
Interplanting of Corn (Zea mays L.) Shifts Nitrogen Utilization by Promoting Rhizosphere Microbial Nitrogen Nitrification
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design and Crop Management
2.2. Plant Measurements
2.3. Soil Chemical Analysis
2.4. Macrogenomes of Rhizosphere Soil Microorganisms
2.5. Statistical Analyses
3. Results
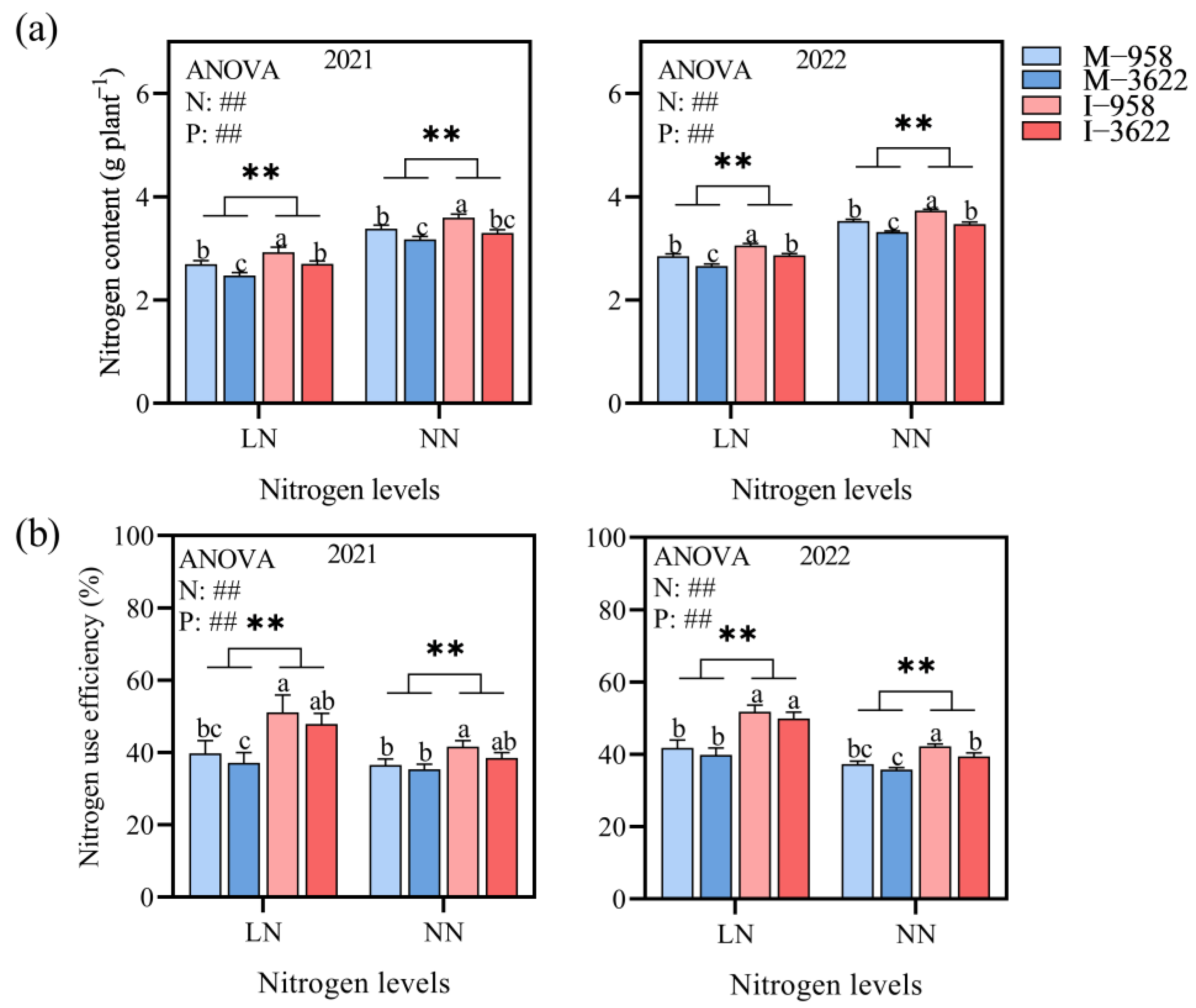
3.1. Nitrogen Utilization of Corn
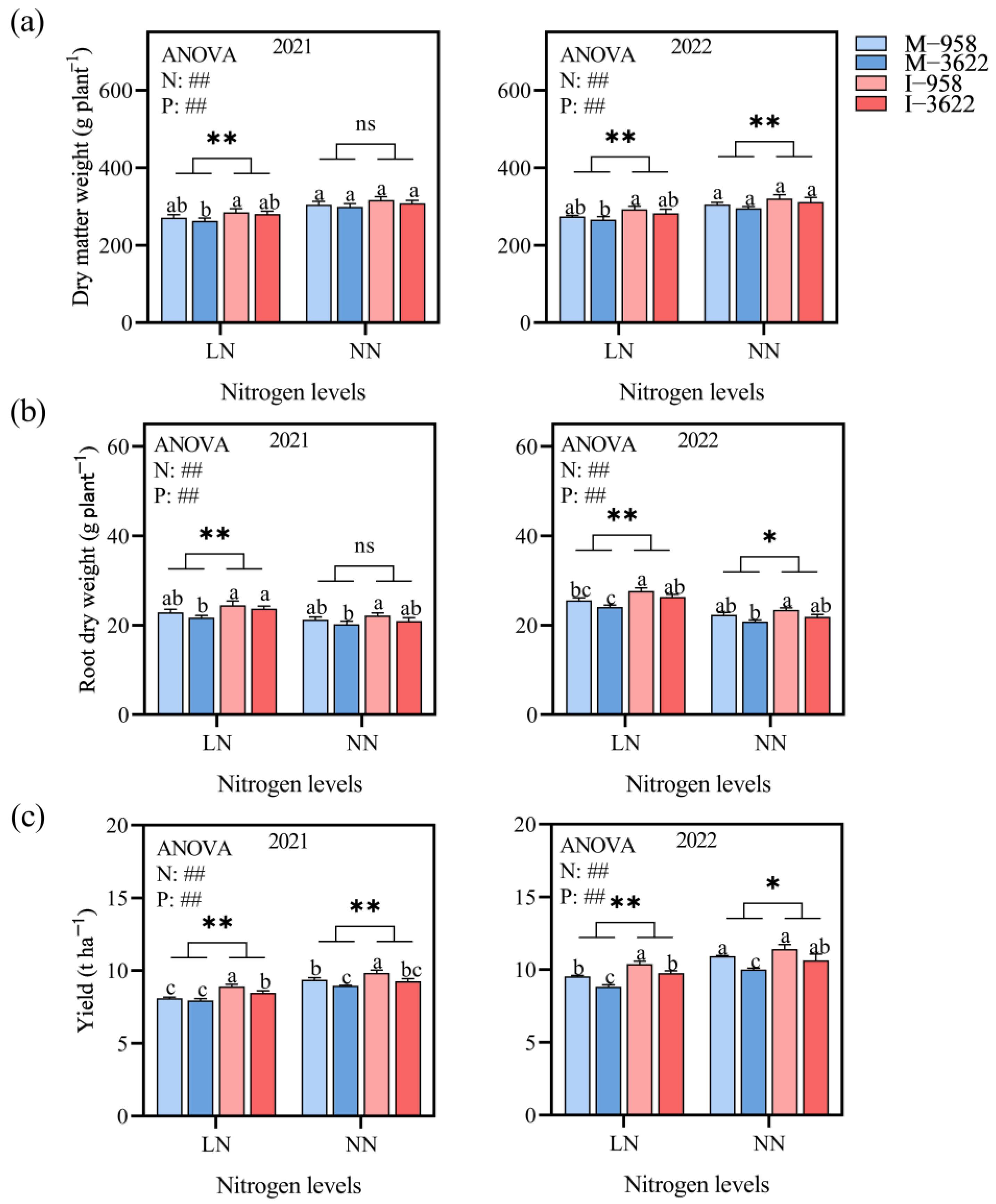
3.2. Crop Growth in Interplanting
3.3. Soil Nutrient and Net Potential Mineralization
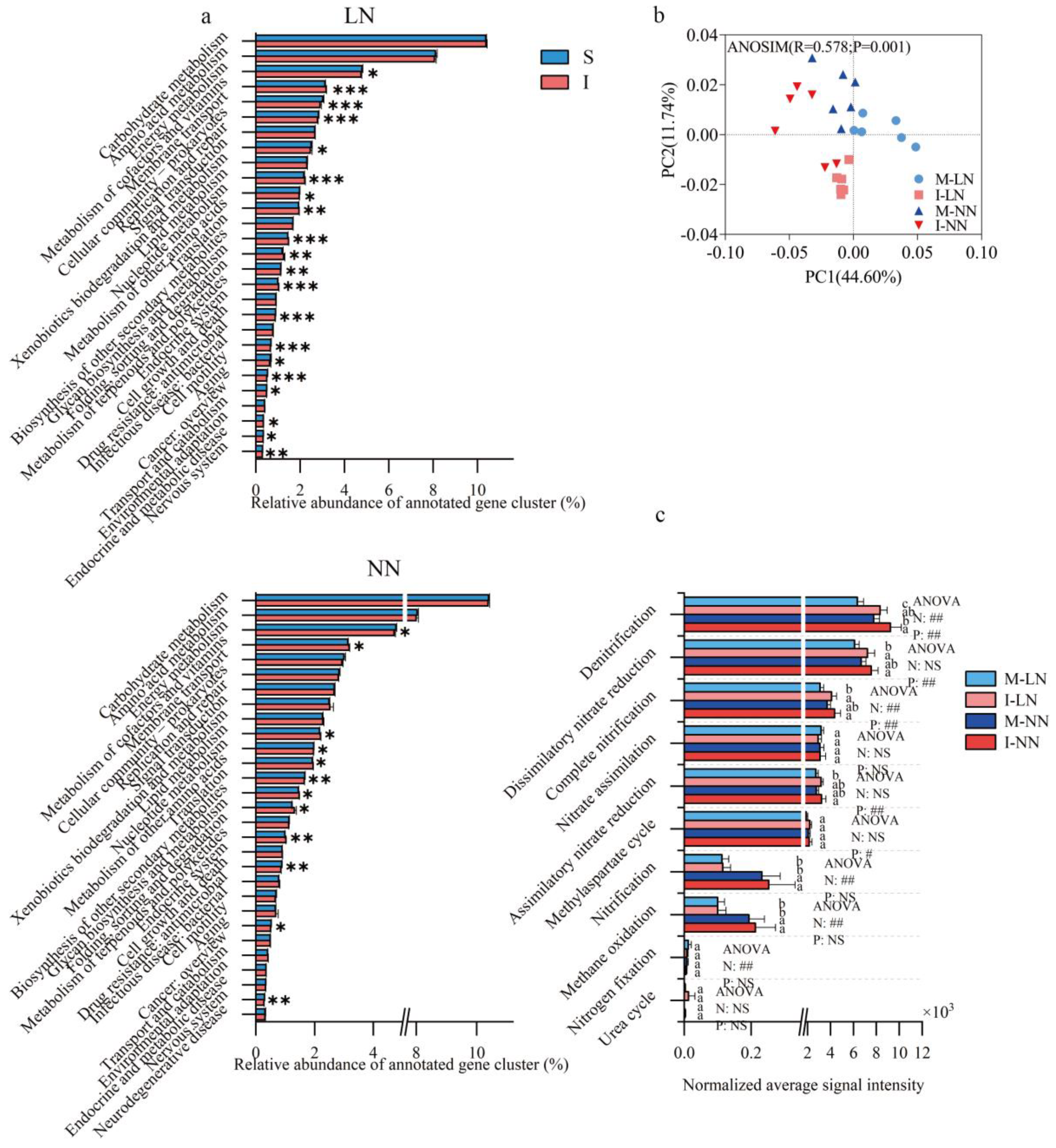
3.4. Soil Microbial Function of Nitrogen Cycle Pathway
3.5. Relative Abundance of Genes in Nitrogen Cycle Pathway
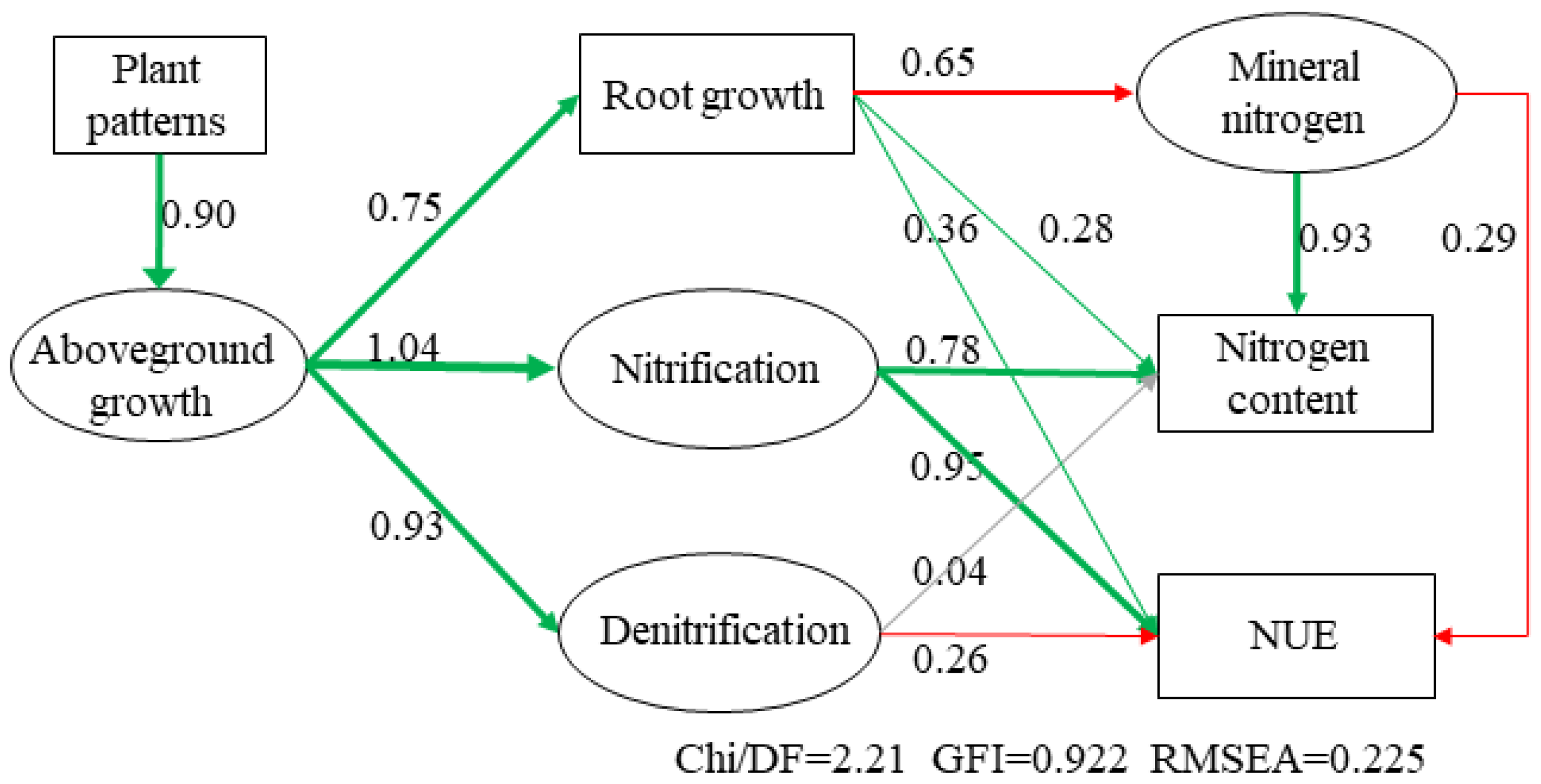
3.6. Relationships between Plant Nitrogen Content, Microbial Nitrogen Cycling, and NUE
4. Discussion
4.1. Interplanting Promoted Plant Growth and Nitrogen Utilization
4.2. Interplanting Enhanced Nitrogen Accumulation and Nitrogen Cycling Function in Rhizosphere
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- National Bureau of Statistics. China Statistical Yearbook—2022. 2022. Available online: http://www.stats.gov.cn/ (accessed on 6 September 2023).
- Malhi, S.S.; Lemke, R.; Wang, Z.H.; Chhabra, B.S. Tillage, nitrogen and crop residue effects on crop yield, nutrient uptake, soil quality, and greenhouse gas emissions. Soil Tillage Res. 2006, 90, 171–183. [Google Scholar] [CrossRef]
- Wang, S.; Huang, X.L.; Zhang, Y.; Yin, C.B.; Richel, A. The effect of corn straw return on corn production in Northeast China: An integrated regional evaluation with meta-analysis and system dynamics. Resour. Conserv. Recycl. 2021, 167, 105402. [Google Scholar] [CrossRef]
- Bai, Y.C.; Chang, Y.Y.; Hussain, M.; Lu, B.; Zhang, J.P.; Song, X.B.; Lei, X.S.; Pei, D. Soil chemical and microbiological properties are changed by long term chemical fertilizers that limit ecosystem functioning. Microorganisms 2020, 8, 694. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Zhang, W.S.; Chen, Y.X.; Zhang, W.; Liu, D.Y.; Gao, X.P.; Chen, X.P. Nitrogen leaching and grey water footprint affected by nitrogen fertilization rate in maize production: A case study of southwest China. J. Sci. Food Agric. 2021, 101, 6064–6073. [Google Scholar] [CrossRef]
- Zhou, J.; Guan, D.W.; Zhou, B.K.; Zhao, B.S.; Ma, M.C.; Qin, J.; Jiang, X.; Chen, S.F.; Cao, F.M.; Shen, D.L.; et al. Influence of 34-years of fertilization on bacterial communities in an intensively cultivated black soil in northeast China. Soil Biol. Biochem. 2015, 90, 42–51. [Google Scholar] [CrossRef]
- Hu, X.J.; Liu, J.J.; Wei, D.; Zhu, P.; Cui, X.A.; Zhou, B.K.; Chen, X.L.; Jin, J.; Liu, X.B.; Wang, G.H. Effects of over 30-year of different fertilization regimes on fungal community compositions in the black soils of northeast China. Agric. Ecosyst. Environ. 2017, 248, 113–122. [Google Scholar] [CrossRef]
- Chen, L.; Li, K.K.; Shi, W.J.; Wang, X.L.; Wang, E.T.; Liu, J.F.; Sui, X.H.; Mi, G.H.; Tian, C.F.; Chen, W.X. Negative impacts of excessive nitrogen fertilization on the abundance and diversity of diazotrophs in black soil under maize monocropping. Geoderma 2021, 393, 114999. [Google Scholar] [CrossRef]
- Treseder, K.K. Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef]
- Zhong, Y.Q.W.; Yan, W.M.; Shangguan, Z.P. Impact of long-term N additions upon coupling between soil microbial community structure and activity, and nutrient-use efficiencies. Soil Biol. Biochem. 2015, 91, 151–159. [Google Scholar] [CrossRef]
- Martin, F.S.; Noordwijk, M.V. Is native timber tree intercropping an economically feasible alternative for smallholder farmers in the philippines? Aust. J. Agric. Resour. Econ. 2011, 55, 257–272. [Google Scholar] [CrossRef]
- Beedy, T.L.; Snapp, S.S.; Akinnifesi, F.K.; Sileshi, G.W. Impact of Gliricidia sepium intercropping on soil organic matter fractions in a maize-based cropping system. Agric. Ecosyst. Environ. 2010, 138, 139–146. [Google Scholar] [CrossRef]
- Dang, K.; Gong, X.W.; Zhao, G.; Wang, H.L.; Ivanistau, A.; Feng, B.L. Intercropping alters the soil microbial diversity and community to facilitate nitrogen assimilation: A potential mechanism for increasing proso millet grain yield. Front. Microbiol. 2020, 11, 601054. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, S.; Zhang, X.; Mao, Q.G.; Li, X.Z.; You, Y.M.; Wang, J.X.; Zheng, M.H.; Zhang, W.; Lu, X.K.; et al. Nitrogen addition reduces soil bacterial richness, while phosphorus addition alters community composition in an old-growth N-rich tropical forest in southern China. Soil Biol. Biochem. 2018, 127, 22–30. [Google Scholar] [CrossRef]
- Yu, L.L.; Luo, S.S.; Gou, Y.G.; Xu, X.; Wang, J.W. Structure of rhizospheric microbial community and N cycling functional gene shifts with reduced N input in sugarcane-soybean intercropping in South China. Agric. Ecosyst. Environ. 2021, 314, 107413. [Google Scholar] [CrossRef]
- Cuartero, J.; Pascual, J.A.; Vivo, J.M.; Özbolat, O.; Sánchez-Navarro, V.; Egea-Cortines, M.; Zornoza, R.; Mena, M.M.; Garcia, E.; Ros, M. A first-year melon/cowpea intercropping system improves soil nutrients and changes the soil microbial community. Agric. Ecosyst. Environ. 2022, 328, 107856. [Google Scholar] [CrossRef]
- Bargaz, A.; Noyce, G.L.; Fulthorpe, R.; Carlsson, G.; Furze, J.R.; Jensen, E.S.; Dhiba, D.; Isaac, M.E. Species interactions enhance root allocation, microbial diversity and P acquisition in intercropped wheat and soybean under P deficiency. Appl. Soil Ecol. 2017, 120, 179–188. [Google Scholar] [CrossRef]
- Nyawade, S.O.; Karanja, N.N.; Gachene, C.K.K.; Gitari, H.I.; Schulte-Geldermann, E.; Parker, M.L. Short-term dynamics of soil organic matter fractions and microbial activity in smallholder potato-legume intercropping systems. Appl. Soil Ecol. 2019, 142, 123–135. [Google Scholar] [CrossRef]
- Yang, Y.D.; Feng, X.M.; Hu, Y.G.; Zeng, Z.H. The diazotrophic community in oat rhizosphere: Effects of legume intercropping and crop growth stage. Front. Agric. Sci. Eng. 2019, 6, 162–171. [Google Scholar] [CrossRef]
- Zhao, F.Y.; Sun, Z.X.; Feng, L.S.; Zhang, Y.Y.; Feng, C.; Bai, W.; Zheng, J.M.; Zhang, Z.; Yang, N.; Cai, Q.; et al. Biological N fixation but not mineral N fertilization enhances the accumulation of N in peanut soil in maize/peanut intercropping system. J. Agric. Food Res. 2022, 10, 100365. [Google Scholar] [CrossRef]
- Yang, L.; Luo, Y.; Lu, B.L.; Zhou, G.P.; Chang, D.N.; Gao, S.J.; Zhang, J.D.; Che, Z.X.; Cao, W.D. Long-term maize and pea intercropping improved subsoil carbon storage while reduced greenhouse gas emissions. Agric. Ecosyst. Environ. 2023, 349, 108444. [Google Scholar] [CrossRef]
- Chen, W.Z.; Kang, Z.M.; Yang, Y.N.; Li, Y.S.; Qiu, R.L.; Qin, J.H.; Li, H.S. Interplanting of rice cultivars with high and low Cd accumulation can achieve the goal of “repairing while producing” in Cd-contaminated soil. Sci. Total Environ. 2022, 851, 158229. [Google Scholar] [CrossRef]
- Reiss, E.R.; Drinkwater, L.E. Cultivar mixtures: A meta-analysis of the effect of intraspecific diversity on crop yield. Ecol. Appl. 2018, 28, 62–77. [Google Scholar] [CrossRef]
- Li, C.H.; Su, X.H.; Sun, D.L. Ecological and physiological effects of intercropping compound populations with different genotypes of maize. Acta Ecol. Sin. 2002, 22, 2096–2103. Available online: https://www.ecologica.cn/stxb/article/abstract/021212 (accessed on 19 September 2023). (In Chinese).
- Duan, P.F.; Zhao, D. Study on ecological effects of intercropping different genotypes of maize on disease resistance and stable yield. J. Maize Sci. 2017, 25, 107–112. Available online: http://www.ymkx.com.cn/jms/article/abstract/20170617 (accessed on 23 September 2023). (In Chinese).
- Zhu, Y.Y.; Chen, H.; Fan, J.H.; Wang, Y.Y.; Li, Y.; Chen, J.B.; Fan, J.X.; Yang, S.S.; Hu, L.P.; Leung, H.; et al. Genetic diversity and disease control in rice. Nature 2000, 406, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Grettenberger, I.M.; Tooker, J.F. Moving beyond resistance management toward an expanded role for seed mixtures in agriculture. Agric. Ecosyst. Environ. 2015, 208, 29–36. [Google Scholar] [CrossRef]
- Ma, J.F.; Ma, Y.; Wei, X.T.; Feng, L.S.; Chen, H.; Hong, D.F.; Tang, Z.H.; Zhang, X.S. Quality performance of two genotypic maize intercropping populations after wind disaster lodging. Shandong Agric. Sci. 2015, 47, 36–38. (In Chinese) [Google Scholar] [CrossRef]
- Li, H.P.; Liu, K.; Li, Z.B.; Zhang, M.B.; Zhang, Y.E.; Li, S.Y.; Wang, X.L.; Zhou, J.L.; Zhao, Y.L.; Liu, T.X.; et al. Mixing trait-based corn (Zea mays L.) cultivars increases yield through pollination synchronization and increased cross-fertilization. Crop J. 2023, 11, 291–300. [Google Scholar] [CrossRef]
- Tao, J.J.; Wang, H.B.; Zhu, Z.Y.; Tan, J.F.; Wang, Y.L. Effect of Different genotype summer maize intercropping on yield and nitrogen absorption and utilization. Acta Agric. Boreali-Sin. 2016, 31, 185–191. (In Chinese). Available online: http://www.hbnxb.net/CN/10.7668/hbnxb.2016.06.029 (accessed on 15 September 2023).
- Liu, T.X.; Li, C.H.; Ma, X.M.; Zhao, X.; Liu, S.Y. Effects of intercropping different genotypes of maize on leaf senescence, grain yield, and quality. Chin. J. Plant Ecol. 2008, 32, 914–921. (In Chinese) [Google Scholar] [CrossRef]
- Li, X.L.; Guo, L.G.; Zhou, B.Y.; Tang, X.M.; Chen, C.C.; Zhang, L.; Zhang, S.Y.; Li, C.F.; Xiao, K.; Dong, W.X.; et al. Characterization of low-N responses in maize (Zea mays L.) cultivars with contrasting nitrogen use efficiency in the north China plain. J. Integr. Agric. 2019, 18, 2141–2152. [Google Scholar] [CrossRef]
- Gao, J.; Lei, M.; Yang, L.J.; Wang, P.; Tao, H.B.; Huang, S.B. Reduced row spacing improved yield by optimizing root distribution in maize. Eur. J. Agron. 2021, 127, 126291. [Google Scholar] [CrossRef]
- Roley, S.S.; Duncan, D.S.; Liang, D.; Garoutte, A.; Jackson, R.D.; Tiedje, J.M.; Robertson, G.P. Associative nitrogen fixation (ANF) in switchgrass (Panicum virgatum) across a nitrogen input gradient. PLoS ONE 2018, 13, e0197320. [Google Scholar] [CrossRef] [PubMed]
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Principal coordinate analysis and non-metric multidimensional scaling. In Analysing Ecological Data; Springer: New York, NY, USA, 2007; pp. 259–264. [Google Scholar]
- Field, A. Discovering Statistics Using IBM SPSS Statistic; Sage Publications Limited: London, UK, 2024. [Google Scholar]
- OriginPro, Version 2023; OriginLab Corporation: Northampton, MA, USA. Available online: https://www.originlab.com (accessed on 10 September 2023).
- Li, C.J.; Hoffland, E.; Kuyper, T.W.; Yu, Y.; Zhang, C.C.; Li, H.G.; Zhang, F.S.; Werf, V.D.W. Syndromes of production in intercropping impact yield gains. Nat. Plants 2020, 6, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Wang, L.; Sun, X.; Wang, X.C.; Pu, T.; Yang, H.; Rengel, Z.; Liu, W.G.; Yang, W.Y. Improved post-silking light interception increases yield and P-use efficiency of maize in maize/soybean relay strip intercropping. Field Crops Res. 2021, 262, 108054. [Google Scholar] [CrossRef]
- Shi, Z.S.; Zhu, M.; Li, F.H.; Wang, Z.B. Research on yield-increasing of different kinds of maize. J. Maize Sci. 2008, 16, 107–109. (In Chinese). Available online: http://www.ymkx.com.cn/jms/article/abstract/08020107 (accessed on 21 September 2023).
- Hou, S.; Ren, H.; Fan, F.L.; Zhao, M.; Zhou, W.B.; Zhou, B.Y.; Li, C.F. The effects of plant density and nitrogen fertilization on maize yield and soil microbial communities in the black soil region of Northeast China. Geoderma 2023, 430, 116325. [Google Scholar] [CrossRef]
- Craine, J.M. Competition for Nutrients and Optimal Root Allocation. Plant Soil 2006, 285, 171–185. [Google Scholar] [CrossRef]
- Chen, Y.B.; Dong, P.F.; Jia, X.C.; Miao, Z.Y.; Wang, J.J.; Wang, Q.; Yang, Q.; Zhao, W.L. Effects of intercropping on root distribution, nutrient accumulation and yield of maize with different root architecture. J. Nucl. Agric. 2023, 37, 594–605. (In Chinese) [Google Scholar]
- Huang, Y.; Wu, Q.; Deng, M.Y.; He, X.Q.; Jin, Y.; Wang, J.; Luo, K.; Xie, X.; Wang, T.; Yong, T.W. Effects of root interaction on root growth, leaf photosynthetic characteristics and biomass of maize under two intercropping systems of maize and legumes. J. Sichuan Agric. Univ. 2020, 38, 513–519. (In Chinese) [Google Scholar] [CrossRef]
- Yu, R.P.; Yang, H.; Xing, Y.; Zhang, W.P.; Lambers, H.; Li, L. Belowground processes and sustainability in agroecosystems with intercropping. Plant Soil 2022, 476, 263–288. [Google Scholar] [CrossRef]
- Wang, G.H.; Sheng, L.C.; Zhao, D.; Sheng, J.D.; Wang, X.R.; Liao, H. Allocation of nitrogen and carbon is regulated by nodulation and mycorrhizal networks in soybean/maize intercropping system. Front. Plant Sci. 2016, 7, 1901. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, T.R.; Vadez, V. Physiological traits for crop yield improvement in low N and P environments. Food Secur. Nutr.-Stress. Environ. Exploit. Plants’ Genet. Capab. 2002, 245, 9–23. [Google Scholar] [CrossRef]
- Yu, L.; Tang, Y.; Wang, Z.; Gou, Y.; Wang, J. Nitrogen-cycling genes and rhizosphere microbial community with reduced nitrogen application in maize/soybean strip intercropping. Nutr. Cycl. Agroecosystems 2019, 113, 35–49. [Google Scholar] [CrossRef]
- Wang, X.; Gao, Y.; Zhang, H.; Shao, Z.; Sun, B.; Gao, Q. Enhancement of rhizosphere citric acid and decrease of NO3−/NH4+ ratio by root interactions facilitate N fixation and transfer. Plant Soil 2020, 447, 169–182. [Google Scholar] [CrossRef]
- Tang, Y.Q.; Zhang, X.Y.; Li, D.D.; Wang, H.M.; Chen, F.S.; Fu, X.L.; Fang, X.M.; Sun, X.M.; Yu, G.R. Impacts of nitrogen and phosphorus additions on the abundance and community structure of ammonia oxidizers and denitrifying bacteria in Chinese fir plantations. Soil Biol. Biochem. 2016, 103, 284–293. [Google Scholar] [CrossRef]
- Shi, Y.L.; Liu, X.R.; Zhang, Q.W. Effects of combined biochar and organic fertilizer on nitrous oxide fluxes and the related nitrifier and denitrifier communities in a saline-alkali soil. Sci. Total Environ. 2019, 686, 199–211. [Google Scholar] [CrossRef]
- Kumar, V.V. Plant Growth-Promoting Microorganisms: Interaction with Plants and Soil; Springer International Publishing: Cham, Switzerland, 2016; Volume 1, pp. 1–16. [Google Scholar] [CrossRef]
- Liu, J.J.; Yu, Z.H.; Yao, Q.; Sui, Y.Y.; Shi, Y.; Chu, H.Y.; Tang, C.X.; Franks, A.E.; Jin, J.; Liu, X.B.; et al. Ammonia-oxidizing archaea show more distinct biogeographic distribution patterns than ammonia-oxidizing bacteria across the black soil zone of northeast China. Front. Microbiol. 2018, 9, 171. [Google Scholar] [CrossRef]
- Ouyang, Y.; Evans, S.E.; Friesen, M.L.; Tiemann, L.K. Effect of nitrogen fertilization on the abundance of nitrogen cycling genes in agricultural soils: A meta-analysis of field studies. Soil Biol. Biochem. 2018, 127, 71–78. [Google Scholar] [CrossRef]
- Sun, R.B.; Wang, F.H.; Hu, C.S.; Liu, B.B. Metagenomics reveals taxon-specific responses of the nitrogen-cycling microbial community to long-term nitrogen fertilization. Soil Biol. Biochem. 2021, 156, 108214. [Google Scholar] [CrossRef]
- Li, B.; Li, Y.Y.; Wu, H.M.; Zhang, F.F.; Li, C.J.; Li, X.X.; Lambers, H.; Li, L. Root exudates drive interspecific facilitation by enhancing nodulation and N2 fixation. Proc. Nat. Acad. Sci. USA 2016, 113, 6496–6501. [Google Scholar] [CrossRef] [PubMed]
- Brooker, R.W.; Bennett, A.E.; Cong, W.; Daniell, T.J.; George, T.S.; Hallett, P.D.; Hawes, C.; Iannetta, P.M.; Jones, H.G.; Karley, A.J.; et al. Improving intercropping: A synthesis of research in agronomy, plant physiology and ecology. New Phytol. 2015, 206, 107–117. [Google Scholar] [CrossRef]
- Tang, Y.L.; Yu, L.L.; Guan, A.M.; Zhou, X.Y.; Wang, Z.G.; Guo, Y.G.; Wang, J.W. Soil mineral nitrogen residual and yield-scaled soil N2O emissions lowered by reducing nitrogen application and intercropping with soybean for sweet maize production in southern China. J. Integr. Agric. 2017, 16, 2586–2596. [Google Scholar] [CrossRef]
- Pan, H.; Qin, Y.; Wang, Y.; Liu, S.; Yu, B.; Song, Y.; Wang, X.; Zhu, G. Dissimilatory nitrate/nitrite reduction to ammonium (DNRA) pathway dominates nitrate reduction processes in rhizosphere and non-rhizosphere of four fertilized farmland soil. Environ. Res. 2020, 186, 109612. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nitrogen | Planting | SMC | NH4+ | NO3− | AP | AK | TN | TP | TK |
|---|---|---|---|---|---|---|---|---|---|
| Level | Pattern | (%) | (mg·kg−1) | (mg·kg−1) | (mg·kg−1) | (mg·kg−1) | (g·kg−1) | (g·kg−1) | (g·kg−1) |
| LN | M | 21.25 ± 0.11 b | 5.75 ± 0.08 a | 9.4 ± 0.08 a | 26.1 ± 0.48 a | 196.37 ± 2.76 a | 1.29 ± 0.01 a | 0.76 ± 0.04 a | 4.41 ± 0.19 a |
| I | 22.25 ± 0.50 a | 5.53 ± 0.08 b | 8.94 ± 0.21 b | 25.73 ± 0.39 a | 190.67 ± 2.54 a | 1.32 ± 0.04 a | 0.74 ± 0.03 a | 4.22 ± 0.11 a | |
| NN | M | 19.67 ± 0.33 b | 8.41 ± 0.08 a | 17.44 ± 0.39 a | 24.86 ± 0.6 a | 191.88 ± 3.57 a | 1.34 ± 0.01 a | 0.73 ± 0.02 a | 4.4 ± 0.12 a |
| I | 23.13 ± 0.24 a | 8.28 ± 0.09 a | 15.4 ± 0.24 b | 24.71 ± 0.48 a | 189.31 ± 3.06 a | 1.34 ± 0.01 a | 0.71 ± 0.03 a | 4.37 ± 0.13 a | |
| ANOVA | |||||||||
| N | ns | ** | ** | ** | ns | * | ns | ns | |
| P | ** | ** | ** | ns | ns | ns | ns | ns | |
| N × P | ** | ns | ** | ns | ns | ns | ns | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, Z.; Shang, H.; Lin, M.; Song, R.; He, J.; Li, X.; Sun, L.; Li, X.; Guo, H.; Li, Y.; et al. Interplanting of Corn (Zea mays L.) Shifts Nitrogen Utilization by Promoting Rhizosphere Microbial Nitrogen Nitrification. Agronomy 2024, 14, 586. https://doi.org/10.3390/agronomy14030586
Miao Z, Shang H, Lin M, Song R, He J, Li X, Sun L, Li X, Guo H, Li Y, et al. Interplanting of Corn (Zea mays L.) Shifts Nitrogen Utilization by Promoting Rhizosphere Microbial Nitrogen Nitrification. Agronomy. 2024; 14(3):586. https://doi.org/10.3390/agronomy14030586
Chicago/Turabian StyleMiao, Zhengyan, Haipeng Shang, Mengjie Lin, Rui Song, Jiashuai He, Xinmei Li, Leikang Sun, Xiaoyong Li, Hangzhao Guo, Yuxia Li, and et al. 2024. "Interplanting of Corn (Zea mays L.) Shifts Nitrogen Utilization by Promoting Rhizosphere Microbial Nitrogen Nitrification" Agronomy 14, no. 3: 586. https://doi.org/10.3390/agronomy14030586