
Screening of New Potential Probiotics Strains against Photobacterium damselae Subsp. piscicida for Marine Aquaculture

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation of Potential Probiotic Strains
2.2. Antagonistic Effect of Potential Probiotic Strains against Photobacterium damselae Subsp. piscicida
2.3. Production of Antibacterial Substances of Probiotics against Pathogens
2.4. Fish Bile and pH Resistance
2.5. Adhesion in Intestinal and Skin Mucus
2.6. Growth in Intestinal and Skin Mucus
2.7. Probiotic Strain Safety
2.8. Potential Probiotic Administration in Experimental Diet
2.9. Analysis of Gene Expression

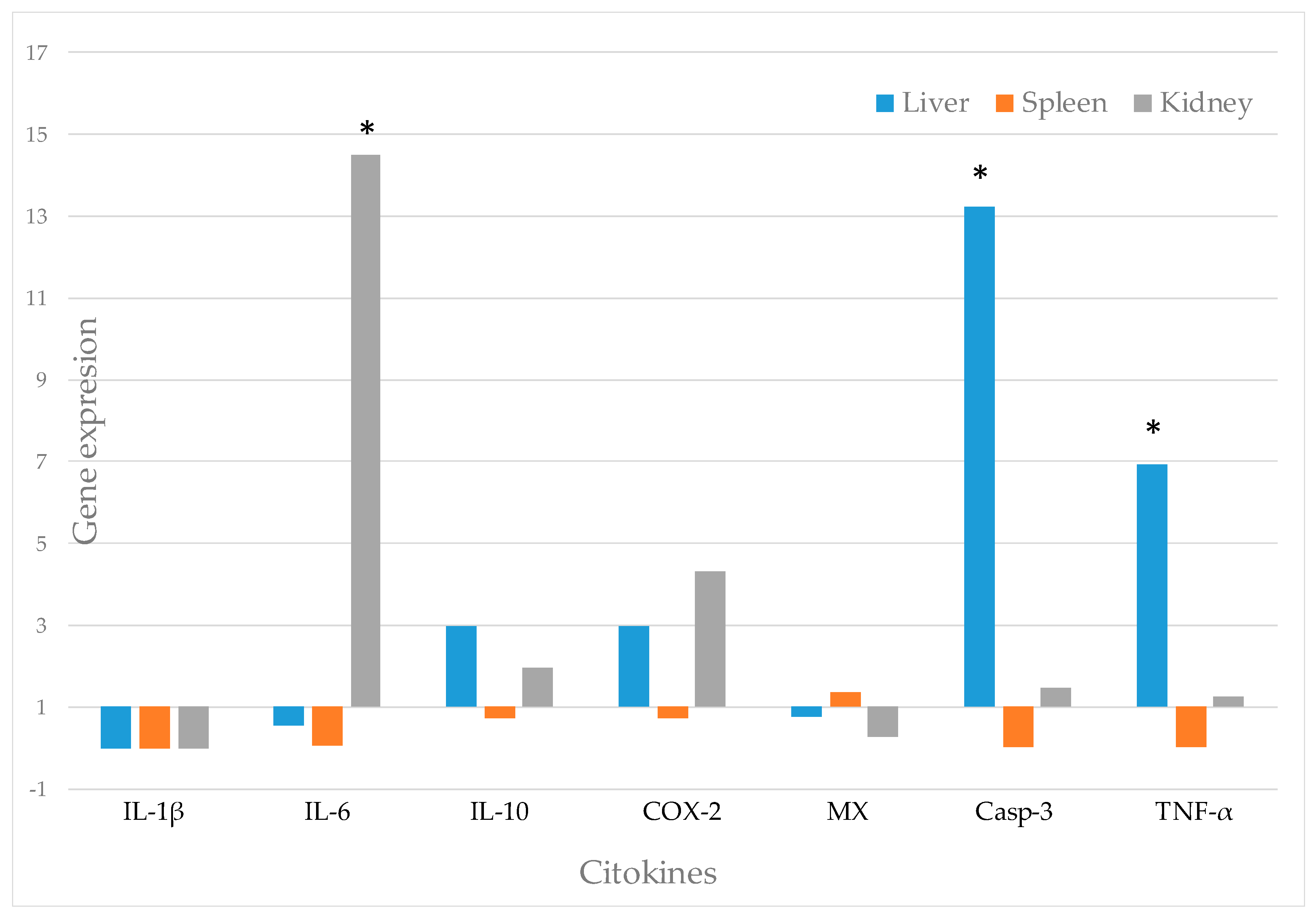{kind=link}
| Gene | Primer Sequences | Hybridization Temperature | Source |
|---|---|---|---|
| β-actin | Forward 5′ATGTGGATCAGCAAGCAGG-3′ | 57.7 °C | AJ537421 Genne runer |
| Reverse 5′AGAAATGTGTGGTGTGGTCG-3′ | |||
| IL-1β | Forward 5′-ATTACCCACCACCCACTGAC-3′ | 57.7 °C | AJ269472 Genne runer |
| Reverse 5′-TCTCTTCCACTATGCTCTCCAG-3′ | |||
| IL-6 | Forward 5′-ACTTCCAAAACATGCCCTGA-3′ | 59.3 °C | AM490062 (Sepulcre et al., 2007) [22] |
| Reverse 5′- CCGCTGGTCAGTCTAAGGAG-3′ | |||
| IL-10 | Forward 5′-ACCCCGTTCGCTTGCCA-3′ | 59.3 °C | AM268529 (Picchietti et al., 2009) [23] |
| Reverse 5′-CATCTGGTGACATCACTC-3′ | |||
| Casp-3 | Forward 5’-ACGAAGCAGGTCAATCATCC-3’ | 59.3 °C | DQ345774 Genne runer |
| Reverse 5’-GCAGTTTAAGGGTATCCAGAGC-3’ | |||
| TNF-α | Forward 5′-GCCAAGCAAACAGCAGGAC-3′ | 60 °C | DQ200910 Genne runer |
| Reverse 5′-ACAGCGGATATGGACGGTG-3′ | |||
| COX-2 | Forward 5′-AGCACTTCACCCACCAGTTC-3′ | 59.3 °C | AJ630649 (Sepulcre et al., 2007) [22] |
| Reverse 5′-AAGCTTGCCATCCTTGAAGA-3′ | |||
| Mx | Forward 5′-GGTCAAGGAGCAGATCAAACAG-3′ | 57.7 °C | AM228974 Genne runer |
| Reverse 5′-CTCGCATCAGGTTAGGGAATC-3′ |
2.10. Statistical Analysis
3. Results
3.1. Antagonistic Effect of Potential Probiotic Strains
3.2. Production of Antibacterial Substances
3.3. Fish Bile and pH Resistance
3.4. Adhesion in Intestinal and Skin Mucus
3.5. Growth in Intestinal and Skin Mucus
3.6. Probiotic Strain Safety
3.7. Analysis of Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leong, J.C.; Fryer, J.L. Viral vaccines for aquaculture. Ann. Rev. Fish Dis. 1993, 3, 225–240. [Google Scholar] [CrossRef]
- Sica, M.G.; Brugnoni, L.I.; Marucci, P.L.; Cubitto, M.A. Characterization of probiotic properties of lactic acid bacteria isolated from an estuarine environment for application in rainbow trout (Oncorhynchus mykiss, Walbaum) farming. Antonie van Leeuwenhoek 2012, 101, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Kesarcodi-Watson, A.; Kaspar, H.; Lategan, M.J.; Gibson, L. Probiotics in aquaculture: The need, principles and mechanisms of action and screening processes. Aquaculture 2008, 274, 1–14. [Google Scholar] [CrossRef]
- Ringø, E.; Løvmo, L.; Kristiansen, M.; Bakken, Y.; Salinas, I.; Myklebust, R.; Olsen, R.E.; Mayhew, T.M. Lactic acid bacteria vs. pathogens in the gastrointestinal tract of fish: A review. Aquac. Res. 2010, 41, 451–467. [Google Scholar] [CrossRef]
- Toranzo, A.E.; Magariños, B.; Romalde, J.L. A review of the main bacterial fish diseases in mariculture systems. Aquaculture 2004, 246, 37–61. [Google Scholar] [CrossRef]
- Hagi, T.; Hoshino, T. Screening and characterization of potential probiotic lactic acid bacteria from cultured common carp intestine. Biosci. Biotechnol. Biochem. 2009, 73, 1479–1483. [Google Scholar] [CrossRef] [Green Version]
- Abdelkhalek, N.K.; Eissa, I.A.; Ahmed, E.; Kilany, O.E.; El-Adl, M.; Dawood, M.A.O.; Hassan, A.M.; Abdel-Daim, M.M. Protective role of dietary Spirulina platensis against diazinon-induced Oxidative damage in Nile tilapia; Oreochromis niloticus. Environ. Toxicol. Pharmacol. 2017, 54, 99–104. [Google Scholar] [CrossRef]
- Yan, J.; Guo, C.; Dawood, M.A.O.; Gao, J. Effects of dietary chitosan on growth, lipid metabolism, immune response and antioxidant-related gene expression in Misgurnus anguillicaudatus. Benef. Microbes 2017, 8, 439–449. [Google Scholar] [CrossRef]
- Sorroza, L.; Padilla, D.; Acosta, F.; Román, L.; Grasso, V.; Vega, J.; Real, F. Characterization of the probiotic strain Vagococcus fluvialis in the protection of European sea bass (Dicentrarchus labrax) against vibriosis by Vibrio anguillarum. Vet. Microbiol. 2012, 155, 369–373. [Google Scholar] [CrossRef]
- Etyemez, M.; Balcázar, J.L. Isolation and characterization of bacteria with antibacterial properties from Nile tilapia (Oreochromis niloticus). Res. Vet. Sci. 2016, 105, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Balcázar, J.L.; Vendrell, D.; De Blas, I.; Ruiz-Zarzuela, I.; Gironés, O.; Múzquiz, J.L. In vitro competitive adhesion and production of antagonistic compounds by lactic acid bacteria against fish pathogens. Vet. Microbiol. 2007, 122, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, G.; Ray, A.K. The advancement of probiotics research and its application in fish farming industries. Res. Vet. Sci. 2017, 115, 66–77. [Google Scholar] [CrossRef]
- Lazado, C.C.; Caipang, C.M.A. Mucosal immunity and probiotics in fish. Fish Shellfish Immunol. 2014, 39, 78–89. [Google Scholar] [CrossRef]
- Austin, B.; Baudet, E.; Stobie, M. Inhibition of bacterial fish pathogens by Tetraselmis suecica. J. Fish Dis. 1992, 15, 55–61. [Google Scholar] [CrossRef]
- Croxatto, A.; Prod’hom, G.; Greub, G. Applications of MALDI-TOF mass spectrometry in clinical diagnostic microbiology. FEMS Microbiol. Rev. 2012, 36, 380–407. [Google Scholar] [CrossRef]
- Bizzini, A.; Greub, G. Matrix-assisted laser desorption ionization time-offlight mass spectrometry, a revolution in clinical microbial identification. Clin. Microbiol. Infect. 2010, 16, 1614–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikoskelainen, S.; Salminen, S.; Bylund, G.; Ouwehand, A.C. Characterization of the properties of human- and dairy-derived probiotics for prevention of infectious diseases in fish. Appl. Environ. Microbiol. 2001, 67, 2430–2435. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Austin, B. Characterization of probiotic carnobacteria isolated from rainbow trout (Oncorhynchus mykiss) intestine. Lett. Appl. Microbiol. 2008, 47, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Olsson, J.C.; Westerdahl, A.; Conway, P.L.; Kjelleberg, S. Intestinal colonization potential of turbot (Scophthalmus maximus)- and dab (Limanda limanda)-associated bacteria with inhibitory effects against Vibrio anguillarum. Appl. Environ. Microbiol. 1992, 58, 551–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irianto, A.; Austin, B. Use of probiotic to control furunculosis in rainbow trout (Oncorhynchus mykiss). J. Fish Dis. 2002, 25, 333–342. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sepulcre, M.P.; Sarropoulou, E.; Kotoulas, G.; Meseguer, J.; Mulero, V. Vibrio anguillarum evades the immune response of the bony fish sea bass (Dicentrarchus labrax L.) through the inhibition of leukocyte respiratory burst and down-regulation of apoptotic caspases. Mol. Immunol. 2007, 44, 3751–3757. [Google Scholar] [CrossRef] [PubMed]
- Picchietti, S.; Fausto, A.M.; Randelli, E.; Carnevali, O.; Taddei, A.R.; Buonocore, F.; Scapigliati, G.; Abelli, L. Early treatment with Lactobacillus delbrueckii strain induces an increase in intestinal T-cells and granulocytes and modulates immune-related genes of larval Dicentrarchus labrax (L.). Fish Shellfish Immunol. 2009, 26, 368–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardones, F.O.; Paredes, F.; Medina, M.; Tello, A.; Valdivia, V.; Ibarra, R.; Correa, J.; Gelcich, S. Identification of research gaps for highly infectious diseases in aquaculture: The case of the endemic Piscirickettsia salmonis in the Chilean salmon farming industry. Aquaculture 2018, 482, 211–220. [Google Scholar] [CrossRef]
- Serrano, P.H. Responsible Use of Antibiotics in Aquaculture; Food & Agriculture Organization: Rome, Italy, 2005; p. 469. [Google Scholar]
- Suzuki, S.; Pruden, A.; Virta, M.; Zhang, T. Antibiotic resistance in aquatic systems. Front. Microbiol. 2017, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef]
- Cheng, G.; Hao, H.; Xie, S.; Wang, X.; Dai, M.; Huang, L.; Yuan, Z. Antibiotic alternatives: The substitution of antibiotics in animal husbandry? Front. Microbiol. 2014, 5, 217. [Google Scholar] [CrossRef] [Green Version]
- Defoirdt, T.; Sorgeloos, P.; Bossier, P. Alternatives to antibiotics for the control of bacterial disease in aquaculture. Curr. Opin. Microbiol. 2011, 14, 251–258. [Google Scholar] [CrossRef]
- Hjelm, M.; Bergh, Ø.; Riaza, A.; Nielsen, J.; Melchiorsen, J.; Jensen, S.; Duncan, H.; Ahrens, P.; Birkbeck, H.; Gram, L. Selection and identification of autochthonous potential probiotic bacteria from Turbot larvae (Scophthalmus maximus) rearing units. Syst. Appl. Microbiol. 2004, 27, 360–371. [Google Scholar] [CrossRef] [Green Version]
- Makridis, P.; Martins, S.; Vercauteren, T.; Van Driessche, K.; Decamp, O.; Dinis, M.T. Evaluation of candidate probiotic strains for gilthead sea bream larvae (Sparus aurata) using an in vivo approach. Lett. Appl. Microbiol. 2005, 40, 274–277. [Google Scholar] [CrossRef] [Green Version]
- Nayak, S.K. Probiotics and immunity: A fish perspective. Fish Sellfish Immunol. 2010, 29, 2–14. [Google Scholar] [CrossRef]
- Robertson, P.A.W.; O’Dowd, C.; Burrells, C.; Williams, P.; Austin, B. Use of Carnobacterium sp. as a probiotic for Atlantic salmon (Salmo salar L.) and rainbow trout (Oncorhynchus mykiss, Walbaum). Aquaculture 2000, 185, 235–243. [Google Scholar] [CrossRef]
- Nikoskelainen, S.; Ouwehand, A.C.; Bylund, G.; Salminen, S.; Lilius, E. Immune enhancement in rainbow trout (Oncorhynchus mykiss) by potential probiotic bacteria (Lactobacillus rhamnosus). Fish Shellfish Immunol. 2003, 15, 443–452. [Google Scholar] [CrossRef]
- Nikolopoulou, D.; Moutou, K.; Fountoulaki, E.; Venou, B.; Adamidou, S.; Alexis, M. Patterns of gastric evacuation, digesta characteristics and pH changes along the gastrointestinal tract of gilthead sea bream (Sparus aurata L.) and European sea bass (Dicentrarchus labrax L.). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2011, 158, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Fjellheim, A.J.; Klinkenberg, G.; Skjermo, J.; Aasen, I.M.; Vadstein, O. Selection of candidate probionts by two different screening strategies from Atlantic cod (Gadus morhua L.) larvae. Vet. Microbiol. 2010, 144, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Vine, N.G.; Leukes, W.D.; Kaiser, H. Probiotics in marine larviculture. FEMS Microbiol. Rev. 2006, 30, 404–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Grześkowiak, Ł.; Salminen, S. Probiotic strains and their combination inhibit in vitro adhesion of pathogens to pig intestinal mucosa. Curr. Microbiol. 2007, 55, 260–265. [Google Scholar] [CrossRef]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef] [Green Version]
- Merrifield, D.L.; Dimitroglou, A.; Foey, A.; Davies, S.J.; Baker, R.T.M.; Bøgwald, J.; Castex, M.; Ringø, E. The current status and future focus of probiotic and prebiotic applications for salmonids. Aquaculture 2010, 302, 1–18. [Google Scholar] [CrossRef]
- Sharifuzzaman, S.M.; Abbass, A.; Tinsley, J.W.; Austin, B. Subcellular components of probiotics Kocuria SM1 and Rhodococcus SM2 induce protective immunity in rainbow trout (Oncorhynchus mykiss, Walbaum) against Vibrio anguillarum. Fish Shellfish Immunol. 2011, 30, 347–353. [Google Scholar] [CrossRef]
- Sharifuzzaman, S.M.; Austin, B. Development of protection in rainbow trout (Oncorhynchus mykiss, Walbaum) to Vibrio anguillarum following use of the probiotic Kocuria SM1. Fish Sellfish Immunol. 2010, 29, 212–216. [Google Scholar] [CrossRef]
- Sharifuzzaman, S.M.; Austin, B. Influence of probiotic feeding duration on disease resistance and immune parameters in rainbow trout. Fish Shellfish Immunol. 2009, 27, 440–445. [Google Scholar] [CrossRef]
- Panigrahi, A.; Kiron, V.; Satoh, S.; Hirono, I.; Kobayashi, T.; Sugita, H.; Puangkaew, J.; Aoki, T. Immune modulation and expression of cytokine genes in rainbow trout Oncorhynchus mykiss upon probiotic feeding. Dev. Comp. Immunol. 2007, 31, 372–382. [Google Scholar] [CrossRef]
- Pérez-Sánchez, T.; Balcázar, J.L.; Merrifield, D.L.; Carnevali, O.; Gioacchini, G.; de Blas, I.; Ruiz-Zarzuela, I. Expression of immune-related genes in rainbow trout (Oncorhynchus mykiss) induced by probiotic bacteria during Lactococcus garvieae infection. Fish Sellfish Immunol. 2011, 31, 196–201. [Google Scholar] [CrossRef]
- Hasan, M.T.; Jang, W.J.; Lee, B.J.; Kim, K.W.; Hur, S.W.; Lim, S.G.; Bai, S.C.; Kong, I.S. Heat-killed Bacillus sp. SJ-10 probiotic acts as a growth and humoral innate immunity response enhancer in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2019, 88, 424–431. [Google Scholar] [CrossRef]
- Román, L.; Acosta, F.; Padilla, D.; El Aamri, F.; Bravo, J.; Vega, B.; Rodriguez, E.; Vega, J.; Déniz, S.; Real, F. The in vitro immunomodulatory effect of extracellular products (ECPs) of Vagococcus fluvialis L21 on European sea bass (Dicentrarchus labrax) leucocytes. Fish Sellfish Immunol. 2015, 42, 517–521. [Google Scholar] [CrossRef]
- Nurmi, J.T.; Puolakkainen, P.A.; Rautonen, N.E. Bifidobacterium lactis sp. 420 upregulates cyclooxygenase (Cox)-1 and down-regulates Cox-2 gene expression in a Caco-2 cell culture model. Nutr. Cancer 2005, 51, 83–92. [Google Scholar] [CrossRef]
- Cassatella, M.A.; Meda, L.; Gasperini, S.; Calzetti, F.; Bonora, S. Interleukin (IL-10) upregulates IL-1 receptor antagonist production from lipopolysaccharide-stimulated human polymorphonuclear leukocytes by delaying mRNA degradation. J. Exp. Med. 1994, 179, 1695–1699. [Google Scholar] [CrossRef]
- Raida, M.K.; Buchmann, K. Development of adaptive immunity in rainbow trout, Oncorhynchus mykiss (Walbaum) surviving an infection with Yersinia ruckeri. Fish Shellfish Immunol. 2008, 25, 533–541. [Google Scholar] [CrossRef]
- Feng, J.; Chang, X.; Zhang, Y.; Yan, X.; Zhang, J.; Nie, G. Effects of Lactococcus lactis from Cyprinus carpio L. as probiotics on growth performance, innate immune response and disease resistance against Aeromonas hydrophila. Fish Shellfish Immunol. 2019, 93, 73–81. [Google Scholar] [CrossRef]
- Yi, Y.; Zhang, Z.; Zhao, F.; Liu, H.; Yu, L.; Zha, J.; Wang, G. Probiotic potential of Bacillus velezensis JW: Antimicrobial activity against fish pathogenic bacteria and immune enhancement effects on Carassius auratus. Fish Shellfish Immunol. 2018, 78, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S. Apoptosis by Death Factor. Cell 1997, 88, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Caipang, C.M.A.; Hirono, I.; Aoki, T. Modulation of the early immune response against viruses by a teleostean interferon regulatory factor-1 (IRF-1). Comp. Biochem. Physiol. Part A 2009, 152, 440–446. [Google Scholar] [CrossRef]
- Acosta, F.; Lockhart, K.; Gahlawat, S.K.; Real, F.; Ellis, A.E. Mx expression in Atlantic salmon (Salmo salar L.) parr in response to Listonella anguillarum bacterin, lipopolysaccharide and chromosomal DNA. Fish Shellfish Immunol. 2004, 17, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.; Acosta, F.; Padilla, D.; Grasso, V.; Real, F. Mx expression in gilthead sea bream (Sparus aurata L.) in response to poly I: C, bacterial LPS and chromosomal DNA: Preliminary study. Fish Shellfish Immunol. 2011, 31, 170–172. [Google Scholar] [CrossRef]

| Pathogenic Strains | Reference | Source |
|---|---|---|
| Ph. damselae subsp. piscicida C2 | IUSA | Sparus aurata |
| Ph. damselae subsp. piscicida 17911 | ATCC | Perca fluviatilis |
| Ph. damselae subsp. piscicida DI21 | ATCC | Sparus aurata |
| Ph. damselae subsp. piscicida 94/99 | IUSA | Sparus aurata |
| Ph. damselae subsp. piscicida EP04 | IUSA | Sparus aurata |
| Pathogenic Strains Tested | Potential Probiotic Strains | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| Ph. damselae subsp. piscicida C2 | + | - | - |
| Ph. damselae subsp. piscicida ATCC 17911 | + | - | - |
| Ph. damselae subsp. piscicida DI21 | + | - | - |
| Ph. damselae subsp. piscicida 94/99 | + | - | - |
| Ph. damselae subsp. piscicida EP04 | + | + | + |
| pH | Potential Probiotic Strains | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| pH 7 | 100 | 100 | 100 |
| pH 6 | 91.6 ± 3.3 | 71.8 * ± 2.2 | 85.7 ± 3.5 |
| pH 5 | 74.3 * ± 4.5 | 41.5 * ± 3.5 | 65.5 * ± 1.2 |
| pH 4 | 50.4 * ± 10.6 | 25.2 * ± 2.7 | 55.6 * ± 0.8 |
| pH 3 | 6.3 * ± 3.8 | 4.9 * ± 1.8 | 11.5 * ± 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez Falcón, A.; Padilla, D.; Real, F.; Ramos Sosa, M.J.; Acosta-Hernández, B.; Sánchez Henao, A.; García-Álvarez, N.; Rosario Medina, I.; Silva Sergent, F.; Déniz, S.; et al. Screening of New Potential Probiotics Strains against Photobacterium damselae Subsp. piscicida for Marine Aquaculture. Animals 2021, 11, 2029. https://doi.org/10.3390/ani11072029
Gutiérrez Falcón A, Padilla D, Real F, Ramos Sosa MJ, Acosta-Hernández B, Sánchez Henao A, García-Álvarez N, Rosario Medina I, Silva Sergent F, Déniz S, et al. Screening of New Potential Probiotics Strains against Photobacterium damselae Subsp. piscicida for Marine Aquaculture. Animals. 2021; 11(7):2029. https://doi.org/10.3390/ani11072029
Chicago/Turabian StyleGutiérrez Falcón, Ana, Daniel Padilla, Fernando Real, María José Ramos Sosa, Begoña Acosta-Hernández, Andrés Sánchez Henao, Natalia García-Álvarez, Inmaculada Rosario Medina, Freddy Silva Sergent, Soraya Déniz, and et al. 2021. "Screening of New Potential Probiotics Strains against Photobacterium damselae Subsp. piscicida for Marine Aquaculture" Animals 11, no. 7: 2029. https://doi.org/10.3390/ani11072029