Isolation and Characterization of a Variant Psedorabies Virus HNXY and Construction of rHNXY-∆TK/∆gE

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells, Viruses and Plasmids
2.2. Virus Isolation and Purification
2.3. Neutralizing Test
2.4. PCR Amplification of Full-Length gB, gE, gC, gD, TK, RR1 and RR2 of HNXY and Phylogenetic Analysis
2.5. One-Step Growth Analysis
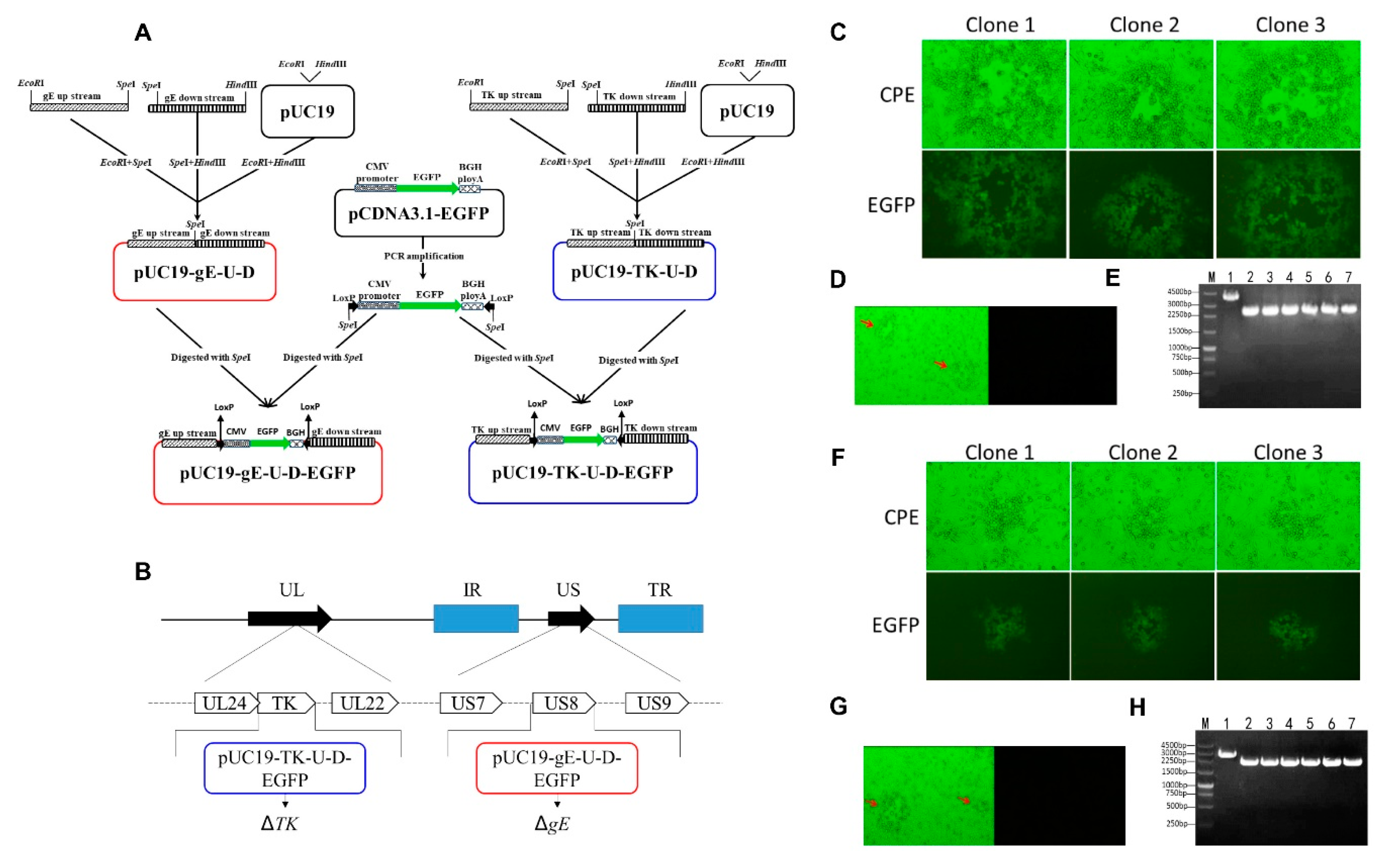
2.6. Construction of Transfer Vectors
2.7. Generation of rHNXY-∆TK/∆gE
2.8. Animal Experiment
2.8.1. Experimental Infection of Mice
2.8.2. Experimental Infection of Pigs
2.9. Accession Numbers
2.10. Statistical Analyses
3. Results
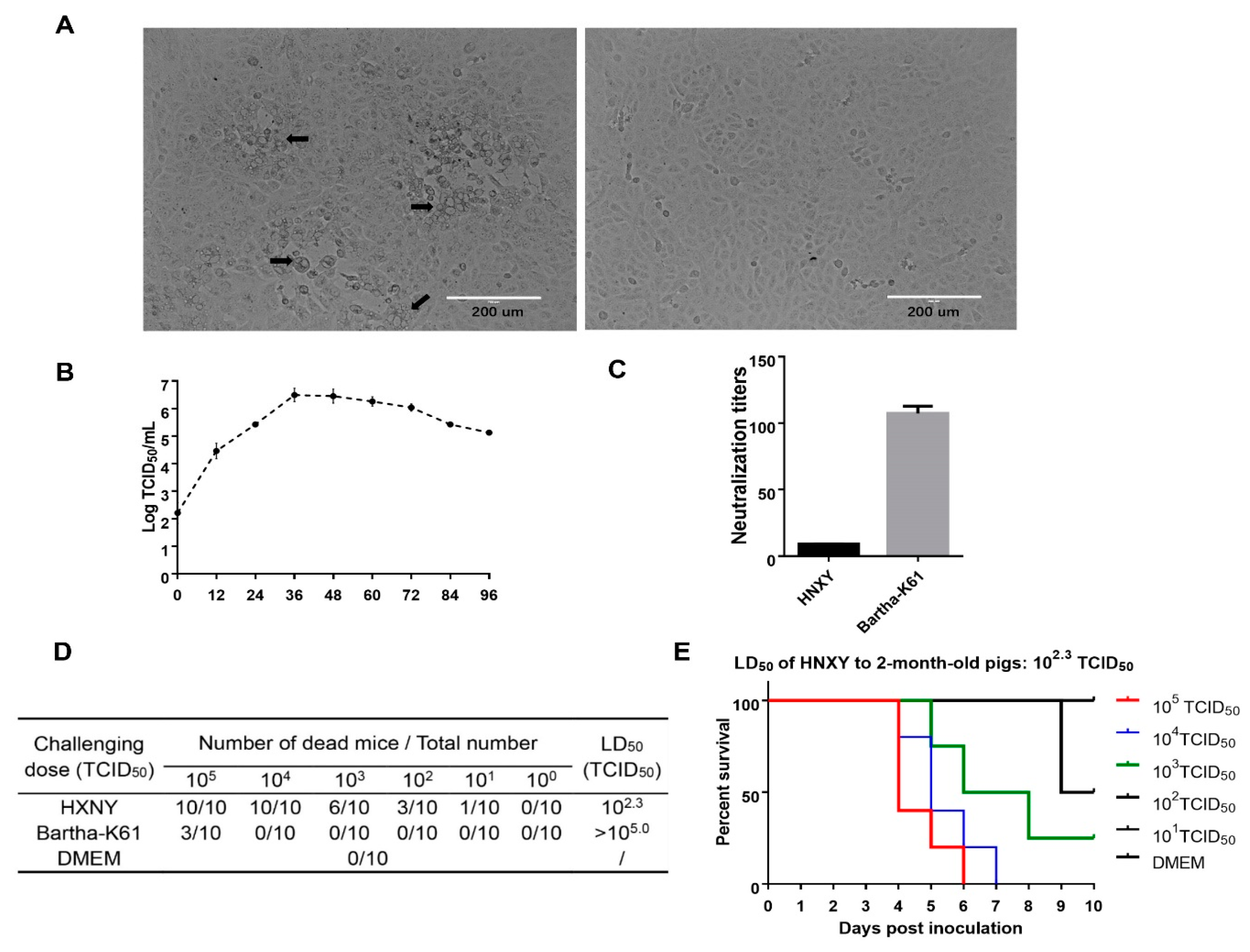
3.1. Virus Isolation
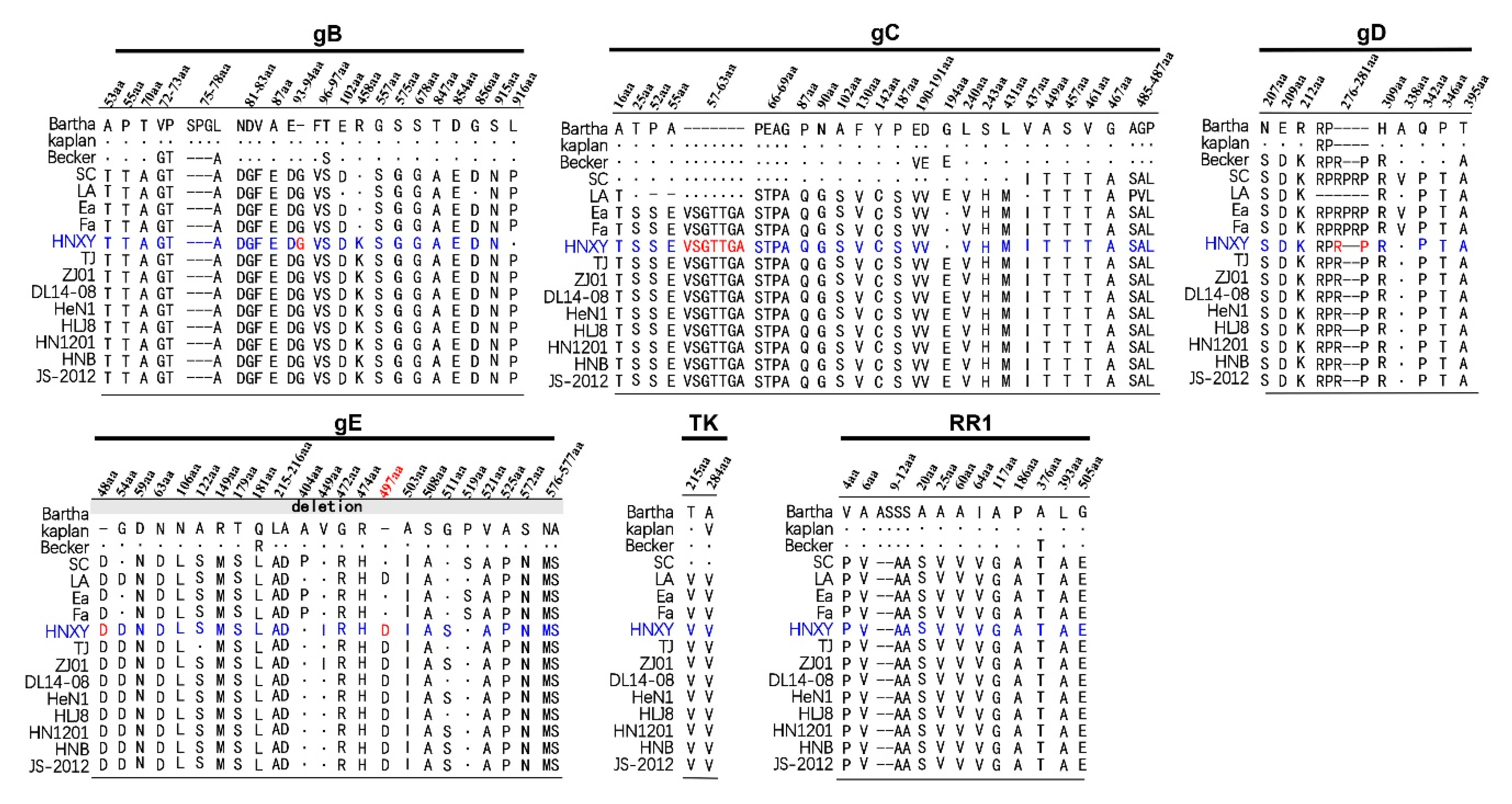
3.2. Genetic Variation Analysis of gB, gE, gC, gD, TK, RR1 and RR2 Proteins
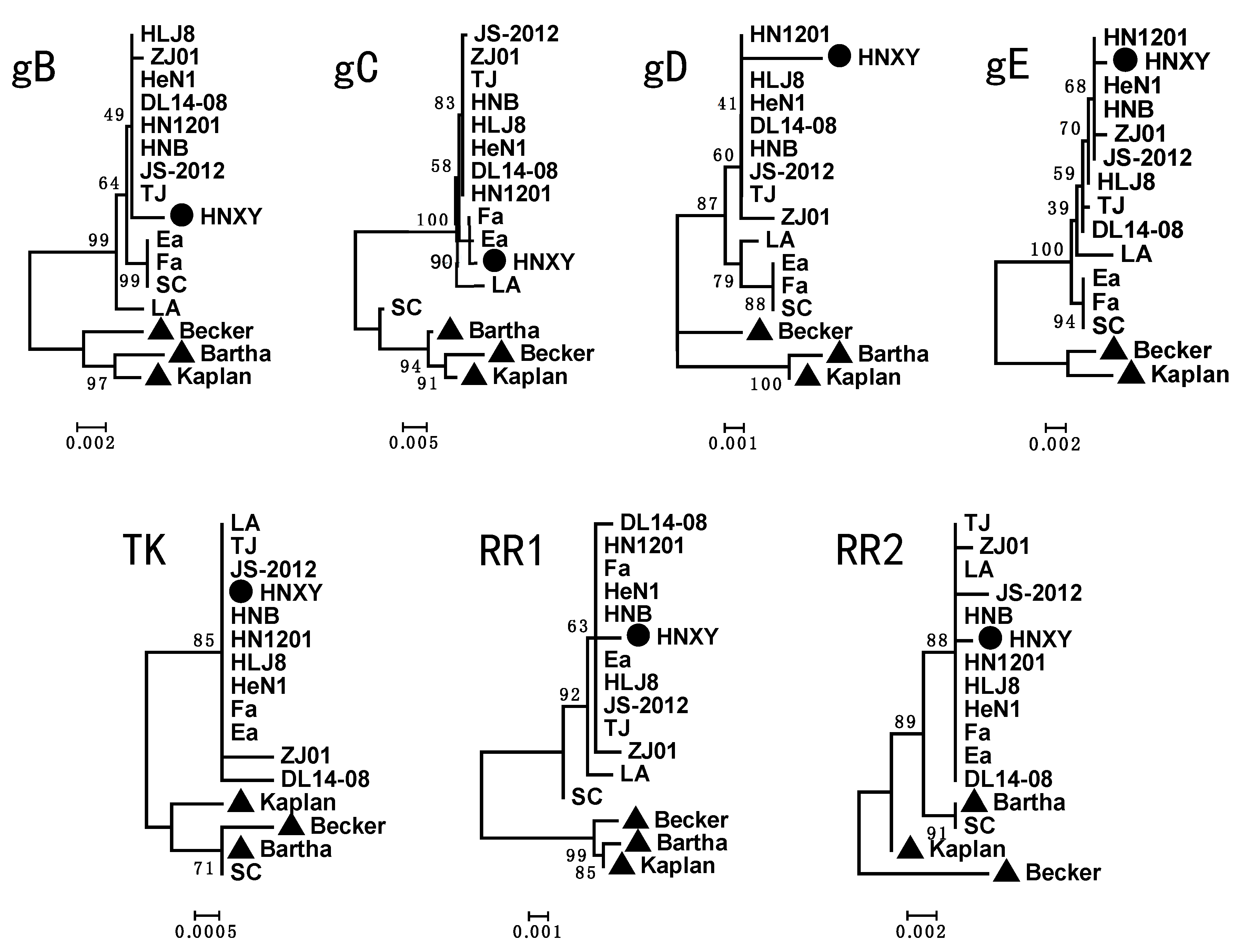
3.3. Phylogenetic Tree Analysis
3.4. Generation of rHNXY-∆TK/∆gE
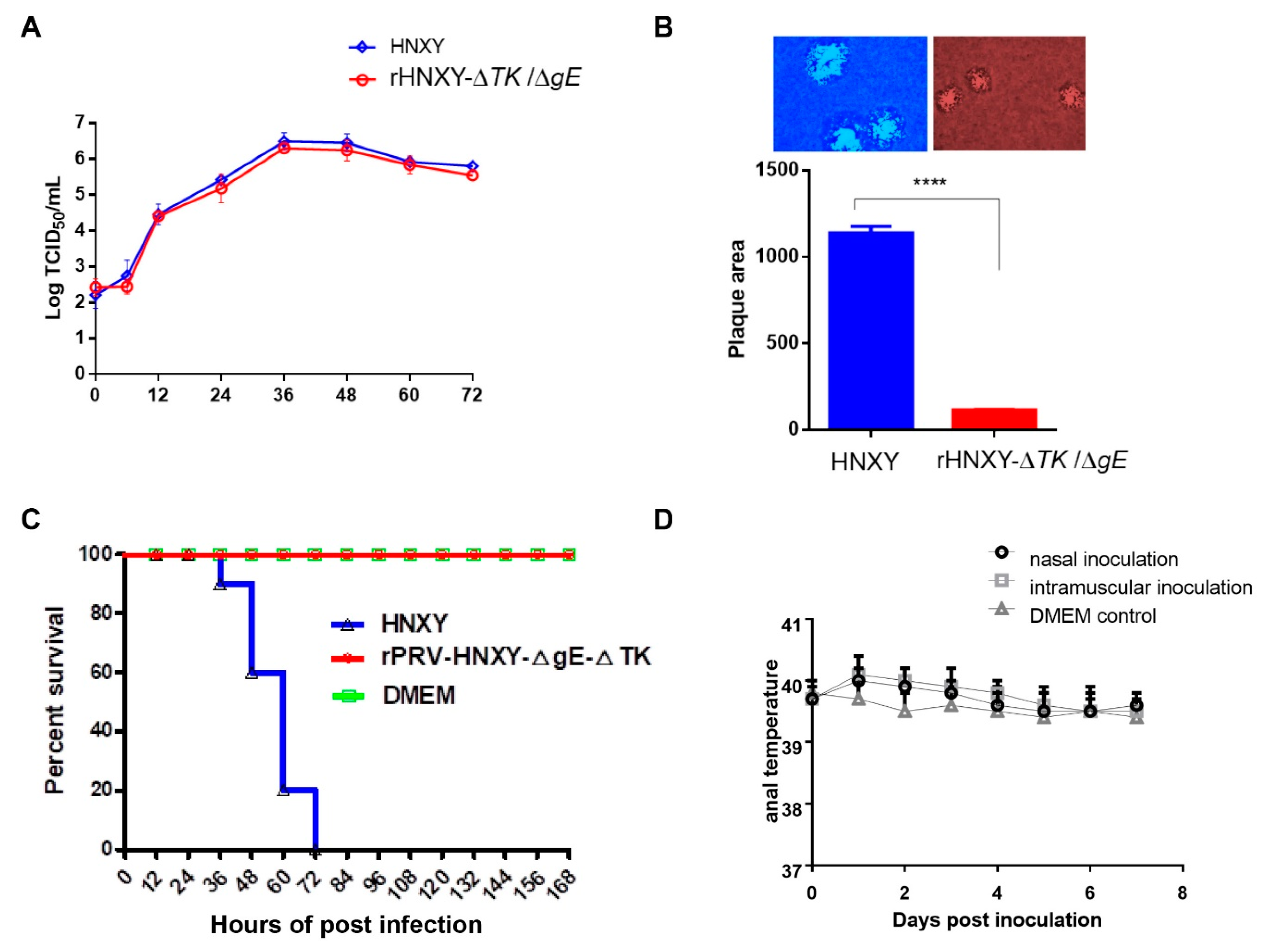
3.5. Growth Properties of rHNXY-∆TK/∆gE and HNXY In Vitro
3.6. Pathogenicity of rHNXY-∆TK/∆gE and HNXY In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhou, M.; Wu, X.; Jiang, D.; Sui, C.; Chen, L.; Cong, X.; Xin, X.; Wang, G.; Li, Y.; Tian, F.; et al. Characterization of a moderately pathogenic pseudorabies virus variant isolated in China, 2014. Infect. Genet. Evol. 2019, 68, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Marcaccini, A.; Lopez, P.M.; Quiroga, M.I.; Bermudez, R.; Nieto, J.M.; Aleman, N. Pseudorabies virus infection in mink: A host-specific pathogenesis. Vet. Immunol. Immunop. 2008, 124, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol. Mol. Biol. R. 2005, 69, 462–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, C.; Zhang, Q.Z.; Tian, Z.J.; Zheng, H.; Zhao, K.; Liu, F.; Guo, J.C.; Tong, W.; Jiang, C.G.; Wang, S.J.; et al. Genomic characterization of emergent pseudorabies virus in China reveals marked sequence divergence: Evidence for the existence of two major genotypes. Virology 2015, 483, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, G.S.; Mendoza, E.S.; Pujols, R.J.; Badiola, S.I.; Ciprian, C.A.; Hernandez, B.E. Biological properties of three Mexican isolates of Aujeszky’s disease virus. Arch. Med. Res. 1997, 28, 349–354. [Google Scholar]
- Sozzi, E.; Moreno, A.; Lelli, D.; Cinotti, S.; Alborali, G.L.; Nigrelli, A.; Luppi, A.; Bresaola, M.; Catella, A.; Cordioli, P. Genomic characterization of pseudorabies virus strains isolated in Italy. Transbound. Emerg. Dis. 2014, 61. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Sozzi, E.; Grilli, G.; Gibelli, L.R.; Gelmetti, D.; Lelli, D.; Chiari, M.; Prati, P.; Alborali, G.; Boniotti, M.B.; et al. Detection and molecular analysis of Pseudorabies virus strains isolated from dogs and a wild boar in Italy. Veter Microbiol. 2015, 177, 359–365. [Google Scholar] [CrossRef]
- Tomislav, K.; Dragan, B.; Jelena, P.; Danko, D.; Lorena, J.; Besi, R.; Tomislav, B. Characterisation of pseudorabies virus in domestic pigs and wild boars in Croatia. Acta Vet. Hung. 2014, 62, 512–519. [Google Scholar]
- Yamane, I.; Ishizeki, S.; Yamazaki, H. Aujeszky’s disease and the effects of infection on Japanese swine herdproductivity: A cross-sectional study. J. Vet. Med. Sci. 2015, 77, 579–582. [Google Scholar] [CrossRef]
- Pacini, M.I.; Forzan, M.; Cilia, G.; Bernardini, L.; Marzoli, F.; Pedonese, F.; Bandecchi, P.; Fratini, F.; Mazzei, M. Detection of pseudorabies virus in wild boar foetus. Animals 2020, 10, 366. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.L.; Wu, Y.H.; Li, H.F.; Li, S.Y.; Fan, S.Y.; Wu, H.L.; Li, Y.J.; Lü, Y.L.; Han, J.; Zhang, W.C.; et al. Clinical experience and next-generation sequencing analysis of encephalitis caused by pseudorabies virus. Zhonghua Yi Xue Za Zhi 2018, 98, 1152–1157. [Google Scholar] [PubMed]
- Ai, J.-W.; Weng, S.-S.; Cheng, Q.; Cui, P.; Li, Y.-J.; Wu, H.-L.; Zhu, Y.-M.; Xu, B.; Zhang, W.-H. Human Endophthalmitis Caused By Pseudorabies Virus Infection, China, 2017. Emerg. Infect. Dis. 2018, 24, 1087–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Wang, X.; Xie, C.; Ding, S.; Yang, H.; Guo, S.; Li, J.; Qin, L.; Ban, F.; Wang, D.; et al. OUP accepted manuscript. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- An, T.Q.; Peng, J.M.; Tian, Z.J.; Zhao, H.Y.; Li, N.; Liu, Y.M.; Chen, J.Z.; Leng, C.L.; Sun, Y.; Chang, D.; et al. Pseudorabies virus variant in Bartha-K61-vaccinated pigs, China, 2012. Emerg. Infect. Dis. 2013, 19, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Dong, J.; Wang, J.; Hou, C.; Sun, H.; Yang, W.; Bai, J.; Jiang, P. A novel inactivated gE/gI deleted pseudorabies virus (PRV) vaccine completely protects pigs from an emerged variant PRV challenge. Virus Res. 2015, 195, 57–63. [Google Scholar] [CrossRef]
- Luo, Y.; Li, N.; Cong, X.; Wang, C.H.; Du, M.; Li, L.; Zhao, B.; Yuan, J.; Liu, D.D.; Li, S.; et al. Pathogenicity and genomic characterization of a pseudorabies virus variant isolated from Bartha-K61-vaccinated swine population in China. Vet. Microbiol. 2014, 174, 107–115. [Google Scholar] [CrossRef]
- Tong, W.; Liu, F.; Zheng, H.; Liang, C.; Zhou, Y.J.; Jiang, Y.F.; Shan, T.L.; Gao, F.; Li, G.X.; Tong, G.Z. Emergence of a Pseudorabies virus variant with increased virulence to piglets. Vet. Microbiol. 2015, 181, 236–240. [Google Scholar] [CrossRef]
- Wang, C.H.; Yuan, J.; Qin, H.Y.; Luo, Y.; Cong, X.; Li, Y.; Chen, J.; Li, S.; Sun, Y.; Qiu, H.J. A novel gE-deleted pseudorabies virus (PRV) provides rapid and complete protection from lethal challenge with the PRV variant emerging in Bartha-K61-vaccinated swine population in China. Vaccine 2014, 32, 3379–3385. [Google Scholar] [CrossRef]
- Cong, X.; Lei, J.L.; Xia, S.L.; Wang, Y.M.; Li, Y.; Li, S.; Luo, Y.; Sun, Y.; Qiu, H.J. Pathogenicity and immunogenicity of a gE/gI/TK gene-deleted pseudorabies virus variant in susceptible animals. Vet. Microbiol. 2016, 182, 170–177. [Google Scholar] [CrossRef]
- Tong, W.; Li, G.; Liang, C.; Liu, F.; Tian, Q.; Cao, Y.; Li, L.; Zheng, X.; Zheng, H.; Tong, G. A live, attenuated pseudorabies virus strain JS-2012 deleted for gE/gI protects against both classical and emerging strains. Antivir. Res. 2016, 130, 110–117. [Google Scholar] [CrossRef]
- Yin, Y.; Xu, Z.; Liu, X.; Li, P.; Yang, F.; Zhao, J.; Fan, Y.; Sun, X.; Zhu, L. A live gI/gE-deleted pseudorabies virus (PRV) protects weaned piglets against lethal variant PRV challenge. Virus Genes 2017, 53, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.M.; Zhou, Q.; Song, W.B.; Sun, E.C.; Zhang, M.M.; He, Q.G.; Chen, H.C.; Wu, B.; Liu, Z.F. Novel pseudorabies virus variant with defects in TK, gE and gI protects growing pigs against lethal challenge. Vaccine 2015, 33, 5733–5740. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.D.; Liu, J.T.; Wang, T.Y.; An, T.Q.; Sun, M.X.; Wang, S.J.; Fang, Q.Q.; Hou, L.L.; Tian, Z.J.; Cai, X.H. Live attenuated pseudorabies virus developed using the CRISPR/Cas9 system. Virus Res. 2016, 225, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Xiao, Y.; Yang, Q.; Wang, Y.; Sun, Z.; Zhang, C.; Yan, S.; Wang, J.; Guo, L.; Yan, H.; et al. Construction of a gE-Deleted Pseudorabies Virus and Its Efficacy to the New-Emerging Variant PRV Challenge in the Form of Killed Vaccine. Biomed. Res. Int. 2015, 2015, 684945. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Guo, L.; Jia, X.; Wang, T.; Wang, J.; Sun, Z.; Wang, L.; Li, X.; Tan, F.; Tian, K. Construction of a triple gene-deleted Chinese Pseudorabies virus variant and its efficacy study as a vaccine candidate on suckling piglets. Vaccine 2015, 33, 2432–2437. [Google Scholar] [CrossRef]
- Liang, X.; Sun, L.; Yu, T.; Pan, Y.; Wang, D.; Hu, X.; Fu, Z.; He, Q.; Cao, G. A CRISPR/Cas9 and Cre/Lox system-based express vaccine development strategy against re-emerging Pseudorabies virus. Sci. Rep. 2016, 6, 19176. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.Q.; Tong, W.; Zheng, H.; Li, L.W.; Li, G.X.; Gao, F.; Wang, T.; Liang, C.; Ye, C.; Wu, J.Q.; et al. Variations in glycoprotein B contribute to immunogenic difference between PRV variant JS-2012 and Bartha-K61. Vet. Microbiol. 2017, 208, 97–105. [Google Scholar] [CrossRef]
- Klupp, B.G.; Fuchs, W.; Weiland, E.; Mettenleiter, T.C. Pseudorabies virus glycoprotein L is necessary for virus infectivity but dispensable for virion localization of glycoprotein H. J. Virol. 1997, 71, 7687–7695. [Google Scholar] [CrossRef] [Green Version]
- Freuling, C.M.; Muller, T.F.; Mettenleiter, T.C. Vaccines against pseudorabies virus (PrV). Vet. Microbiol. 2017, 206, 3–9. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, S.; Xu, Q.; Wu, J.; Zhai, X.; Li, S.; Wang, J.; Ni, J.; Yuan, L.; Song, X.; et al. Investigation on pseudorabies prevalence in Chinese swine breeding farms in 2013–2016. Trop. Anim. Health Pro. 2018, 50, 1279–1285. [Google Scholar] [CrossRef]
- Sun, Y.; Liang, W.; Liu, Q.; Zhao, T.; Zhu, H.; Hua, L.; Peng, Z.; Tang, X.; Stratton, C.W.; Zhou, D.; et al. Epidemiological and genetic characteristics of swine pseudorabies virus in mainland China between 2012 and 2017. PeerJ 2018, 6, e5785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.T.; Liu, H.M.; Guo, Z.D.; Du, J.M.; Zhao, J.; Chen, L.; Yang, X.; Wang, X.W.; Yao, H.X.; Wang, C.Q. Investigation of etiology of massive infection with porcine pseudorabies virus in Henan and neighboring Provinces. Chin. J. Virol. 2014, 30, 441–449. [Google Scholar]
- Gu, Z.; Hou, C.; Sun, H.; Yang, W.; Dong, J.; Bai, J.; Jiang, P. Emergence of highly virulent pseudorabies virus in southern China. Can. J. Vet. Res. 2015, 79, 221–228. [Google Scholar] [PubMed]
- Mukamoto, M.; Watanabe, I.; Kobayashi, Y.; Icatlo, F.C.; Ishii, H.; Kodama, Y. Immunogenicity in Aujeszky’s disease virus structural glycoprotein gVI (gp50) in swine. Vet. Microbiol. 1991, 29, 109–121. [Google Scholar] [CrossRef]
- Riviere, M.; Tartaglia, J.; Perkus, M.E.; Norton, E.K.; Bongermino, C.M.; Lacoste, F.; Duret, C.; Desmettre, P.; Paoletti, E. Protection of mice and swine from pseudorabies virus conferred by vaccinia virus-based recombinants. J. Virol. 1992, 66, 3424–3434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ober, B.T.; Teufel, B.; Wiesmuller, K.H.; Jung, G.; Pfaff, E.; Saalmuller, A.; Rziha, H.J. The porcine humoral immune response against pseudorabies virus specifically targets attachment sites on glycoprotein gC. J. Virol. 2000, 74, 1752–1760. [Google Scholar] [CrossRef] [Green Version]
- De Wind, N.; Peeters, B.P.; Zuderveld, A.; Gielkens, A.L.; Berns, A.J.; Kimman, T.G. Mutagenesis and characterization of a 41-kilobase-pair region of the pseudorabies virus genome: Transcription map, search for virulence genes, and comparison with homologs of herpes simplex virus type 1. Virology 1994, 200, 784–790. [Google Scholar] [CrossRef]
- Fan, J.; Zeng, X.; Zhang, G.; Wu, Q.; Niu, J.; Sun, B.; Xie, Q.; Ma, J. Molecular characterization and phylogenetic analysis of pseudorabies virus variants isolated from Guangdong province of southern China during 2013–2014. J. Vet. Sci. 2016, 17, 369–375. [Google Scholar] [CrossRef]
- Li, A.; Lu, G.; Qi, J.; Wu, L.; Tian, K.; Luo, T.; Shi, Y.; Yan, J.; Gao, G.F. Structural basis of nectin-1 recognition by pseudorabies virus glycoprotein D. PLoS Pathoens. 2017, 13, e1006314. [Google Scholar] [CrossRef] [Green Version]
- Mettenleiter, T.C. Pseudorabies (Aujeszky’s disease) virus: State of the art. August 1993. Acta Vet. Hung. 1994, 42, 153–177. [Google Scholar]
- Moormann, R.J.; de Rover, T.; Briaire, J.; Peeters, B.P.; Gielkens, A.L.; van Oirschot, J.T. Inactivation of the thymidine kinase gene of a gI deletion mutant of pseudorabies virus generates a safe but still highly immunogenic vaccine strain. J. Gen. Virol. 1990, 71, 1591–1595. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Details |
|---|---|---|
| gE-U-F | 5-ccggaattccctcctcgccgccctgaccctg-3 | upstream homologous fragments flanking gE gene |
| gE-U-R | 5-ggactagtgacggagataaaacgccaccca-3 | |
| gE-D-F | 5-ggactagtataccgggagaaccggt-3 | downstream homologous fragments flanking gE gene |
| gE-D-R | 5-cccaagcttaggagcggttgtgga-3 | |
| TK-U-F | 5-ccggaattcccggttgcccacgtacag-3 | upstream homologous fragments flanking TK gene |
| TK-U-R | 5-ggactagtcccggcgcgcttccgg-3 | |
| TK-D-F | 5-ggactagtccctcgcccctcccaccc-3 | downstream homologous fragments flanking TK gene |
| TK-D-R | 5-cccaagcttccgggtcctcgccgaa-3 | |
| loxP-EGFP-F | 5-ggactagtataacttcgtatagcatacattatacgaagttattagttattaatagta-3 | EGFP expression cassette |
| loxP-EGFP-R | 5-ggactagtataacttcgtataatgtatgctatacgaagttatagccatagagcccac-3 | |
| gE-U-L | 5-agcccggtccgtagcctccgcagtac-3 | gE deletion verification |
| gE-D-R | 5-cctccgtccactcgccggcgt-3 | |
| TK-U-L | 5-ggtgcaccaggtgcaggcacag-3 | TK deletion verification |
| TK-D-R | 5-ggcgacggtcgcccgcgcgagg-3 |
| Gene | Compared with Bartha, Kaplan and Becker | Compared with SC, LA, Ea, Fa | Compared with PRV Isolated after 2012 in China | |||
|---|---|---|---|---|---|---|
| Nucleotide Sequence Homology | Amino Acid Sequence Homology | Nucleotide Sequence Homology | Amino Acid Sequence Homology | Nucleotide Sequence Homology | Amino Acid Sequence Homology | |
| gB | 98.2%–98.4% | 96.4%–97.2% | 99.5%–99.6% | 98.7%–98.9% | 99.7%–99.8% | 99.3%–99.9% |
| gC | 95.9%–96.2% | 91.7%–92.5% | 96.9%–99.9% | 94.0%–99.4% | 99.5%–99.6% | 98.6%–98.8% |
| gD | 98.5%–98.8% | 96.0%–97.5% | 99.2%–99.4% | 98.3%–98.5% | 99.4%–99.6% | 98.3%–98.5% |
| gE | 97.6%–97.8% | 95.5%–95.7% | 99.4%–99.5% | 98.8%–99.3% | 99.7%–99.9% | 99.0%–99.7% |
| TK | 99.6%–99.7% | 99.1%–99.4% | 99.7%–100% | 99.1%–99.7% | 99.9%–100% | 99.4%–99.7% |
| RR1 | 98.6% | 96.8%–97.1% | 99.7%–99.9% | 99.2%–99.5% | 99.7%–99.9% | 99.2%–99.5% |
| RR2 | 98.4%–99.5% | 97.4%–99.0% | 99.5%–99.9% | 99.0%–99.3% | 99.7%–99.9% | 99.0%–99.3% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, F.; Lv, Y.; Zhang, S.; Liu, L.; Wu, Y.; Zhao, P.; Peng, Z.; Liu, S.; Zhang, Z.; Li, W. Isolation and Characterization of a Variant Psedorabies Virus HNXY and Construction of rHNXY-∆TK/∆gE. Animals 2020, 10, 1804. https://doi.org/10.3390/ani10101804
Wu F, Lv Y, Zhang S, Liu L, Wu Y, Zhao P, Peng Z, Liu S, Zhang Z, Li W. Isolation and Characterization of a Variant Psedorabies Virus HNXY and Construction of rHNXY-∆TK/∆gE. Animals. 2020; 10(10):1804. https://doi.org/10.3390/ani10101804
Chicago/Turabian StyleWu, Fengsun, Yujin Lv, Shijun Zhang, Lingling Liu, Yuchen Wu, Pandeng Zhao, Zhifeng Peng, Shengli Liu, Zhonghua Zhang, and Wengang Li. 2020. "Isolation and Characterization of a Variant Psedorabies Virus HNXY and Construction of rHNXY-∆TK/∆gE" Animals 10, no. 10: 1804. https://doi.org/10.3390/ani10101804