
Improving Survival of Juvenile Scalloped Spiny Lobster (Panulirus homarus) and Crucifix Crab (Charybdis feriatus) Using Shelter and Live Prey

 ,
, 
 ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Palinurid Lobster Experiment
2.3. Crucifix Crab Experiment
2.4. Statistical Analysis
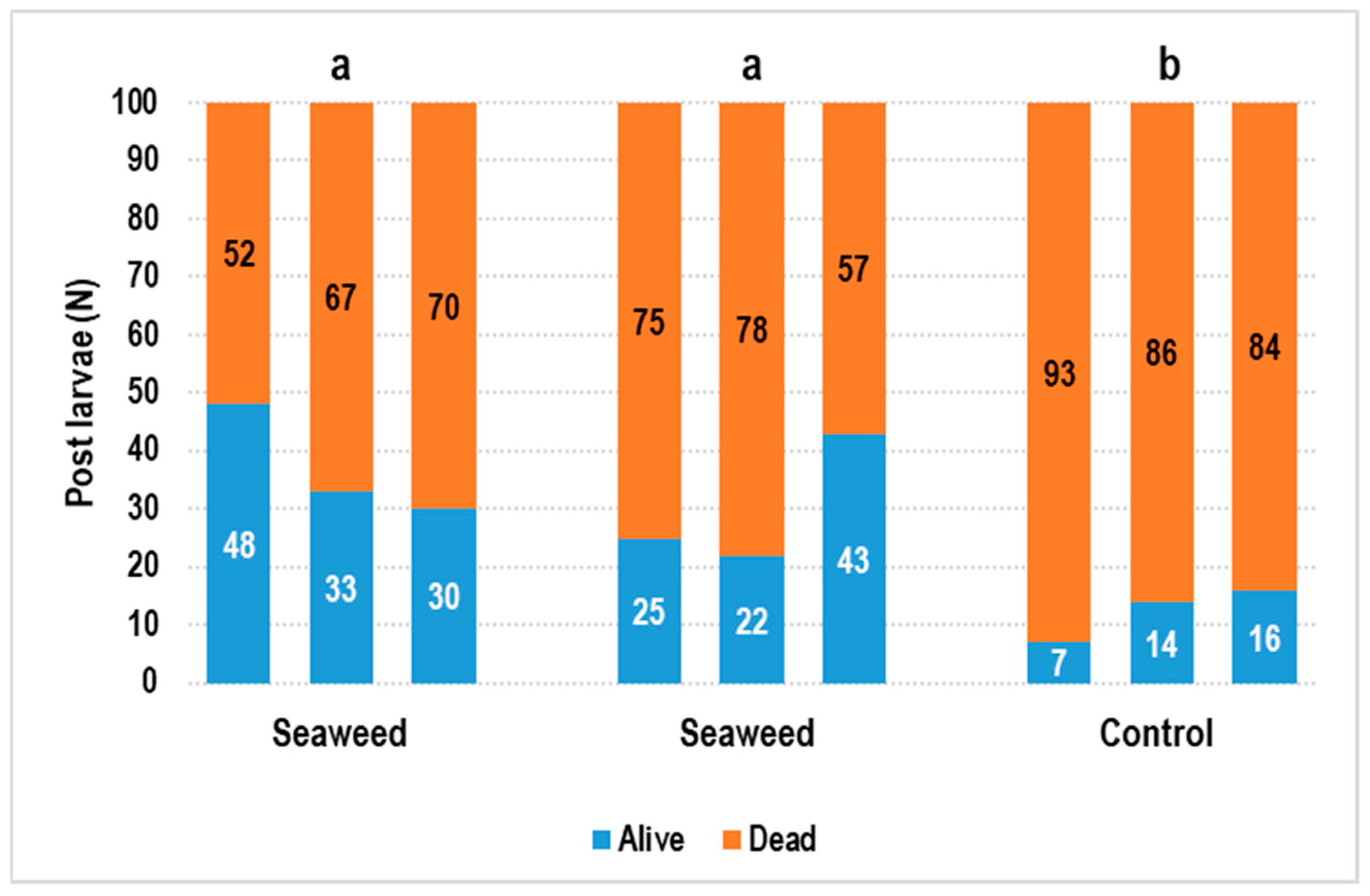
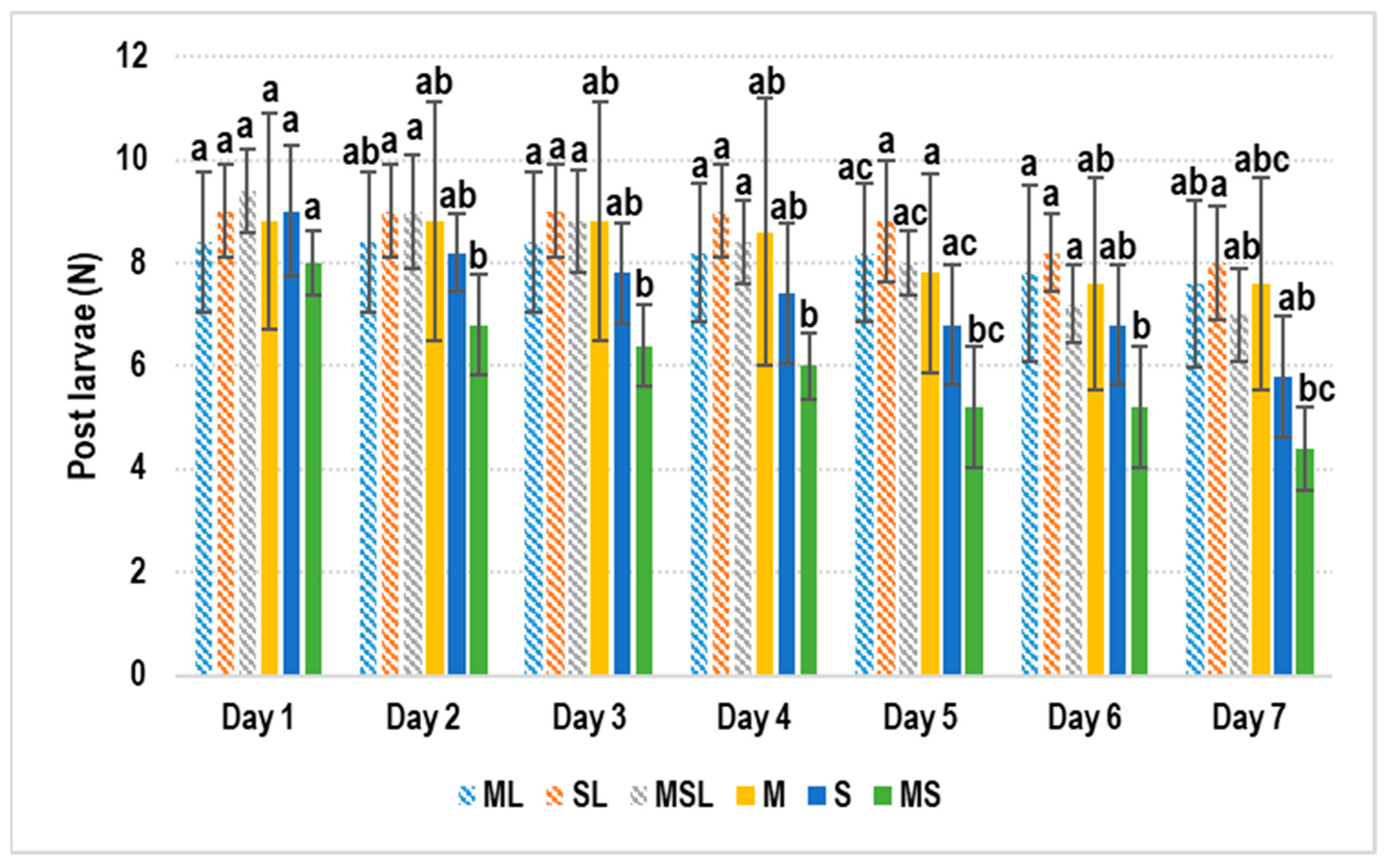
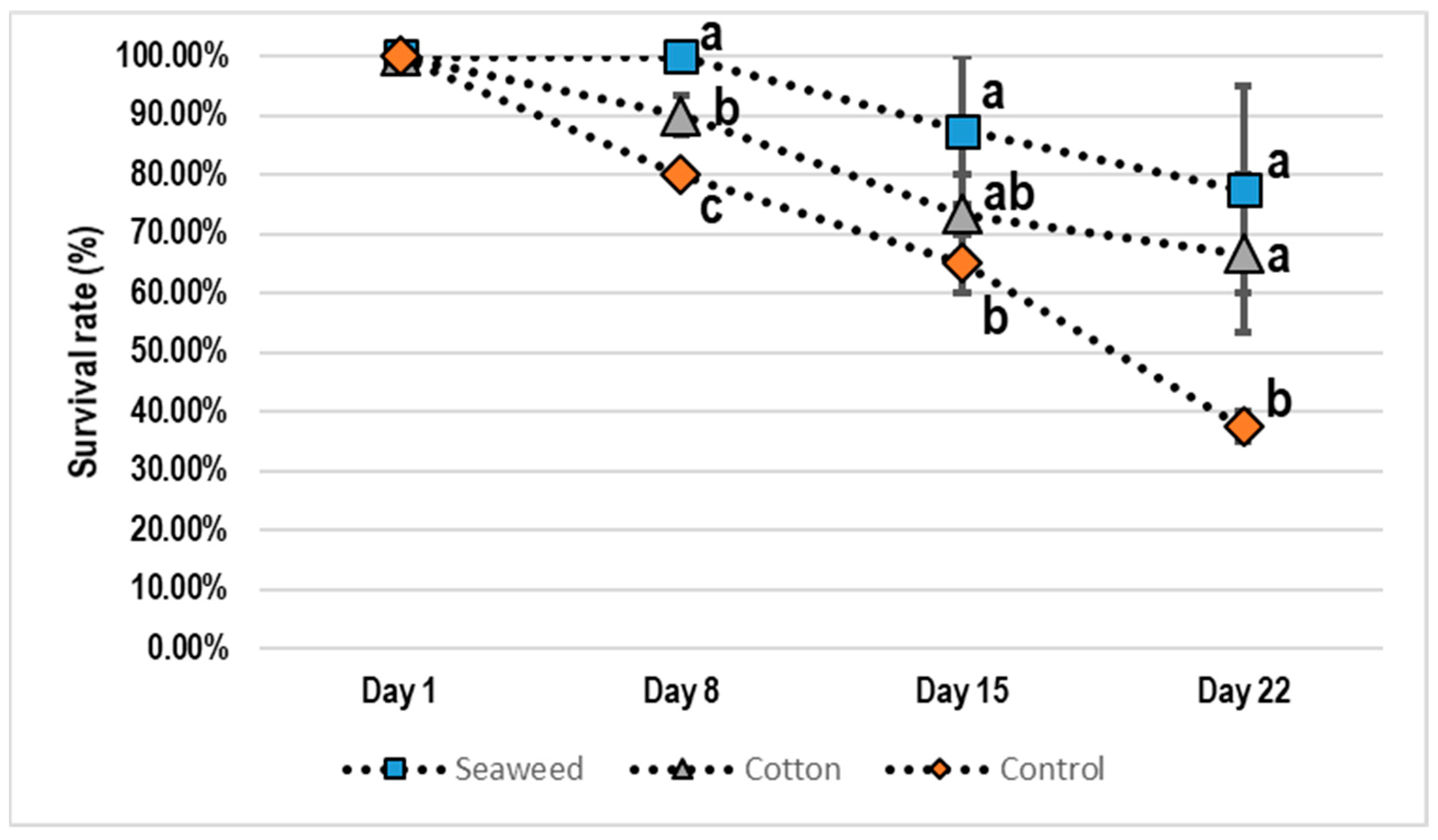
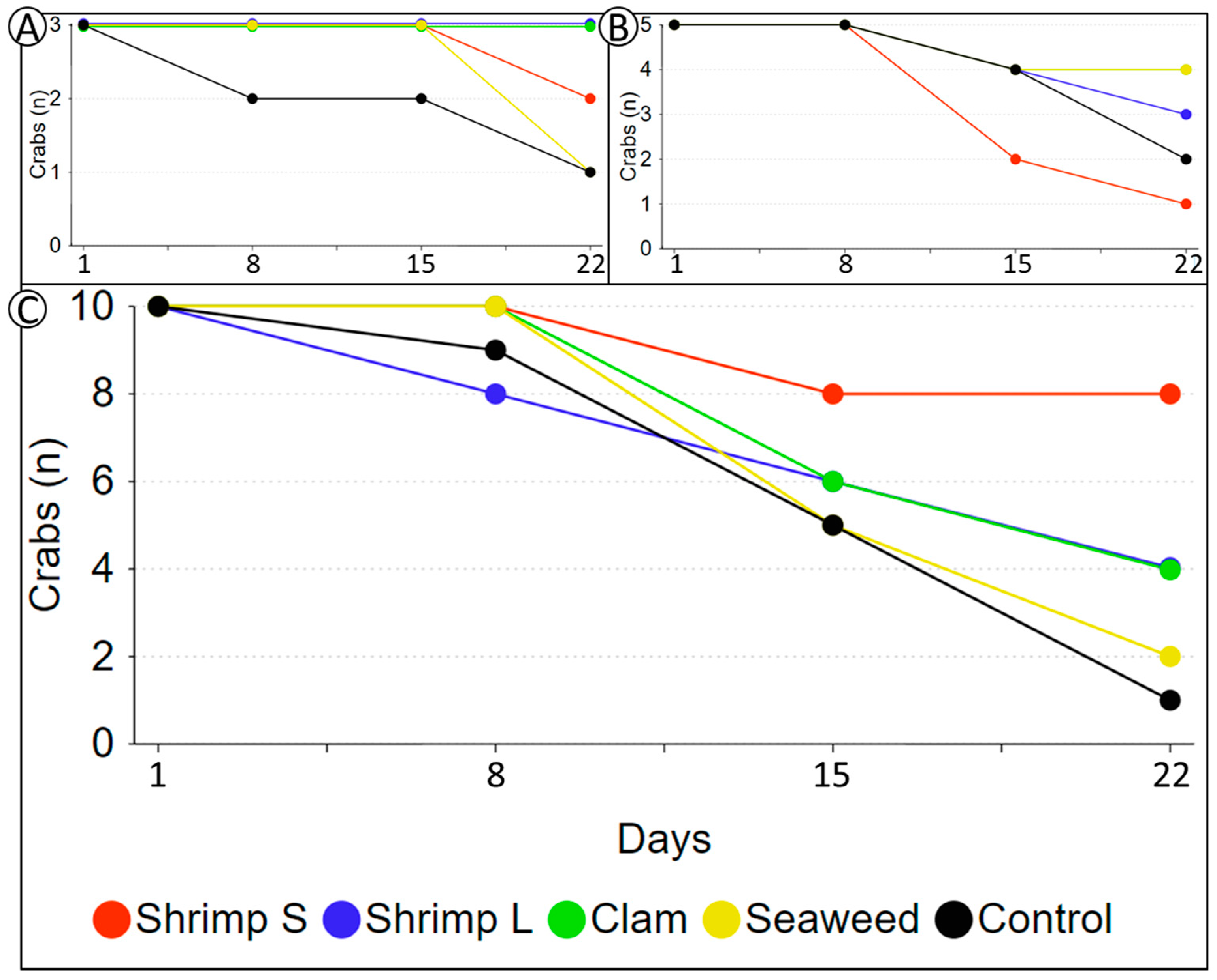
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- FAO. GLOBEFISH Highlights October 2019 ISSUE, with Jan.–Jun. 2019 Statistics–A quarterly update on world seafood markets. In Globefish Highlights; Lem, A., Ryder, J., Souza, M.C.D., Pierce, R.M., Eds.; FAO: Rome, Italy, 2019; Volume 4, p. 62. [Google Scholar]
- Vijayakumaran, M.; Radhakrishnan, E. Effects of food density on feeding and moulting of phyllosoma larvae of the spiny lobster Panulirus homarus (Linnaeus). Proc. Symp. Coast. Aquac. 1986, 4, 1281–1285. [Google Scholar]
- Matsuda, H.; Yamakawa, T. The complete development and morphological changes of larval Panulirus longipes (Decapoda, Palinuridae) under laboratory conditions. Fish Sci. 2000, 66, 278–293. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, H.; Takenouchi, T. New tank design for larval culture of Japanese spiny lobster, Panulirus japonicus. N. Z. J. Mar. Freshw. Res. 2005, 39, 279–285. [Google Scholar] [CrossRef]
- Sekiguchi, H.; Inoue, N. Larval recruitment and fisheries of the spiny lobster Panulirus japonicus coupling with the Kuroshio subgyre circulation in the western North Pacific: A review. J. Mar. Biol. Ass. India 2010, 52, 195–207. [Google Scholar]
- Smith, G.; Kenway, M.; Hall, M. Starvation and recovery ability of phyllosoma of the tropical spiny lobsters Panulirus ornatus and P. homarus in captivity. J. Mar. Biol. Ass. India 2010, 52, 249–256. [Google Scholar]
- Goldstein, J.S.; Matsuda, H.; Matthews, T.R.; Abe, F.; Yamakawa, T. Development in culture of larval spotted spiny lobster Panulirus guttatus (Latreille, 1804) (Decapoda: Achelata: Palinuridae). J. Crust. Biol. 2019, 39, 574–581. [Google Scholar] [CrossRef]
- García-Echauri, L.L.; Liggins, G.; Cetina-Heredia, P.; Roughan, M.; Coleman, M.A.; Jeffs, A. Future ocean temperature impacting the survival prospects of post-larval spiny lobsters. Mar. Environ. Res. 2020, 156, 104918. [Google Scholar] [CrossRef]
- Williams, M.; Primavera, J. Choosing tropical portunid species for culture, domestication and stock enhancement in the Indo-Pacific. Asian Fish. Sci. 2001, 14, 121–142. [Google Scholar]
- Baylon, J.; Suzuki, H. Effects of changes in salinity and temperature on survival and development of larvae and juveniles of the crucifix crab Charybdis feriatus (Crustacea:Decapoda:Portunidae). Aquaculture 2007, 269, 390–401. [Google Scholar] [CrossRef]
- Herrnkind, W.F.; Butler, M.J., IV. Factors regulating settlement and microhabitat use by spiny lobsters Panulirus argus. Mar. Ecol. Prog. Ser. 1986, 34, 23–30. [Google Scholar] [CrossRef]
- Eggleston, D.B.; Lipcius, R.N.; Miller, D.L.; Coba-Cetina, L. Shelter scaling regulates survival of juvenile Caribbean spiny lobster Panulirus argus. Mar. Ecol. Prog. Ser. 1990, 62, 79–88. [Google Scholar] [CrossRef]
- Lozano-Alvarez, E.; Briones-Fourzán, P.; Negrete-Soto, F. An evaluation of concrete block structures as shelter for juvenile Caribbean spiny lobsters, Panulirus Argus. Bull. Mar. Sci. 1994, 55, 351–362. [Google Scholar]
- Johnston, D.; Melville-Smith, R.; Hendriks, B.; Maguire, G.B.; Phillips, B. Stocking density and shelter type for the optimal growth and survival of western rock lobster Panulirus cygnus (George). Aquaculture 2006, 260, 114–127. [Google Scholar] [CrossRef] [Green Version]
- Sumbing, M.V.; Al-Azad, S.; Estim, A.; Mustafa, S. Growth performance of spiny lobster Panulirus ornatus in land-based Integrated Multi-Trophic Aquaculture (IMTA) system. Trans. Sci. Technol. 2016, 3, 143–149. [Google Scholar]
- Supriyono, E.; Prihardianto, R.W.; Nirmala, K. The stress and growth responses of spiny lobster Panulirus homarus reared in recirculation system equipped by PVC shelter. Aquacult. Aquarium Conserv. Legis. 2017, 10, 147–155. [Google Scholar]
- Baeza, J.A.; Childress, M.J.; Ambrosio, L.J. Chemical sensing of microhabitat by pueruli of the reef-dwelling Caribbean spiny lobster Panulirus argus: Testing the importance of red algae, juveniles, and their interactive effect. Bull. Mar. Sci. 2018, 94, 603–618. [Google Scholar] [CrossRef] [Green Version]
- Syafrizal, S.; Jones, C.M.; Permana, I.G.; Utomo, N.B.P. Effect of feeding frequency on survival and growth of juvenile spiny lobster Panulirus versicolor in Indonesia. Aquacult. Aquarium Conserv. Legis. 2018, 11, 1427–1434. [Google Scholar]
- Oniam, V.; Arkronrat, W.; Mohamed, N.B. Effect of feeding frequency and various shelter of blue swimming crab larvae, Portunus pelagicus (Linnaeus, 1758). Songklanakarin J. Sci. Technol. 2015, 37, 129–134. [Google Scholar]
- Liu, D.; Guo, X.; Zhong, D.; Wang, F.; Li, L.; Hu, N. Prey density and a conspecific competitor influence multiple predator effects in a crab-clam foraging system. Aquaculture 2018, 493, 264–271. [Google Scholar] [CrossRef]
- Huang, H.; Wang, Y.; Cheng, J.; Zhou, G.; Yang, C.; Song, X.; Dong, H. Optimization of mixed aquiculture for eriocheir sinensis and ecological impact in Taihu lake. Oceanol. Limnol. Sin. 2006, 37, 430. [Google Scholar]
- Gao, X.; Endo, H.; Agatsuma, Y. Seasonal changes in photosynthesis, growth, nitrogen accumulation, and salinity tolerance of Chaetomorpha crassa (Cladophorales, Chlorophyceae). J. Appl. Phycol. 2018, 30, 1905–1912. [Google Scholar] [CrossRef]
- Joll, L.M. Foregut evacuation of four foods by the western rock lobster, Panulirus cygnus, in aquaria. Mar. Freshw. Res. 1982, 33, 939–943. [Google Scholar] [CrossRef]
- Castañeda-Fernández-de-Lara, V.; Serviere-Zaragoza, E.; Hernández-Vázquez, S.; Butler, M.J. Feeding ecology of juvenile spiny lobster, Panulirus interruptus, on the Pacific coast of Baja California Sur, Mexico. N. Z. J. Mar. Freshw. Res. 2005, 39, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Mashaii, N.; Rajabipour, F.; Shakouri, A. Feeding habits of the scalloped spiny lobster, Panulirus homarus (Linnaeus, 1758)(Decapoda: Palinuridae) from the South East Coast of Iran. Turk. J. Fish. Aquat. Sci. 2011, 11, 45–54. [Google Scholar] [CrossRef]
- Ton Nu Hai, A.; Speelman, S. Economic-environmental trade-offs in marine aquaculture: The case of lobster farming in Vietnam. Aquaculture 2020, 516, 734593. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Subject | Treatment |
|---|---|---|
| Panulirus homarus | Shelter | Seaweed |
| Cotton filter | ||
| Prey | ML: Fresh clam and live shrimp | |
| SL: Fresh shrimp and live shrimp | ||
| MSL: Fresh clam, fresh shrimp, and live shrimp | ||
| M: Fresh clam | ||
| S: Fresh shrimp | ||
| MS: Fresh clam and fresh shrimp | ||
| Charybdis feriatus | Shelter | Seaweed |
| Cotton filter | ||
| Prey | Live shrimp S | |
| Live shrimp L | ||
| clam |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, C.-H.; Huang, P.-Y.; Chang, Y.-C.; Pan, Y.-J.; Azra, M.N.; Chen, L.-L.; Hsu, T.-H. Improving Survival of Juvenile Scalloped Spiny Lobster (Panulirus homarus) and Crucifix Crab (Charybdis feriatus) Using Shelter and Live Prey. Animals 2021, 11, 370. https://doi.org/10.3390/ani11020370
Ma C-H, Huang P-Y, Chang Y-C, Pan Y-J, Azra MN, Chen L-L, Hsu T-H. Improving Survival of Juvenile Scalloped Spiny Lobster (Panulirus homarus) and Crucifix Crab (Charybdis feriatus) Using Shelter and Live Prey. Animals. 2021; 11(2):370. https://doi.org/10.3390/ani11020370
Chicago/Turabian StyleMa, Chia-Huan, Po-Yu Huang, Yung-Cheng Chang, Yen-Ju Pan, Mohamad Nor Azra, Li-Li Chen, and Te-Hua Hsu. 2021. "Improving Survival of Juvenile Scalloped Spiny Lobster (Panulirus homarus) and Crucifix Crab (Charybdis feriatus) Using Shelter and Live Prey" Animals 11, no. 2: 370. https://doi.org/10.3390/ani11020370