
Thermoregulatory Responses and Performance of Dairy Calves Fed Different Amounts of Colostrum

 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, and Treatments
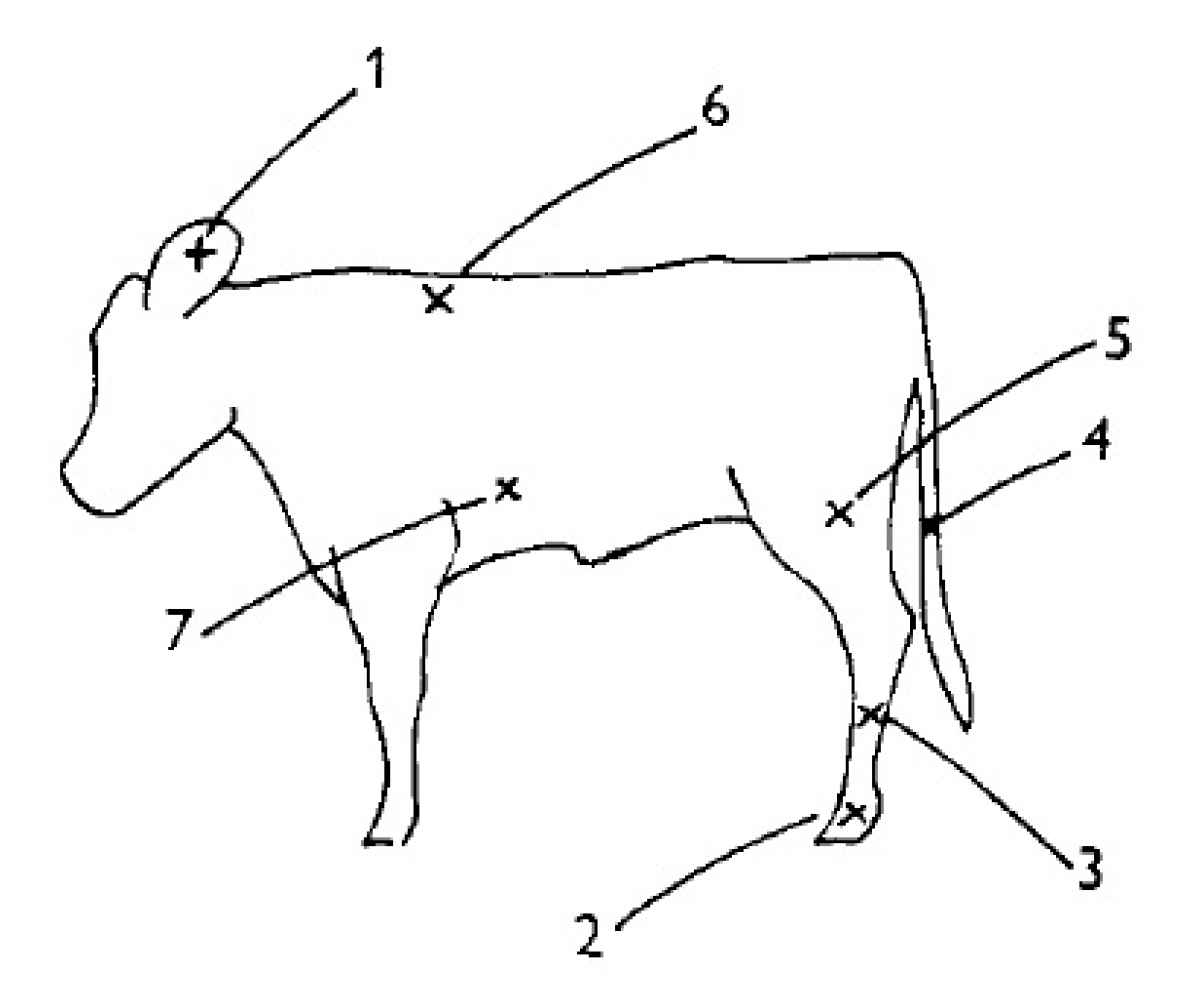
2.2. Cold Challenge
2.3. Calf Performance
2.4. Blood Sampling and Analysis of Metabolites and Hormones
2.5. Statistical Analysis
3. Results
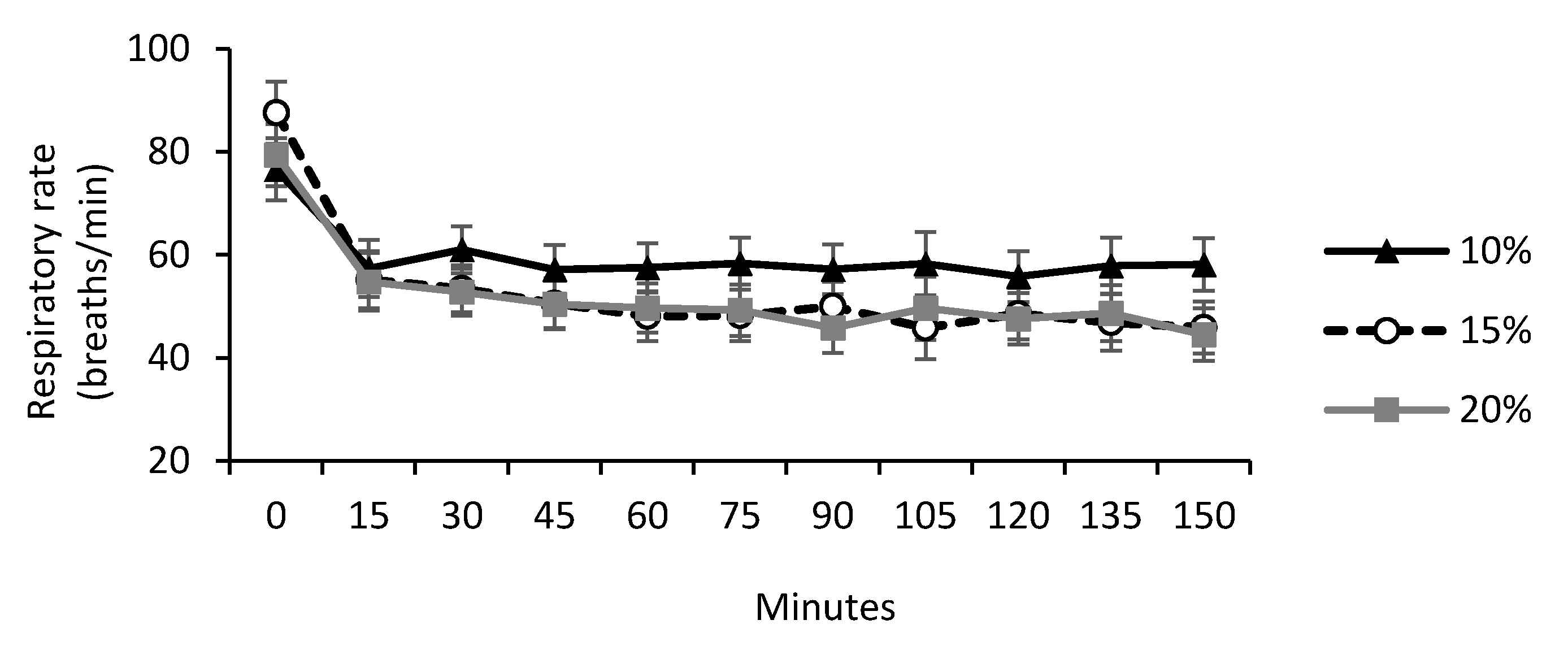
3.1. Cold Challenge
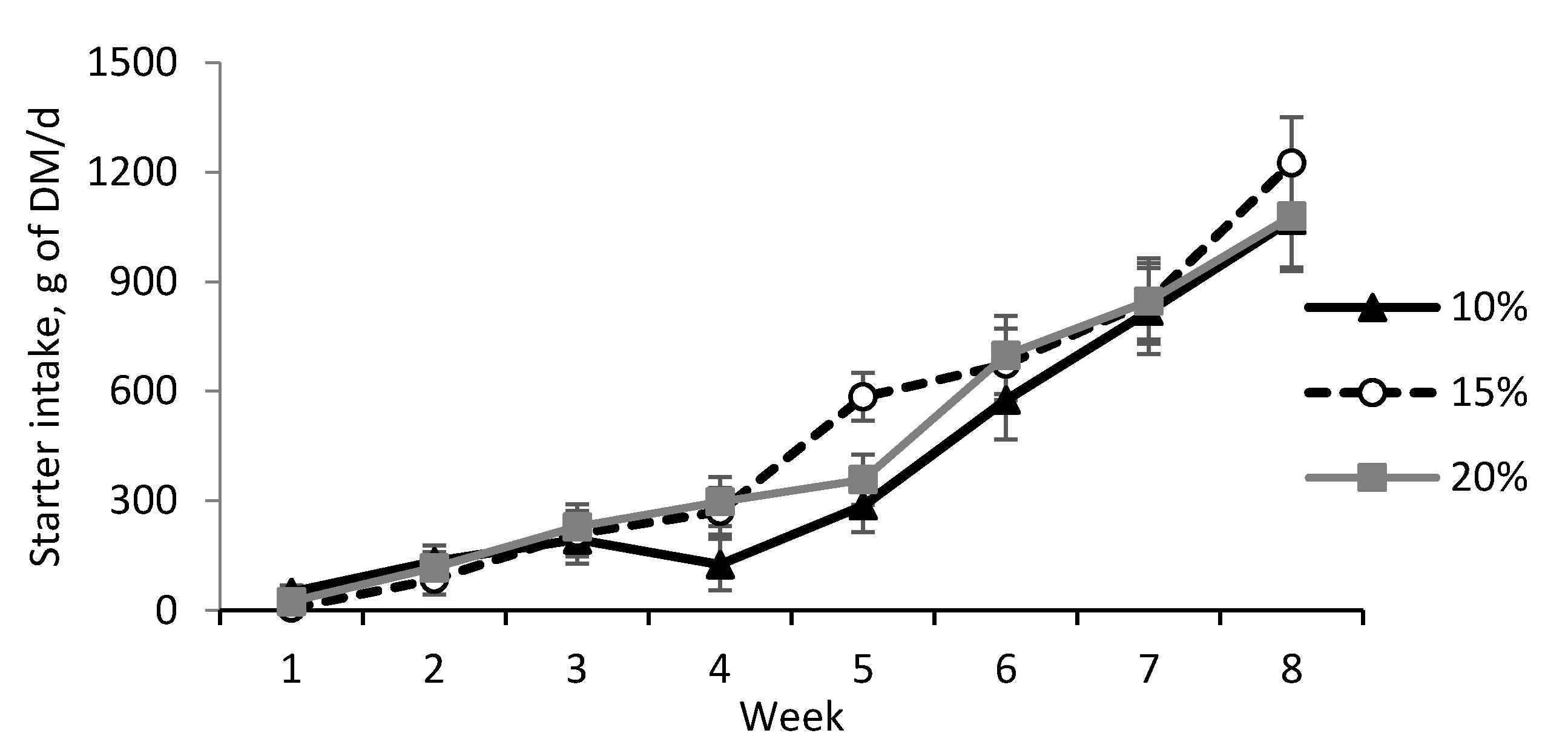
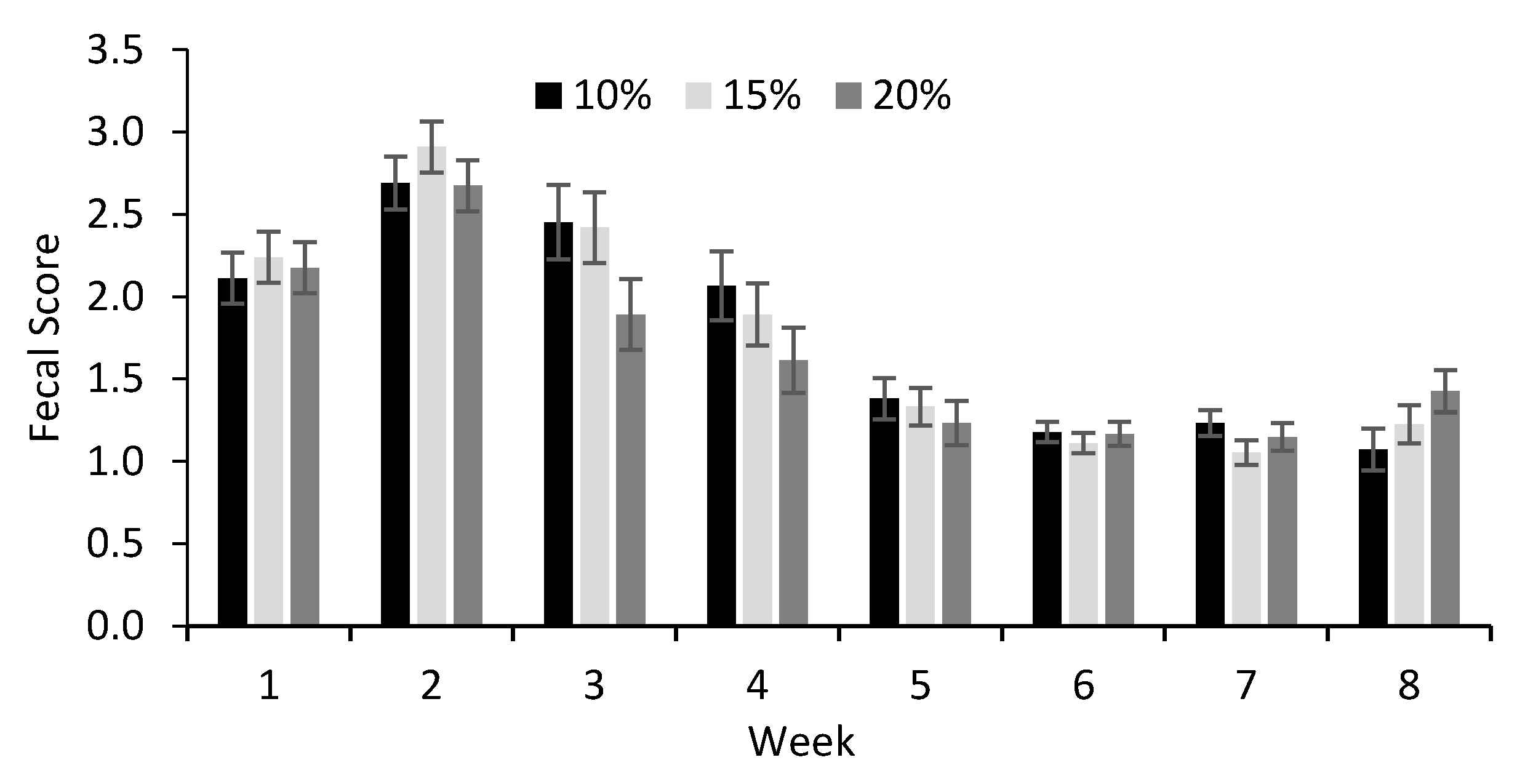
3.2. Performance, Metabolism, and Health
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Godden, S. Colostrum management for dairy calves. Vet. Clin. N. Am. Food Anim. Pract. 2008, 24, 19–39. [Google Scholar] [CrossRef]
- Osaka, I.; Matsui, Y.; Terada, F. Effect of the mass of immunoglobulin (Ig)G intake and age at first colostrum feeding on serum IgG concentration in Holstein calves. J. Dairy Sci. 2014, 97, 6608–6612. [Google Scholar] [CrossRef] [PubMed]
- Hammon, H.M.; Schiessler, G.; Nussbaum, A.; Blum, J.W. Feed intake patterns, growth performance, and metabolic and endocrine traits in calves fed unlimited amounts of colostrum and milk by automate, starting in the neonatal period. J. Dairy Sci. 2002, 85, 3352–3362. [Google Scholar] [CrossRef]
- Lombard, J.; Urie, N.J.; Garry, F.; Godden, S.; Quigley, J.D.; Earleywine, T.; McGuirk, S.; Moore, D.; Branan, M.; Chamorro, M.; et al. Consensus recommendations on calf- and herd-level passive immunity in dairy calves in the United States. J. Dairy Sci. 2020, 103, 7611–7624. [Google Scholar] [CrossRef] [PubMed]
- Faber, S.N.; Faber, N.E.; Mccauley, T.C.; Ax, R.L. Case Study: Effects of colostrum ingestion on lactational performance. Prof. Anim. Sci. 2005, 21, 420–425. [Google Scholar] [CrossRef]
- Hammon, H.M.; Steinhoff-Wagner, J.; Schönhusen, U.; Metges, C.C.; Blum, J.W. Energy metabolism in the newborn farm animal with emphasis on the calf: Endocrine changes and responses to milk-born and systemic hormones. Domest. Anim. Endocrinol. 2012, 43, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.L.; Drackley, J.K. The Development, Nutrition, and Management of the Young Calf; Iowa State University Press: Ames, IA, USA, 1998; ISBN 9780813829807. [Google Scholar]
- Bellows, R.A. Factors affecting calf survival. In Proceedings of the 15th Range Beef Cow Symposium, Rapid City, SD, USA, 9 December 1997; Volume 152, pp. 141–150. [Google Scholar]
- Diesch, T.; Mellor, D.; Stafford, K.; Ward, R. The physiological and physical status of single calves at birth in a dairy herd in New Zealand. N. Z. Vet. J. 2004, 52, 250–255. [Google Scholar] [CrossRef]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Silva, F.L.M.; Bittar, C.M.M. Thermogenesis and some rearing strategies of dairy calves at low temperature—A review. J. Appl. Anim. Res. 2019, 47, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Harper, M.-E.; Antoniou, A.; Bevilacqua, L.; Bezaire, V.; Monemdjou, S. Cellular energy expenditure and the importance of uncoupling1. J. Anim. Sci. 2002, 80, E90–E97. [Google Scholar] [CrossRef]
- Herpin, P.; Louveau, I.; Damon, M.; Le Dividich, J. Chapter 14 Environmental and hormonal regulation of energy metabolism in early development of the pig. In Biology of Growing Animals; Elsevier: Amsterdam, The Netherlands, 2005; Volume 3, pp. 351–374. [Google Scholar]
- Vermorel, M.; Dardillat, C.; Vernet, J.; Demigne, C. Energy metabolism and thermoregulation in the newborn calf. Ann. Rech. Vet. 1983, 14, 382–389. [Google Scholar]
- Himms-Hagen, J. Brown adipose tissue thermogenesis: Interdisciplinary studies. FASEB J. 1990, 4, 2890–2898. [Google Scholar] [CrossRef] [PubMed]
- Danijela, K. Endocrine and metabolic adaptations of calves to extra-uterine life. Acta Vet. Brno. 2015, 65, 297–318. [Google Scholar] [CrossRef]
- Bartier, A.L.; Windeyer, M.C.; Doepel, L. Evaluation of on-farm tools for colostrum quality measurement. J. Dairy Sci. 2015, 98, 1878–1884. [Google Scholar] [CrossRef]
- Dos Santos, G.; da Silva, J.T.; da Rocha Santos, F.H.; Bittar, C.M.M. Nutritional and microbiological quality of bovine colostrum samples in Brazil. Rev. Bras. Zootec. 2017, 46, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Jimenez, E.; Blaxter, K.L. The metabolism and thermal regulation of calves in the first month of life. Br. J. Nutr. 1962, 16, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Bellows, R.A.A.; Lammoglia, M.A.A. Effects of severity of dystocia on cold tolerance and serum concentrations of glucose and cortisol in neonatal beef calves. Theriogenology 2000, 53, 803–813. [Google Scholar] [CrossRef]
- Jolles, S.; Borrell, R.; Zouwail, S.; Heaps, A.; Sharp, H.; Moody, M.; Selwood, C.; Williams, P.; Phillips, C.; Hood, K.; et al. Calculated globulin (CG) as a screening test for antibody deficiency. Clin. Exp. Immunol. 2014, 177, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Pozos, R.S.; Danzl, D.F. Human Physiological responses to cold stress and hypothermia. In Medical Aspects of Harsh Environments; Pandolf, K.B., Burr, R.E., Eds.; Walter Reed Army Medical Center Borden Institute: Washington, DC, USA, 2001; Volume 1, pp. 351–382. [Google Scholar]
- Rauprich, A.B.E.; Hammon, H.M.; Blum, J.W. Influence of feeding different amounts of first colostrum on metabolic, endocrine, and health status and on growth performance in neonatal calves. J. Anim. Sci. 2000, 78, 896. [Google Scholar] [CrossRef] [Green Version]
- Himms-Hagen, J. Brown adipose tissue metabolism and thermogenesis. Annu. Rev. Nutr. 1985, 5, 69–94. [Google Scholar] [CrossRef]
- Vermorel, M.; Vernet, J.; Saido; Dardillat, C.; Demigne, C. Energy metabolism and thermoregulation in the newborn calf; variations during the first day of life and differences between breeds. Can. J. Anim. Sci. 1989, 69, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Hillman, N.H.; Kallapur, S.G.; Jobe, A.H. Physiology of transition from intrauterine to extrauterine Life. Clin. Perinatol. 2012, 39, 769–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, K.C.; Rajkovich, N.B.; White-Newsome, J.L.; Larsen, L.; O’Neill, M.S. Preventing cold-related morbidity and mortality in a changing climate. Maturitas 2011, 69, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Lammoglia, M.A.; Bellows, R.A.; Grings, E.E.; Bergman, J.W.; Short, R.E.; MacNeil, M.D. Effects of feeding beef females supplemental fat during gestation on cold tolerance in newborn calves. J. Anim. Sci. 1999, 77, 824. [Google Scholar] [CrossRef] [Green Version]
- Lammoglia, M.A.; Bellows, R.A.; Grings, E.E.; Bergman, J.W. Effects of prepartum supplementary fat and muscle hypertrophy genotype on cold tolerance in newborn calves. J. Anim. Sci. 1999, 77, 2227. [Google Scholar] [CrossRef]
- Klingenspor, M.; Fromme, T.; Bast, A.; Bolze, F.; Li, Y.; Maurer, S.; Schweizer, S.; Willershäuser, M.; Fromme, T. Brown adipose tissue. In Adipose Tissue Biology; Springer Press: New York, NY, USA, 2017; pp. 91–147. [Google Scholar]
- Knowles, T.G.; Warriss, P.D.; Vogel, K. Stress physiology of animals during transport. In Livestock Handling and Transport; CABI: Wallingford, UK, 2014; pp. 399–420. ISBN 9781780643212. [Google Scholar]
- Omidi-Mirzaei, H.; Khorvash, M.; Ghorbani, G.R.; Moshiri, B.; Mirzaei, M.; Pezeshki, A.; Ghaffari, M.H. Effects of the step-up/step-down and step-down milk feeding procedures on the performance, structural growth, and blood metabolites of Holstein dairy calves. J. Dairy Sci. 2015, 98, 7975–7981. [Google Scholar] [CrossRef]
- Bassett, J.M.; Alexander, G. Insulin, growth hormone and corticosteroids in neonatal lambs. Neonatology 1971, 17, 112–125. [Google Scholar] [CrossRef]
- Gruber, S.L.; Tatum, J.D.; Engle, T.E.; Chapman, P.L.; Belk, K.E.; Smith, G.C. Relationships of behavioral and physiological symptoms of preslaughter stress to beef longissimus muscle tenderness1. J. Anim. Sci. 2010, 88, 1148–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.; Dahl, G.E. Invited review: Heat stress effects during late gestation on dry cows and their calves. J. Dairy Sci. 2013, 96, 4079–4093. [Google Scholar] [CrossRef] [PubMed]
- Weaver, D.M.; Tyler, J.W.; VanMetre, D.C.; Hostetler, D.E.; Barrington, G.M. Passive transfer of colostral immunoglobulins in Calves. J. Vet. Intern. Med. 2000, 14, 569. [Google Scholar] [CrossRef]
- Velibor, S.; Anna, N.-J.; Gy, H.; Horea, S.; Dragan, G.; Danijela, K. The plasma levels of triiodothyronine, thyroxine and cortisol in newborn calves. Acta Vet. Brno. 2002, 52, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Bach, A. Ruminant nutrition symposium: Optimizing performance of the offspring: Nourishing and managing the dam and postnatal calf for optimal lactation, reproduction, and immunity1,2. J. Anim. Sci. 2012, 90, 1835–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Bach, A.; Weary, D.M.; von Keyserlingk, M.A.G. Invited review: Transitioning from milk to solid feed in dairy heifers. J. Dairy Sci. 2016, 99, 885–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelgadir, I.E.O.; Morrill, J.L.; Higgins, J.J. Effect of roasted soybeans and corn on performance and ruminal and blood metabolites of dairy calves. J. Dairy Sci. 1996, 79, 465–474. [Google Scholar] [CrossRef]
- Miyazaki, T.; Okada, K.; Miyazaki, M. Short communication: Neonatal calves coagulate first-milking colostrum and produce a large curd for efficient absorption of immunoglobulins after first ingestion. J. Dairy Sci. 2017, 100, 7262–7270. [Google Scholar] [CrossRef] [Green Version]
- Conneely, M.; Berry, D.P.; Murphy, J.P.; Lorenz, I.; Doherty, M.L.; Kennedy, E. Effect of feeding colostrum at different volumes and subsequent number of transition milk feeds on the serum immunoglobulin G concentration and health status of dairy calves. J. Dairy Sci. 2014, 97, 6991–7000. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, A.P.; de Toledo, A.F.; Cezar, A.M.; Coelho, M.G.; Virginio Júnior, G.F.; Poczynek, M.; Silva, M.D.; Haines, D.M.; Campos, M.; Bittar, C.M.M. Passive transfer and neonatal health in dairy calves receiving maternal colostrum and/or a colostrum replacer. Livest. Sci. 2020, 240, 104158. [Google Scholar] [CrossRef]
- Hulbert, L.E.; Moisá, S.J. Stress, immunity, and the management of calves1. J. Dairy Sci. 2016, 99, 3199–3216. [Google Scholar] [CrossRef] [Green Version]
- Jezek, J.; Nemec, M.; Staric, J.; Klinkon, M. Age related changes and reference intervals of haematological variables in dairy calves. Bull. Vet. Inst. Puławy 2011, 55, 471–478. [Google Scholar]
- Reber, A.J.; Hippen, A.R.; Hurley, D.J. Effects of the ingestion of whole colostrum or cell-free colostrum on the capacity of leukocytes in newborn calves to stimulate or respond in one-way mixed leukocyte cultures. Am. J. Vet. Res. 2005, 66, 1854–1860. [Google Scholar] [CrossRef]
- Hammon, H.M.; Steinhoff-Wagner, J.; Flor, J.; Schönhusen, U.; Metges, C.C. Lactation biology symposium: Role of colostrum and colostrum components on glucose metabolism in neonatal calves1,2. J. Anim. Sci. 2013, 91, 685–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item 1 | Treatments 2 | SEM | p-Value 3 | ||||
|---|---|---|---|---|---|---|---|
| 10% | 15% | 20% | Treat | Time | Treat * Time | ||
| Heart rate *, beats/min | 85.1 | 84.2 | 82.1 | 4.66 | 0.88 | 0.25 | 0.17 |
| Respiratory rate *, breaths/min | 59.6 | 52.8 | 52.1 | 4.58 | 0.45 | <0.001 | 0.09 |
| Shivering score * | 2.2 a | 2.0 ab | 1.7 b | 0.19 | 0.10 | <0.001 | 0.77 |
| Rectal temperature *, °C | 37.7 b | 38.1 a | 38.0 a | 0.08 | 0.06 | <0.001 | 0.32 |
| Skin Surface Temperature *, °C | |||||||
| Ear | 19.4 | 19.9 | 20.8 | 1.14 | 0.66 | <0.001 | 0.72 |
| Prescapular | 30.1 b | 30.7 ab | 31.9 a | 0.6 | 0.09 | 0.11 | 0.54 |
| Thorax wall | 32.5 | 33.0 | 33.4 | 0.45 | 0.36 | <0.001 | 0.83 |
| Thigh | 30.5 | 30.0 | 30.6 | 1.1 | 0.87 | <0.001 | 0.97 |
| Shin | 26.0 | 27.8 | 27.8 | 0.93 | 0.23 | <0.001 | 0.62 |
| Foot | 23.2 | 26.8 | 24.6 | 1.14 | 0.11 | <0.001 | 0.11 |
| Tail | 17.4 | 22.5 | 20.9 | 1.18 | 0.18 | <0.001 | 0.34 |
| Selected Blood Metabolites ** | |||||||
| Total protein, g/dL | 6.4 b | 6.7 ab | 7.1 a | 0.23 | 0.09 | <0.001 | 0.86 |
| Albumin, g/dL | 2.5 b | 2.4 ab | 2.4 a | 0.05 | 0.08 | 0.01 | 0.48 |
| Globulin, g/dL | 3.86 b | 4.36 ab | 4.73 a | 0.22 | 0.03 | <0.001 | 0.92 |
| Glucose, mg/dL | 125.6 | 127.1 | 141.3 | 8.51 | 0.34 | <0.001 | 0.90 |
| Lactate, mg/dL | 46.7 a | 44.9 a | 34.6 b | 3.85 | 0.07 | 0.03 | 0.17 |
| NEFA, mmol/L | 0.5 | 0.6 | 0.5 | 0.05 | 0.20 | <0.001 | 0.73 |
| Alk. Phosphatase, UI | 174.1 | 187.1 | 153.7 | 23.7 | 0.33 | <0.001 | 1.00 |
| Blood hormones | |||||||
| Insulin **, μUI | 3.8 | 4.7 | 4.0 | 0.68 | 0.61 | 0.01 | 0.71 |
| Cortisol ¥, µg/dL | 6.8 | 7.1 | 8.1 | 0.85 | 0.32 | 0.12 | 0.75 |
| T3 §, ng/dL | 4.2 | 3.5 | 4.3 | 0.55 | 0.56 | 0.81 | 0.35 |
| T4 §, ng/dL | 1.3 | 1.2 | 1.1 | 0.19 | 0.53 | 0.30 | 0.36 |
| Item | Treatments 1 | SEM | p-Value 2 | ||||
|---|---|---|---|---|---|---|---|
| 10% | 15% | 20% | Treat | Age 3 | Treat *Age | ||
| Starter intake, g DM/d | 407.1 | 488.2 | 456.5 | 50.05 | 0.52 | <0.001 | 0.08 |
| Total intake, g DM/d | 1098.0 | 1178.4 | 1156.2 | 77.66 | 0.65 | <0.001 | 0.14 |
| Body weight, kg | 47.1 | 50.9 | 50.2 | 4.88 | 0.22 | <0.001 | 0.73 |
| Average gain, g/d | 481.4 | 544.4 | 552.8 | 55.24 | 0.48 | <0.001 | 0.42 |
| Feed efficiency | 0.4 | 0.4 | 0.5 | 0.03 | 0.20 | <0.001 | 0.42 |
| Heart girth, cm | 84.1 b | 86.9 a | 86.6 a | 2.65 | 0.05 | <0.001 | 0.10 |
| Hip width, cm | 22.9 | 23.6 | 23.3 | 0.69 | 0.18 | <0.001 | 0.34 |
| Withers height, cm | 82.7 | 84.2 | 82.6 | 1.88 | 0.32 | <0.001 | 0.94 |
| Item 1 | Treatments 2 | SEM | p-Value 3 | ||||
|---|---|---|---|---|---|---|---|
| 10% | 15% | 20% | Treat | Age 4 | Treat *Age | ||
| Total protein, g/dL | 5.4 | 5.5 | 5.6 | 0.09 | 0.32 | <0.001 | 0.17 |
| Albumin, g/dL | 2.8 | 2.8 | 2.9 | 2.83 | 0.30 | <0.001 | 0.52 |
| Glucose, mg/dL | 98.6 | 107.9 | 102.7 | 6.00 | 0.54 | <0.001 | 0.95 |
| Lactate, mg/dL | 13.5 | 14.2 | 14.3 | 0.89 | 0.80 | <0.001 | 0.30 |
| BHB, mmol/L | 0.07 b | 0.09 a | 0.09 a | 0.005 | 0.09 | <0.001 | 0.50 |
| NEFA, mmol/L | 0.21 a | 0.16 b | 0.19 ab | 0.19 | 0.08 | <0.001 | 0.49 |
| Item | Treatments 1 | SEM | p-Value 2 | ||||
|---|---|---|---|---|---|---|---|
| 10% | 15% | 20% | Treat | Age | Treat * Age | ||
| Fecal score | |||||||
| 24 h after birth | 1.0 b | 1.5 a | 1.5 a | 0.18 | 0.07 | - | - |
| Preweaning | 1.8 | 1.8 | 1.7 | 0.07 | 0.42 | <0.001 | 0.46 |
| Diarrhea days | 10.3 | 12.4 | 9.3 | 2.08 | 0.28 | - | - |
| Rectal temperature, °C | 38.3 | 38.3 | 38.3 | 0.06 | 0.87 | <0.001 | 0.27 |
| Hematocrit, % | 20.1 | 20.0 | 20.5 | 0.95 | 0.74 | 0.03 | 0.96 |
| Erythrocytes, 106/µL | 7.1 | 7.2 | 6.9 | 0.27 | 0.65 | <0.001 | 0.23 |
| MCV 3, µm | 29.8 | 28.4 | 30.9 | 0.96 | 0.17 | <0.001 | 0.51 |
| Leukocytes, 103/µL | 6.3 b | 6.3 b | 7.4 a | 0.37 | 0.02 | 0.02 | 0.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, F.L.M.; Miqueo, E.; Silva, M.D.d.; Torrezan, T.M.; Rocha, N.B.; Salles, M.S.V.; Bittar, C.M.M. Thermoregulatory Responses and Performance of Dairy Calves Fed Different Amounts of Colostrum. Animals 2021, 11, 703. https://doi.org/10.3390/ani11030703
Silva FLM, Miqueo E, Silva MDd, Torrezan TM, Rocha NB, Salles MSV, Bittar CMM. Thermoregulatory Responses and Performance of Dairy Calves Fed Different Amounts of Colostrum. Animals. 2021; 11(3):703. https://doi.org/10.3390/ani11030703
Chicago/Turabian StyleSilva, Fernanda Lavinia Moura, Evangelina Miqueo, Marcos Donizete da Silva, Thaís Manzoni Torrezan, Nathalia Brito Rocha, Márcia Saladini Vieira Salles, and Carla Maris Machado Bittar. 2021. "Thermoregulatory Responses and Performance of Dairy Calves Fed Different Amounts of Colostrum" Animals 11, no. 3: 703. https://doi.org/10.3390/ani11030703