Effect of Green Algae Chaetomorpha antennina Extract on Growth, Modulate Immunity, and Defenses against Edwardsiella tarda Infection in Labeo rohita

 ,
,  ,
, 


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Experimental Setup
2.2. Preparation of Basal Diet
2.3. Preparation of MECA
2.4. Experimental Design
2.5. Blood Collection and Sample Analysis for Immunological Parameters
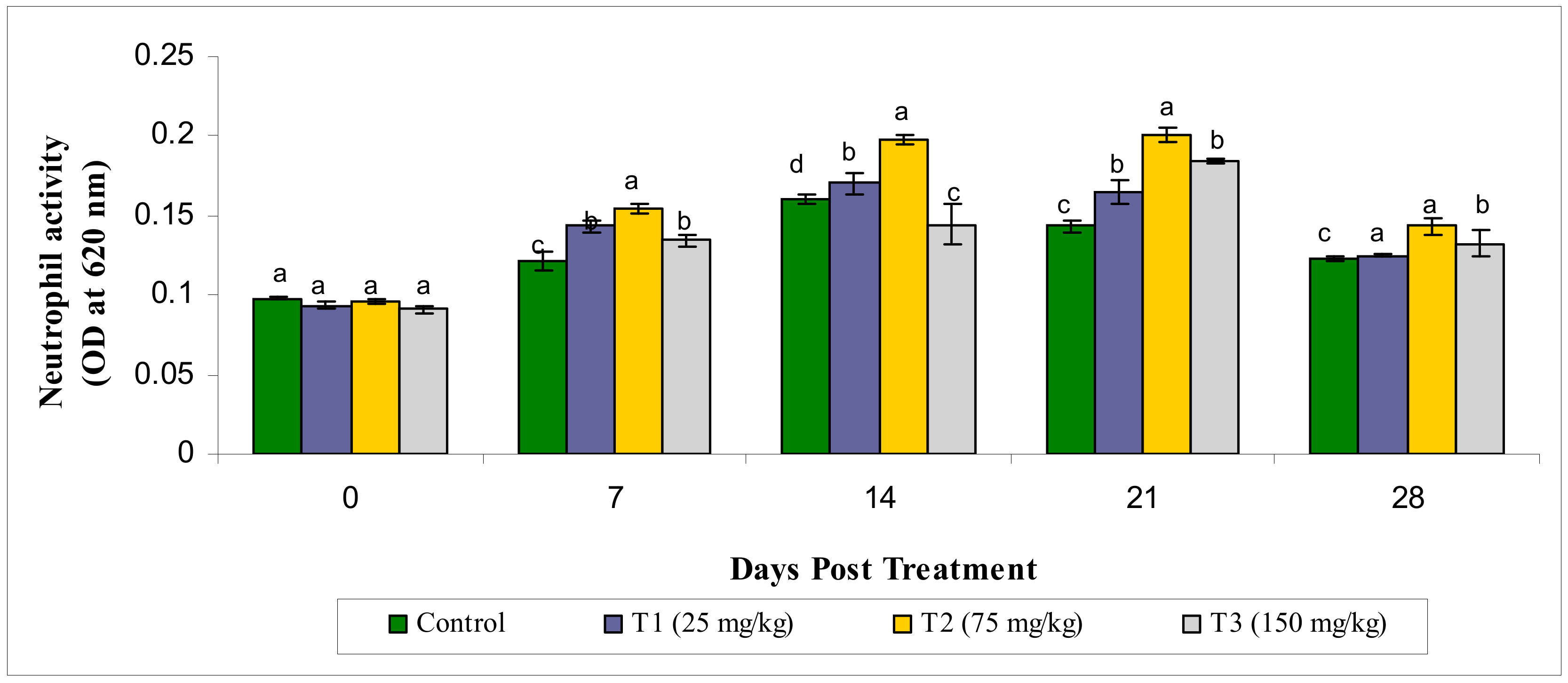
2.5.1. Neutrophil Activity
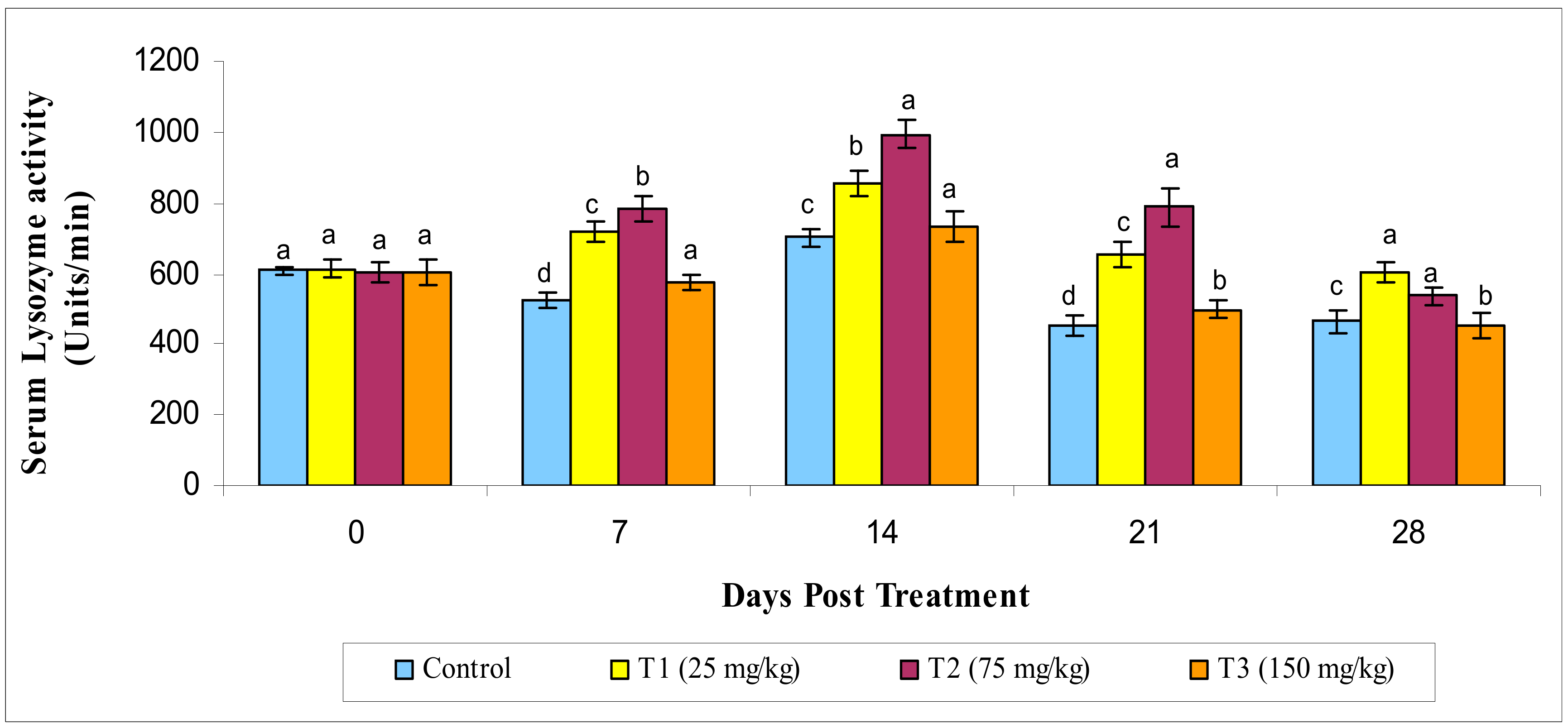
2.5.2. Serum Lysozyme Activity
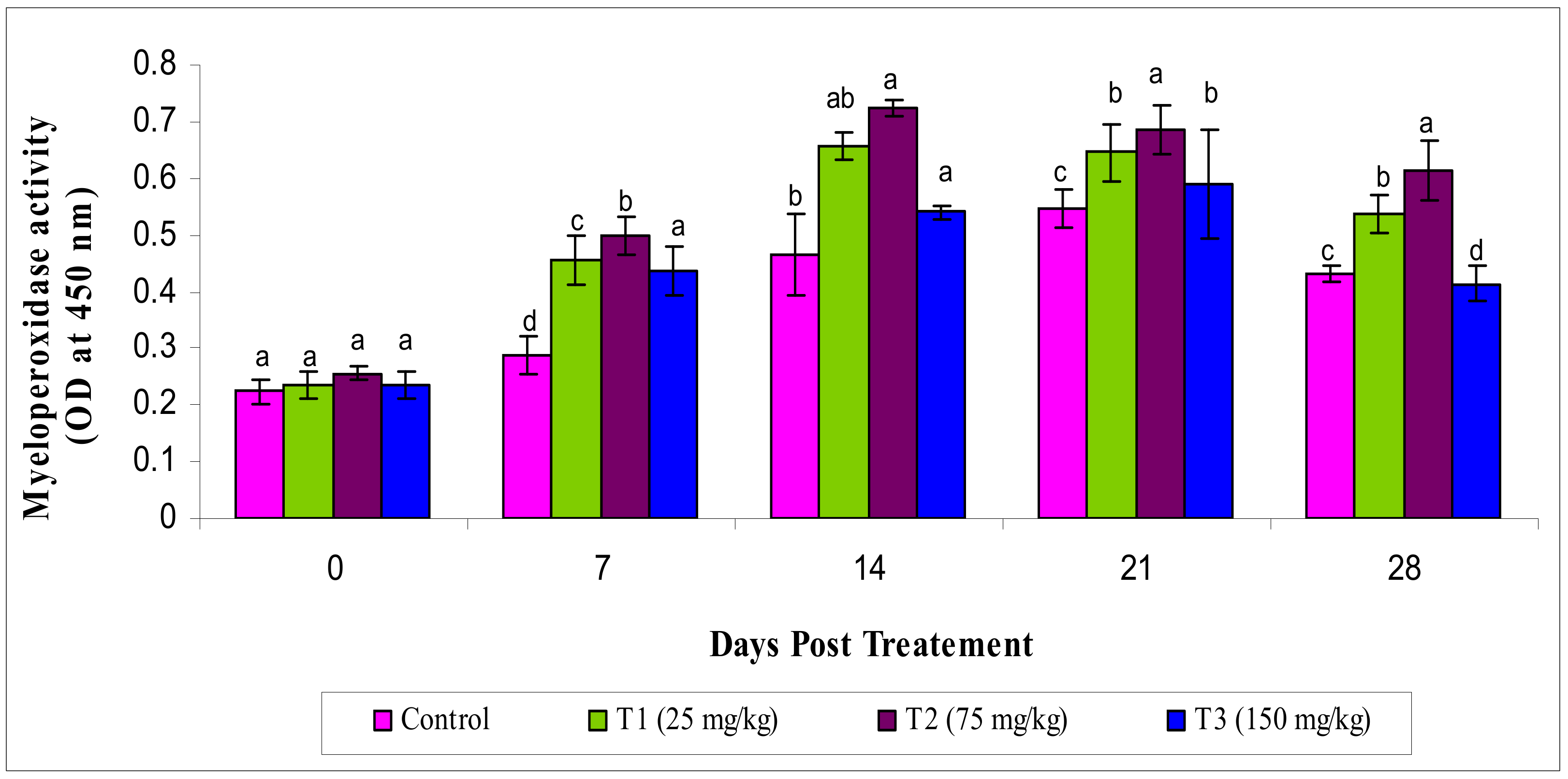
2.5.3. Myeloperoxidase (MPO) Activity
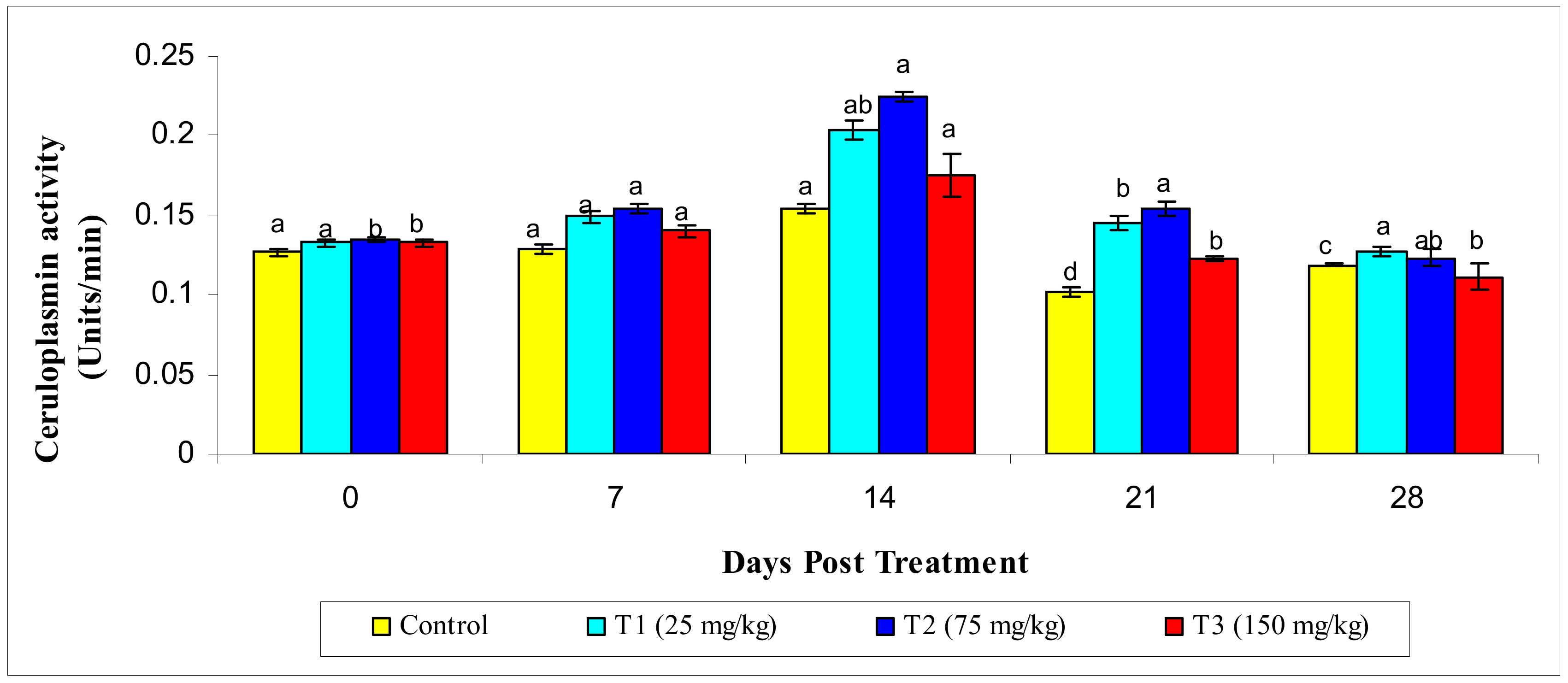
2.5.4. Ceruloplasmin Activity
2.5.5. Serum Antiprotease Activity
2.6. Preparation of Viable Leukocytes from Peripheral Blood
2.7. Reactive Oxygen Species Production
2.8. Reactive Nitrogen Species Production
2.9. Disease Resistance
2.10. Statistical Analysis
3. Results
3.1. Growth Response
3.2. Immunological Response
3.2.1. Neutrophil Activity
3.2.2. Lysozyme Activity
3.2.3. Myeloperoxidase Activity
3.2.4. Ceruloplasmin Activity
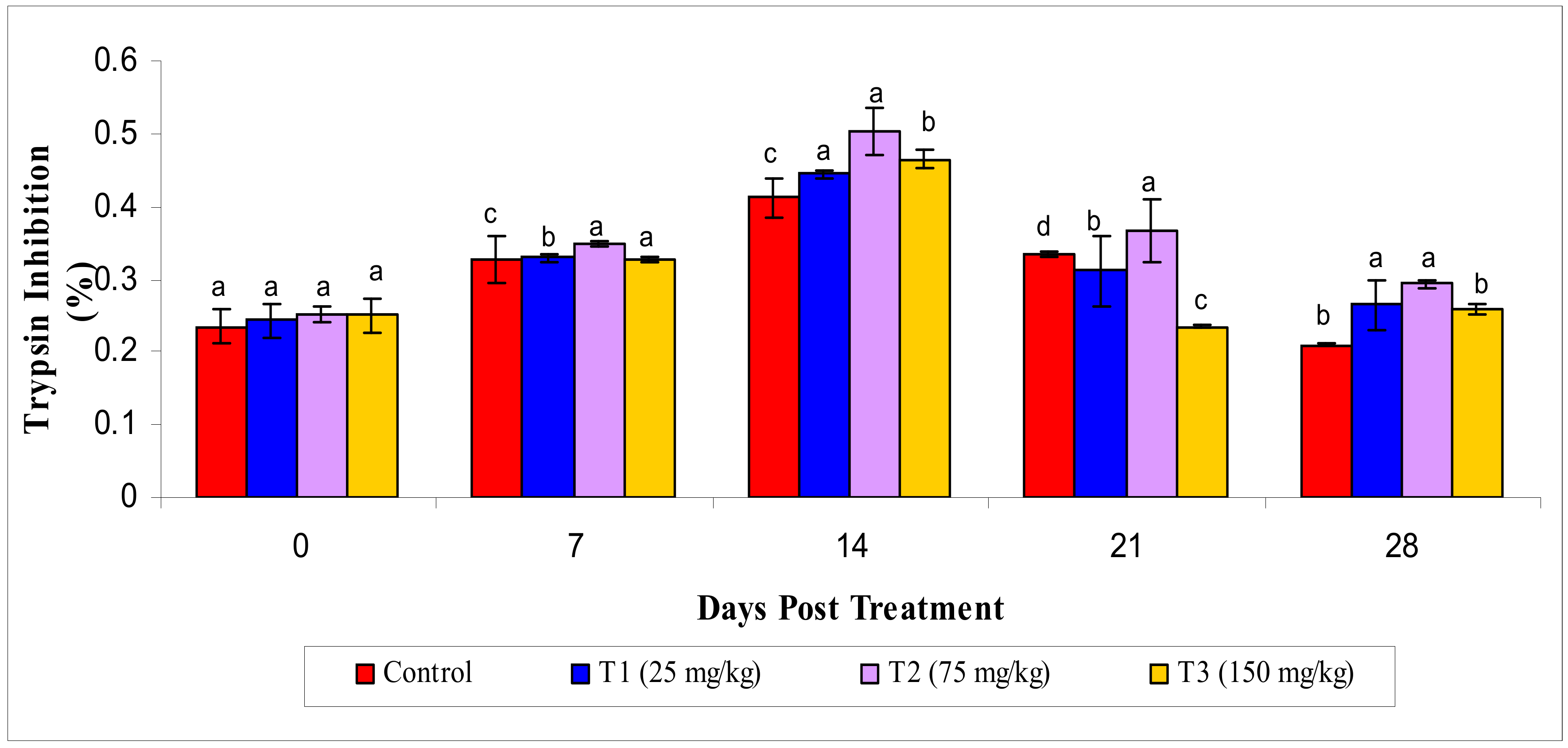
3.2.5. Antiprotease Activity
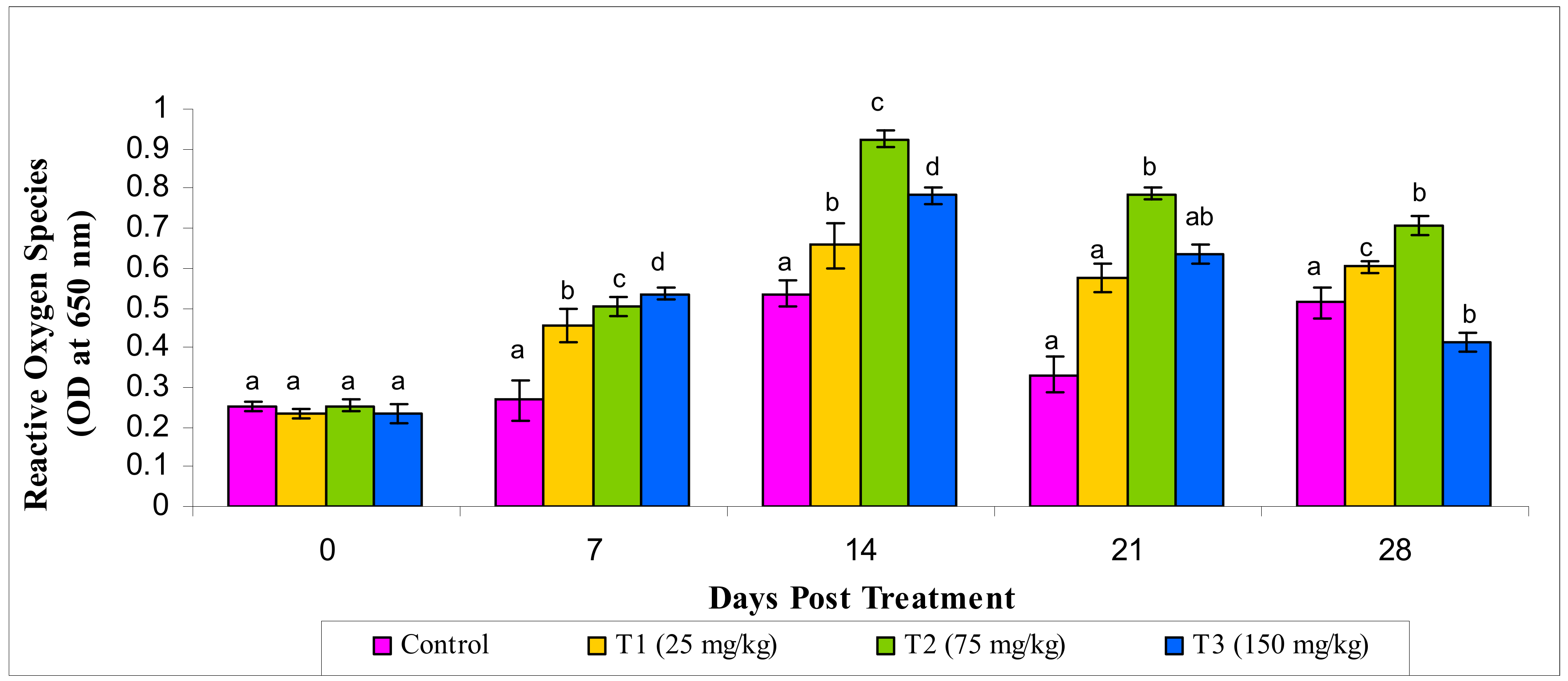
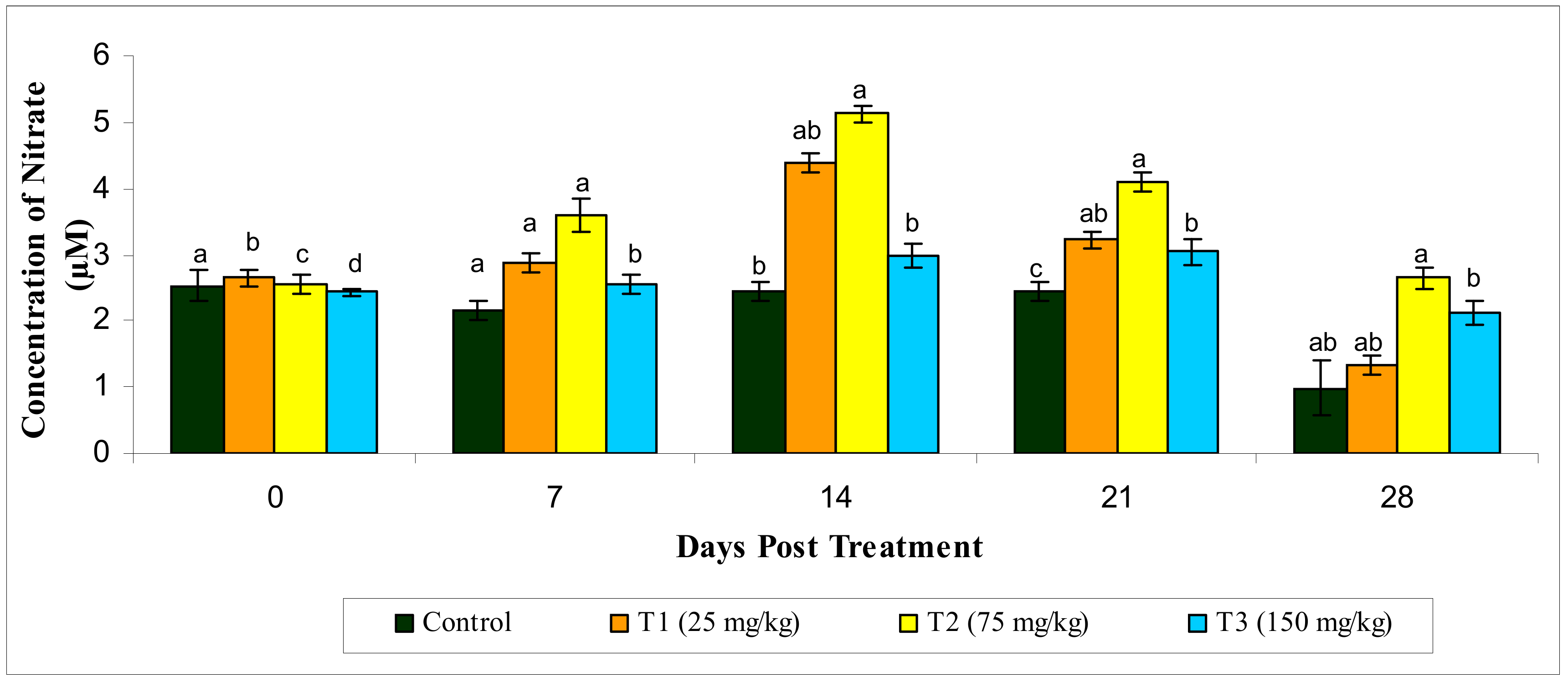
3.2.6. Reactive Oxygen Species and Reactive Oxygen Species
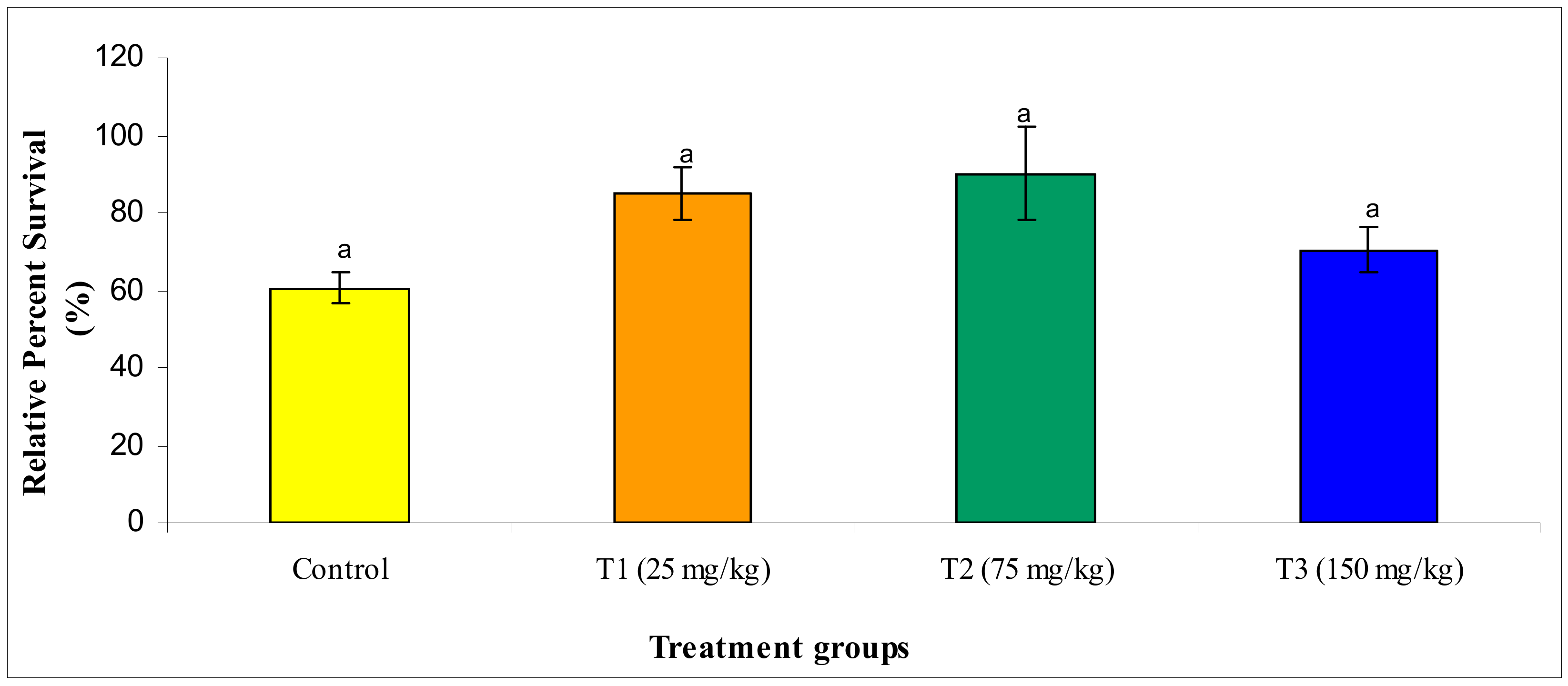
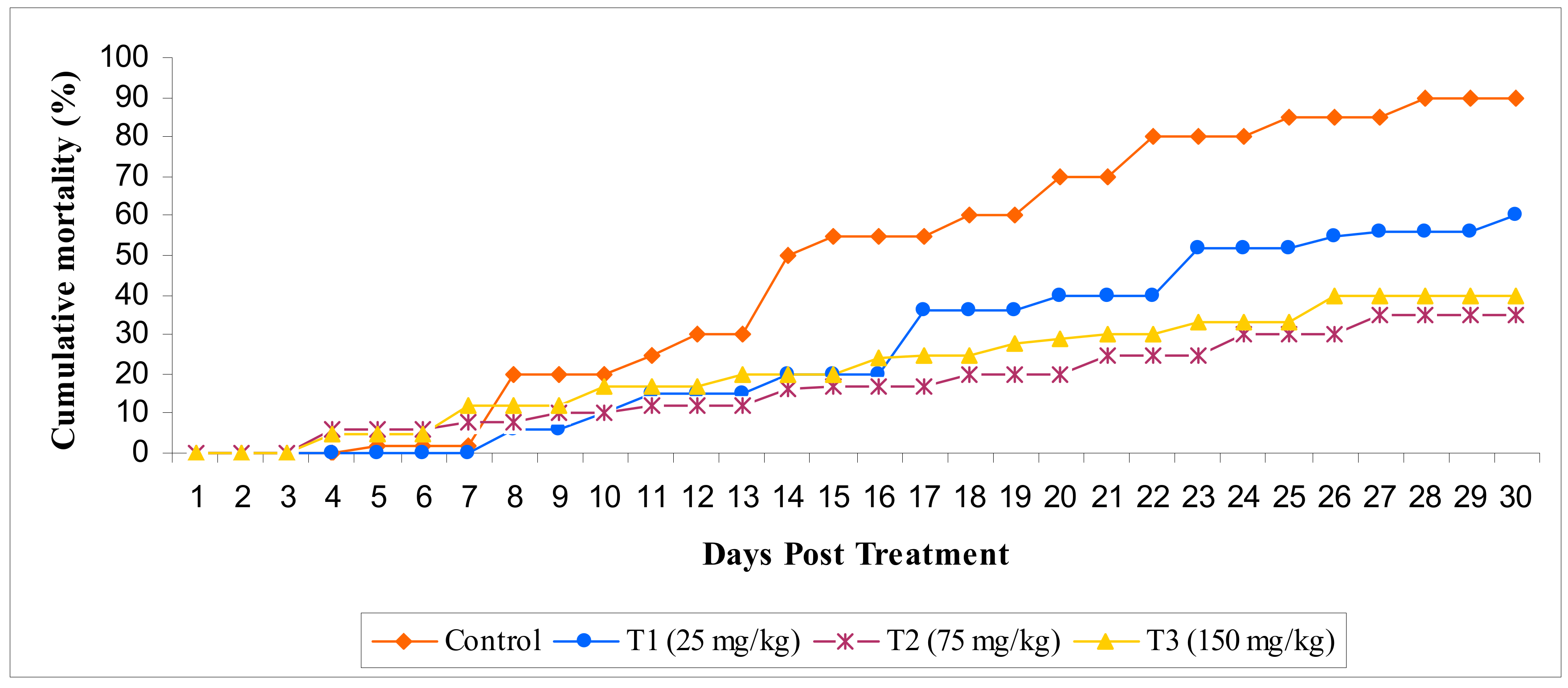
3.2.7. Challenge Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kar, B.; Mohanty, J.; Hemaprasanth, K.P.; Sahoo, P.K. The immune response in rohu, Labeorohita (Actinopterygii: Cyprinidae) to Argulussiamensis (Branchiura: Argulidae) infection: Kinetics of immune gene expression and innate immune response. Aquac. Res. 2013, 46, 1292–1308. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. Bacterial identification for publication: When is enough enough? J. Clin. Microbiol. 2002, 40, 1887–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanty, B.R.; Sahoo, P.K. Edwardsiellosis in fish: A brief review. J. Biosci. 2007, 32, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Castro, N.; Toranzo, A.E.; Barja, J.L.; Nunez, S.; Magarinos, B. Characterization of Edwardsiellatarda strains isolated from turbot, Psetta maxima (L.). J. Fish Dis. 2006, 29, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Sakai, M. Current research status of fish immunostimulants. Aquaculture 1999, 172, 63–92. [Google Scholar] [CrossRef]
- Siwicki, A.K.; Anderson, D.P.; Rumsey, G.L. Dietary intake of immunostimulants by rainbow trout affects non-specific immunity and protection against furunculosis. Vet. Immu. Immunopathol. 1994, 41, 125–139. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microl. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Jyotirmayee, P.; Sachidananda, D.; Das Basanta, K. Antibacterial activity of freshwater microalgae: A review. Afr. J. Pham. Pharmal. 2004, 8, 809–818. [Google Scholar] [CrossRef]
- Genovese, G.; Leitner, S.; Minicante, S.A.; Lass-Flörl, C. The Mediterranean red alga Asparagopsistaxiformis has antifungal activity against Aspergillus species. Mycoses 2013, 56, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Dudgeon, D. The contribution of scientific information to the conservation and management of freshwater biodiversity in tropical Asia. In Aquatic Biodiversity: A Celebratory Volume in Honour of Henri J. Dumont; Martens, K., Ed.; Springer: Dordrecht, The Netherlands, 2003; pp. 295–314. [Google Scholar]
- Sapna, M.; Swati, R.; Anil, K.; Meena, V. Medicinal attributes of Acacia nilotica Linn, a comprehensive review on ethno pharmacological claims. Int. J. Pharm. Life Sci. 2011, 2, 830–837. [Google Scholar]
- Fernando, S.; Kim, M.; Son, K.T.; Jeong, Y.; Jeon, Y.J. Antioxidant activity of marine algal polyphenolic compounds: A mechanistic approach. J. Med. Food. 2016, 19, 615–628. [Google Scholar] [CrossRef]
- Iwai, K. Antidiabetic and antioxidant effects of polyphenols in brown alga Ecklonia stolonifera in genetically diabetic KK- A(Y) Mice. Plant Foods Hum. Nutr. 2008, 63, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Murakami-Funayama, K.; Miyashita, K. Anti-obesity and anti-diabetic effects of fucoxanthin on diet-induced obesity conditions in a murine model. Mol. Med. Rep. 2009, 2, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, M.; Miyashita, T.; Nishikawa, S.; Emi, S.; Tsukui, T.; Beppu, F.; Okada, T.; Miyashita, K. Fucoxanthin regulates adipocytokine mRNA expression in white adipose tissue of diabetic/obese KK- Ay mice. Arch. Biochem. Biophys. 2010, 504, 17–25. [Google Scholar] [CrossRef]
- Tsuyoshi, S.; Takeshi, Y.; Takao, Y. Utilization of waste Porphyra products as eco-friendly feed ingredients. Bull. Fish. Res. Agency 2010, 29, 19–27. [Google Scholar]
- Wassef, E.A.; El-Masry, M.H.; Mikhail, F.R. Growth enhancement and muscle structure of striped mullet, (Mugilcephalus L.) fingerlings by feeding algal meal-based diets. Aquac. Res. 2001, 32, 315–322. [Google Scholar] [CrossRef]
- Nair, R.; Chabhadiya, R.; Chanda, S. Marine algae screening for a potent antibacterial agent. J. Herb. Pharmacother. 2007, 7, 73–86. [Google Scholar] [CrossRef]
- Athukorala, Y.; Lee, K.W.; Kim, S.K.; Jeon, Y.J. Anticoagulant activity of marine green and brown algae collected from Jeju Island in Korea. Bioresour. Technol. 2006, 98, 1711–1716. [Google Scholar]
- Richards, T.J.; Kern, E.R.; Glasgow, L.A.; Overall, J.C.; Deign, E.F.; Hatch, M.T. Antiviral activity of extracts from marine algae, Antimicrobial Agents. Chemotherapy 1978, 14, 24–30. [Google Scholar]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar]
- National Research Council (NRC). Nutrient Requirements of Fish; The National Academies Press: Washington, DC, USA, 1993. [Google Scholar]
- Sattanathan, G.; Thanapal, P.; Padmapriya, S.S.; Vijaya, A.A.; Park, S.; Kim, I.H.; Balasubramanian, B. Influences of dietary inclusion of algae Chaetomorphaaerea enhanced growth performance, immunity, haematological response and disease resistance of Labeorohita challenged with Aeromonashydrophila. Aquac. Rep. 2020, 17, 1–8. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Kirubakaran, C.J.W.; Alexander, C.P.; Michael, R.D. Enhancement of non-specific immune responses and disease resistance on oral administration of Nyctanthesarbortristis seed extract in Oreochromismossambicus (Peters). Aquac. Res. 2010, 41, 1630–1639. [Google Scholar] [CrossRef]
- Michael, R.D.; Srinivas, S.D.; Sailendri, K. A rapid method for repetitive bleeding in fish. Indian. J. Exp. Biol. 1994, 32, 838–839. [Google Scholar]
- Stasiack, A.S.; Bauman, C.P. Neutrophil activity as a potent indicator for concomitant analysis. Fish Shellfish Immun. 1996, 37, 53–59. [Google Scholar]
- Parry, R.M.; Chandan, R.C.; Shahani, K.M. A rapid and sensitive assay of muramidase. Proc. Soc. Exp. Biol. Med. 1965, 119, 384–386. [Google Scholar] [CrossRef]
- Hutchinson, T.H.; Manning, M.J. Seasonal trends in serum lysozyme activity and total protein concentration in dab (Limandalimanda L.) sampled from Lyme Bay, UK. Fish Shellfish Immun. 1996, 6, 473–482. [Google Scholar] [CrossRef]
- Quade, M.J.; Roth, J.A. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunopathol. 1997, 58, 239–248. [Google Scholar] [CrossRef]
- Sahoo, P.K.; Kumari, J.; Mishra, B.K. Non-specific immune responses in juveniles of Indian major carps. J. Appl. Ichthyol. 2005, 21, 151–155. [Google Scholar] [CrossRef]
- Dautremepuits, C.; Betoulle, S.; Paris-Palacios, S.; Vernet, G. Humoral immune factors modulated by copper and chitosan in healthy and parasitised carp (Cyprinuscarpio L.) by Ptychobothrium sp. (Cestoda). Aquat. Toxicol. 2004, 68, 325–338. [Google Scholar] [CrossRef]
- Rao, Y.V.; Chakrabarti, R. Enhanced anti-protease in Labeorohita fed with diet containing herbal ingredients. Indian J. Clin. Biochem. 2004, 19, 132–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divyanageswari, M.; Christybapita, D.; Dinakaran Michael, R. Enhancement of nonspecific immunity and disease resistance in Oreochromismossambicus by Solanum trilobatum leaf fractions. Fish Shellfish Immun. 2007, 23, 249–259. [Google Scholar] [CrossRef]
- Swain, P.; Behyra, A.; Dash, D.; Nayak, S.K. Serum antibody response of Indian major carp, Labeorohita to three species of pathogenic bacteria, Aeromonas hdyrophila, Edwadsiellatarda and Pesudomonasfluorescens. Vet. Immunol. Immunopathol. 2007, 117, 137–141. [Google Scholar] [CrossRef]
- Anderson, D. Immunostimulants, Adjuvants, and Vaccine carriers in fish: Applications to Aquaculture. Annual. Rev. Fish Dis. 1992, 2, 281–307. [Google Scholar] [CrossRef]
- Mukherjee, P.K. Quality Control of Herbal Drugs: An Approach to Evaluation of Botanicals; Business Horizons Phar—Maceutical: New Delhi, India, 2002; p. 800. [Google Scholar]
- Couso, N.; Castro, R.; Magariños, B.; Obach, A.; Lamas, J. Effect of oral administration of glucans on the resistance of gilthead seabream to pasteurellosis. Aquaculture 2003, 219, 99–109. [Google Scholar] [CrossRef]
- Sakai, M.; Kamiya, H.; Atusuta, S.; Kobayashi, M. Immunomodulatory effects on rainbow trout, Oncorhynchus mykiss injected with the extracts of abalone, Haliotis discus hannai. J. Appl. Ichthyol. 1991, 7, 54–59. [Google Scholar] [CrossRef]
- Punitha, S.M.J.; Babum, M.M.; Sivaram, V.; Shankar, V.S.; Dhas, S.A.; Mahesh, T.C.; Immuanel, G.; Citrarasu, T. Immunostimulating influence of herbal biomedicines on nonspecific immunity in Grouper Epinephelustauvina juveniles against Vibrioharveyi infection. Aquac. Int. 2008, 16, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Citarasu, T.; Sivaram, V.; Immanuel, G.; Rout, N.; Murugan, V. Influence of selected Indian immunostimulant herbs against white spot syndrome virus (WSSV) infection in black tiger shrimp, Penaeusmonodon with reference to haematological, biochemical and immunological changes. Fish Shellfish Immun. 2006, 21, 372–384. [Google Scholar] [CrossRef]
- Sahu, S.; Das, B.K.; Mishra, B.K.; Pradhan, J.; Sarangi, N. Effect of Allium sativum on the immunity of Labeorohita infected with Aeromonashydrophila. J. Appl. Ichthyol. 2007, 23, 80–86. [Google Scholar] [CrossRef]
- Rao, Y.V.; Das, B.K.; Jyotirmayee, P.; Chakrabarti, R. Effect of Achyranthesaspera on the immunity and survival of Labeorohita infected with Aeromonashydrophila. Fish Shellfish Immun. 2006, 20, 263–273. [Google Scholar]
- Salton, M.; Ghuysen, J.M. The structure of di-and tetra-saccharides released from cell walls by lysozyme and Streptomyces F1 enzyme and the β (1→4) N-acetylhexosaminidase activity of these enzymes. Biochim. Biophys. Acta 1959, 32, 552–554. [Google Scholar] [CrossRef]
- Irwin, D.M.; Gong, Z.M. Molecular evolution of vertebrate goose-type lysozyme genes. J. Mol. Evol. 2003, 56, 234–242. [Google Scholar] [CrossRef]
- Sun, B.; Wang, G.; Xie, H.; Gao, Q.; Nie, P. Gene structure of goose-type lysozyme in the mandarin fish Sinipercachuatsi with analysis on the lytic activity of its recombinant in Escherichiacoli. Aquaculture 2006, 252, 106–113. [Google Scholar] [CrossRef]
- Jian, J.; Wu, Z. Effects of traditional Chinese medicine on nonspecific immunity and disease resistance of large yellow croaker, Pseudosciaenacrocea (Richardson). Aquaculture 2003, 218, 1–9. [Google Scholar] [CrossRef]
- Yin, G.; Jeney, G.; Racz, T.; Xu, P.; Jun, X.; Jeney, Z. Effect of two Chinese herbs (Astragalus radix and Scutellariaradix) on non specific immune response of tilapia, Oreochromisniloticus. Aquaculture 2006, 253, 39–47. [Google Scholar] [CrossRef]
- Amar, E.C.; Kiron, V.; Satoh, S.; Watanabe, T. Enhancement of innate immunity in rainbow trout (Oncorhynchus mykiss Walbaum) associated with dietary intake of carotenoids from natural products. Fish Shellfish Immun. 2004, 16, 527–537. [Google Scholar] [CrossRef]
- Grattendick, K.; Stuart, R.; Roberts, E.; Lincoln, J.; Lefkowitz, S.S.; Bollen, A. Alveolar macrophage activation by myeloperoxidase: A model for exacerbation of lung inflammation. Am. J. Respir. Cell Mol. Biol. 2002, 26, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Lau, D.; Mollnau, H.; Eiserich, J.P.; Freeman, B.A.; Daiber, A.; Gehling, U.M.; Brümmer, J.; Rudolph, V.; Münzel, T.; Heitzer, T.; et al. Myeloperoxidase mediates neutrophil activation by association with CD11b/CD18 integrins. Proc. Natl. Acad. Sci. USA 2005, 102, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Bilen, S.; Unal, S.; Güvensoy, H. Effects of oyster mushroom (Pleurotusostreatus) and nettle (Urticadioica) methanolic extracts on immune responses and resistance to Aeromonas hydrophila in rainbow trout (Oncorhynchus mykiss). Aquaculture 2016, 454, 90–94. [Google Scholar] [CrossRef]
- Alexander, C.P.; Kirubakaran, C.J.W.; Michael, R.D. Water soluble fraction of Tinosporacordifolialeaves enhanced the non-specific immune mechanisms and disease resistance in Oreochromismossambicus. Fish Shellfish Immun. 2010, 29, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Neumann, N.F.; Stafford, J.L.; Barreda, D.; Ainsworth, A.J.; Belosevic, M. Antimicrobial mechanisms of fish phagocytes and their role in host defense. Dev. Comp. Immunol. 2001, 25, 807–825. [Google Scholar] [CrossRef]
- Walsh, C.J.; Toranto, J.D.; Gilliland, C.T.; Noyes, D.R.; Bo-dine, A.B.; Luer, C.A. Nitric oxide production by nurse shark (Ginglymostomacirratum) and clearnose skate (Raja eglanteria) peripheral blood leucocytes. Fish Shellfish Immun. 2006, 20, 40–46. [Google Scholar] [CrossRef]
- Sahoo, P.; Mahapatra, K.D.; Saha, J.; Barat, A.; Sahoo, M.; Mohanty, B.; Gjerde, B.; Odegård, B.; Rye, M.; Salte, R. Family association between immune parameters and resistance to Aeromonashydrophila infection in the Indian major carp, Labeorohita. Fish Shellfish Immun. 2008, 25, 163–169. [Google Scholar] [CrossRef]
- Ellis, A. Innate host defense mechanisms of fish against viruses and bacteria. Dev. Comp. Immunol. 2001, 25, 827–839. [Google Scholar] [CrossRef]
- Mohanty, B.; Sahoo, P.; Mahapatra, B.; Saha, J. Innate immune responses in families of Indian major carp, Labeorohita, differing in their resistance to Edwardsiellatarda infection. Curr. Sci. 2007, 24, 1270–1274. [Google Scholar]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.S. Inonotus obliquus containing diet enhances the innate immune mechanism and disease resistance in olive flounder Paralichythysolivaceus against Uronemamarinum. Fish Shellfish Immun. 2012, 32, 1148–1154. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Kim, M.C.; Kim, J.S.; Balasundaram, C.; Heo, M.S. Effect of Coriolus versicolor supplemented diet on innate immune response and disease resistance in kelp grouper Epinephelusbruneus against Listonella anguillarum. Fish Shellfish Immun. 2012, 32, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Leiro, J.M.; Castro, R.; Arranz, J.A.; Lamas, J. Immunomodulating activities of acidic sulphatedpolysaccha- rides obtained from the seaweed Ulva rigida C. Agardh. Int. Immunopharmacol. 2007, 7, 879–888. [Google Scholar] [CrossRef]
- Plumb, J.A. Edwardsiella septicaemias. In Fish Diseases and Disorders, Viral, Bacterial, and Fungal Infections; Woo, P.T.K., Bruno, D.W., Eds.; Centre for Agriculture and Bioscience International: Oxon, UK, 1999; pp. 479–521. [Google Scholar]
- Park, K.H.; Jeong, H.D. Enhanced resistance against Edwardsiella tarda infection in tilapia (Oreochromisniloticus) by administration of protein-bound polysaccharide. Aquaculture 1996, 143, 135–143. [Google Scholar] [CrossRef]
- Jacobson, D.M.; Andersen, R.A. The discovery of mixotrophy in photosynthetic species of Dinophysis (Dinophyceae): Light and electron microscopical observations of food vacuoles in Dinophysis acuminata, D. norvegica and two heterotrophic dinophysoid dinoflagellates. Phycologia 1994, 33, 97–110. [Google Scholar] [CrossRef]
- Shanmughapriya, S.; Manilal, A.; Sujith, S.; Selvin, J.; Seghal Kiran, G.; Natarajaseenivasan, K. Antimicrobial activity of seaweeds extracts against multiresistant pathogens. Ann. Microbiol. 2008, 58, 535–541. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control (0 mg/kg) | T1 (25 mg/kg) | T2 (75 mg/kg) | T3 (150 mg/kg) |
|---|---|---|---|---|
| Initial Weight (g) | 50.51 ± 0.95 | 50.88 ± 1.48 | 50.61 ± 1.03 | 50.94 ± 2.20 |
| Final Weight (g) | 73.69 a ± 6.39 | 81.2 a ± 8.15 | 90.29 a ± 3.96 | 86.07 ac ± 4.81 |
| Weight Gain | 22.83 a ± 0.29 | 30.32 ab ± 0.30 | 39.67 a ± 0.19 | 35.13 b ± 0.21 |
| Specific Growth rate | 1.34 a ± 0.24 | 1.86 ab ± 0.29 | 1.87 b ± 0.25 | 2.07 b ± 0.4 |
| Feed conversion rate | 0.60 a ± 0.062 | 0.51 c ± 0.132 | 0.43 c ± 0.076 | 0.39 c ± 0.066 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sattanathan, G.; Tamizhazhagan, V.; Padmapriya, S.; Liu, W.-C.; Balasubramanian, B. Effect of Green Algae Chaetomorpha antennina Extract on Growth, Modulate Immunity, and Defenses against Edwardsiella tarda Infection in Labeo rohita. Animals 2020, 10, 2033. https://doi.org/10.3390/ani10112033
Sattanathan G, Tamizhazhagan V, Padmapriya S, Liu W-C, Balasubramanian B. Effect of Green Algae Chaetomorpha antennina Extract on Growth, Modulate Immunity, and Defenses against Edwardsiella tarda Infection in Labeo rohita. Animals. 2020; 10(11):2033. https://doi.org/10.3390/ani10112033
Chicago/Turabian StyleSattanathan, Govindharajan, Vairakannu Tamizhazhagan, Swaminathan Padmapriya, Wen-Chao Liu, and Balamuralikrishnan Balasubramanian. 2020. "Effect of Green Algae Chaetomorpha antennina Extract on Growth, Modulate Immunity, and Defenses against Edwardsiella tarda Infection in Labeo rohita" Animals 10, no. 11: 2033. https://doi.org/10.3390/ani10112033