-3 polyunsaturated fatty acids on immunoglobulins, cytokines, fatty acid composition, and performance of lactating sows and suckling piglets. J. Anim. Sci. Biotechnol. 2012, 3, 43. [Google Scholar] [CrossRef] [Green Version]
- Le Dividich, J.; Charneca, R.; Thomas, F. Relationship between birth order, birth weight, colostrum intake, acquisition of passive immunity and pre-weaning mortality of piglets. Span. J. Agric. Res. 2017, 15, e0603. [Google Scholar] [CrossRef] [Green Version]
- Quesnel, H.; Farmer, C.; Devillers, N. Colostrum intake: Influence on piglet performance and factors of variation. Livest. Sci. 2012, 146, 105–114. [Google Scholar] [CrossRef]
- Quesnel, H. Colostrum: Roles in piglet performance and production by the sow. In Proceedings of the VI SINSUI–Simposio Internacional de Suinocultura, Porto Alegre, Brazil, 10–13 May 2011; pp. 1–12. [Google Scholar]
- Amdi, C.; Krogh, U.; Flummer, C.; Oksbjerg, N.; Hansen, C.F.; Theil, P. Intrauterine growth restricted piglets defined by their head shape ingest insufficient amounts of colostrum. J. Anim. Sci. 2013, 91, 5605–5613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schodl, K.; Revermann, R.; Winckler, C.; Fuerst-Waltl, B.; Leeb, C.; Willam, A.; Knapp, P.; Pfeiffer, C. Assessment of Piglet Vitality by Farmers—Validation of A Scoring Scheme and Estimation of Associated Genetic Parameters. Animals 2019, 9, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, E.; Jarvis, S.; D’eath, R.; Ross, D.; Robson, S.; Farish, M.; Nevison, I.; Lawrence, A.; Edwards, S. Investigating the behavioural and physiological indicators of neonatal survival in pigs. Theriogenology 2008, 69, 773–783. [Google Scholar] [CrossRef]
- Rooney, H.B.; O’driscoll, K.; O’doherty, J.V.; Lawlor, P.G. Effect of increasing dietary energy density during late gestation and lactation on sow performance, piglet vitality, and lifetime growth of offspring. J. Anim. Sci. 2020, 98, skz379. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Lee, J.H.; Lee, S.H.; Seok, M.-J.; Kim, D.W.; Kang, B.N.; Johnson, T.J.; Isaacson, R.E.; Kim, H.B. Piglet gut microbial shifts early in life: Causes and effects. J. Anim. Sci. Biotech. 2019, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Macpherson, A.J.; de Agüero, M.G.; Ganal-Vonarburg, S.C. How nutrition and the maternal microbiota shape the neonatal immune system. Nat. Rev. Immunol. 2017, 17, 508. [Google Scholar] [CrossRef]
- Al Nabhani, Z.; Dulauroy, S.; Lécuyer, E.; Polomack, B.; Campagne, P.; Berard, M.; Eberl, G. Excess calorie intake early in life increases susceptibility to colitis in adulthood. Nat. Metab. 2019, 1, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hansen, C.; Mullan, B.; Pluske, J. Nutrition and pathology of weaner pigs: Nutritional strategies to support barrier function in the gastrointestinal tract. Anim. Feed Sci. Technol. 2012, 173, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Guevarra, R.B.; Hong, S.H.; Cho, J.H.; Kim, B.-R.; Shin, J.; Lee, J.H.; Kang, B.N.; Kim, Y.H.; Wattanaphansak, S.; Isaacson, R.E. The dynamics of the piglet gut microbiome during the weaning transition in association with health and nutrition. J. Anim. Sci. Biotechnol. 2018, 9, 54. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Hu, H.; Zijlstra, R.T.; Zheng, J.; Gänzle, M.G. Metagenomic reconstructions of gut microbial metabolism in weanling pigs. Microbiome 2019, 7, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vienola, K.; Jurgens, G.; Vuorenmaa, J.; Apajalahti, J. Tall oil fatty acid inclusion in the diet improves performance and increases ileal density of lactobacilli in broiler chickens. Br. Poult. Sci. 2018, 59, 349–355. [Google Scholar] [CrossRef]
- Kettunen, H.; Van Eerden, E.; Lipiński, K.; Rinttilä, T.; Valkonen, E.; Vuorenmaa, J. Dietary resin acid composition as a performance enhancer for broiler chickens. J. Appl. Anim. Nutr. 2017, 5, 1–8. [Google Scholar] [CrossRef]
- Theil, P.K.; Flummer, C.; Hurley, W.; Kristensen, N.B.; Labouriau, R.; Sørensen, M.T. Mechanistic model to predict colostrum intake based on deuterium oxide dilution technique data and impact of gestation and prefarrowing diets on piglet intake and sow yield of colostrum. J. Anim. Sci. 2014, 92, 5507–5519. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.; Junnikkala, S.; Valros, A.; Peltoniemi, O.; Oliviero, C. Validation of Brix refractometer to estimate colostrum immunoglobulin G content and composition in the sow. Animal 2016, 10, 1728–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, P.A.; Aho, V.T.; Paulin, L.; Pekkonen, E.; Auvinen, P.; Scheperjans, F. Oral and nasal microbiota in Parkinson’s disease. Parkinsonism Relat. Disord. 2017, 38, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Zakrzewski, M.; Proietti, C.; Ellis, J.J.; Hasan, S.; Brion, M.-J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome–environment interactions. Bioinformatics 2017, 33, 782–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyriakis, S. New aspects of the prevention and/or treatment of the major stress induced diseases of the early weaned piglet. Pig News Inf. 1989, 10, 177–181. [Google Scholar]
- Varley, M.A. The Neonatal Pig: Development and Survival; CAB International: Wallingford, UK, 1995. [Google Scholar]
- Suiryanrayna, M.V.; Ramana, J. A review of the effects of dietary organic acids fed to swine. J. Anim. Sci. Biotechnol. 2015, 6, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguirre, M.; Vuorenmaa, J.; Valkonen, E.; Kettunen, H.; Callens, C.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F.; Goossens, E. In-feed resin acids reduce matrix metalloproteinase activity in the ileal mucosa of healthy broilers without inducing major effects on the gut microbiota. Vet. Res. 2019, 50, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sipponen, A.; Peltola, R.; Jokinen, J.J.; Laitinen, K.; Lohi, J.; Rautio, M.; Männistö, M.; Sipponen, P.; Lounatmaa, K. Effects of Norway spruce (Picea abies) resin on cell wall and cell membrane of Staphylococcus aureus. Ultrastruct. Pathol. 2009, 33, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Partanen, K. Organic acids-their efficacy and modes of action in pigs. Gut Environ. Pigs/A. Piva KE Bach Knudsen JE Lindberg 2001, 2, 201–217. [Google Scholar]
- Piva, A.; Morlacchini, M.; Casadei, G.; Gatta, P.P.; Biagi, G.; Prandini, A. Sodium butyrate improves growth performance of weaned piglets during the first period after weaning. Ital. J. Anim. Sci. 2002, 1, 35–41. [Google Scholar] [CrossRef]
- Bonetti, A.; Tugnoli, B.; Piva, A.; Grilli, E. Towards zero zinc oxide: Feeding strategies to manage post-weaning diarrhea in piglets. Animals 2021, 11, 642. [Google Scholar] [CrossRef]
- Hasan, S.; Junnikkala, S.; Peltoniemi, O.; Paulin, L.; Lyyski, A.; Vuorenmaa, J.; Oliviero, C. Dietary supplementation with yeast hydrolysate in pregnancy influences colostrum yield and gut microbiota of sows and piglets after birth. PLoS ONE 2018, 13, e0197586. [Google Scholar] [CrossRef] [Green Version]
- Patterson, R.; Connor, M.; Krause, D.; Nyachoti, C.J.A. Response of piglets weaned from sows fed diets supplemented with conjugated linoleic acid (CLA) to an Escherichia coli K88+ oral challenge. Animal 2008, 2, 1303–1311. [Google Scholar] [CrossRef] [Green Version]
- Bee, G. Dietary conjugated linoleic acid consumption during pregnancy and lactation influences growth and tissue composition in weaned pigs. J. Nutr. 2000, 130, 2981–2989. [Google Scholar] [CrossRef] [Green Version]
- Larson, M.A.; Wei, S.H.; Weber, A.; Mack, D.R.; McDonald, T.L. Human serum amyloid A3 peptide enhances intestinal MUC3 expression and inhibits EPEC adherence. Biochem. Biophys. Res. Commun. 2003, 300, 531–540. [Google Scholar] [CrossRef]
- Poonsuk, K.; Zhang, J.; Chen, Q.; Gonzalez, W.; da Silva Carrion, L.C.; Sun, Y.; Ji, J.; Wang, C.; Main, R.; Zimmerman, J. Quantifying the effect of lactogenic antibody on porcine epidemic diarrhea virus infection in neonatal piglets. Vet. Microbiol. 2016, 197, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Kielland, C.; Rootwelt, V.; Reksen, O.; Framstad, T. The association between immunoglobulin G in sow colostrum and piglet plasma. J. Anim. Sci. 2015, 93, 4453–4462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lallès, J.-P.; Boudry, G.; Favier, C.; Le Floc’h, N.; Luron, I.; Montagne, L.; Oswald, I.P.; Pié, S.; Piel, C.; Sève, B. Gut function and dysfunction in young pigs: Physiology. Anim. Res. 2004, 53, 301–316. [Google Scholar] [CrossRef] [Green Version]
- Lalles, J.-P.; Bosi, P.; Smidt, H.; Stokes, C.R. Nutritional management of gut health in pigs around weaning. Proc. Nutr. Soc. 2007, 66, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Hansberry, D.R.; Shah, K.; Agarwal, P.; Agarwal, N. Fecal myeloperoxidase as a biomarker for inflammatory bowel disease. Cureus 2017, 9, e1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krawisz, J.; Sharon, P.; Stenson, W. Quantitative assay for acute intestinal inflammation based on myeloperoxidase activity: Assessme nt of inflammation in rat and hamster models. Gastroenterology 1984, 87, 1344–1350. [Google Scholar] [CrossRef]
- Bontempo, V.; Jiang, X.; Cheli, F.; Lo Verso, L.; Mantovani, G.; Vitari, F.; Domeneghini, C.; Agazzi, A. Administration of a novel plant extract product via drinking water to post-weaning piglets: Effects on performance and gut health. Animal 2014, 721–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, R.; Li, X.; Li, J.; Ji, W.; Zeng, X.; Bao, J. Exposure to the environmental pollutant ammonia causes changes in gut microbiota and inflammatory markers in fattening pigs. Ecotoxicol. Environ. Saf. 2021, 208, 111564. [Google Scholar] [CrossRef] [PubMed]
- Pearce, S.C.; Mani, V.; Boddicker, R.L.; Johnson, J.S.; Weber, T.E.; Ross, J.W.; Rhoads, R.P.; Baumgard, L.H.; Gabler, N.K. Heat stress reduces intestinal barrier integrity and favors intestinal glucose transport in growing pigs. PLoS ONE 2013, 8, e70215. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.-S.; Hirai, S.; Goto, T.; Kuroyanagi, K.; Lee, J.-Y.; Uemura, T.; Ezaki, Y.; Takahashi, N.; Kawada, T. Dehydroabietic acid, a phytochemical, acts as ligand for PPARs in macrophages and adipocytes to regulate inflammation. Biochem. Biophys. Res. Commun. 2008, 369, 333–338. [Google Scholar] [CrossRef]
- Lai, C.; Yin, J.; Li, D.; Zhao, L.; Qiao, S.; Xing, J. Conjugated linoleic acid attenuates the production and gene expression of proinflammatory cytokines in weaned pigs challenged with lipopolysaccharide. J. Nutr. 2005, 135, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, N.; Favier, C.; Techer, C.; Guillou, D. Factors influencing faecal myeloperoxidase in piglets from trials without in-feed therapeuticals. In Proceedings of the Zero Zinc Summit, Copenhagen, Denmark, 17–18 June 2019; pp. 3–12. [Google Scholar]
- Schiavon, S.; Tagliapietra, F.; Bailoni, L.; Bortolozzo, A. Effects of sugar beet pulp on growth and health status of weaned piglets. Ital. J. Anim. Sci. 2004, 3, 337–351. [Google Scholar] [CrossRef]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Burrough, E. Swine dysentery: Etiopathogenesis and diagnosis of a reemerging disease. Vet. Pat. 2017, 54, 22–31. [Google Scholar] [CrossRef]
- Hampson, D.; Trott, D. A Review-Intestinal spirochaetal infections of pigs: An overview with an Australian perspective. In Proceedings of the Sixth Biennial Conference of the Australasian Pig Science Association (APSA), Canberra, Australia, 26–29 November 1995; pp. 139–169. [Google Scholar]
- Fujio-Vejar, S.; Vasquez, Y.; Morales, P.; Magne, F.; Vera-Wolf, P.; Ugalde, J.A.; Navarrete, P.; Gotteland, M. The gut microbiota of healthy chilean subjects reveals a high abundance of the phylum verrucomicrobia. Front. Microbiol. 2017, 8, 1221. [Google Scholar] [CrossRef] [Green Version]
- Carrique-Mas, J.; Bryant, J.; Cuong, N.; Hoang, N.; Campbell, J.; Hoang, N.; Dung, T.; Duy, D.; Hoa, N.; Thompson, C. An epidemiological investigation of Campylobacter in pig and poultry farms in the Mekong delta of Vietnam. Epidemiol. Infect. 2014, 142, 1425–1436. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Song, P.; Fan, P.; Hou, C.; Thacker, P.; Ma, X. Dietary sodium butyrate decreases postweaning diarrhea by modulating intestinal permeability and changing the bacterial communities in weaned piglets. J. Nutr. 2015, 145, 2774–2780. [Google Scholar] [CrossRef] [PubMed]
- La Reau, A.J.; Suen, G. The Ruminococci: Key symbionts of the gut ecosystem. J. Microbiol. 2018, 56, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Koh, H.-W.; Kim, M.S.; Lee, J.-S.; Kim, H.; Park, S.-J. Changes in the swine gut microbiota in response to porcine epidemic diarrhea infection. Microbes Environ. 2015, 30, 284–287. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Peng, Q.; Chen, Y.; Zhou, X.; Zhang, F.; Li, A.; Huang, D.; Wu, Q.; Ye, Y.; He, H. Altered gut microbiota profiles in sows and neonatal piglets associated with porcine epidemic diarrhea virus infection. Sci. Rep. 2017, 7, 17439. [Google Scholar] [CrossRef]
- Loubinoux, J.; Bronowicki, J.-P.; Pereira, I.A.; Mougenel, J.-L.; Le Faou, A.E. Sulfate-reducing bacteria in human feces and their association with inflammatory bowel diseases. FEMS Microbiol. Ecol. 2002, 40, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Mach, N.; Berri, M.; Estellé, J.; Levenez, F.; Lemonnier, G.; Denis, C.; Leplat, J.J.; Chevaleyre, C.; Billon, Y.; Doré, J. Early-life establishment of the swine gut microbiome and impact on host phenotypes. Environ. Microbiol. Rep. 2015, 7, 554–569. [Google Scholar] [CrossRef]
- Ahn, S.; Jin, T.-E.; Chang, D.-H.; Rhee, M.-S.; Kim, H.J.; Lee, S.J.; Park, D.-S.; Kim, B.-C. Agathobaculum butyriciproducens gen. nov. sp. nov., a strict anaerobic, butyrate-producing gut bacterium isolated from human faeces and reclassification of Eubacterium desmolans as Agathobaculum desmolans comb. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 3656–3661. [Google Scholar] [CrossRef]
- Sassone-Corsi, M.; Raffatellu, M. No vacancy: How beneficial microbes cooperate with immunity to provide colonization resistance to pathogens. J. Immunol. 2015, 194, 4081–4087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eeckhaut, V.; Machiels, K.; Perrier, C.; Romero, C.; Maes, S.; Flahou, B.; Steppe, M.; Haesebrouck, F.; Sas, B.; Ducatelle, R. Butyricicoccus pullicaecorum in inflammatory bowel disease. Gut 2013, 62, 1745–1752. [Google Scholar] [CrossRef]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the genetic basis of fibrolytic specialization by Lachnospiraceae and Ruminococcaceae in diverse gut communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Kim, J.-S.; Lee, K.C.; Suh, M.K.; Han, K.-I.; Eom, M.K.; Lee, J.H.; Park, S.-H.; Kang, S.W.; Park, J.-E.; Oh, B.S. Mediterraneibacter butyricigenes sp. nov., a butyrate-producing bacterium isolated from human faeces. J. Microbiol. 2019, 57, 38–44. [Google Scholar] [CrossRef]
- Surana, N.; Kasper, D. Lachnospiraceae protect from colitis by regulating colonic group 3 innate lymphoid cells. Eur. J. Immunol. 2016, 46, 346. [Google Scholar]
- Sun, J.; Du, L.; Li, X.; Zhong, H.; Ding, Y.; Liu, Z.; Ge, L. Identification of the core bacteria in rectums of diarrheic and non-diarrheic piglets. Sci. Rep. 2019, 9, 18675. [Google Scholar] [CrossRef] [PubMed]
- Kubasova, T.; Davidova-Gerzova, L.; Babak, V.; Cejkova, D.; Montagne, L.; Le-Floc’h, N.; Rychlik, I. Effects of host genetics and environmental conditions on fecal microbiota composition of pigs. PLoS ONE 2018, 13, e0201901. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xie, Y.; Zhong, R.; Liu, L.; Lin, C.; Xiao, L.; Chen, L.; Zhang, H.; Beckers, Y.; Everaert, N. Effects of xylo-oligosaccharides on growth and gut microbiota as potential replacements for antibiotic in weaning piglets. Front. Microbiol. 2021, 12, 355. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | RAC Treatment | Control | p-Value |
|---|---|---|---|
| Farrowing and sow characteristics | |||
| Number of sows | 20 | 20 | |
| Gestation length, day | 117.01 ± 0.30 | 117.51 ± 0.30 | 0.25 |
| Farrowing duration, min | 331.45 ± 41.95 | 296.9 ± 36.91 | 0.54 |
| Sow back fat, pregnancy | 10.4 ± 0.31 | 10.4 ± 0.33 | 1.00 |
| Sow back fat, farrowing | 11.65 ± 0.31 | 11.3 ± 0.28 | 0.40 |
| Sow back fat, weaning | 8.15 ± 0.16 | 8.2 ± 0.22 | 0.85 |
| Colostrum characteristics | |||
| Colostrum yield, g | 6197.36 ± 205.03 | 6017.64 ± 196.50 | 0.53 |
| Colostrum brix value | 29.79 ± 0.78 | 28.36 ± 0.75 | 0.19 |
| Colostrum intake, g | 444.91 ± 7.86 | 452.44 ± 9.49 | 0.54 |
| Colostrum serum amyloid A (SAA), mg/L | 673.83 ± 71.47 | 536.73 ± 57.90 | 0.14 |
| Litter characteristics | |||
| Total born/litter size | 15 ± 0.54 | 13.5 ± 0.67 | 0.09 |
| Live-born piglets | 14.15 ± 0.50 | 13.09 ± 0.20 | 0.35 |
| Stillborn piglets | 0.85 ± 0.23 | 0.15 ± 0.08 | 0.01 * |
| Piglet vitality | 1.37 ± 0.03 | 1.39 ± 0.03 | 0.55 |
| Weaned piglets | 12.75 | 12.25 | 0.54 |
| Treatment Group | Treatment | ADG | Estimate | (95% Conf. Interval) | p-Value |
|---|---|---|---|---|---|
| CFT (383 piglets) | CC | 313 | |||
| CR | 313 | 0.45 | −14.70–15.60 | 0.95 | |
| RC | 318 | 5.15 | −10.20–20.50 | 0.51 | |
| RR | 322 | 8.71 | −6.52–23.94 | 0.26 | |
| PWFT (285 piglets) | CCC | 272 | |||
| CCR | 273 | 1.12 | −20.71–22.95 | 0.92 | |
| CRC | 266 | −6.27 | −28.51–15.98 | 0.58 | |
| CRR | 278 | 5.65 | −17.09–28.40 | 0.63 | |
| RCC | 258 | −14.30 | −35.26–6.66 | 0.18 | |
| RCR | 283 | 10.63 | −11.77–33.03 | 0.35 | |
| RRC | 289 | 17.23 | −5.18–39.63 | 0.13 | |
| RRR | 306 | 33.78 | 11.21–56.35 | 0.003 ** |
| Outcome Variable | Predictor Variable | Odds Ratio | Confidence Interval | p-Value |
|---|---|---|---|---|
| Post-weaning diarrhea (7 weeks) | Piglet diarrhea at weaning | 11.60 | 1.25–107.28 | 0.031 * |
| RAC | 0.52 | 0.29–0.93 | 0.027 * | |
| Mortality at post-weaning (7 weeks) | BW at weaning | |||
| Small | 1 | 0.031 * | ||
| Average | 0.20 | 0.02–2.46 | ||
| Large | 0.09 | 0.01–1.09 | ||
| Piglet diarrhea at weaning | 8.63 | 1.34–59.04 | 0.026 * |
| Taxa (Genus) | Fecal MPO | ADG | CCC Mean | RCC Mean | RCR Mean | RRR Mean | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|
| Pearson’s Index | p-Value | Pearson’s Index | p-Value | ||||||
| Bacteroidales__unclassified | 0.24 | 0.10 | −0.18 | 0.23 | 0.79 | 1.40 | 0.81 | 0.99 | 0.00 * |
| Blautia | −0.12 | 0.42 | 0.33 | 0.02 | 2.22 | 1.52 | 2.70 | 2.33 | 0.00 * |
| Butyricicoccus | −0.20 | 0.18 | 0.16 | 0.29 | 1.34 | 1.18 | 1.75 | 1.70 | 0.02 * |
| Clostridiales__unclassified | 0.10 | 0.49 | −0.02 | 0.88 | 1.27 | 1.43 | 1.39 | 1.41 | 0.86 |
| Clostridium__sensu__stricto | 0.51 | 0.00 * | −0.20 | 0.18 | 4.22 | 4.23 | 3.00 | 3.30 | 0.14 |
| Dorea | −0.27 | 0.07 | 0.10 | 0.51 | 0.86 | 0.69 | 0.86 | 0.80 | 0.30 |
| Faecalibacterium | −0.30 | 0.04 * | 0.47 | 0.00 | 1.49 | 1.18 | 2.05 | 1.68 | 0.05 * |
| Firmicutes__unclassified | 0.30 | 0.04 * | −0.15 | 0.30 | 0.54 | 0.91 | 0.63 | 0.71 | 0.19 |
| Gemmiger | −0.35 | 0.02 * | 0.52 | 0.00 | 1.23 | 0.72 | 1.42 | 1.42 | 0.00 * |
| Holdemanella | −0.10 | 0.53 | −0.09 | 0.57 | 0.76 | 0.43 | 0.80 | 0.66 | 0.01 * |
| Lachnospiraceae__unclassified | −0.24 | 0.11 | 0.51 | 0.00 | 2.96 | 2.37 | 3.82 | 3.42 | 0.01 * |
| Lactobacillus | −0.29 | 0.054 | −0.03 | 0.84 | 4.64 | 4.40 | 4.10 | 3.95 | 0.51 |
| Limosilactobacillus | −0.11 | 0.45 | −0.02 | 0.88 | 2.29 | 2.19 | 1.76 | 2.14 | 0.58 |
| Mediterraneibacter | −0.15 | 0.33 | −0.04 | 0.78 | 0.90 | 0.64 | 1.18 | 0.86 | 0.00 * |
| Megasphaera | −0.35 | 0.02 * | 0.20 | 0.19 | 1.42 | 1.59 | 1.75 | 1.78 | 0.70 |
| Prevotella | −0.36 | 0.01 * | 0.27 | 0.07 | 1.51 | 1.67 | 2.13 | 2.25 | 0.27 |
| Romboutsia | 0.43 | 0.00 * | −0.33 | 0.02 | 0.72 | 1.02 | 0.64 | 0.57 | 0.13 |
| Ruminococcaceae__unclassified | 0.23 | 0.13 | −0.04 | 0.78 | 1.88 | 2.64 | 2.10 | 2.48 | 0.05 * |
| Ruminococcus | −0.20 | 0.18 | 0.14 | 0.36 | 0.84 | 0.98 | 1.13 | 1.03 | 0.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uddin, M.K.; Hasan, S.; Mahmud, M.R.; Peltoniemi, O.; Oliviero, C. In-Feed Supplementation of Resin Acid-Enriched Composition Modulates Gut Microbiota, Improves Growth Performance, and Reduces Post-Weaning Diarrhea and Gut Inflammation in Piglets. Animals 2021, 11, 2511. https://doi.org/10.3390/ani11092511
Uddin MK, Hasan S, Mahmud MR, Peltoniemi O, Oliviero C. In-Feed Supplementation of Resin Acid-Enriched Composition Modulates Gut Microbiota, Improves Growth Performance, and Reduces Post-Weaning Diarrhea and Gut Inflammation in Piglets. Animals. 2021; 11(9):2511. https://doi.org/10.3390/ani11092511
Chicago/Turabian StyleUddin, Md Karim, Shah Hasan, Md. Rayhan Mahmud, Olli Peltoniemi, and Claudio Oliviero. 2021. "In-Feed Supplementation of Resin Acid-Enriched Composition Modulates Gut Microbiota, Improves Growth Performance, and Reduces Post-Weaning Diarrhea and Gut Inflammation in Piglets" Animals 11, no. 9: 2511. https://doi.org/10.3390/ani11092511