1. Introduction
In Spain, two hen trunks have historically been differentiated; the Atlantic trunk, generally comprising larger-format dual-purpose birds, with red earlobes and brown-shelled eggs, and the Mediterranean trunk, consisting of lighter individuals, with white earlobes and of a white-shelled egg-laying morphotype [
1]. The aforementioned features have been considered by breeders on a regular basis for breed ascription and animal classification. This segregation of the Atlantic and Mediterranean trunks would later be supported from a molecular perspective through the estimation of genetic distances using microsatellite markers [
2].
As a result, natural and human selection led to a high heterogeneity and variability of morphological characteristics in avian breeds [
3,
4]. Such high heterogeneity was promoted when breeding objectives (meat, eggs, or dual-purpose breeds) and, hence, morphological characteristics started to differ and polarize among populations to adapt to environment requirements at the minimum biological cost. These differentiation processes determined breeds to base their adaptability strategies on their particular enhanced body features [
5].
Andalusia (Southern Spain) is influenced by the Mediterranean climate, with maximum temperatures rising above 40 °C in summer, as reported by the Spanish State Meteorological Agency (AEMET). In this context, very high temperatures are present from late spring on and last for the whole summer. Among the breeds in the area, two laying hen genotypes have traditionally configured poultry production under backyard and extensive systems: the Utrerana and Sureña avian breeds [
6,
7].
The Utrerana and Sureña avian breeds share a common geographic location, socioeconomic context, and history. In addition, four varieties of plumage color are present in both breeds: White, Franciscan, Black, and Partridge in the Utrerana breed; White, Franciscan, Black, Partridge, Blue, and Splash in the Sureña Breed. However, the Sureña hen has a larger format than most Mediterranean hen breeds [
8,
9].
These widely accessible low-capital/input investment birds were historically kept in sustainable systems for decades, thus becoming the source of production of high-biological-value proteins in rural livelihoods until globalization called for the intensification of animal production [
10,
11].
As a direct consequence, the population census of Spanish breeds suffered a regression due to the introduction of selected commercial strains of birds with a higher production during the last half of the 20th century [
12,
13]. In this way, the Utrerana avian breed became classified as an endangered breed, according to Royal Decree 45/2019 of 8 February, while the Sureña avian breed is in the process of being included in the Official Livestock Breeds Catalog of the Ministry of Agriculture, Fisheries, and Environment (MAPA) of Spain.
Consumers’ interest in quality food products revolved around market demands as a conscious response to the drawbacks implied by intensive production. Food alternatives produced through sustainable production systems became popular, provided these systems were characterized by a low impact on the environment and human health while they also considered animal welfare [
14]. Increased demands soon translated into commercial chains starting to request differentiated products, whose properties significantly differed from products obtained through hybrid commercial strains [
15].
For local producers to be able to fulfill market demands, products and the elements needed to ensure their constant supply must be defined through breed characterization zoometrically, genetically, or even productively. Contextually, the characterization of local populations, as well as the relationship among already established breeds, can provide pieces of evidence on the mechanism and events that contributed to the origin and development of native poultry breeds in the south region of Spain, as well as the adaptive mechanisms that may have permitted their survival in time [
16]. Additionally, breed standardization could be an important tool for the evaluation of birds within their flocks and determine certain measurements for the selection of the best animals [
17]. In this regard, morphometric and phaneroptic approaches may be fundamental in poultry management as they are fast and economically profitable [
18].
This information altogether enables the correct development and implementation of the administrative structures needed to guarantee the stability and future viability of breeds through the development conservation and breeding programs, as well as the sustainable commercialization of their products once censuses are enough.
In this context, this study aimed to determine the contribution of quantitative and qualitative morphological-related traits to the zoometric characterization through the development of a discriminant canonical analysis (DCA), as a tool that permits determining phenotypic variability in the Andalusian avian breeds and within their varieties, as a strategy to support the standardization of native breeds and implement conservation strategies that ensure the consolidation of local genotypes as recognized breeds.
2. Materials and Methods
2.1. Animals, Sample Size, and Distribution
Biometric data were collected from 608 adult birds (from 1 to 7 years old, 1.94 ± 0.75 years), 473 hens (77.80%), and 135 roosters (22.20%), belonging to different varieties of Utrerana and Sureña breeds, as described in
Figure 1. The sample size accounted for at least 20 times as many observations as variables. As this assumption was fulfilled, the study sample permitted to obtain reliable estimates of the canonical factor loadings for interpretation and to draw valid conclusions [
19].
The sample was collected at 16 farms across the seven provinces in Andalusia (Cádiz, Córdoba, Granada, Huelva, Jaén, Málaga, and Sevilla). All animals were reared under extensive backyard conditions.
National guidelines for the care and the use of laboratory and farm animals, and avian-specific codes for good practices were followed during the data collection. For this, standards consistent with European Union legislation (2010/63/EU, from 22 September 2010) as transposed into Spanish law (Royal Decree Law 53/2013, from 1 February 2013). The study protocol was submitted to The Ethics Committee of Animal Experimentation of the University of Córdoba (Spain) and deemed exempt from review.
2.2. Biometric Measurement Collection
Biometrical analysis was performed in each bird, measuring 27 quantitative and five qualitative variables, following the procedure for morphological characterization of native chicken breeds described in previous studies [
20,
21]. A summary of the quantitative biometric variables and how to measure them is shown in
Table 1. All corporal measurements were taken on the right side of the animal.
Figure 2 shows details of the head measurements taken. A suspended electronic scale (measurement precision = 5 g; Kern CH50K100, Kern & Sohn, Balingen, Germany), a Vernier scale (Electro DH M 60.205, Barcelona, Spain), and a tape measure were used for measurement collection.
The following qualitative traits were evaluated in the present study: eye color, beak color, presence or absence of spurs, tarsus color, and nail color. Moreover, skull ratio, ocular ratio, beak ratio, and tarsus ratio were computed, as shown in
Table 2.
2.3. Normality and Kruskall–Wallis Tests
The Shapiro–Francia W’ test (for 50 < n < 2500 samples) was used to discard gross violations of the normality assumption. The Shapiro–Francia W’ test was performed using the Shapiro–Francia normality routine of the test and distribution graphics package of the Stata Version 16.0 software (College Station, TX, USA). The normality test suggested normality assumption was not met. Hence, a nonparametric approach was followed. The Kruskal–Wallis H test was performed to detect differences in the median across sexes and genotypes. The Kruskal–Wallis H Test reported medians to significantly differ across all possibilities for sex and breed/variety combinations. Consequently, a separate DCA was performed for males and females.
2.4. Discriminant Canonical Analysis (DCA)
In the present research, 36 explanatory variables were used to perform the DCA: body weight, ornithological measurement, wingspan, skull length, skull width, ocular length, ocular width, beak length, beak width, comb length, comb width, number of spikes in the comb, earlobe length, earlobe width, wattle length, wattle width, neck length, back sternum length, tail length, thigh length, folding wing length, tarsus length, anteroposterior tarsus diameter, lateromedial tarsus diameter, eye color, beak color, presence or absence of spurs, tarsus color, nail color, skull ratio, ocular ratio, beak ratio, and tarsus ratio. In each sex, the breed and variety of the bird were used as classification criteria to measure the variability in morphological traits between and within the used classification groups and establish and outline population clusters [
22,
23].
The statistical analysis issued a set of discriminant functions that could be used as a tool to determine the clustering patterns described by the population sample through a linear combination of morphological-related traits. Furthermore, this canonical tool was used to plot pairs of canonical variables and graphically depict the group differences into an easily interpretable territorial map. Regularized forward stepwise multinomial logistic regression algorithms were used to perform the variable selection. Priors were regularized following the group sizes computed from the prior probability option in SPSS v26.0 software (IBM, Armonk, NY, USA), instead of considering them to be equal, thus preventing groups with different sample sizes from affecting the quality of the classification [
24].
Previous studies have reported DCA to be robust and its outputs to be consistent when sample sizes among groups were highly unequal. Potential distortion effects derived from unequal sample sizing can be palliated using at least 20 samples for every four or five predictors. Additionally, the maximum number of independent variables must be n − 2 (where n = simple size). The present design was developed aiming at meeting these requirements sufficiently, to ensure the validity of the conclusions drawn.
Before discriminant analysis, independence of regressors was ensured by multicollinearity analysis. The same variables were chosen by the forward and the backward stepwise selection methods. Hence, the progressive selection method was chosen as preferable since it is less time-consuming than the backward selection method.
The discriminant routine of the Classify package of SPSS v26.0 software (IBM, Armonk, NY, USA) and the discriminant analysis routine of the analyzing data package of XLSTAT 2014 (Pearson Edition) (Addinsoft, Paris, France) were used to perform the DCA.
2.4.1. Multicollinearity Preliminary Testing
Redundancies in the variables used were identified after performing the multicollinearity assumption before running the DCA. Multicollinearity analysis seeks to avoid the overinflation of the explanatory potential of variance due to the inclusion of an unnecessarily large number of variables. As an indicator of multicollinearity, the variance inflation factor was calculated using the following formula:
where
R2 is the coefficient of determination of the regression equation.
A recommended maximum
VIF value of 5 was used in the study, as suggested by Rogerson [
25]. Tolerance (1 −
R2) is the amount of variability in a certain independent variable that is not explained by the rest [
26]. When tolerance values are lower than 0 and, simultaneously,
VIF values ≥10, multicollinearity must be considered troublesome.
VIF was computed using the discriminant analysis routine of the analyzing data package of XLSTAT 2014 (Pearson Edition).
2.4.2. Canonical Correlation Dimension Determination
Pearson’s ρ was used to interpret canonical correlations. The maximum number of canonical correlations between two sets of variables is the number of variables in the smaller set. Although most of the relationships between different sets are explained by the first canonical correlation, all canonical correlations must be considered. Dimensions with canonical correlation values of ≥0.30 may be statistically significant.
2.4.3. Discriminant Canonical Analysis Efficiency
Wilks’ lambda test was used to evaluate variables that significantly contribute to the discriminant function. When Wilks’ lambda approximates to 0, the contribution of the variable to a discriminant function increases. The chi-square statistic was considered to test the significance of Wilks’ lambda. If the significance is below 0.05, the function can be concluded to adequately explain the group adscription [
27].
2.4.4. Discriminant Canonical Analysis Model Reliability
Pillai’s trace criterion was used in the discriminant function analysis to test the assumption of equal covariance matrices. This is the only acceptable test that must be used in cases of unequal sample sizes [
28]. Pillai’s trace criterion was calculated using the discriminant analysis routine of the analyzing data package of XLSTAT 2014 (Pearson Edition). A significance below 0.05 indicates significant statistical differences in the dependent variables across the levels of independence; hence, application of DCA is feasible.
2.4.5. Variable Dimensionality Reduction
A preliminary principal component analysis (PCA) was computed to minimize overall variables into few meaningful variables that contributed to the morphological characterization of males and females in different genotypes. PCA was performed automatically using the discriminant analysis routine of the analyzing data package XLSTAT 2014 (Pearson Edition) (Addinsoft, Paris, France).
2.4.6. Canonical Coefficient and Loading Interpretation and Spatial Representation
The percentage of allocation of an individual within its group (defined by its genotype) was calculated using a discriminant function analysis. Values ≥|0.40| in the discriminant loading of a variable were considered to be significantly discriminant. Thus, nonsignificant variables were excluded from the function using stepwise procedures. Higher values for absolute coefficients for each particular variable determine better discriminating power. Afterward, data were standardized following the premises reported by Manly and Alberto [
29], and Mahalanobis distances were calculated using the following formula:
where
D2ij is the distance between population
i and
j,
Υi and
Υj are the means of variable
x in the
i-th and
j-th populations, respectively, and
COV−1 is the inverse of the covariance matrix of measured variable
x. The squared Mahalanobis distance matrix was converted into a Euclidean distance matrix.
Afterward, dendrograms were built using the underweighted paired-group method arithmetic averages (UPGMA) from the Rovira i Virgili University, Tarragona, Spain, and the Phylogeny procedure of MEGA X 10.0.5 from the Institute of Molecular Evolutionary Genetics, The Pennsylvania State University, State College, PA, USA.
2.4.7. Discriminant Function Cross-Validation
The percentage of correctly classified cases can be defined as the hit ratio. The leave-one-out cross-validation procedure was used to consider if the discriminant functions can be validated. Classification accuracy is achieved when the classification rate is at least 25% higher than obtained by chance.
Press’s Q statistic can support these results since it can be used to compare the discriminating power of the cross-validated function, as follows:
where
n is the number of observations in the sample;
n’ is the number of observations correctly classified, and
K is the number of groups.
The value of the Press’s Q statistic must be compared with the critical value of 6.63 for χ2 with a degree of freedom in a significance of 0.01. When Press’s Q exceeds the critical value of χ2 = 6.63, the cross-validated classification can be regarded as significantly better than chance.
2.5. Data Mining CHAID Decision Tree
The chi-squared automatic interaction detection (CHAID) decision tree (DT) data mining method was used for classification, prediction, interpretation, and discrete categorized data manipulation. The tree routine of the Classify package of SPSS v26.0 software (IBM, Armonk, NY, USA) was used. Each internal node was built in the tree around a zoometric or phaneroptic trait (input variables), while a chi-squared test significance split criterion (p < 0.05 at least) was fulfilled in the so-called pre-pruning process.
Breiman, et al. [
30] suggested that pre- or post-pruning methods prevent over-dimension of trees to prevent the failure to pursue the addition of traits (branches) which add significantly to the overall fit. As a result, a tree that exhaustively depicts the significant relationships across independent variables is one from which those nodes that do not contribute to the overall prediction have been discarded. Furthermore, CHAID additionally penalizes model complexity. In this regard, the Bonferroni inequality significant adjustment for significance levels was used.
Breiman’s method uses chi-squared tests to determine to configure the tree building process. Each branch represents an outcome of the test (in a number of two or more), and each leaf node (or terminal node) represents a category level of the target variable (breed/variety). The root node in the tree is the one that is located at the top. The decisions are made at each node, and each record of data continues through the tree along a path until the record reaches a leaf or terminal node of the tree [
31].
Afterward, cross-validation was performed to validate the set of predictors considered measuring the differences between the prediction error for a tree applied to a new sample and a training sample. Cross-validation of the decision tree was performed using the “complexity parameter” and cross-validated error to estimate how accurately the model performs data prediction. Tenfold cross-validation [
32] was performed using every sample record in the training sample and study data. The resubstitution error rate measures the proportion of original observations that were misclassified by various subsets of the original tree.
Tenfold cross-validation was used to obtain a cross-validated error rate, from which the optimal tree was selected to prevent bias and outlier overfitting. Tenfold cross-validation involves creating 10 random subsets of the original data, setting one portion aside as a test set, constructing a tree for the remaining (10 − 1) portions, and evaluating the tree using the test portion. This was repeated for all portions, and an estimate of the error was evaluated. Adding up the error across the 10 portions represented the cross-validated error rate. Afterward, the tree yielding the lowest cross-validated error rate was selected as the tree that best fit the data.
4. Discussion
Differential sex-linked hormonal and genetic regulation patterns of the expression of growth have been reported to occur in local poultry breeds [
35,
36]. Dimorphism and dichromatism could be a consequence of sexual selection and might provide an adaptative advantage of one population over others. For instance, in the context of the conditions found in rustic backyard environments, even if there is a lower selective pressure focused toward production, male-to-male competition has induced roosters to increase the size, giving an advantage against the opponent [
37].
In the context of multizoometric and phaneroptic analyses, it has been suggested that it is necessary to check for the different relationships across explanatory variables and select independent variables that do not overlap when deciding on the factors which determine the efficiency of predictive models [
23]. High correlations between skull length and skull width (i.e., skull ratio) were revealed by the multicollinearity analysis since the formula for skull ratio calculation comprises the aforementioned measurements. The same happened with anteroposterior (in both sexes) and lateromedial (only in hens) tarsus diameters as the elements which determine the tarsus ratio. The calculation formula of beak ratio, which includes the remaining beak measurements, was eliminated from further analysis due to multicollinearity problems (
VIF > 5).
Lastly, the ocular width variable was discarded from the analysis of male individuals since this variable is contained within the formula of ocular ratio (
VIF > 5). These results are supported by those in Ning et al. [
38], who found multicollinearity problems when formulae were developed after the inclusion of explanatory variables which were already included.
Phaneroptic variables have been reported to be highly significantly interrelated [
39]. Even if most qualitative variables were discarded after the multicollinearity analysis, nail color in hen and beak color in roosters were the only qualitative variables that remained in the DCA. Thus, results suggest that multicollinearity problems between different qualitative measurements in birds may have occurred.
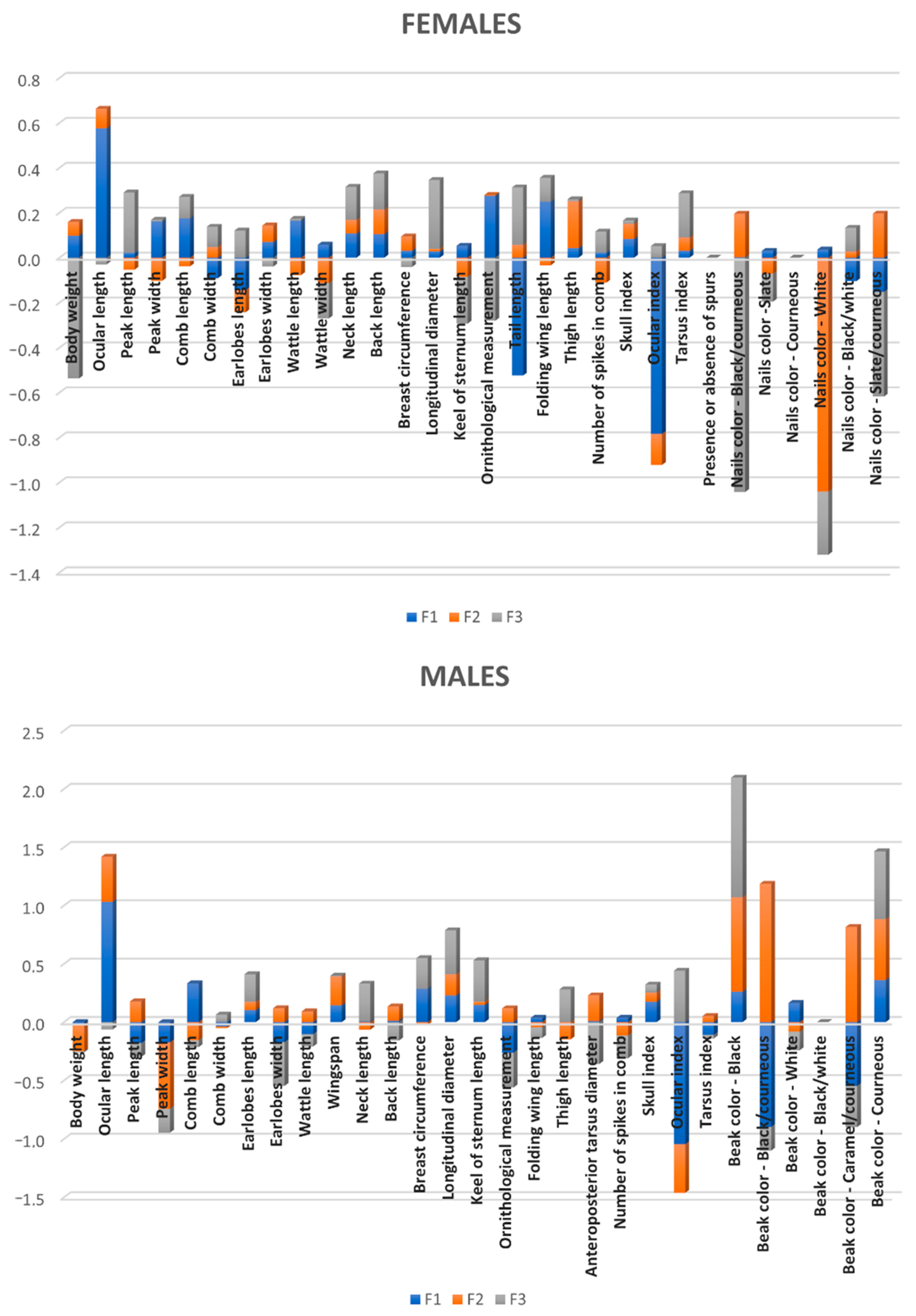
White nails was reported to be the best discriminating feature in hens (
Table 6). Only seven individuals of White, Splash, and Franciscan Sureña showed dark nails, while no hen of White and Franciscan varieties showed nails of a different color than white. In roosters, black/corneous and white colors in the beak were also reported to have high discriminant power.
Previous studies have reported that phaneroptic features are somehow correlated in native chicken breeds, provided they may derive from the expression of the same gene background across the body parts [
40]. Additionally, it has been suggested that these qualitative traits have significant effects on other quantitative traits such as body weight and daily gain in chicken [
40,
41].
Our results are indicative of the fact that qualitative variables, with high discriminant ability to discern among local hen genotypes, must be considered as efficient selection criteria in breeding programs, as an effective method to identify the individuals presenting the most desirable production-related characteristics at the most convenient earlier age.
Furthermore, certain phaneroptic variables may be associated with consumers’ trends and their cultural preferences. For instance, while North American consumers have strong preferences for white-skin meat [
42], meat from dark-skin poultry is preferred by producers and consumers in South America [
14]. Hence, multivariety breeds accounting for a wide variety of feather and skin color patterns such as Utrerana and Sureña could satisfy the needs of a wider scope of targets in different market niches.
Feather coloration strongly conditions the camouflage abilities of birds. In this regard, Dohner [
43] suggested that the less aggressive strains developed for confinement may be less self-sufficient and may not be as alert to predators. In hens, this has been ascribed to the association of specific quantitative trait loci with behavioral traits [
44]. As an example, birds carrying the ancestral junglefowl allele (i) of the
PMEL17 locus are black, while White Leghorn (I) birds are white (with heterozygotes frequently being less pigmented).
Contextually,
i/
i alleles carriers have been reported to be more vocal, less prone to develop fearful attitudes toward humans, and more aggressive, social, and explorative (enhanced foraging behavior) [
44]. These enhanced behavioral features may make these dark-colored breeds less susceptible to predation by hawks [
43]. The
PMEL17 locus has simultaneously been associated with feather-pecking and bullying behavior toward counterparts [
45], with darker birds tending to be rather affected by feather-pecking than their white counterparts [
46]. It is still unknown whether feather-pecking may exclusively be attributed to plumage color or to the behavior of
i/
i carrier individuals to become targets of pecking attacks.
Alternatively, Tickell [
47] stated that coloration-related costs in higher rates of bird predation may also translate into the enhancement of other tactics for evading capture [
6]. This was reflected in our study (
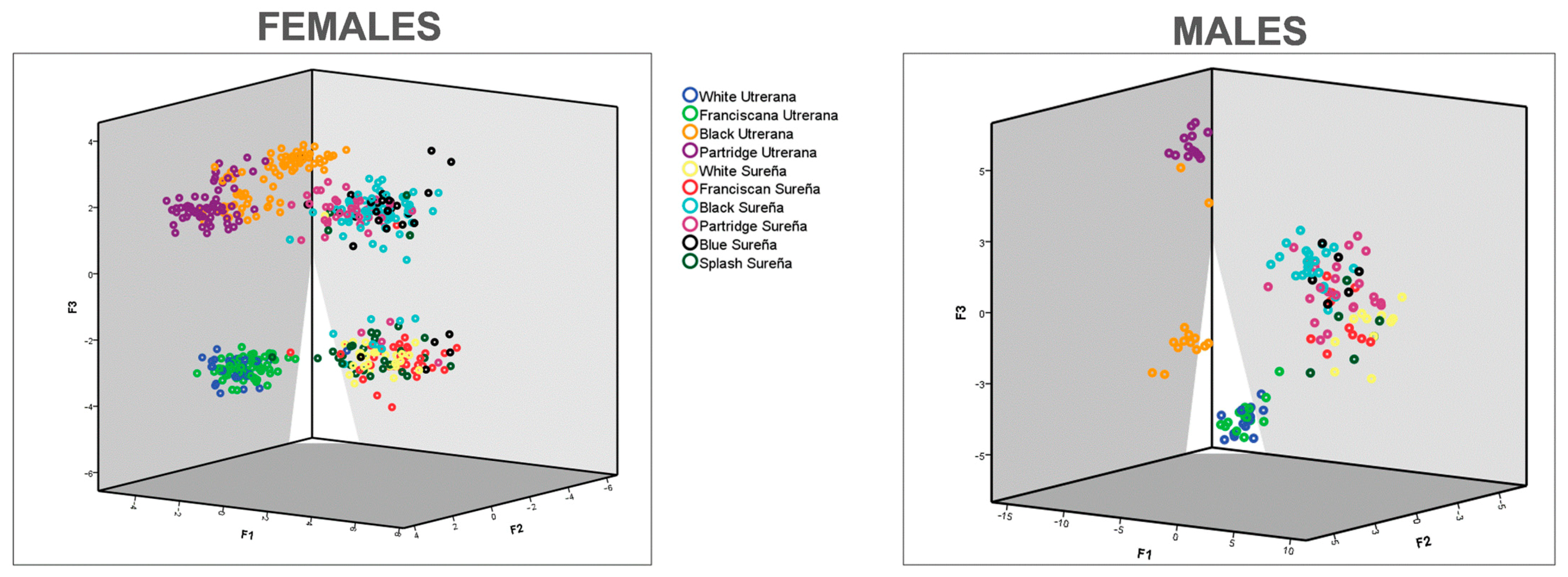
Figure 5 and
Figure 6) with Sureña presenting smaller ocular indices in comparison to Utrerana hens, albeit with darker Sureña individuals being closer to white Utrerana animals and white-feathered Sureña located further away when morphological traits were considered.
Ocular ratio was ranked second and first regarding its discriminant ability in hens and roosters, respectively. The relevance of ocular ratio may be ascribed to higher adaptability to the environment and improved capacity to seek food as a result of improved vision skills. Indeed, except for certain occasions, birds have a highly developed vision.
In relationship to the size of the skull, the avian eye is very large. While humans have an eye relative size of 5% with respect the skull, in hens, 50% of the cranial volume is occupied by the orbit [
48]. High visual acuity is advantageous for hens relying heavily on their ability to navigate surroundings to find and acquire food, to identify potential mates, and to quickly escape from predators [
49,
50]. Hall and Ross [
51] reported that the light level, which is highly correlated with bird activity pattern, has a more significant influence on eye shape and body size than other factors, such as phylogeny.
Birds with a higher adaptation to darkness habits, such as brooding and nesting abilities, exhibit larger axial and corneal lengths and, therefore, a higher eye size diameter than the rest of the birds [
52,
53]. On the other hand, larger individuals with larger eyes have the potential for more sensitive and acute vision than smaller individuals with smaller eyes. This could suggest that the Sureña breed, with a significantly larger eye size, has a sharper vision. However, each breed has developed an ideal eye design for conditions in which it is produced. Larger eyes need more brain space for information processing. Therefore, evaluation of ocular size in each breed must be performed taking into account body size [
54]. Thus, the higher size of Sureña eyes could be mainly ascribed to a proportionally larger body shape.
It has also been suggested that lower values for ocular ratio may act as an adaptation to optimal antipredator behavior since larger ocular width could suppose an advantage in the lateral visual field [
55,
56]. Thus, results obtained in the present study may suggest that Utrerana eyes make it more adapted to survival in free-range systems. Furthermore, smaller birds have developed rather improved adaptative qualities such as hardiness, agility, scavenging ability, and less time needed for flight [
57]. The Utrerana breed, with lower body weight and ocular ratio, may be better adapted to free-range systems through its enhanced rusticity, even if the literature indicates that both breeds can easily thrive and are well adapted to the environmental conditions present in these alternative production systems [
9,
58].
Back length was the third best discriminant variable in hens. These results agree with those presented by previous research [
59,
60]. In this sense, back length has been reported to be highly correlated with other important traits. As a consequence, it plays an important role as a linear body measurement when the aim is to predict for body weight, as well as to develop and to implement productive selection strategies during breeding in laying hens.
Size-related parameters such as body weight (in hens) and wingspan (in roosters) play a pivotal role in the classification of individuals (
Table 6 and
Table 7). These traits allow us to delimitate those animals belonging to the Sureña breed. Sureña individuals typically account for larger body sizes than Utrerana individuals.
Lighter hens have been reported to present higher egg productions and lower feed conversion rates and, therefore, a better laying ability [
61]. On the other hand, breeds characterized by larger individuals may be prone to become dual-purpose genotypes in alternative production systems, in which both sexes are reared together, to later, at an advanced age, separate males for final fattening and slaughtering, while females are kept during several laying cycles [
62,
63]. Bearing this in mind, focusing efforts on the selection of the Utrerana breed toward an egg production aptitude and Sureña as a dual-purpose breed may be the most effective and profitable productive alternative.
Although Sureña and Utrerana breeds were presumably selected from a common origin [
8], the graphic representation of the observations assessed in the present study (
Figure 4) reports a clear differentiation of morphological characteristics between the two breeds. While three clear clusters are shown in Utrerana breed (Partridge, Black, and Franciscan/White varieties), the closeness of the six varieties of the Sureña avian breed suggests a likely lack of reproductive management and crossbreeding among the different varieties of this breed.
This proves that, once official breed recognition occurs, an incorrect application of a breeding program in local breeds can lead to a deterioration of the phenotypic and genotypic identity of the individuals, which directly results in the partial or total loss of the genetic pool of these local resources [
64,
65].
Contextually, Partridge Utrerana was reported to be the most differentiated variety from all studied varieties. These results are supported by those in Macrì et al. [
6], who reported Partridge Utrerana individuals to be placed the farthest away from the rest of Utrerana varieties.
More than 75% of hens in each Utrerana variety were correctly classified (
Supplementary Table S3), except for the individuals of the White variety, whereby 50% of hens were notably classified as Franciscan Utrerana hens. This Utrerana White/Franciscan misclassification is supported by the results in
Figure 5 and
Figure 6. Franciscan and White Utrerana varieties were closely clustered (
Figure 5 and
Figure 6). This finding may indirectly indicate reminiscences of hybridization between White and Franciscan Utrerana varieties, with both presenting white legs and beak, which may be the result of the attempts of breeders to decrease the consanguinity within the White Utrerana variety, given that this variety has historically been the subpopulation accounting for the smallest census and that which faces the highest endangerment risk [
22].
Blue Sureña variety females were those for which a rather frequent misclassification rate occurred (
Supplementary Table S3). This finding may stem from the fact that breeding practices performed in the area may seek the obtention of individuals presenting blue plumage patterns through a cross between other varieties, such as Black or Splash [
66].
Biometric studies have been performed worldwide to make breed characterization feasible and to be considered during the implementation of conservation strategies and policies [
16]. This suggests that the preservation of the breed diversity may be one of the motor elements ensuring the future survival of a breed. This future survival may rely on the enhancement of a breed’s ability to cover a wider scope of market demands, thereby reaching a broader audience [
67]. The present methodological proposal is framed in the context of opportunity and resurgence of a potential production industry that intends to lay the base for a sustainable selective breeding program in avian breeds. Certain easily measurable traits, such as phaneroptic variables and ocular ratio, can efficiently play a pivotal role in the classification of birds. In this context, the discriminant tool designed in the present research allows efficiently classifying individuals considering biometric and phaneroptic traits. This is supported by the 71.82% and 81.48% of individuals correctly ascribed to their prior hen breed/variety cluster.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}