Development of wheat—Hordeum chilense Chromosome 2Hch Introgression Lines Potentially Useful for Improving Grain Quality Traits


Departamento de Genética, Escuela Técnica Superior de Ingeniería Agronómica y de Montes, Edificio Gregor Mendel, Campus de Rabanales, Universidad de Córdoba, CeiA3, ES-14071 Córdoba, Spain
*
Author to whom correspondence should be addressed.
Agronomy 2019, 9(9), 493; https://doi.org/10.3390/agronomy9090493
Submission received: 25 July 2019
/
Revised: 16 August 2019
/
Accepted: 23 August 2019
/
Published: 28 August 2019
(This article belongs to the Special Issue Chromosome Manipulation for Plant Breeding Purposes)
Abstract
:The chromosome 2Hch of Hordeum chilense. has the potential to improve seed carotenoid content in wheat as it carries a set of endosperm carotenoid-related genes. We have obtained structural changes in chromosome 2Hch in a common wheat (Triticum aestivum L. “Chinese Spring”) background by crossing a wheat double disomic substitution 2Hch(2D) and 7Hch(7D) line with a disomic addition line carrying chromosome 2Cc from Aegilops cylindrica Host.. Seven introgressions of chromosome 2Hch into wheat were characterized by fluorescence in situ hybridization (FISH) and DNA markers. Chromosome-specific simple sequence repeats (SSRs) were used for identifying wheat chromosomes. In addition, we tested 82 conserved orthologous set (COS) markers for homoeologous group 2, of which 65 amplified targets in H. chilense and 26 showed polymorphism between H. chilense and wheat. A total of 24 markers were assigned to chromosome 2Hch with eight allocated to 2HchS and 16 to 2HchL. Among the seven introgressions there was a disomic substitution line 2Hch(2D), a ditelosomic addition line for the 2HchL arm, an isochromosome for the 2HchL arm, a homozygous centromeric 2HchS·2DL translocation, a double monosomic 2HchS·2DL plus 7HchS·D translocation, a homozygous centromeric 7HchS·2HchL translocation and, finally, a 2HchL·7HchL translocation. Wheat—H. chilense macrosyntenic comparisons using COS markers revealed that H. chilense chromosome 2Hch exhibits synteny to wheat homoeologous group 2 chromosomes, and the COS markers assigned to this chromosome will facilitate alien gene introgression into wheat. The genetic stocks developed here include new wheat—H. chilense recombinations which are useful for studying the effect of chromosome 2Hch on grain quality traits.
1. Introduction
Narrow genetic diversity often limits the improvement of many traits in wheat. The introgression of genes from wild relatives to wheat has become a widely recognized genetic approach for increasing genetic diversity and, hence, the need to explore primary, secondary and tertiary gene pools of wheat has grown [1,2]. Hordeum chilense Roem. et Schultz. is a diploid wild barley that exhibits advantageous agronomic and quality characteristics [3,4,5,6]. Furthermore, its high crossability with other species of the tribe Triticeae, such as both durum and common wheat, [3,4,5] make it useful in cereal breeding.
Addition and substitution lines of alien chromosomes in a common wheat background, are useful for introgressing alien chromosomal segments carrying genes of agronomical interest into wheat. Chromosome addition lines of H. chilense in the Triticum aestivum L. cultivar “Chinese Spring” have been obtained including five for chromosomes 1Hch, 4Hch, 5Hch, 6Hch and 7Hch and the ditelosomic addition line for the short arm of chromosome 2Hch [7]. Fertile wheat lines carrying deletions and translocations involving chromosome 3Hch from H. chilense have also been obtained [8]. However, no addition or substitution lines in a common wheat background have been developed for chromosome 2Hch.
The location of agronomic traits on specific H. chilense chromosomes have been carried using these available wheat—H. chilense addition and substitution lines, such as resistance to greenbug (Schizaphis graminum Rond.) [9] and endosperm prolamins located on chromosome 1Hch [10,11]; resistance to Septoria tritici on chromosome 4Hch [12]; tolerance to salt on chromosomes 1Hch, 4Hch and 5Hch [13]; fertility restoration on chromosome 6Hch [14]; and carotenoid content on chromosome 7Hch [15]. Wheat—H. chilense translocation or recombinant lines have also been generated using both addition and substitution lines of H. chilense chromosomes in wheat background [8,11,16,17,18].
Chromosome 2Hch has the potential to improve seed carotenoid content in wheat. Genetic studies of yellow pigment content (YPC) in H. chilense revealed that chromosome 2Hch showed a significant association with YPC [19] and four endosperm carotenoid-related genes have been genetically mapped to chromosome 2Hch, such as geranyl geranyl pyrophosphate synthase (Ggpps1) for geranylgeranyl diphosphate synthesis, zeta-carotene desaturase (Zds), beta-carotene hydroxylase 3 (Hyd3) from the carotenoid biosynthetic pathway and polyphenol oxidase 1 gene (Ppo1) implicated in plant tissue enzymatic browning [20].
Molecular markers that are able to distinguish H. chilense chromosome 2Hch in wheat background provide a useful tool for selection. The conserved orthologous set (COS) [21] represents an important reservoir of markers that allow comparative studies with wheat and barley and their transference to H. chilense is a main goal.
The aims of this work were the following: (a) to obtain wheat—H. chilense chromosome 2Hch introgression lines; (b) to characterize the lines obtained by fluorescence in situ hybridization (FISH) and chromosome-specific simple sequence repeat (SSR) markers; (c) to transfer COS markers to H. chilense and to determine their arm location within 2Hch and (d) to compare the arm location with wheat and barley homoeologous group 2.
2. Materials and Methods
2.1. Plant Material
A “Chinese Spring” (CS) wheat—H. chilense double 2Hch(2D)-7Hch(7D) disomic substitution line, previously obtained at the University of Córdoba (results not shown), was used for inducing structural changes in chromosome 2Hch using gametocidal chromosome 2Cc from Aegilops cylindrica host. The double 2Hch(2D) and 7Hch(7D) disomic substitution line was obtained by pollinating tritordeum (the fertile amphiploid between H. chilense and T. turgidum L., AABBHchHch, 2n = 6x = 42) with a wheat disomic addition line for gametocidal chromosome 2Cc from Ae. cylindrica Host. following the breeding procedure described in [8]. The double substitution 2Hch(2D) and 7Hch(7D) line was pollinated with the wheat disomic addition line for the gametocidal chromosome 2Cc from Ae. cylindrica. The F1 plants monosomic for 2Hch, 7Hch and 2Cc were selfed for four generations.
2.2. Fluorescence In Situ Hybridization (FISH)
The excised root tips were pretreated with ice water for 24 h and then fixed in acetic ethanol: acetic acid (3:1, v/v), as described previously [8]. The FISH protocol was carried out as described by [22]. The pAs1 sequence (1 kb) isolated from Aegilops tauschii Coss. [23] and H. chilense genomic DNA were used as probes. The pAs1 probe hybridizes to D-genome chromosomes of wheat [24] and Hch-genome chromosomes from H. chilense [25]. The pAs1 probe and H. chilense DNA were labeled with biotin-16-dUTP (Roche Diagnostics, Switzerland) and with digoxigenin-11-dUTP (Roche Diagnostics, Switzerland), respectively, by nick translation. Three plants per each introgression line were analyzed.
Biotin- and digoxigenin-labelled probes were detected with streptavidin-Cy3 conjugates (Sigma, St. Louis, MO, USA) and antidigoxigenin FITC (Roche Diagnostics) antibodies, respectively. The chromosomes were counterstained with DAPI (4′,6-diamidino-2-phenylindole) and mounted in Vectashield mounting medium (Vector laboratories, Inc., Burlingame, CA, USA). A Leica DMR epifluorescence microscope was used for signal visualization. Images were captured with a Leica DFC7000T camera and processed with LEICA application suite v4.0 software (Leica, Germany).
2.3. Molecular Marker Analysis
A total of 82 COS markers from wheat homoeologous group 2 [21] were studied for their utility in H. chilense (File S1). H. chilense (line H7) and common wheat CS were used as controls. The CTAB method [26] was used for DNA extraction of young leaf tissue. The concentration of each sample was estimated using a NanoDrop 1000 Spectrophotometer (Thermo Scientific, Waltham, MA, USA). Amplifications were made using a TGradient thermocycler (Biometra, Göttingen, Germany) with 60 ng of template DNA in a 25 µl volume reaction containing 5 µl of 10× PCR Buffer, 0.5 µM of each primer, 1.5–2.0 mM MgCl2, 0.3 mM dNTPs and 0.25 U of Taq DNA polymerase (BIOTOOLS B&M Laboratories, Madrid, Spain). The PCR conditions of COS markers were as follows: 4 min at 94°C, followed by 35 cycles of 45 s at 94°C, 50s at 58°C annealing temperature, 50 s at 72°C, and a final extension step of 7 min at 72 °C.
In addition, four chromosome-specific SSR markers for the wheat D-genome were used for molecular characterization of the introgression lines [27,28]. Xgwm261 and Xgwm157 markers were used to detect 2DS and 2DL chromosome arms, respectively. Xcfd66 and Xbarc111 were used to detect 7DS and 7DL chromosome arms, respectively. Amplifications were carried out as described at GrainGenes [29] One plant from each introgression line was used for the molecular characterization. “Chinese Spring”, H. chilense, a ditelosomic 2HchS line, a ditelosomic 7HchS line, a ditelosomic 7HchL line and disomic substitution line CS 7Hch(7D) were used as controls. Ditelosomic 2HchS and ditelosomic 7HchS lines were provided by the John Innes Centre (UK). The ditelosomic 7HchL line and disomic CS 7Hch(7D) substitution line were obtained previously [17].
The amplified products were resolved using 2% agarose gels (SSRs) or polyacrylamide gels (10%, w/v; C: 2.67%) (COS) and stained with ethidium bromide or SafeView Nucleic Acid Stain (NBS Biologicals, Huntingdon, UK) incorporated in the gel. A 100 bp DNA ladder (Solis BioDyne, Tartu, Estonia) was used as a standard molecular weight marker. Kodak Digital Science 1D software (version 2.0) was used to determine the amplicon lengths.
2.4. Comparative Mapping
The orthologous relationship between the 2A, 2B, and 2D genome chromosomes of bread wheat and the 2Hch chromosome from H. chilense has been studied from the genomic perspective of wheat as described previously [30]. For the construction of the physical map, the expressed sequence tag (EST) source sequences (File S2) were used as queries in BLASTn searches against the wheat reference pseudomolecules [31] to identify the start positions (bp) of the ESTs. In this study, BLAST hits with E values smaller than 1e−10, identity % > 58.44 and alignment length > 100 bp were considered significant. The genomic start positions in bp of the best hits in wheat pseudomolecules (File S3) were used to construct a physical map of the polymorphic COS markers. The wheat reference genome sequence [31] was used to determine the centromere positions for 2A, 2B and 2D wheat chromosomes. Both the length in bp of wheat pseudomolecules, as well as the start genomic positions of the ESTs, were converted to pixels. Then, the data from the BLASTn searches were used to construct a physical map for 2A, 2B, and 2D wheat chromosomes showing the position of the source EST of the COS markers assigned to H. chilense chromosome 2Hch.
The rice locus (RAP) [21] was used to locate the COS markers in the barley genome zipper [32]. The RAP locus identifier was retrieved using the ID Converter tool [33]. The full-length barley cDNA corresponding to each rice locus was used for determination of the barley Unigene corresponding to each COS marker. The Unigene sequences were aligned in Barleymap [34] to obtain their positions in the International Barley Sequencing Consortium map [32,35].
3. Results
3.1. Cytogenetic and Molecular Characterization of Wheat—H. chilense Introgression Lines Involving Chromosome 2Hch
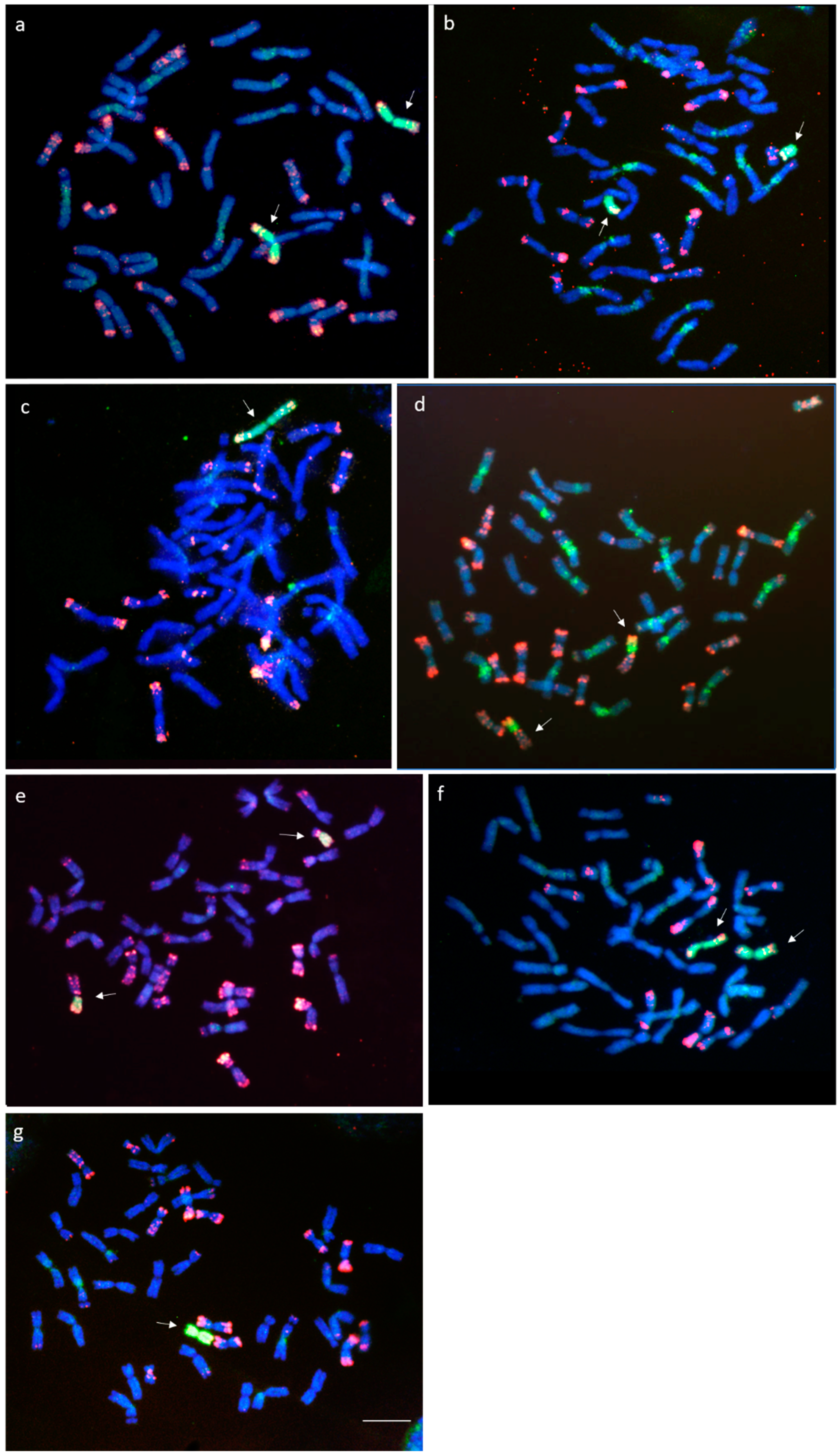
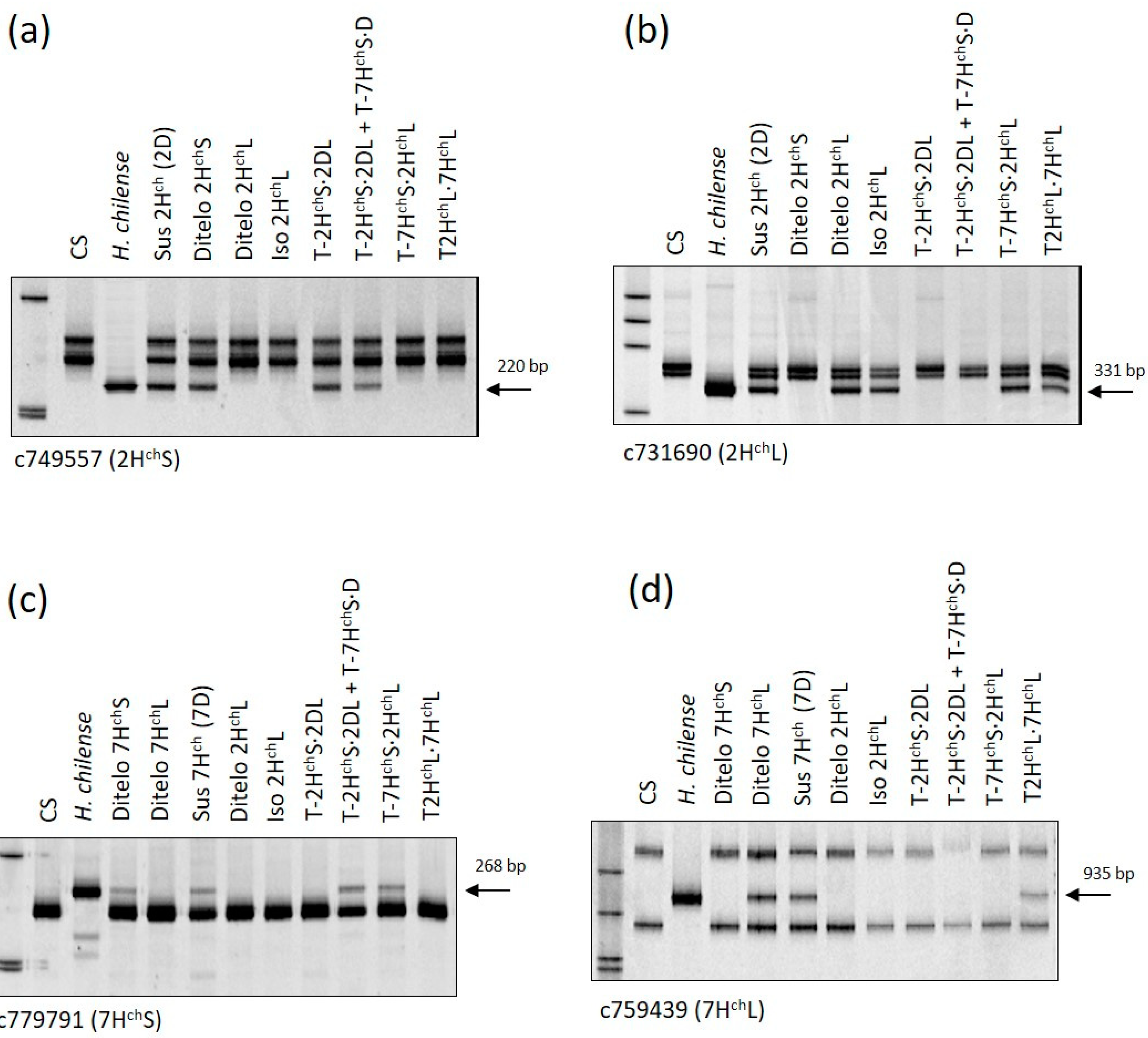
The pAs1 and H. chilense genomic DNA used as probes in FISH analysis allowed the identification of a pair of 2Hch chromosomes and the absence of the wheat 2D chromosome pair in one line with 42 chromosomes. This result indicated that this line was disomic for the substitution 2Hch(2D) (Figure 1a). The absence of 2D was tested using Xgwm261-2DS (Figure 2a) and Xgwm157-2DL (Figure 2b) molecular markers. A pair of telocentric chromosomes was identified by FISH in one line with 42 + 2t chromosomes (Figure 1b). To determine the chromosome arm involved in each introgression line, we used the c749557 COS marker mapped on the 2HchS arm and c731690 mapped on 2HchL, respectively (see Section 3.2). The presence of the c731690 marker for 2HchL and the absence of the c749557 marker for 2HchS showed that this line was ditelosomic for the 2HchL arm (Figure 3a,b).
FISH analysis revealed a line apparently carrying chromosome 2Hch (Figure 1c). Marker c731690 for 2HchL was amplified in this line, but there was no amplification of the c749557 marker for 2HchS (Figure 3a,b). These results suggested that a 2HchL·2HchL isochromosome was present in this line, and it was named Iso 2HchL. Both the ditelosomic 2HchL and Iso 2HchL lines were nullisomic for chromosome 2D, as demonstrated by the absence of amplification of both Xgwm261-2DS (Figure 2a) and Xgwm157-2DL (Figure 2b) molecular markers.
We identified two lines carrying centromeric translocations involving the 2HchS chromosome arm and wheat chromosomes. One of these lines was homozygous for the 2HchS·2DL translocation (Figure 1d). The other translocation line was a double monosomic for 2HchS·2DL and 7HchS·D translocations (Figure 1e). Chromosome-specific SSR markers confirmed the absence of 2DS (Figure 2a) and the presence of 2DL (Figure 2b) in both lines. Amplification of the c749557 marker (Figure 3a) and the absence of amplification of the c731690 marker (Figure 3b) demonstrated the presence of 2HchS and the absence of 2HchL, respectively, in both lines. COS markers c779791 and c759439, previously assigned to 7HchS and 7HchL, respectively [18], were used to detect introgression from chromosome 7Hch (Figure 3c,d). Amplification of the c779791 marker specific for the 7HchS arm (Figure 3c) indicated the presence of 7HchS translocated to an unidentified wheat fragment. The presence of pAs1 signals on the wheat small fragment indicated that the chromosome 7HchS arm was translocated to an unidentified D-genome chromosome (Figure 1e). The absence of amplification of chromosome-specific markers Xcfd66-7DS and Xbarc111-7DL demonstrated the absence of a 7D chromosome pair in this line (Figure 2c,d).
In the remaining two lines, two centromeric translocations involving 2Hch and 7Hch H. chilense chromosomes were detected. One line was homozygous for the 7HchS·2HchL translocation (Figure 1f) and the other one was monosomic for the 2HchL·7HchL translocation. Both translocation lines were nullisomic for chromosome 2D (Figure 2a,b). Chromosome-specific SSR marker patterns for 2D (Xgwm261-2DS and Xgwm157-2DL) and 7D (Xcfd66-7DS and Xbarc111-7DL) genome chromosomes are given in Figure 2a,b and Figure 2c,d, respectively. Chromosome-specific marker results for chromosome 7HchS and 7HchL are given in Figure 3c,d. Table 1 shows the chromosome constitutions of all the H. chilense introgression lines. All lines were vigorous and seed set.
3.2. Transferability and Chromosome Location of COS Markers in H. chilense
The transferability to H. chilense of 83 COS markers from wheat homoeologous group 2 was studied (File S1). First, all 83 markers were screened for polymorphisms (size polymorphisms or presence and absence) between H. chilense and common wheat. Of the 83 markers, 65 (78.3%) consistently amplified H. chilense products and 26 (40.0% of the total) were polymorphic between H. chilense and wheat (Table 2). Twenty-four of these 26 polymorphic markers were mapped to chromosome 2Hch, as demonstrated by their presence in the wheat—H. chilense 2Hch(2D) substitution line. We were unable to map the remaining two markers because they did not amplify products in any of the available wheat—H. chilense addition lines. Of the 24 COS markers mapped on chromosome 2Hch, eight were located on 2HchS and 16 were located on 2HchL, as demonstrated by their presence and absence in 2HchS or 2HchL ditelosomic lines, respectively. Table 2 summarize the characterization and chromosome arm location of wheat COS markers on H. chilense chromosome 2Hch. Figure 3a,b shows examples of amplification of homoeologous group 2 COS markers.
3.3. Wheat—H. chilense Group 2 Homoeology
To investigate wheat—H. chilense group 2 macrosyntenic relationships, the source ESTs of the 24 polymorphic COS markers mapped on chromosome 2Hch were BLASTed to the sequences of the wheat chromosomes [31]. All EST markers showed hits on wheat pseudomolecules (File S3). To produce a physical map (Figure 4), the start positions of the alignments of the best hits on the A, B, and D genomes were extracted. All markers assigned to 2HchS and 2HchL were located in the same arm of wheat homoeologous group 2 chromosomes (Table 2, Figure 4).
To determine the positions of COS markers in Barley-maps (Table 2) the align tool implemented in Barleymap [34] was used to determine the positions of COS markers in barley maps (Table 2). Four of the COS markers were not found in the barley genome zipper [32] so their position in barley could not be determined. A good correspondence between H. chilense and barley was found for the arm locations of COS markers (Table 2).
4. Discussion
A set of introgression lines involving chromosome 2Hch from H. chilense in common wheat background was produced in this work using the gametocidal chromosome 2Cc from Ae. cylindrica. Gametocidal genes have been used to produce structural chromosome aberrations in wheat [36,37], barley [38,39] and rye [40]. In H. chilense, structural changes have been previously obtained for chromosomes 1Hch, 3Hch, 4Hch and 7Hch [8,11,17,18,41] and have been useful in determining the locations of genes and markers in this species. Breaks at both centromeric and interstitial regions of chromosomes have been induced by gametocidal chromosome 2Cc [36]. In the present study, telocentric and translocations between chromosome 2Hch from H. chilense and wheat chromosomes have been generated
Alien addition and translocation lines are an ideal template for PCR-based mapping to assign molecular markers to chromosomes of the wild relatives of wheat [8,11,41,42,43]. Using gene-based conserved orthologous set (COS) markers on wheat—H. chilense introgression lines obtained in this work, we assigned a total of 24 markers to H. chilense chromosome 2Hch. A 78.3% transference rate of COS markers to H. chilense chromosome 2Hch was found. Since COS markers were intended for comparative studies among grasses, the high rate of transferability obtained in this work was expected [21]. Similar rates of transference of COS markers to H. chilense chromosome 7Hch have been found previously [17]. COS markers have also been transferred successfully to other Triticeae species such as Agropyron cristatum [29,41] and Aegilops spp. [44].
The relevance of chromosome 2Hch for endosperm carotenoid content has been highlighted by previous work. Association studies for YPC allowed the identification of three main chromosome regions for YPC variation in H. chilense, with the largest one located on chromosome 2Hch and smaller regions detected on chromosomes 3Hch and 7Hch [20]. Four candidate genes associated with YPC were genetically mapped to chromosome 2Hch: both Ggpps1 and Zds were tightly linked and mapped near the centromere, while Hyd3 and Ppo1 were mapped to the long arm of chromosome 2Hch [20]. Furthermore, a significant QTL at the distal part of chromosome 2Hch has also been found where no carotenoid-related genes have been mapped [19,20]. The importance of chromosome 2HchL in grain carotenoid content has also been revealed in the new cereal tritordeum (amphiploid derived from a cross between the wild barley H. chilense and durum wheat), which has higher carotenoid pigment content in its grain than durum or bread wheat [45,46].
Chromosome 7Hch from H. chilense confers the capacity to accumulate higher carotene concentration in seeds [15]. The Psy1 gene controlling the first step of the carotenoid biosynthetic pathway was mapped to 7HchS [47]. Wheat—H. chilense chromosome 7Hchintrogression lines have been developed [17,18,48], and all the genetic stocks carrying Psy1 from H. chilense show increased carotenoid content relative to common wheat [48,49]. The obtention in this work of translocation T7HchS·2HchL could be of interest for studying the effect of the Psy1 gene located on the 7HchS arm and both the Hyd3 and Ppo1 genes mapped to the long arm of chromosome 2Hch [20]. Furthermore, the lines carrying T2HchS·2DL and T2HchS·2DL + T7HchS·D translocations could be of interest for studying the effect of both the Ggpps1 and Zds genes mapped to the short arm of 2Hch, in the absence and presence of the Psy1 gene from H. chilense, respectively.
The transference of desirable genes from wild relatives to wheat can be restrict by linkage drag and the lack of compensation for the wheat chromatin substituted. The identification of alien chromosomal regions carrying the genes of interest and the analysis of their homoeologous relationships with wheat chromosomes can overcome that difficulty. It has been pointed out that only translocations produced by homoeologous recombination are beneficial for wheat improvement [50,51]. In this work, wheat—H. chilense macrosyntenic comparisons using COS markers revealed that H. chilense chromosome 2Hch exhibits good synteny with wheat homoeologous group 2 chromosomes. Comparative mapping of carotenoid-related genes mapped to chromosome 2Hch also showed good collinearity between H. chilense and Triticeae species [20,46]. The 24 COS markers assigned to chromosome 2Hch in this work will facilitate introgression of alien genes associated with this chromosome into wheat.
5. Conclusions
We used in situ hybridization and genotyping to characterize genetic stocks harboring chromosome 2Hch from H. chilense in a wheat background. As far as we know, no previous translocation lines involving chromosome 2Hchintrogressed into wheat have been described. The cytogenetic stocks developed here may constitute an important resource for studying the effect of chromosome 2Hch on wheat grain color. In addition, these cytogenetic stocks allowed the localization of a set of conserved orthologous set (COS) markers to specific arms in chromosome 2Hch.The genomic position of orthologous unigene EST-contigs used for the COS marker design revealed a good macrosyntenic relationship between H. chilense chromosome 2Hch and wheat homoeologous group 2. The new wheat—H. chilense recombinations are useful for genetic studies and might also serve as a bridge for transferring genes associated with yellow pigment into a wheat background.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/9/9/493/s1, File S1: COS markers used in this work together with their primer sequences, File S2: Source of the COS markers assigned to the H. chilense chromosome 2Hch, File S3: Results of BLASTn search for COS markers assigned to H. chilense chromosome 2Hch in the reference sequences of hexaploid wheat group 2 chromosomes (www.wheatgenome.org/) and the start positions (bp) of the marker-specific ESTs.
Author Contributions
A.C. conceived and designed the study; C.P. performed the experiments; A.C. analysed the data and wrote the paper: all authors have read and approved the final manuscript.
Funding
This research was supported by grants AGL2014-53195-R and RTI2018-093367-B-I00 from the Spanish State Research Agency (Ministry of Science, Innovation and Universities), co-financed by the European Regional Development Fund (FEDER) from the European Union.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hajjar, R.; Hodgkin, T. The use of wild relatives in crop improvement: A survey of developments over the last 20 years. Euphytica 2007, 156, 13. [Google Scholar] [CrossRef]
- Prohems, J.; Gramazio, P.; Plazas, M.; Dempewil, H.; Kilian, B.; Díez, M.J.; Fita, A.; Herraiz, F.J.; Rodríguez-Burruezo, A.; Soler, S.; et al. Introgressiomics: A new approach for using wild relatives in breeding for adaptation to climate change. Euphytica 2017, 213, 158. [Google Scholar] [CrossRef]
- Martín, A.; Martín, L.M.; Cabrera, A.; Ramírez, M.C.; Giménez, M.J.; Rubiales, D.; Hernández, P.; Ballesteros, J. The potential of Hordeum chilense in breeding Triticeae species. In Triticeae III; Jaradat, A.A., Humphreys, M., Eds.; Science Publishers Inc.: Enfield, UK, 1998; pp. 377–386. [Google Scholar]
- Martín, A.; Cabrera, A.; Hernández, P.; Ramírez, M.C.; Rubiales, D.; Ballesteros, J. Prospect for the use of Hordeum chilense in durum wheat breeding. In Durum Wheat Improvement in the Mediterranean Region: New Challenges; Rojo, C., Nachit, M., Di-Fonzo, N., Araus, J., Eds.; Options Méditerranées: Zaragoza, Spain, 2000; pp. 111–115. [Google Scholar]
- Martín, A.; Cabrera, A. Cytogenetics of Hordeum chilense: Current status and considerations with reference to breeding. Cytogenet. Genome Res. 2005, 109, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.B.; Guzman, C. Interspecific and intergeneric hybridization as a source of variation for wheat grain quality improvement. Theor. Appl. Genet. 2018, 131, 225–251. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.E.; Reader, S.M.; Chapman, V. The addition of Hordeum chilense chromosomes to wheat. In Induced Variability in Plant Breeding, Proceedings of International Symposium in Eucarpia; Broertjes, C., Ed.; Pudoc: Wageningen, The Netherlands, 1982; pp. 79–81. [Google Scholar]
- Said, M.; Recio, R.; Cabrera, A. Development and characterisation of structural changes in chromosome 3Hch from Hordeum chilense in common wheat and their use in physical mapping. Euphytica 2012, 188, 429–440. [Google Scholar] [CrossRef]
- Castro, A.M.; Martin, A.; Martin, L.M. Location of genes controlling resistance to greenbug (Scizaphis graminum Rond.) in Hordeum chilense. Plant Breed 1996, 115, 335–338. [Google Scholar] [CrossRef]
- Payne, P.I.; Holt, L.M.; Reader, S.M.; Miller, T.E. Chromosomal location of genes coding for endosperm proteins of Hordeum chilense, determined by two-dimensional electrophoresis of wheat-H. chilense chromosome addition lines. Biochem. Genet. 1987, 25, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Cherif-Mouaki, S.; Said, M.; Alvarez, J.B.; Cabrera, A. Sub-arm location of prolamin and EST-SSR loci on chromosome 1Hch from Hordeum chilense. Euphytica 2011, 178, 63–69. [Google Scholar] [CrossRef]
- Rubiales, D.; Reader, S.M.; Martín, A. Chromosomal location of resistance of Septoria tritici in Hordeum chilense determined by the study of chromosomal addition and substitution lines in “Chinese Spring” wheat. Euphytica 2000, 115, 221–224. [Google Scholar] [CrossRef]
- Forster, B.P.; Phillips, M.S.; Miller, T.E.; Baird, E.; Powell, W. Chromosome location of genes controlling tolerance to salt (NaCl) and vigour in Hordeum vulgare and H. chilense. Heredity 1889, 65, 9–107. [Google Scholar] [CrossRef]
- Martin, A.C.; Atienza, S.G.; Ramírez, M.C.; Barro, F.; Martin, A. Male fertility restoration of wheat in Hordeum chilense cytoplasm is associated with 6HchS chromosome addition. Aust. J. Agric. Res. 2008, 59, 206–213. [Google Scholar] [CrossRef]
- Alvarez, J.B.; Martín, L.M.; Martín, A. Chromosomal localization of genes for carotenoid pigments using addition lines of Hordeum chilense in wheat. Plant Breed. 1998, 117, 287–289. [Google Scholar] [CrossRef]
- Calderón, M.D.C.; Ramírez, M.D.C.; Martín, A.; Prieto, P. Development of Hordeum chilense 4Hch introgression lines in durum wheat: A tool for breeders and complex trait analysis. Plant Breed. 2012, 131, 733–738. [Google Scholar] [CrossRef]
- Mattera, M.G.; Avila, M.C.; Atienza, S.G.; Cabrera, A. Cytological and molecular characterization of wheat-Hordeum chilense chromosome 7Hch introgression lines. Euphytica 2015, 203, 165–176. [Google Scholar] [CrossRef]
- Mattera, M.G.; Cabrera, A. Characterization of a set of common wheat-Hordeum chilense chromosome 7Hch introgression lines and its potential use in research on grain quality traits. Plant Breed. 2017, 136, 344–350. [Google Scholar] [CrossRef]
- Atienza, S.G.; Ramírez, C.M.; Hernández, P.; Martín, A. Chromosomal location of genes for carotenoid pigments in Hordeum chilense. Plant Breed. 2004, 123, 303–304. [Google Scholar] [CrossRef]
- Rodríguez-Suárez, C.; Atienza, S.G. Hordeum chilense genome, a useful tool to investigate the endosperm yellow pigment content in the Triticeae. BMC Plant Biol. 2012, 12, 200. [Google Scholar] [CrossRef]
- Quraishi, U.M.; Abrouk, M.; Bolot, S.; Pont, C.; Throude, M.; Guilhot, N.; Confolent, C.; Bortolini, F.; Praud, S.; Murigneux, A.; et al. Genomics in cereals: From genome-wide conserved orthologous set (COS) sequences to candidate genes for trait dissection. Func. Integr. Genom. 2009, 9, 473–484. [Google Scholar] [CrossRef]
- Cabrera, A.; Martín, A.; Barro, F. In-Situ Comparative Mapping (ISCM) of Glu-1 loci in Triticum and Hordeum. Chromosome Res. 2002, 10, 49–54. [Google Scholar] [CrossRef]
- Rayburn, A.L.; Gill, B.S. Molecular identification of the D-genome chromosomes of wheat. J. Hered. 1986, 77, 253–255. [Google Scholar] [CrossRef]
- Mukai, Y.; Nakahara, Y.; Yamamoto, M. Simultaneous discrimination of the three genomes in hexaploid wheat by multicolor fluorescence in situ hybridization using total genomic and highly repeated DNA probes. Genome 1993, 36, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, A.; Friebe, B.; Jiang, J.; Gill, B.S. Characterization of Hordeum chilense chromosomes by C-banding and in situ hybridization using highly repeated DNA probes. Genome 1995, 38, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acid Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Röder, M.S.; Korzun, V.; Wandehake, K.; Planschke, J.; Tixier, M.H.; Leroy, P.; Ganal, M.W. A microsatellite map of wheat. Genetics 1998, 149, 2007–2023. [Google Scholar] [PubMed]
- Sourdille, P.; Singh, S.; Cadalen, T.; Brown-Guedira, G.; Gay, G.; Qi, L.; Gill, B.S.; Dufour, P.; Murigneux, A.; Bernard, M. Microsatellite-based deletion bin system for the establishment of genetic-physical map relationships in wheat (Triticum aestivum L.). Func. Integr. Genom. 2004, 4, 12–25. [Google Scholar] [CrossRef] [PubMed]
- GrainGenes. Available online: http://wheat.pw.usda.gov/cgi-bin/graingenes/browse.cgi?class=marker (accessed on 1 December 2018).
- Said, M.; Copete, A.; Gaál, E.; Molnár, I.; Cabrera, A.; Doležel, J.; Vrána, J. Uncovering macrosyntenic relationships between tetraploid Agropyron cristatum and bread wheat genomes using COS markers. Theor. Appl. Genet. 2019. [Google Scholar] [CrossRef] [PubMed]
- International Wheat Genome Sequencing Consortium. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef]
- Mayer, K.F.X.; Martis, M.; Hedley, P.E.; Simková, H.; Liu, H.; Morris, J.A.; Steuernagel, B.; Taudien, S.; Roessner, S.; Gundlach, H.; et al. Unlocking the barley genome by chromosomal and comparative genomics. Plant Cell 2011, 23, 1249–1263. [Google Scholar] [CrossRef]
- RAP-DB The Rice Annotation Project Database. Available online: https://rapdb.dna.affrc.go.jp/tools/converter (accessed on 22 November 2018).
- Cantalapiedra, C.P.; Boudiar, R.; Casas, A.M.; Igartua, E.; Contreras-Moreira, B. Barleymap: Physical and genetic mapping of nucleotide sequences and annotation of surrounding loci in barley. Mol. Breed. 2014, 35, 13. [Google Scholar] [CrossRef]
- International Barley Genome Sequencing Consortium. A physical, genetic and functional sequence assembly of the barley genome. Nature 2012, 491, 711–716. [Google Scholar] [CrossRef]
- Endo, T.R. Induction of chromosomal structural changes by a chromosome of Aegilops cylindrica L. in common wheat. J. Hered. 1988, 79, 366–370. [Google Scholar] [CrossRef]
- Endo, T.R.; Gill, B.S. The deletion stocks of common wheat. J. Hered. 1996, 87, 295–307. [Google Scholar] [CrossRef]
- Endo, T.R. Cytological dissection of barley genome by the gametocidal system. Breed. Sci. 2009, 59, 481–486. [Google Scholar] [CrossRef] [Green Version]
- Nasuda, S.; Kikkawa, Y.; Ashida, T.; RafiqulIslam, A.K.M.; Sato, K.; Endo, R. Chromosomal assignment and deletion mapping of barley EST markers. Genes Genet. Syst. 2005, 80, 357–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friebe, B.; Kynast, R.G.; Gill, B.S. Gametocidal factor induced structural rearrangements in rye chromosomes added to common wheat. Chromosome Res. 2000, 8, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Said, M.; Cabrera, A. A physical map of chromosome 4Hch from Hordeum chilense containing SSR, STS and EST-SSR molecular markers. Euphytica 2009, 167, 253–259. [Google Scholar] [CrossRef]
- Ochoa, V.; Madrid, E.; Said, M.; Rubiales, D.; Cabrera, A. Molecular and cytogenetic characterization of a common wheat-Agropyron cristatum chromosome translocation conferring resistance to leaf rust. Euphytica 2015, 201, 89–95. [Google Scholar] [CrossRef]
- Copete, A.; Cabrera, A. Chromosomal location of genes for resistance to powdery mildew in Agropyron cristatum and mapping of Conserved Orthologous Set molecular markers. Euphytica 2017, 213, 189–297. [Google Scholar] [CrossRef]
- Molnár, I.; Vrána, J.; Burešová, V.; Cápal, P.; Farkas, A.; Cseh, A.; Kubaláková, M.; Molnár-Láng, M.; Doležel, J. Dissecting the U, M, S and C genomes of wild relatives of bread wheat (Aegilops spp.) into chromosomes and exploring their synteny with wheat. Plant J. 2016, 88, 452–467. [Google Scholar] [CrossRef]
- Atienza, S.G.; Ballesteros, J.; Martín, A.; Hornero-Méndez, D. Genetic variability of carotenoid concentration and degree of esterification among tritordeum (xTritordeumAscherson et Graebner) and durum wheat accessions. J. Agric. Food Chem. 2007, 55, 4244–4251. [Google Scholar] [CrossRef]
- Rodríguez-Suárez, C.; Atienza, S.G. Polyphenol oxidase genes in Hordeum chilense and implications in tritordeum breeding. Mol. Breed. 2014, 34, s11032–s11014. [Google Scholar] [CrossRef]
- Atienza, S.G.; Ávila, C.M.; Martín, A. The development of a PCR-based marker for PSY1 from Hordeum chilense, a candidate gene for carotenoid content accumulation in tritordeum seeds. Crop Pasture Sci. 2007, 58, 767–773. [Google Scholar] [CrossRef]
- Rey, M.D.; Calderón, M.D.C.; Rodrigo, M.J.; Zacarías, L.; Alós, E.; Prieto, P. Novel bread wheat lines enriched in carotenoids carrying Hordeum chilense chromosome arms in the ph1b background. PLoS ONE 2015, 10, e0134598. [Google Scholar] [CrossRef] [PubMed]
- Mattera, M.G.; Cabrera, A.; Hornero-Méndez, D.; Atienza, S.G. Lutein esterification in wheat endosperm is controlled by the homoeologous group 7 and is increased by the simultaneous presence of chromosomes 7D and 7Hch from Hordeum chilense. Crop. Pasture Sci. 2015, 66, 912–921. [Google Scholar] [CrossRef]
- Friebe, B.; Jiang, J.; Raupp, W.J.; McIntosch, R.A.; Gill, B.S. Characterization of wheat alien translocations conferring resistance to diseases and pests: Current status. Euphytica 2006, 91, 59–87. [Google Scholar] [CrossRef]
- Qi, L.; Friebe, B.; Zhang, P.; Gill, B.S. Homoeologous recombination, chromosome engineering and crop improvement. Chromosome Res. 2007, 15, 3–19. [Google Scholar] [CrossRef]
Figure 1.
Fluorescence in situ hybridization (FISH) with the pAs1 repetitive (red) and H. chilense genomic DNA (green) probes to mitotic metaphase of wheat—H. chilense introgression lines involving chromosome 2Hch. (a) Disomic substitution 2Hch (2D); (b) Ditelosomic 2HchL; (c) Isochromosome 2HchL; (d) Translocation 2HchS·2DL; (e) Translocation 2HchS·2DL + T7HchS·D; (f) Translocation 7HchS·2HchL; (g) Translocation 2HchL·7HchL. Bar = 10 µm.
Figure 1.
Fluorescence in situ hybridization (FISH) with the pAs1 repetitive (red) and H. chilense genomic DNA (green) probes to mitotic metaphase of wheat—H. chilense introgression lines involving chromosome 2Hch. (a) Disomic substitution 2Hch (2D); (b) Ditelosomic 2HchL; (c) Isochromosome 2HchL; (d) Translocation 2HchS·2DL; (e) Translocation 2HchS·2DL + T7HchS·D; (f) Translocation 7HchS·2HchL; (g) Translocation 2HchL·7HchL. Bar = 10 µm.

Figure 2.
Molecular characterization of introgression lines with wheat chromosome-specific simple sequence repeats (SSR) markers. (a) Xgwm261-2DS; (b) Xgwm157-2DL; (c) Xcfd66-7DS and (d) Xbarc111-7DL.
Figure 2.
Molecular characterization of introgression lines with wheat chromosome-specific simple sequence repeats (SSR) markers. (a) Xgwm261-2DS; (b) Xgwm157-2DL; (c) Xcfd66-7DS and (d) Xbarc111-7DL.

Figure 3.
Examples of PCR amplification profiles used for identifying chromosome 2Hch and 7Hch arms in the introgression lines. (a) c749557 mapped on the short arm of chromosome 2Hch; (b) c731690 mapped on the long arm of chromosome 2Hch; (c) c779791 mapped on the short arm of chromosome 7Hch; and (d) c759439 mapped on the long arm of chromosome 7Hch. “Chinese Spring” (CS), H. chilense (Hch), ditelo 2HchS, ditelo 7HchS, ditelo 7HchL and disomic substitution line CS 7Hch(7D) were used as controls.
Figure 3.
Examples of PCR amplification profiles used for identifying chromosome 2Hch and 7Hch arms in the introgression lines. (a) c749557 mapped on the short arm of chromosome 2Hch; (b) c731690 mapped on the long arm of chromosome 2Hch; (c) c779791 mapped on the short arm of chromosome 7Hch; and (d) c759439 mapped on the long arm of chromosome 7Hch. “Chinese Spring” (CS), H. chilense (Hch), ditelo 2HchS, ditelo 7HchS, ditelo 7HchL and disomic substitution line CS 7Hch(7D) were used as controls.

Figure 4.
Visualization of wheat-H. chilense orthologous regions from the perspective of wheat homoeologous chromosome group 2. Physical map of the source expressed sequence tags (ESTs) of the COS-markers (left), the genomic positions on wheat pseudomolecules (kb) are on the right.
Figure 4.
Visualization of wheat-H. chilense orthologous regions from the perspective of wheat homoeologous chromosome group 2. Physical map of the source expressed sequence tags (ESTs) of the COS-markers (left), the genomic positions on wheat pseudomolecules (kb) are on the right.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Chromosome constitutions of wheat—H. chilense introgression lines involving chromosome 2Hch.
Table 1.
Chromosome constitutions of wheat—H. chilense introgression lines involving chromosome 2Hch.
| Line | Type of Aberration | H. chilense Introgressions | No. of D Chromosomes | No. of A/B Chromosomes | Total No. of Chromosomes |
|---|---|---|---|---|---|
| Sus 2Hch (2D) | Substitution 1 | 2Hch | 12 (2D pair absent) | 28 | 42 |
| Ditelo 2HchL | Telosome 1 | 2HchL | 12 (2D pair absent) | 30 | 42 + 2 telos |
| Iso 2HchL | Isochromosome 2 | 2HchL | 12 (2D pair absent) | 30 | 42 + iso |
| T2HchS·2DL | Translocation 1 | 2HchS | 12 + 2T (2DL absent) | 30 | 42 + 2T |
| T2HchS·2DL + T7HchS·D | Translocation 3 | 2HchS+7HchS | 8 + 2T (7D pair absent) | 28 | 36 + 2T |
| T7HchS·2HchL | Translocation 1 | 2HchL+7HchS | 9 (2D pair absent) | 30 | 39 + 2T |
| T2HchL·7HchL | Translocation 2 | 2HchL+7HchL | 12 (2D pair absent) | 30 | 42 + 1T |
1 disomic; 2 monosomic; 3 double monosomic
Table 2.
Characterization and chromosome localization of wheat conserved orthologous set (COS) markers on H. chilense chromosome 2Hch.
Table 2.
Characterization and chromosome localization of wheat conserved orthologous set (COS) markers on H. chilense chromosome 2Hch.
| Marker | Product Size in T. aestivum | Product Size in H. chilense | Arm Location in H. chilense | Chromosome Location in Wheat 1 | Location in Wheat (cm) 1 | Location in Barley (cm) 2 |
|---|---|---|---|---|---|---|
| c723421 | 262–234 | 242 | 2HchS | 2BS | 28.1 | 63.5 |
| c754613 | 775 | 750 | 2HchS | 2AS-2BS-2DS | 31.7 | 46.3 |
| c745448 | 329 | 313–364 | 2HchS | 2BS | 32.1 | 52.5 |
| c77095 | 997–1075 | 1186 | 2HchS | 2AS-2BS-2DS | 35.9 | 47.7 |
| c741602 | 887 | 925 | 2HchS | 2AS-2BS | 45.8 | 56.3 |
| c751379 | 889 | 911 | 2HchS | 2AS-2BS | 56.3 | 59.2 |
| c733078 | 419 | 393 | 2HchS | 2BS-2DS | 68.2 | Not found |
| c756234 | 432 | 311 | - | 2AS-2BS | 63.9 | 52.2 |
| c749557 | 251–268 | 220 | 2HchS | 2AS-2DS | 69.0 | 59.2 |
| c740970 | 229 | 239 | 2HchL | 2AL-2BL-2DL | 140.2 | 63.5 |
| c744070 | 204 | 221 | 2HchL | 2AL-2BL-2DL | 143.7 | Not found |
| c729382 | 602 | 463 | 2HchL | 2BL-2DL | 145.5 | 69.2 |
| c751189 | 805 | 851 | 2HchL | 2AL-2BL-2DL | 145.8 | 70.54 |
| c749297 | 257 | 247 | 2HchL | 2AL-2BL-2DL | 149.3 | 71.1 |
| BF291656 | 920 | 821 | 2HchL | 2AL-2BL-2DL | 152.0 | 98.6 |
| c760814 | 554602 | 2HchL | 2DL | 167.4 | Not found | |
| c747571 | 330 | 370 | 2HchL | 2AL-2BL-2DL | 173.2 | 122.2 |
| c743473 | 252 | 276 | 2HchL | 2AL-2BL-2DL | 173.5 | 82.75 |
| c779794 | 521–602 | 661 | 2HchL | 2AL-2BL | 176.0 | 113.5 |
| c795564 | 573 | 629 | 2HchL | 2AL | 178.5 | 113.5 |
| c730704 | 377 | 393 | 2HchL | 2DL | 179.4 | 156.7 |
| c741642 | 366-390 | 377 | 2HchL | 2BL-2DL | 187.3 | 126.0 |
| c746642 | 843–889 | 864 | 2HchL | 2AL-2BL-2DL | 189.5 | 156.7 |
| c747195 | 508 | 668 | 2HchL | 2AL-2BL-2DL | 190.2 | Not found |
| c731690 | 342–355 | 320 | 2HchL | 2BL-2DL | 191.7 | 102.8 |
| c756123 | 398–520 | 442 | - | 2AL-2BL-2DL | 198.2 | 136.8 |
1 Quraishi et al. (2009); 2 Position in barley determined using Barleymap (http://floresta.eead.csic.es).
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Palomino, C.; Cabrera, A. Development of wheat—Hordeum chilense Chromosome 2Hch Introgression Lines Potentially Useful for Improving Grain Quality Traits. Agronomy 2019, 9, 493. https://doi.org/10.3390/agronomy9090493
AMA Style
Palomino C, Cabrera A. Development of wheat—Hordeum chilense Chromosome 2Hch Introgression Lines Potentially Useful for Improving Grain Quality Traits. Agronomy. 2019; 9(9):493. https://doi.org/10.3390/agronomy9090493
Chicago/Turabian StylePalomino, Carmen, and Adoracion Cabrera. 2019. "Development of wheat—Hordeum chilense Chromosome 2Hch Introgression Lines Potentially Useful for Improving Grain Quality Traits" Agronomy 9, no. 9: 493. https://doi.org/10.3390/agronomy9090493
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.