
Origanum dictamnus Essential Oil in Vapour or Aqueous Solution Application for Pepper Fruit Preservation against Botrytis cinerea

Department of Agricultural Sciences, Biotechnology and Food Science, Cyprus University of Technology, Limassol 3036, Cyprus
Agronomy 2024, 14(2), 257; https://doi.org/10.3390/agronomy14020257
Submission received: 20 December 2023
/
Revised: 5 January 2024
/
Accepted: 23 January 2024
/
Published: 25 January 2024
(This article belongs to the Special Issue It Runs in the Family: The Importance of the Lamiaceae Family Species)
Abstract
:The use of synthetic sanitizers for fresh commodities preservation is of concern, with eco-friendly alternatives, including essential oils (EOs), attracting research and industry interest. Dittany (Origanum dictamnus—DIT) oil was applied, either through vapour or dipping, on pepper fruit or in vitro against Botrytis cinerea, and compared to untreated (control) or chlorine (CHL)-treated fruits stored at 11 °C. Direct DIT vapours (up to 6 d) suppressed lesion growth, spore germination, and spore production compared to the untreated fruits. The antimicrobial properties of EOs were evidenced in pre-exposed-to-DIT oil vapours (residual effect), resulting in fruit lesion suppression. However, DIT-pretreated fruits had the same spore production and spore germination as the control and CHL applications. In in vitro tests, DIT vapours decreased colony growth and spore production when fungi were grown on Potato Dextrose Agar (PDA) or PDA pre-exposed to DIT following B. cinerea inoculation. This evidenced that the disease suppression after DIT vapour application primarily affected the interaction of the fruit–pathogen and/or residual responses on fruit tissue and/or PDA media. Fungal biomass in Potato Dextrose Broth (PDB) was evaluated after DIT and CHL applications (10–50–100–500–5000 μL L−1) and decreased with the DIT oil. Additionally, DIT or CHL sanitary dipping on pepper was ineffective against B. cinerea compared to vapour application. Therefore, DIT vapours revealed antimicrobial properties and could be an alternative postharvest sanitiser. DIT oil application should also be evaluated at semi-commercial scale for further optimizations, prior to commercialisation.
1. Introduction
Fresh produce preservation is still challenging and attracting the attention of researchers and application strategies on the postharvest community. The need for better fresh commodities’ commercial storage/transit conditions has increased recently due to the financial costs associated with spoiling food, consumer worries regarding the safety of foods containing synthetic substances, and an increase in fresh produce consumption [1]. The challenge is to control the spread of postharvest diseases and preserve fruit quality and safety. The most often utilized sanitizers for postharvest washing are chlorine and peroxyacetic acid [2], because of their well-known antimicrobial properties and simplicity of use [3,4]. In fact, commonly, fresh produce is preserved with chemical treatments, such as disinfectants containing chlorine or bromine [2,5]. However, existing sanitation means have given rise to significant health and environmental issues because of the possible cancer-causing residues from pesticide inputs and their lack of effectiveness on a variety of microorganisms [6,7]. Innovative preservation solutions, both environmentally and consumer-safe, are therefore, required.
Natural components have received great attention and application in the food industry [6,8,9]. The majority of natural products are easily accessible, safe for the environment, less likely to cause pest resistance, less dangerous for non-target organisms, less detrimental to plant growth, and, best of all, sometimes, less costly than certain chemical treatments [10,11,12]. Essential oils (EOs), hydrosol (Hyd; enriched water after the distillation process), and extracts from aromatic plants that have antimicrobial properties have garnered significant attention lately [2,13,14,15]. These products are thought to be safe for humans and the environment [16]. They can be used as agrochemicals [11] by controlling pathogens and/or microorganisms that produce toxins in food [17] but also observing stimulatory effects on crops [18]. Both EOs and their constituents reveal efficiency against insects [19,20], nematodes, fungi, and bacteria [15,21,22,23].
One advantage of EOs is their bioactivity as a volatile mean, which could make them effective fumigants for the preservation of commodities during storage [1,15,24]. Many research studies have documented the beneficial responses of EOs on the preservation of fresh produce, including strawberry (Fragaria vesca L.), tomato (Solanum lycopersicum L.), cucumber (Cucumis sativus L.), apricot (Prunus armeniaca L.), pear (Pyrus communis L.), apples (Malus domestica L.), and avocado (Persea americana L.) fruit, to name a few [1,9,11,25,26,27,28,29]. Essential oil and/or hydrosol might trigger a defence response, by increasing defence-related enzymes and total phenolics of the fresh commodities [9,15]. The consumer preference for natural plant components, such as EOs, over synthetic fungicides may contribute to the high acceptance of these novel treatments [2].
Origanum dictamnus (L.), commonly well known as dittany (DIT), is an indigenous plant species found in Crete, Greece, and well known for its antimicrobial [30,31], antioxidant [6,32], antiproliferative [6], aphrodisiac, oxytocic, antirheumatic, and healing properties [6,33]. Dittany properties are basically attributed to the main EO components, the phenol carvacrol [6]. Carvacrol is capable of suppressing the growth of bacteria and to inhibit the flux toxin production by Bacillus cereus [17] and control spoilage yeast in foodstuffs [23].
Pepper (Capsicum annuum) crop is widely cultivated, and fruit may be consumed raw or cooked [34,35]. Since it contains ingredients, including capsaicin, phenolic molecules, and vitamins A and C, it has also been linked to a lower risk of cancer and/or other disorders. Peppers are susceptible to low storage temperatures, i.e., less than 7 °C, and to opportunistic microbes at the preharvest and postharvest stages, causing significant fruit deterioration and alterations in secondary metabolism because of physiologic modifications, which promote pathogen growth. These are the main causes of pepper postharvest loss [36]. The encapsulation of chitosan containing Heracleum persicum fruit EO extended sweet bell pepper fruit shelf life due to the antioxidant capacity of the investigated oil [35].
Among the most significant fruit disorders globally, grey mould (Botrytis cinerea Pers.: Fr; teleomorph: Botryotinia fuckeliana) is a widespread and prevalent rot that affects a variety of vegetables [31,37]. The scope of this study was to (1) explore the impact of dittany EO towards the rotting of pepper fruit caused by B. cinerea, as opposed to using chlorine, a widely used sanitizer, (2) explore any residual effect of the EO in pre-treated fruits towards fungi development, and (3) evaluate the EO application method on pepper fruit vapours versus dipping.
2. Materials and Methods
2.1. Plant Material and Microorganism
Pepper (C. annuum cv. Bounty F1) fruits were purchased from a commercial greenhouse in Limassol, Cyprus. Plants were grown in soil, under vertical scheme for a period of 6 months, under 2-branch pruning scheme, with common cultivation practices applied. Fruits were selected to have uniform size, colour, ripeness (“immature-green” stage), and checked for no defects or injuries. Essential oils of dittany (Origanum dictamnus) were employed, while chlorine (CHL; 5% NaOCl) was utilized as a commercial chemical sanitizer.
Essential oils were obtained from dittany plants grown in nature, harvested in a hilly area of Crete-Chania, dried in air-ventilated oven at 35 °C, and then EOs were hydrodistilled by using a Clevenger apparatus for 2.5 h [38]. The composition of EO was evaluated using Gas Chromatography–Mass Spectrometry (GC-MS) according to Xylia et al. [14]. Essential oil components were identified through the comparison of EO’s retention time indices and mass spectra with authentic standards when assessable and based on the comparison of their mass spectra with spectral data of Mass Spectra Library NIST08 and published sources [39].
B. cinerea isolated from tomato (strain number: 169558) was supplied by CABI (CABI Bioscience UK Centre, Bakeham Lane, Egham, UK) and aseptically sub-cultured in Potato Dextrose Agar (PDA); it was then purified, and cultures were stored at 4 °C for long-term use, as described previously [31].
2.2. Experimental Set up and Applications
The EOs and CHL impacts on fungal biomass development and sporulation were tested in Potato Dextrose Broth (PDB) in various levels (10–50–100–500–5000 μL L−1). For the in vitro studies, two mycelia plugs were inoculated in 30 mL PDB medium containing EOs or CHL in the above-mentioned concentrations in a 250 mL glass container. The containers were closed and parafilm-sealed after the inoculation.
Regarding the fungal growth, DIT (0.35 mL L−1) oil was diluted in distilled H2O (with 5% (v/v) Tween-20) and mixed with PDA media before solidification. DIT oil application was compared with CHL (48 mL L−1) in PDA as chemical control and an untreated PDA, as a negative control treatment. The DIT and CHL concentrations were based on previous research [40,41]. The complete experimental set up is presented at Table 1. For the in vitro studies, the disc diffusion method was employed for fungal colony growth on PDA medium, as described previously [31]. In brief, PDA media inoculated with a mycelial disc from 4–5-day-old culture of B. cinerea and aliquots (100 μL) of each volatile solution (DIT or CHL) were soaked into a filter paper (3.0 cm × 3.0 cm). The paper discs were placed on the inverted lid of each Petri dish; the lids were closed and sealed with parafilm. The number of spores produced and spores’ viability were tested by adding aliquots of 30 mL of DIT or CHL solutions, in individual small tubes, which were placed inside the plastic containers just before the lids were closed. Wet filter paper was placed in each container to maintain high relative humidity (RH ~95%) during the storage period.
Two sub-experiments were carried out for pepper fruit, as detailed in Section 2.4 and Section 2.5, in order to investigate the proper applications in vivo (either volatiles or dipping). Twelve distinct fruits were subjected to each volatile compound treatment in sub-experiment I, resulting in three treatments: untreated control, chemical control treated with CHL (48 mL L−1), and fruit treated with DIT (0.35 mL L−1). Fruits were placed into 4.5 L polystyrene containers (2 fruit/container) with snap-on lids for each individual experiment. Filter paper (Whatman No. 1; 3.0 cm × 3.0 cm) was soaked with 5 mL of the desired concentration of each volatile solution, and filter paper was placed inside in the containers. Moreover, wet filter paper with distilled H2O was used for 95% RH maintenance. The volatiles were allowed to vaporize inside the containers spontaneously at 20 °C for 2 h. Control samples were handled similarly apart from the volatile treatment (controls consisted of distilled H2O with 5% (v/v) Tween-20). In vitro and in vivo experiments were repeated twice.
The sub-experiment II involved dipping 12 individual fruit per treatment into the EO or CHL solutions (same concentration as vapour treatments) for 30 min, drying the fruit for 1 h at room temperature (RT) and storing the fruit in ambient air conditions for up to 12 d.
2.3. Impact of Dittany and Chlorine Enrichment on Pathogen Growth In Vitro
Two mycelial plugs (1 cm2), obtained from the periphery of 4–5 d old culture of B. cinerea at 25 °C, were placed in the centre of containers with 30 mL PDB. Following inoculation, containers were incubated for 9 d in the dark at 11 °C and 95% RH. Fungal biomass was collected, dried, and weighed, and the results were expressed as dry weight (g), while inhibition of sporulation was calculated (considering the area measured using digital imaging tools (Adobe Photoshop CS6 analysis tools)) compared to the sporulation of the control (without DIT or CHL compounds).
A mycelial plug (2.5 mm diameter), as described above, was placed in the centre of PDA plates. Following inoculation, plates were incubated for 6 d in the containers exposed to air or volatiles in the dark at 11 °C, 95% RH (curative effect). In a second experiment, PDA plates were exposed to air or volatiles for 6 d; then, PDA medium was inoculated with the fungi (as described above), lids replaced on the plates, and plates transferred to air for an additional 6 d period (residual effect). Colony diameter was measured and results monitored as colony area (cm2) development.
2.4. Impact of Dittany and Chlorine Enrichment on Grey Mould Development in Wound-Inoculated Fruit
Two wounds (3 mm diameter and 1–2 mm deep) were made on opposite sides, and at the top and bottom, of commercial immature green pepper, using a sterilised spike. B. cinerea was inoculated as described previously [42], by inserting a mycelial plug (2.5 mm diameter) in each wound of the pepper fruit. The following sub-experiments implemented the following:
- (a)
- Peppers inoculated with B. cinerea were placed in containers and exposed continually to air or volatiles for 6 d or 12 d (examine curative effect).
- (b)
- Peppers inoculated with B. cinerea were placed in containers and exposed to air or volatiles for 6 d and then transferred to air for an additional 6 d (examine residual effect due to partial exposure to volatiles).
- (c)
- Peppers were first exposed to air or volatiles for 6 d, then removed from volatiles and inoculated with B. cinerea and transferred to air for an additional 6 d (examine residual effect on fruit; no fungal exposure to volatiles).
Fumigation was conducted in the dark at 11 °C, 95% RH. Lesion development (expressed in cm2) was measured at the end of the experiments.
2.5. Impact of Sanitary Dips on Grey Mould Development in Wound-Inoculated Fruit
In sub-experiment II, fruits were dipped into DIT or CHL solution, at the same concentrations as the vapour’s treatments. In the first treatment, the fruit inoculated with B. cinerea were incubated for 24 h at RT and then dipped into the EOs or CHL solution for 30 min, dried for 1 h at RT and transferred to air for 12 d (curative effect). In the second treatment, fruits were dipped in the EOs or CHL solutions for 30 min, dried for 1 h at RT, inoculated with B. cinerea, incubated for 24 h at RT, and transferred to storage conditions in air for 12 d (preventative effect). Incubation was performed in the dark at 11 °C, 95% RH and lesion development (cm2) measured every 2 d throughout the storage period. Both experiments were repeated twice.
2.6. Effect of Dittany and Chlorine Enrichment on Spore Production
The B. cinerea spore suspension was collected from a sporulated PDA medium (as described in Tzortzakis, 2010 [42]) by harvesting the spores using an L-shaped spreader. Spores were concentrated and counted using a haemocytometer slide. Plates (PDA medium) were inoculated in the centre with 20 μL spore solution (1.8 × 104 spores mL−1), transferred to containers, and exposed to air or volatiles (CHL or EO) for 10 d (curative effect). In the second experiment, plates were first placed in containers, exposed to air or volatiles (EO or CHL) for 6 d, and then inoculated with freshly prepared fungal suspension (preventative effect). In both experiments, the lids were removed from the plates during incubation. After exposure, the plates were closed and transferred to air for 10 d (residual effect). Following exposure to volatiles (CHL or EO) or air, spores were collected with an L-shaped spreader with 20 mL dH2O (with Tween 80; 0.1% v/v) for 5 min and concentrated to a final 1 mL volume, as described by Tzortzakis [42].
Wounded pepper fruit were inoculated with 15 μL spore suspension (2 × 104 spores mL−1) of B. cinerea in each wound, placed in containers, and exposed to air or volatiles (CHL or EO) for 15 d until spores formed. In the second experiment, healthy fruits were exposed to air or volatiles (CHL or EO) for 6 d, inoculated with B. cinerea (as above), and transferred to air for 15 d until spores formed. Fumigation was performed in the dark at 11 °C, 95% RH. Following exposure to volatiles (CHL or EO) or air, fruit was shaken for 25 min in 80 mL dH2O (with Tween 80 (0.1% v/v) for better spore separation) to remove the spores. Spore suspensions were concentrated to a final volume of 1 mL. A haemocytometer slide was used for microscope quantification counts.
2.7. Effect of Dittany and Chlorine Enrichment on Spore Germination
Spore germination tests were implemented in several treatments:
- (d)
- Spores from 9 d old B. cinerea colonies were collected with an L-shaped spreader and inoculated on PDA medium (2–3 mm thick). Plates were placed in containers, the lids were removed, and they were exposed to air or volatiles (CHL or EO) for 24 h (curative effect).
- (e)
- Plates were placed in containers, the lids removed, and they were exposed to air or volatiles (CHL or EO) for 24 h and then inoculated with the fungal suspension). Following exposure, lids were replaced, and plates were transferred to air for 24 h (residual effect).
- (f)
- Spore suspensions were made from inoculated and volatile-exposed pepper fruits, inoculated on PDA medium, and incubated in air for 24 h in the dark at 11 °C, 95% RH. Measurements were recorded after 24 h incubation. Within each of the 9 replicates (4 sets of 16 squares; 44% of the haemocytometer slide was measured, in two different areas for the PDA medium or the fruit per replicate), 100 spores were examined, and the percentage of germinated spores was calculated.
2.8. Statistical Analysis
Values were presented as means ± standard errors (SE) from 9–12 independent values per treatment. Data were subjected to analysis of variance and Duncan’s Multiple Range test (p < 0.05) using IBM SPSS v.25 for Windows.
3. Results
The composition of the main (>2.0%) EO components is presented at Table 2. Twenty-four components were identified, indicating 99.87% of the oil composition. The main components were carvacrol (69.89%), p-cymene (12.92%), and γ-Terpinene (7.12%), in agreement with previous reports [43]. The main category of the EO was oxygenated monoterpenes, averaged in 72.87%, followed by monoterpene hydrocarbons (24.31%) and then by sesquiterpene hydrocarbons (2.69%).
3.1. Effects of Dittany Oil on Fungal Biomass In Vitro
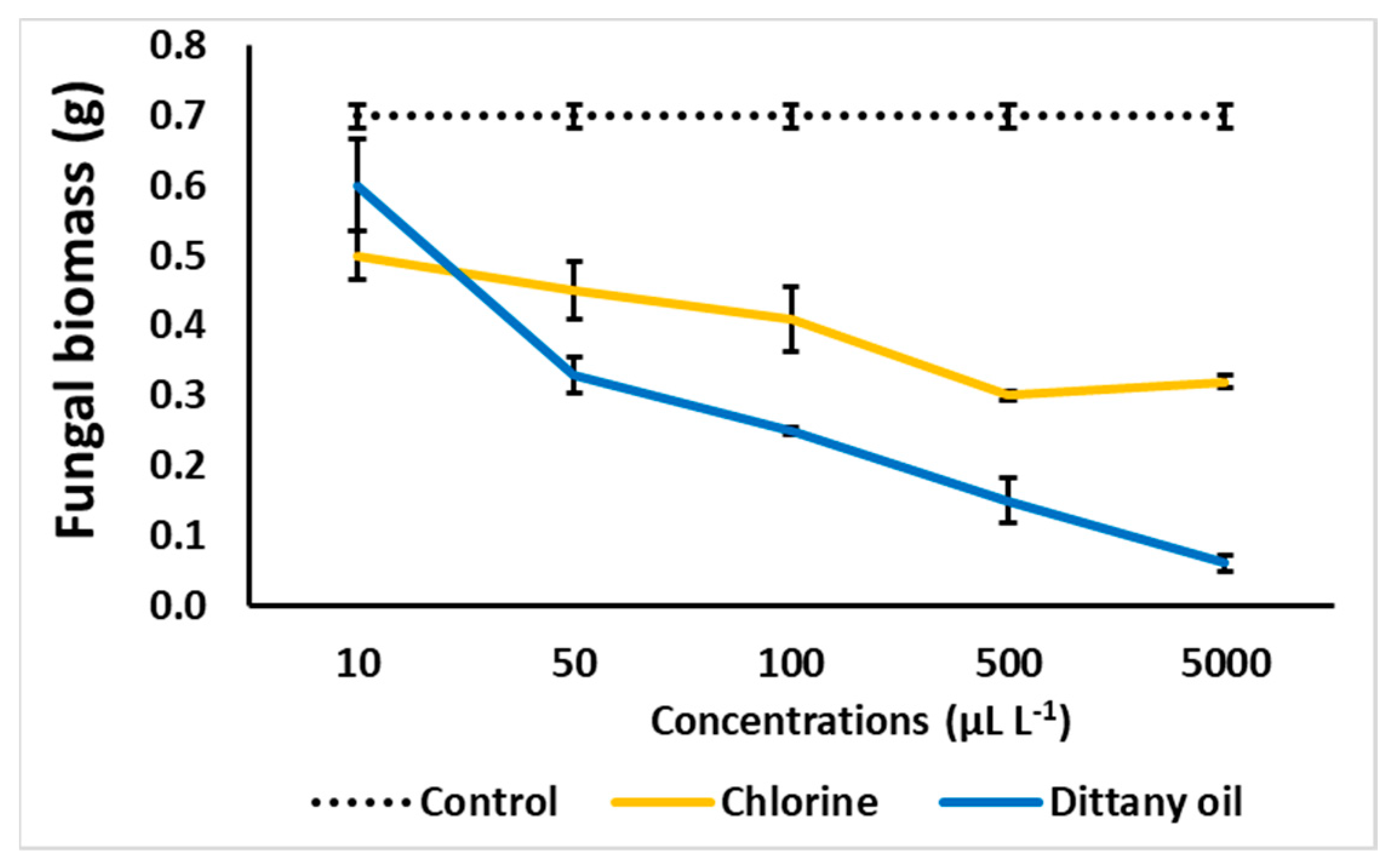
The B. cinerea biomass decreased as the concentration of the chlorine or dittany oil increased in the PDB media (Figure 1). In detail, the application of CHL decreased fungal biomass from 28.4% up to 57.1% at 10 μL L−1 and 500 μL L−1 applications, respectively, compared with the control treatment. The relevant decreases in fungal biomass found after the dittany oil applications were 14.3% at DIT 10 μL L−1 and 91.4% at DIT 500 μL L−1. Spore production accelerated at CHL ≥ 100 μL L−1 but was eliminated in the DIT applications (see Figure S1).
3.2. Effects of Vapours on Grey Mould Development In Vitro
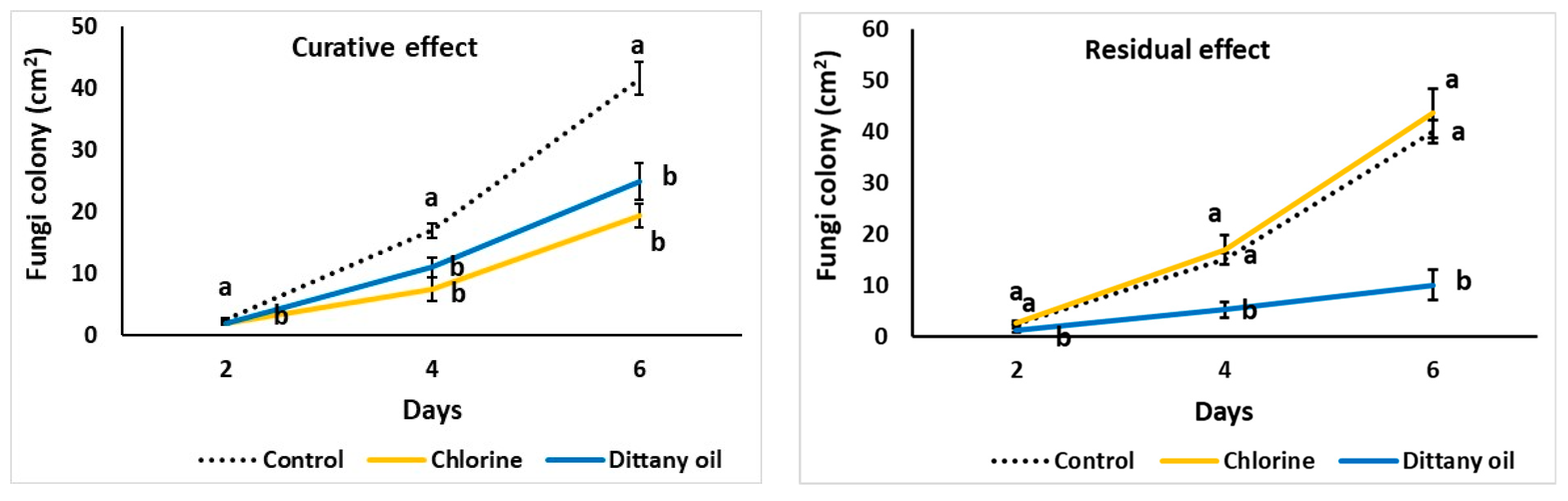
DIT and CHL vapours decreased the B. cinerea colony growth (vegetative stage) in vitro (i.e., fungi inoculated and cultured on PDA prior exposure to volatiles, named “curative effect”) following 4 and 6 d of incubation compared to the control treatment (Figure 2). However, when PDA media were pre-exposed to volatiles (named “residual effect”) and then PDA was inoculated with the fungi, only DIT oil vapor decreased fungal colony growth, while CHL vapours did not affect the colony growth, which was at similar levels as the colony growth in the control treatment (Figure 2).
The effects of CHL or DIT vapours on the reproductive phases of the fungi growth, including spore production and spore germination/viability, are illustrated in Table 3. The number of spores produced decreased (up to 50.8%) in DIT vapour enrichment compared to the control and/or CHL treatments (Table 3). In pre-exposed PDA media to vapours, spore production was suppressed by up to 55.2% and 81.9% for the CHL and DIT applications, respectively, compared with the control. Residual effects were also evidenced for the spore’s viability, as spore germination decreased (up to 43.9%) for vaporized media with CHL or DIT oil. However, spores treated with vapours (vapour exposed PDA medium) did not affect their viability (reproductive phase).
3.3. Effects of Vapours on Grey Mould Development on Pepper Fruit
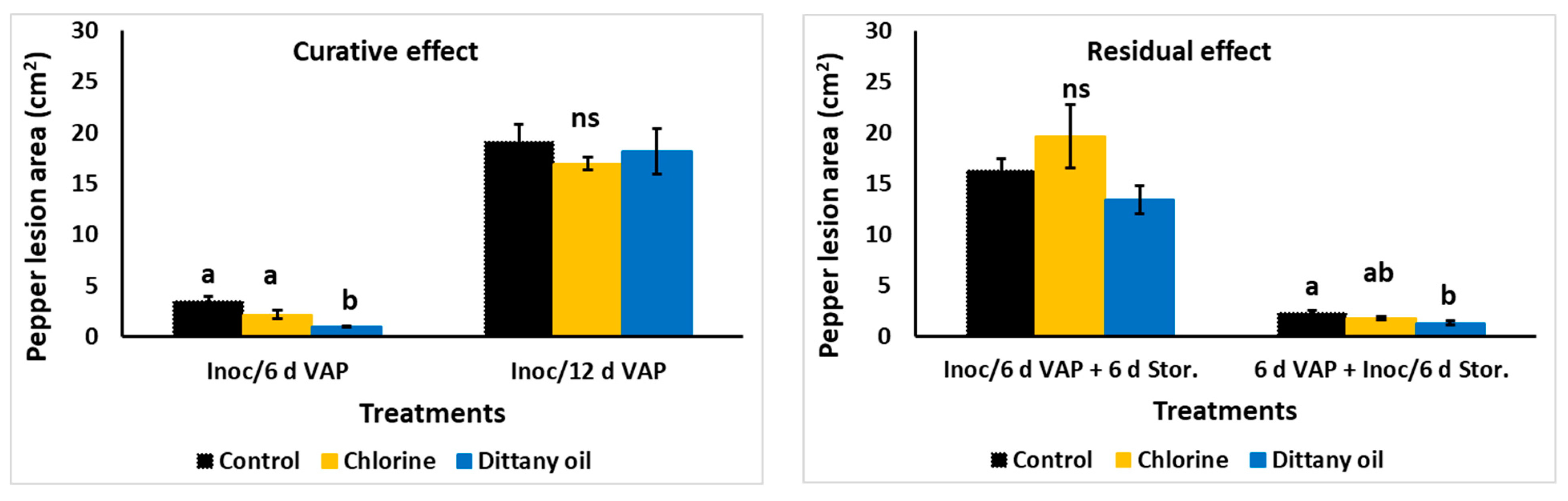
The fungal lesion development of inoculated pepper fruits with B. cinerea decreased after 6 d of storage when subjected to DIT oil vapours; however, this effect did not persist after 12 d of storage at 11 °C and 95% RH (Figure 3). Wound-inoculated fruit subjected to vapours (CHL or DIT) for 6 d and then additionally stored for 6 d in ambient air (in total 12 d of storage) failed to decrease the fungal lesion development, indicating no residual effect after 12 d of storage. However, the fungal lesion area was supressed (up to 44.7%) in pre-exposed fruits to DIT vapours for 6 d followed by B. cinerea inoculation and storage for 6 d in ambient air, indicating a residual lasting effect for the 6 d of storage in ambient air (Figure 3). The fruit exhibited no apparent signs of injury or any other abnormalities.
The impact of vapours on the grey mould development on pepper fruit is shown in Table 4. Notably, the production of spores significantly (p < 0.05) decreased during the exposure of the inoculated fruits to CHL and DIT when compared to the control (peppers exposed to ambient air throughout). However, spore germination decreased in the case of DIT-treated pepper fruits compared to control or CHL-treated fruits during the vapour applications. In peppers pre-exposed to vapours (residual effect), both spores’ production and germination were unaffected and averaged as 1.87 × 105 spores mL−1 and 98.14%, respectively (Table 4).
Indeed, the antimicrobial activity of the tested vapours was evidenced, as when pure CHL and DIT oil was assessed, fungal lesion growth decreased up to 69.4% and 53.0% for CHL and DIT applications, respectively (Table 5). The antimicrobial properties of the CHL and DIT oil were over-expressed at the spore viability, as spore germination decreased up to 97.3% and 99.7% for CHL and DIT applications, respectively.
3.4. Sanitary Dips on Grey Mould Development In Vivo
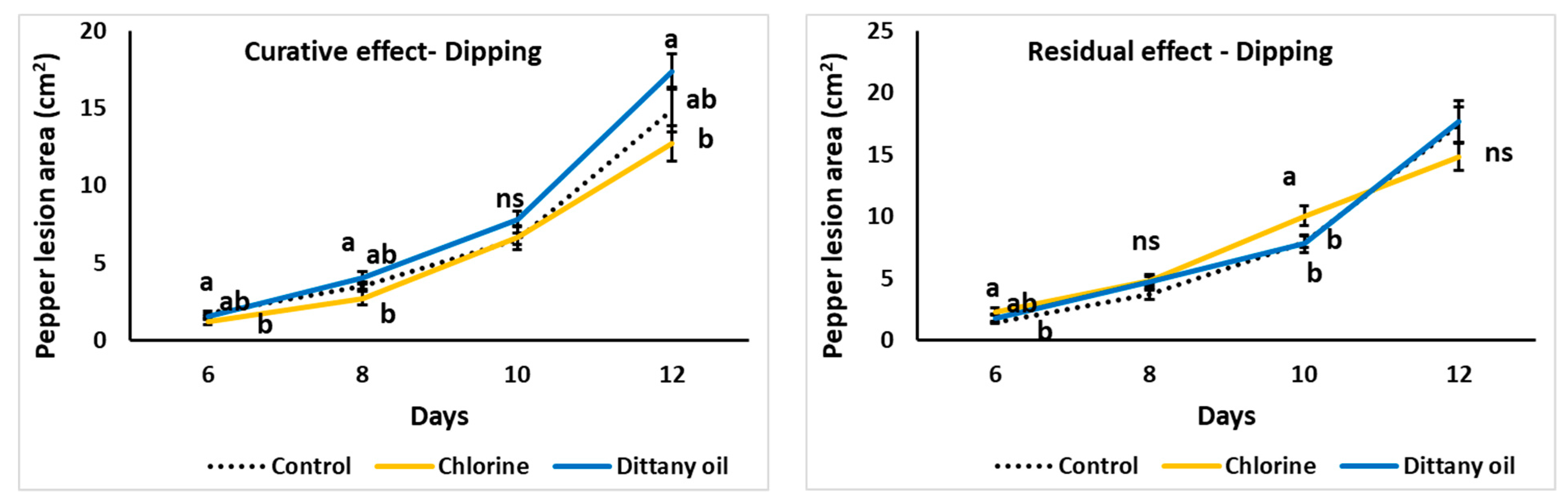
Apart from the vapour application in the present study, sanitary dips of fruit in CHL or DIT solution were also assessed. Sanitary dips on fruits previously inoculated with the fungi did not affect the lesion growth compared to the control treatment, while DIT application slightly accelerated the fungal growth (Figure 4). Similarly, sanitary dips did not reveal any residual effects, as pre-treated fruits with vapours that were inoculated had similar fungi development. Pepper fruits subjected to dipping retained a negligible odour of the sanitary solution (CHL or DIT), but fruits retained their colour as no discolouration was evidenced.
4. Discussion
Fungicide treatments for postharvest decay prevention have been limited in the EU due to severe regulations regarding concerns about food safety [15,44]. Unlike their usage as food flavourings, EOs’ potential use as natural antibacterial, antioxidant, and anticancer agents has received less research and approval, which has limited their use in the industry. This study underscores the potential use of EOs for controlling postharvest diseases caused by the B. cinerea in pepper fruit. Natural volatiles demonstrated efficacy in mitigating grey mould in pepper fruit during both the vegetative (mycelial development) and reproductive (spore germination and production) phases of B. cinerea in vitro. This interference with the fungal disease cycle is crucial for effective disease control. Interestingly, the antifungal efficacy of the DIT oil was evidenced in pre-exposed PDA media to the EOs, indicating a residual effect, which was evidenced for both the vegetative (Figure 2) and reproductive (Table 3) stages of the tested fungi. In previous studies, cinnamon (Cinnamomum zeylanicum L.) oil at ≤500 μL L−1 inhibited several Aspergilus species (A. flavus, A. parasiticus, A. ochraceus) and Fusarium moniliforme on PDA [45], being in line with the current observations. An in vitro test of Origanum vulgare (L.), Thymus capitatus (L.), Cymbopogon citratus (L.), and Coriandrum sativum (L.) vapours (0.5–1.0 mL L−1) inhibited B. cinerea and Alternaria arborescens [46]. The high C. citratus EO levels (2800 μL L−1) completely inhibited Aspergillus flavous, while the production of mycotoxins (aflatoxin) was inhibited at 100 μL L−1 of C. citratus treatments [47].
The antimicrobial properties of the DIT oil mirrored the pepper fruit as well, but the efficacy seemed to last for the first 6 d of application, as longer duration in both curative and residual applications was not promising (Figure 3). In contrast, the antimicrobial properties of DIT oil when applied in eggplant persisted up to 14 d, and the higher the concentration, the greater the suppression against grey mould [31]; this is probably related to the different commodity used. DIT oil has carvacrol as the main component based on the EO analysis (Table 2) and previous reports [43]. Carvacrol has proven antimicrobial and antioxidant properties [48,49], reflecting the DIT oils’ biocidal activities. When an EO mixture that contained carvacrol–oregano–cinnamon at a ratio of 7:1:2 was used, the controlled release of EO vapours from the active box prolonged the storage period of tomatoes and decreased decay [50]. The combination of EOs and/or other postharvest means (i.e., chitosan) improved the efficacy of the preservation of fresh produce [51]. When chitosan and thyme oil were combined in a 3:1 ratio, they controlled Colletotrichum gloeosporioides in avocado fruit during storage [29] so they can be considered a suitable alternative to the currently adopted prochloraz applications. The application of the EOs as nanoemulsions could be a possible strategy for improving the efficiency of natural fungicides [52]. Several studies on tomato, strawberry, and cucumber fruit confirmed the antifungal properties of EOs derived from oregano, thyme, clove, and lemon plants, towards various important postharvest pathogens (B. cinerea, Penicillium italicum, and Penicillium digitatum) [41,53], being in agreement with the results of the current work. Notably, the advantages associated with volatile enrichment persisted in fruit exposed to DIT oil vapours, leading to the suppression of lesion growth and direct impacts on the fungal spore germination and spore production. The viability of spores in pepper fruit, inoculated through wounds, was comparatively less affected than spore production.
Interestingly, a 6 d DIT oil application before inoculation revealed a residual effect and decreased the grey mould development after the fungi inoculation (Figure 3). These observations suggest that vapour applications may exert a lasting effect, possibly through fruit priming in anticipation of subsequent challenges [54] and potentially fostering defence-related metabolism in the fruit [55]. This provided evidence that pre-treating fruit with DIT oil during storage in chilled conditions might prevent the establishment and growth of the fungi. Molecular-based studies would be useful for comprehending the secondary metabolisms and potential mechanisms involved. Indeed, evidence shows that the EOs’ antimicrobial and antioxidant activities are related to the synthesis of phenolic components and the role of phenylalanine ammonia-lyase (PAL) as a primary enzyme that is involved in the phenolics biosynthesis [29]. Similarly, pathogen-related proteins (chitinase and b-1,3-glucanase) have a vital role in plant defence-related mechanisms against fungi by facilitating the biochemical mechanisms involved in fungi cell polymer hydrolysation [29,41]. Thyme oil decreased pathogen development and spore viability by impacting the active sites of enzymes and cellular metabolism [56]. However, DIT oil seems to pose fungistatic effects on the reproductive (spore germination and production) phase of the fungi, rather than fungicidal effects (Table 4). Nevertheless, the spore morphology, the moisture content, and the substrate are recognized as crucial factors influencing fruit susceptibility. The reduced spore germination and production emphasized in this study would impede the fungus’ spread, diminishing its ability to generate spores. Consequently, natural products become highly significant for postharvest sanitation means. In previous studies, EOs’ antimicrobial efficacy has been reported to prolong the storage life of various vegetables and fruits [29,52,57,58].
Essential oils offer a distinct advantage in their bioactivity when applied in the vapour phase, a characteristic that renders them valuable as potential sanitizer for safeguarding stored commodities. In many commodities, sanitary dipping is an effective and straightforward way to preserve fresh commodities, while in some commodities, i.e., grapes and strawberries, the dipping technique is less preferable at it may fail to protect the fresh commodities, accelerating pathogen development. In the present study, sanitary dipping was less successful in fruit sanitation compared to vapour application (Figure 4), while fruits dipped in DIT or CHL solution retained a faint odour in the sanitary solution. It has been reported previously that sage EO dipping application was ineffective for the preservation of pepper fruit [59], while sanitary dipping of kiwi (Actinidia chinensis) fruit against B. cinerea at higher rates (i.e., 500 μL L−1 aqueous solution of Origanum spp. oil) than the ones used in the present study (i.e., 350 μL L−1) revealed a fungal decrease in the fruit flesh but severely reduced fruit quality [60]. Moreover, Satureja montana and Origanum vulgare EO emulsion was effective to control Escherichia coli O157:H7 in the postharvest washing of lettuce [3]; however, the effects were varied according to the different lettuce types, romaine and butterhead versus crisp head lettuce, with greater antimicrobial activity for the latter. High EO levels (i.e., 500 μL L−1) may accelerate fruit metabolism, increase fruit decay, and decrease aroma/marketability, as evidenced on vapourised tomatoes at the breaker ripening stage when exposed to sage (Salvia trilova L.) EO, while lower EO concentration of 50 μL L−1 maintained fruit texture, respiration, and ethylene production [55]. In that study, it was also observed that even 2 d of sage EO application can affect the fruit metabolism and storability, while residual effects were evidenced in pre-exposed tomatoes to sage EO for 7 d, following an additional 7 days of storage at 11 °C and 90% RH [55]. Essential oil addition in foodstuffs might have limitations in terms of not only aroma and taste but also colour and texture, and the general visual perception may be altered [23]. In that sense, EO application should balance the impacts on fresh produce quality, safety, and acceptability with the antimicrobial efficacy of the EO.
The application of DIT EOs ≥ 50 μL L−1 led to a decrease in fungal biomass, which almost zeroed at the highest tested concentration of 5000 μL L−1. The DIT presence affected the fungal development (fungal biomass (Figure 1)) and mycelial growth (Figure 2) per se but also possibly altered the culture medium composition/properties. This was confirmed by exposing the PDA medium to vapours prior the fungi inoculation, where the fungi colony decreased in pre-exposed PDA to DIT vapours, indicating residual effects. Further investigation is needed to examine the different DIT oil concentrations and the time of duration/exposure to vapours. There is no doubt that DIT oil poses strong antimicrobial activity, as pure DIT oil decreased, by almost half, the fungal vegetative growth and, most importantly, nearly prevented the fungi spread, as the spore viability was almost zeroed (Table 5). The mechanism by which volatiles affect fungal spores (germination and production) is not well understood. The EO fungitoxic properties are likely associated with the combined action of various components that the EOs are composed of [25]. This complex matrix of EO components may hinder pathogens from developing resistance, providing a valuable advantage for postharvest storage. Further research is needed to understand any synergistic action of plant molecules, both in vitro and in vivo, and application should be further explored under semi-commercial and full-commercial (i.e., packinghouses) scales and not only under laboratory premises.
Reduced spore production and spore germination, as demonstrated in this study, could effectively inhibit fungal proliferation. This positions EOs as useful postharvest sanitation tools, offering a substitute to commercial chemical-based chlorine [61]. Essential oils, approved as food additives, present a more straightforward registration process for postharvest use compared to novel synthetic pesticides. Thymol, thyme EO, and thyme (as a spice) are involved in the lists of the US Food and Drug Administration as both food and food additives, with thymol first registered as a pesticide in the US in 1964 [62]. When applying EOs, achieving an optimized concentration, duration, and exposure method is crucial to prevent product deterioration. The thickness of fresh commodity is linked to tainting, with thicker fruit skins being less susceptible to such issues [63]. Since the effectiveness of a single natural sanitizer might be low and may not be as broadly effective as synthetic pesticides, during postharvest applications, their efficacy can be enhanced by combining them with specially designed packaging. This approach avoids leaving detectable residues on the surface, addressing concerns about the available sanitation means, which are primarily chlorine-based. Additionally, applying EOs via the vapour phase is likely to be more cost-effective than dipping.
While utilizing EOs, it is important to keep in mind that their effectiveness towards fungal infections and/or other pathogens attacks relies on the EO composition, concentration, methods of applications, timing, pathogen type, and the features of the commodity [64]. Generally, the concentrations of EOs and their components are required to hinder microbial growth to be greater in food compared to culture media. Any commercial application should take this phenomenon into account, as it is linked to interactions among phenolic ingredients and the food matrix [65]. However, the effects of EO application on fresh produce quality can be contradictory. For instance, the use of savoury and thyme EOs in vapour form on peaches and nectarines benefited the fruit’s qualitative features (i.e., less weight loss and significant losses of carotenoid and ascorbic acid content) [66]. However, the high Salvia trilova EO concentrations may have a negative impact on quality factors, including the sensory qualities (i.e., odour, taste, and colour) of the treated fresh produce [55]. Upon utilization, EOs might have direct (lysis of membrane permeability) or indirect (induce resistance) activity. Essential oils, due to their lipophilicity, can easily disrupt the permeability of the membrane and/or enter it [67,68].
5. Conclusions
EOs, as a natural product, are an eco-friendly substitute for chemical preservatives in the food sector, attracting attention as a natural green preservative. The present study demonstrates the potential use of DIT oil for postharvest disease management, both in the vegetative and reproductive phases of Botrytis cinerea. DIT oil decreased fungal lesion growth and prevented the spore germination and spore production to some extent, even better in most cases than the commonly used sanitizer, chlorine, for the postharvest storage of pepper fruit. However, these effects were observed for the 6 d vapour application and not for 12 d, indicating the lasting effect of the vapours on fresh produce preservation. DIT oil application reveals residual effects that could possibly protect the fruit from opportunistic pathogens during the storage of fresh commodities. The EO of DIT may be considered as curative and residual alternatives to typical fresh produce preservation procedures. In comparison to sanitary dips, vapour application was found to be more successful in pepper fruit preservation. Before commercial use, each commodity must be evaluated individually, and the volatile concentration and sanitizing procedure must be optimized.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14020257/s1, Figure S1. Effect of chlorine or dittany essential oil on fungal (Botrytis cinerea) sporulation grown in Potato Dextrose Broth (PDB).
Funding
This research received no external funding.
Data Availability Statement
The authors declare that data are available only upon request.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Cao, Z.; Zhou, D.; Ge, X.; Luo, Y.; Su, J. The role of essential oils in maintaining the postharvest quality and preservation of peach and other fruits. J. Food Biochem. 2022, 46, e14513. [Google Scholar] [CrossRef]
- Domingos, M.M.; Valiati, B.S.; Uliana, D.S.; Leal, R.M.L.; Guimarães, A.P.; Lepaus, B.M.; Geringer, S.J.; de São José, J.F.B. Essential oils against fruit spoilage fungi. In Plant Essential Oils; Prakash, B., Dubey, N.K., de São José, J.F.B., Eds.; Springer: Singapore, 2023; pp. 105–124. ISBN 978-981-99-4369-2. [Google Scholar]
- Pizzo, J.S.; Pelvine, R.A.; da Silva, A.L.B.R.; Mikcha, J.M.G.; Visentainer, J.V.; Rodrigues, C. Use of essential oil emulsions to control Escherichia coli O157:H7 in the postharvest washing of lettuce. Foods 2023, 12, 2571. [Google Scholar] [CrossRef]
- López-Gómez, A.; Ros-Chumillas, M.; Antolinos, V.; Buendía-Moreno, L.; Navarro-Segura, L.; Sánchez-Martínez, M.J.; Martínez-Hernández, G.B.; Soto-Jover, S. Fresh culinary herbs decontamination with essential oil vapours applied under vacuum conditions. Postharvest Biol. Technol. 2019, 156, 110942. [Google Scholar] [CrossRef]
- Yeni, F.; Yavaş, S.; Alpas, H.; Soyer, Y. Most common foodborne pathogens and mycotoxins on fresh produce: A review of recent outbreaks. Crit. Rev. Food Sci. Nutr. 2016, 56, 1532–1544. [Google Scholar] [CrossRef]
- Mitropoulou, G.; Fitsiou, E.; Stavropoulou, E.; Papavassilopoulou, E.; Vamvakias, M.; Pappa, A.; Oreopoulou, A.; Kourkoutas, Y. Composition, antimicrobial, antioxidant, and antiproliferative activity of Origanum dictamnus (dittany) essential oil. Microb. Ecol. Health Dis. 2015, 26, 26543. [Google Scholar] [CrossRef] [PubMed]
- Coroneo, V.; Carraro, V.; Marras, B.; Marrucci, A.; Succa, S.; Meloni, B.; Pinna, A.; Angioni, A.; Sanna, A.; Schintu, M. Presence of trihalomethanes in ready-to-eat vegetables disinfected with chlorine. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess 2017, 34, 2111–2117. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Romero, D.; Castillo, S.; Guillén, F.; Díaz-mula, H.M.; Zapata, P.J.; Valero, D.; Serrano, M. Aloe vera gel coating maintains quality and safety of ready-to-eat pomegranate arils. Postharvest Biol. Technol. 2013, 86, 107–112. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Xylia, P.; Chrysargyris, A. Sage essential oil improves the effectiveness of Aloe vera gel on postharvest quality of tomato fruit. Agronomy 2019, 9, 635. [Google Scholar] [CrossRef]
- Khursheed, A.; Rather, M.A.; Jain, V.; Wani, A.R.; Rasool, S.; Nazir, R.; Malik, N.A.; Majid, S.A. Plant based natural products as potential ecofriendly and safer biopesticides: A comprehensive overview of their advantages over conventional pesticides, limitations and regulatory aspects. Microb. Pathog. 2022, 173, 105854. [Google Scholar] [CrossRef]
- Ibáñez, M.D.; Blázquez, M.A. Phytotoxic effects of commercial essential oils on selected vegetable crops: Cucumber and tomato. Sustain. Chem. Pharm. 2020, 15, 100209. [Google Scholar] [CrossRef]
- Inselberg, H.; do Nascimento Nunes, M.C. Using Cannabidiol as a potential postharvest treatment to maintain quality and extend the shelf life of strawberries. Postharvest Biol. Technol. 2021, 173, 111416. [Google Scholar] [CrossRef]
- Jakubczyk, K.; Tuchowska, A.; Janda-Milczarek, K. Plant hydrolates—Antioxidant properties, chemical composition and potential applications. Biomed. Pharmacother. 2021, 142, 112033. [Google Scholar] [CrossRef]
- Xylia, P.; Clark, A.; Chrysargyris, A.; Romanazzi, G.; Tzortzakis, N. Quality and safety attributes on shredded carrots by using Origanum majorana and ascorbic acid. Postharvest Biol. Technol. 2019, 155, 120–129. [Google Scholar] [CrossRef]
- Cindi, M.D.; Soundy, P.; Romanazzi, G.; Sivakumar, D. Different defense responses and brown rot control in two Prunus persica cultivars to essential oil vapours after storage. Postharvest Biol. Technol. 2016, 119, 9–17. [Google Scholar] [CrossRef]
- Alam, A.; Tripathi, A.; Sharma, V.; Sharma, N. Essential oils: A novel consumer and eco-friendly approach to combat postharvest phytopathogens. J. Adv. Biol. Biotechnol. 2017, 11, 1–16. [Google Scholar] [CrossRef]
- Vergis, J.; Gokulakrishnan, P.; Agarwal, R.K.; Kumar, A. Essential oils as natural food antimicrobial agents: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Chrysargyris, A.; Charalambous, S.; Xylia, P.; Litskas, V.; Stavrinides, M.; Tzortzakis, N. Assessing the biostimulant effects of a novel plant-based formulation on tomato crop. Sustainability 2020, 12, 8432. [Google Scholar] [CrossRef]
- Isman, M.B.; Wilson, J.A.; Bradbury, R. Insecticidal activities of commercial rosemary oils (Rosmarinus officinalis) against larvae of Pseudaletia unipuncta and Trichoplusia ni in relation to their chemical compositions. Pharm. Biol. 2008, 46, 82–87. [Google Scholar] [CrossRef]
- Di Sotto, A.; Gullì, M.; Acquaviva, A.; Tacchini, M.; Di Simone, S.C.; Chiavaroli, A.; Recinella, L.; Leone, S.; Brunetti, L.; Orlando, G.; et al. Phytochemical and pharmacological profiles of the essential oil from the inflorescences of the Cannabis sativa L. Ind. Crop. Prod. 2022, 183, 114980. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Ziaee, M.; Palla, F. Essential oils extracted from different species of the Lamiaceae plant family as prospective bioagents against several detrimental pests. Molecules 2020, 25, 1556. [Google Scholar] [CrossRef]
- Xylia, P.; Chrysargyris, A.; Miltiadous, P.; Tzortzakis, N. Origanum dubium (cypriot oregano) as a promising sanitizing agent against Salmonella enterica and Listeria monocytogenes on tomato and cucumber fruits. Biology 2022, 11, 1772. [Google Scholar] [CrossRef] [PubMed]
- Osimani, A.; Garofalo, C.; Harasym, J.; Aquilanti, L. Use of essential oils against foodborne spoilage yeasts: Advantages and drawbacks. Curr. Opin. Food Sci. 2022, 45, 100821. [Google Scholar] [CrossRef]
- Serrano, M.; Martínez-Romero, D.; Castillo, S.; Guillén, F.; Valero, D. The use of natural antifungal compounds improves the beneficial effect of MAP in sweet cherry storage. Innov. Food Sci. Emerg. Technol. 2005, 6, 115–123. [Google Scholar] [CrossRef]
- Xylia, P.; Chrysargyris, A.; Ahmed, Z.F.R.; Tzortzakis, N. Application of rosemary and eucalyptus essential oils and their main component on the preservation of apple and pear fruits. Horticulture 2021, 7, 479. [Google Scholar] [CrossRef]
- Hassani, A.; Fathi, Z.; Ghosta, Y.; Abdollahi, A.; Meshkatalsadat, M.H.; Marandi, R.J. Evaluation of plant essential oils for control of postharvest brown and gray mold rots on apricot. J. Food Saf. 2012, 32, 94–101. [Google Scholar] [CrossRef]
- Mohammadi, L.; Ramezanian, A.; Tanaka, F.; Tanaka, F. Impact of Aloe vera gel coating enriched with basil (Ocimum basilicum L.) essential oil on postharvest quality of strawberry fruit. J. Food Meas. Charact. 2021, 15, 353–362. [Google Scholar] [CrossRef]
- Rabiei, V.; Shirzadeh, E.; Rabbiangourani, H.; Sharafi, Y. Effect of thyme and lavender essential oils on the qualitative and quantitative traits and storage life of apple “Jonagold” cultivar. J. Med. Plant Res. 2011, 5, 5522–5527. [Google Scholar]
- Bill, M.; Sivakumar, D.; Korsten, L.; Thompson, A.K. The efficacy of combined application of edible coatings and thyme oil in inducing resistance components in avocado (Persea americana Mill.) against anthracnose during post-harvest storage. Crop. Prot. 2014, 64, 159–167. [Google Scholar] [CrossRef]
- Sakkas, H.; Papadopoulou, C. Antimicrobial activity of basil, oregano, and thyme essential oils. J. Microbiol. Biotechnol. 2017, 27, 429–438. [Google Scholar] [CrossRef]
- Stavropoulou, A.; Loulakakis, K.; Magan, N.; Tzortzakis, N. Origanum dictamnus oil vapour suppresses the development of grey mould in eggplant fruit in vitro. Biomed Res. Int. 2014, 2014, 11. [Google Scholar] [CrossRef]
- Skotti, E.; Anastasaki, E.; Kanellou, G.; Polissiou, M.; Tarantilis, P.A. Total phenolic content, antioxidant activity and toxicity of aqueous extracts from selected Greek medicinal and aromatic plants. Ind. Crop. Prod. 2014, 53, 46–54. [Google Scholar] [CrossRef]
- Chorianopoulos, N.; Kalpoutzakis, E.; Aligiannis, N.; Mitaku, S.; Nychas, G.J.; Haroutounian, S.A. Essential oils of Satureja, Origanum, and Thymus species: Chemical composition and antibacterial activities against foodborne pathogens. J. Agric. Food Chem. 2004, 52, 8261–8267. [Google Scholar] [CrossRef] [PubMed]
- Olatunji, T.L.; Afolayan, A.J. The suitability of chili pepper (Capsicum annuum L.) for alleviating human micronutrient dietary deficiencies: A review. Food Sci. Nutr. 2018, 6, 2239–2251. [Google Scholar] [CrossRef] [PubMed]
- Taheri, A.; Behnamian, M.; Dezhsetan, S.; Karimirad, R. Shelf life extension of bell pepper by application of chitosan nanoparticles containing Heracleum persicum fruit essential oil. Postharvest Biol. Technol. 2020, 170, 111313. [Google Scholar] [CrossRef]
- González-Saucedo, A.; Barrera-Necha, L.L.; Ventura-Aguilar, R.I.; Correa-Pacheco, Z.N.; Bautista-Baños, S.; Hernández-López, M. Extension of the postharvest quality of bell pepper by applying nanostructured coatings of chitosan with Byrsonima crassifolia extract (L.) Kunth. Postharvest Biol. Technol. 2019, 149, 74–82. [Google Scholar] [CrossRef]
- Romanazzi, G.; Feliziani, E.; Santini, M.; Landi, L. Effectiveness of postharvest treatment with chitosan and other resistance inducers in the control of storage decay of strawberry. Postharvest Biol. Technol. 2013, 75, 24–27. [Google Scholar] [CrossRef]
- Znini, M.; Cristofari, G.; Majidi, L.; Mazouz, H.; Tomi, P.; Paolini, J.; Costa, J. Antifungal activity of essential oil from Asteriscus graveolens against postharvest phytopathogenic fungi in apples. Nat. Prod. Commun. 2011, 6, 1763–1768. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.P. Identification of Essential oil Components by Gas Chromatography/Mass Spectrometry, 5th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; ISBN 9781932633214. [Google Scholar]
- Tzortzakis, N. Impact of cinnamon oil-enrichment on microbial spoilage of fresh produce. Innov. Food Sci. Emerg. Technol. 2009, 10, 97–102. [Google Scholar] [CrossRef]
- Chen, C.; Cai, N.; Chen, J.; Wan, C. Clove essential oil as an alternative approach to control postharvest blue mold caused by Penicillium italicum in citrus fruit. Biomolecules 2019, 9, 197. [Google Scholar] [CrossRef]
- Tzortzakis, N.G. Ethanol, vinegar and Origanum vulgare oil vapour suppress the development of anthracnose rot in tomato fruit. Int. J. Food Microbiol. 2010, 142, 14–18. [Google Scholar] [CrossRef]
- Liolios, C.C.; Graikou, K.; Skaltsa, E.; Chinou, I. Dittany of Crete: A botanical and ethnopharmacological review. J. Ethnopharmacol. 2010, 131, 229–241. [Google Scholar] [CrossRef]
- Martínez-Romero, D.; Serrano, M.; Bailén, G.; Guillén, F.; Zapata, P.J.; Valverde, J.M.; Castillo, S.; Fuentes, M.; Valero, D. The use of a natural fungicide as an alternative to preharvest synthetic fungicide treatments to control lettuce deterioration during postharvest storage. Postharvest Biol. Technol. 2008, 47, 54–60. [Google Scholar] [CrossRef]
- Soliman, K.M.; Badeaa, R.I. Effect of oil extracted from some medicinal plants on different mycotoxigenic fungi. Food Chem. Toxicol. 2002, 40, 1669–1675. [Google Scholar] [CrossRef]
- Plotto, A.; Roberts, D.D.; Roberts, R.G. Evaluation of plant essential oils as natural postharvest disease control of tomato (Lycopersicon esculentum). Acta Hortic. 2003, 628, 737–745. [Google Scholar] [CrossRef]
- Paranagama, P.A.; Abeysekera, K.H.T.; Abeywickrama, K.; Nugaliyadde, L. Fungicidal and anti-aflatoxigenic effects of the essential oil of Cymbopogon citratus (DC.) Stapf. (lemongrass) against Aspergillus flavus Link. isolated from stored rice. Lett. Appl. Microbiol. 2003, 37, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Garcia, I.; Silva-Espinoza, B.A.; Ortega-Ramirez, L.A.; Leyva, J.M.; Siddiqui, M.W.; Cruz-Valenzuela, M.R.; Gonzalez-Aguilar, G.A.; Ayala-Zavala, J.F. Oregano essential oil as an antimicrobial and antioxidant additive in food products. Crit. Rev. Food Sci. Nutr. 2016, 56, 1717–1727. [Google Scholar] [CrossRef] [PubMed]
- de Medeiros Barbosa, I.; da Costa Medeiros, J.A.; de Oliveira, K.Á.R.; Gomes-Neto, N.J.; Tavares, J.F.; Magnani, M.; de Souza, E.L. Efficacy of the combined application of oregano and rosemary essential oils for the control of Escherichia coli, Listeria monocytogenes and Salmonella Enteritidis in leafy vegetables. Food Control. 2016, 59, 468–477. [Google Scholar] [CrossRef]
- Buendía-Moreno, L.; Ros-Chumillas, M.; Navarro-Segura, L.; Sánchez-Martínez, M.J.; Soto-Jover, S.; Antolinos, V.; Martínez-Hernández, G.B.; López-Gómez, A. Effects of an active cardboard box using encapsulated essential oils on the tomato shelf life. Food Bioprocess Technol. 2019, 12, 1548–1558. [Google Scholar] [CrossRef]
- Correa-Pacheco, Z.N.; Bautista-Baños, S.; Valle-Marquina, M.Á.; Hernández-López, M. The Effect of Nanostructured Chitosan and Chitosan-thyme Essential Oil Coatings on Colletotrichum gloeosporioides Growth in vitro and on cv Hass Avocado and Fruit Quality. J. Phytopathol. 2017, 165, 297–305. [Google Scholar] [CrossRef]
- Naserzadeh, Y.; Mahmoudi, N.; Pakina, E. Antipathogenic effects of emulsion and nanoemulsion of cinnamon essential oil against Rhizopus rot and grey mold on strawberry fruits. Foods Raw Mater. 2019, 7, 210–216. [Google Scholar] [CrossRef]
- Vitoratos, A.; Bilalis, D.; Karkanis, A.; Efthimiadou, A. Antifungal activity of plant essential oils against Botrytis cinerea, Penicillium italicum and Penicillium digitatum. Not. Bot. Horti Agrobot. Cluj-Napoca 2013, 41, 86–92. [Google Scholar] [CrossRef]
- Conrath, U.; Pieterse, C.M.J.; Mauch-mani, B. Priming in plant–pathogen interactions. Trends Plant Sci. 2002, 7, 210–216. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Rousos, C.; Xylia, P.; Tzortzakis, N. Vapour application of sage essential oil maintain tomato fruit quality in breaker and red ripening stages. Plants 2021, 10, 2645. [Google Scholar] [CrossRef]
- Arrebola, E.; Sivakumar, D.; Bacigalupo, R.; Korsten, L. Combined application of antagonist Bacillus amyloliquefaciens and essential oils for the control of peach postharvest diseases. Crop. Prot. 2010, 29, 369–377. [Google Scholar] [CrossRef]
- Tornuk, F.; Cankurt, H.; Ozturk, I.; Sagdic, O.; Bayram, O.; Yetim, H. Efficacy of various plant hydrosols as natural food sanitizers in reducing Escherichia coli O157:H7 and Salmonella Typhimurium on fresh cut carrots and apples. Int. J. Food Microbiol. 2011, 148, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Reyes, J.G.; Spadaro, D.; Prelle, A.; Garibaldi, A.; Gullino, M.L. Efficacy of plant essential oils on postharvest control of rots caused by fungi on different stone fruits in vivo. J. Food Prot. 2013, 76, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Tzortzakis, N.; Chrysargyris, A.; Sivakumar, D.; Loulakakis, K. Vapour or dipping applications of methyl jasmonate, vinegar and sage oil for pepper fruit sanitation towards grey mould. Postharvest Biol. Technol. 2016, 118, 120–127. [Google Scholar] [CrossRef]
- Thanassoulopoulos, C.C.; Yanna, L. On the biological control of Botrytis cinerea on kiwifruit cv “Hayward” during storage. Acta Hortic. 1997, 444, 757–764. [Google Scholar] [CrossRef]
- Pablos, C.; Fernández, A.; Thackeray, A.; Marugán, J. Effects of natural antimicrobials on prevention and reduction of bacterial cross-contamination during the washing of ready-to-eat fresh-cut lettuce. Food Sci. Technol. Int. 2017, 23, 403–414. [Google Scholar] [CrossRef]
- Tripathi, P.; Dubey, N.K. Exploitation of natural products as an alternative strategy to control postharvest fungal rotting of fruit and vegetables. Postharvest Biol. Technol. 2004, 32, 235–245. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.M.F.D.; Steffens, C.A.; Argenta, L.C.; do Amarante, C.V.T.; Oster, A.H.; Casa, R.T.; Amarante, A.G.M.; Espíndola, B.P. Essential oils for the postharvest control of blue mold and quality of “Fuji” apples. Pesqui. Agropecu. Bras. 2018, 53, 547–556. [Google Scholar] [CrossRef]
- Karampoula, F.; Giaouris, E.; Deschamps, J.; Doulgeraki, A.I.; Nychas, G.J.E.; Dubois-Brissonnet, F. Hydrosol of Thymbra capitata is a highly efficient biocide against Salmonella enterica serovar Typhimurium biofilms. Appl. Environ. Microbiol. 2016, 82, 5309–5319. [Google Scholar] [CrossRef] [PubMed]
- Santoro, K.; Maghenzani, M.; Chiabrando, V.; Bosio, P.; Gullino, M.L.; Spadaro, D.; Giacalone, G. Thyme and savory essential oil vapor treatments control brown rot and improve the storage quality of peaches and nectarines, but could favor gray mold. Foods 2018, 7, 7. [Google Scholar] [CrossRef]
- Perricone, M.; Arace, E.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Bioactivity of essential oils: A review on their interaction with food components. Front. Microbiol. 2015, 6, 76. [Google Scholar] [CrossRef]
- Snoussi, M.; Noumi, E.; Trabelsi, N.; Flamini, G.; Papetti, A.; De Feo, V. Mentha spicata Essential oil: Chemical composition, antioxidant and antibacterial activities against planktonic and biofilm cultures of vibrio spp. strains. Molecules 2015, 20, 14402–14424. [Google Scholar] [CrossRef]
Figure 1.
Impact of chlorine or dittany oil on fungal biomass (g) of Botrytis cinerea grown in Potato Dextrose Broth (PDB). Inoculated PDB with B. cinerea was exposed to vapours and incubated at 11 °C and 95% RH. Values represent means of measurements made on six independent replicates per treatment.
Figure 1.
Impact of chlorine or dittany oil on fungal biomass (g) of Botrytis cinerea grown in Potato Dextrose Broth (PDB). Inoculated PDB with B. cinerea was exposed to vapours and incubated at 11 °C and 95% RH. Values represent means of measurements made on six independent replicates per treatment.

Figure 2.
Impact of curative or residual vapour enrichment of chlorine or dittany oil on Botrytis cinerea colony growth (cm2) on PDA media. Values represent means (±SE) of measurements made on nine independent plates per treatment. Values followed by the same letter on each day do not differ significantly at p = 0.05.
Figure 2.
Impact of curative or residual vapour enrichment of chlorine or dittany oil on Botrytis cinerea colony growth (cm2) on PDA media. Values represent means (±SE) of measurements made on nine independent plates per treatment. Values followed by the same letter on each day do not differ significantly at p = 0.05.

Figure 3.
Impact of curative or residual vapour (VAP) enrichment of chlorine or dittany oil on Botrytis cinerea lesion growth (cm2) on pepper fruit. Values represent means (±SE) of measurements made on nine pepper fruits per treatment. Values followed by the same letter in each treatment/application do not differ significantly at p = 0.05. The ns indicates no significance.
Figure 3.
Impact of curative or residual vapour (VAP) enrichment of chlorine or dittany oil on Botrytis cinerea lesion growth (cm2) on pepper fruit. Values represent means (±SE) of measurements made on nine pepper fruits per treatment. Values followed by the same letter in each treatment/application do not differ significantly at p = 0.05. The ns indicates no significance.

Figure 4.
Impact of curative or residual dipping-enrichment of chlorine or dittany oil on Botrytis cinerea lesion growth (cm2) on pepper fruits stored at 11 °C and 95% RH. Values represent means (±SE) of measurements made on nine independent fruits per treatment. Values followed by the same letter on each day do not differ significantly at p = 0.05. The ns indicates no significance.
Figure 4.
Impact of curative or residual dipping-enrichment of chlorine or dittany oil on Botrytis cinerea lesion growth (cm2) on pepper fruits stored at 11 °C and 95% RH. Values represent means (±SE) of measurements made on nine independent fruits per treatment. Values followed by the same letter on each day do not differ significantly at p = 0.05. The ns indicates no significance.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The experimental set up of the volatiles (chlorine or dittany essential oil) on Botrytis cinerea development in vitro (PDA medium) or in vivo (pepper fruit). In curative effect (CE) fruits (or plates) inoculated with B. cinerea and then exposed to vapours. A second sample of fruits (or plates) pre-exposed to vapours (residual effect-RE), inoculated with fungi and transferred/stored to ambient air (control). Treatments maintained throughout at 11 °C and 95% RH.
Table 1.
The experimental set up of the volatiles (chlorine or dittany essential oil) on Botrytis cinerea development in vitro (PDA medium) or in vivo (pepper fruit). In curative effect (CE) fruits (or plates) inoculated with B. cinerea and then exposed to vapours. A second sample of fruits (or plates) pre-exposed to vapours (residual effect-RE), inoculated with fungi and transferred/stored to ambient air (control). Treatments maintained throughout at 11 °C and 95% RH.
| In Vitro | Conditions | Treatment | Section |
|---|---|---|---|
| Colony growth (CE-VAP) | at 20 °C for 2 h, at 11 °C for 6 d | Inoc.-6 d in VAP | 2.3 |
| Colony growth (RE-VAP) | at 20 °C for 2 h, at 11 °C for 6 d, at 11 °C for 6 d | 6 d in VAP-Inoc.-6 d in air | 2.3 |
| Spore production (CE-VAP) | at 11 °C for 10 d | Inoc.-10 d in VAP | 2.6 |
| Spore production (RE-VAP) | at 11 °C for 6 d, at 11 °C for 10 d | 6 d in VAP-Inoc.-10 d in air | 2.6 |
| Spore germination (CE-VAP) | at 11 °C for 10 d, at 11 °C for 24 h | Inoc.-10 d in VAP-Inoc. in PDA-24 h in air | 2.7 a |
| Spore germination (RE-VAP) | at 11 °C for 6 d, at 11 °C for 10 d, at 11 °C for 24 h | 6 d in VAP-Inoc.-10 d in air-Inoc. in PDA-24 h in air | 2.7 b |
| In Vivo | |||
| Lesion growth (CE-VAP) | at 20 °C for 2 h, at 11 °C for 12 d | Inoc.-12 d in VAP | 2.4 a |
| Lesion growth (RE-VAP) | at 20 °C for 2 h, at 11 °C for 6 d, at 11 °C for 6 d | Inoc.-6 d in VAP-6 d in air | 2.4 b |
| Lesion growth (RE-VAP) | at 20 °C for 2 h, at 11 °C for 6 d, at 11 °C for 6 d | 6 d in VAP-Inoc.-6 d in air | 2.4 c |
| Lesion growth (CE-DIP) | at 20 °C for 24 h, at 11 °C for 12 d | Inoc.-1 d at RT-30 min in DIP-12 d in air | 2.5 |
| Lesion growth (RE-DIP) | at 20 °C for 24 h, at 11 °C for 12 d | 30 min in DIP-Inoc.-1 d at RT-12 d in air | 2.5 |
| Spore production (CE-VAP) | at 11 °C for 15 d | Inoc.-15 d in VAP | 2.6 |
| Spore production (RE-VAP) | at 11 °C for 6 d, at 11 °C for 15 d | 6 d in VAP-Inoc.-15 d in air | 2.6 |
| Spore germination (CE-VAP) | at 11 °C for 15 d, at 11 °C for 24 h | Inoc.-15 d in VAP-Inoc. in PDA-24 h in air | 2.7 c |
| Spore germination (RE-VAP) | at 11 °C for 6 d, at 11 °C for 10 d, at 11 °C for 24 h | 6 d in VAP-Inoc.-15 d in air-Inoc in PDA-24 h in air | 2.7 c |
VAP: vapours; DIP: dipping; RE: residual effect; CE: curative effect; PDA: potato dextrose agar; Inoc.: inoculation. Letters a, b, c under the “section” heading correspond with sections in the text.
Table 2.
The chemical and percentage composition of the major components from the essential oil of Origanum dictamnus.
Table 2.
The chemical and percentage composition of the major components from the essential oil of Origanum dictamnus.
| No | Compound | R.I. b | Retention Time | Percentage (%) Composition |
|---|---|---|---|---|
| 1 | a-Thujene | 930 | 9.507 | 0.67 |
| 2 | a-Pinene a | 939 | 9.746 | 0.57 |
| 3 | Sabinene | 975 | 11.253 | 0.06 |
| 4 | b-Pinene a | 979 | 11.353 | 0.10 |
| 5 | 1-Octen-3-ol | 979 | 11.530 | 0.10 |
| 6 | b-Myrcene | 990 | 11.933 | 1.11 |
| 7 | a-Phellandrene | 1002 | 12.402 | 0.10 |
| 8 | a-Terpinene | 1017 | 12.855 | 1.49 |
| 9 | p-cymene a | 1024 | 13.163 | 12.92 |
| 10 | Limonene a | 1029 | 13.302 | 0.37 |
| 11 | γ-Terpinene a | 1059 | 14.400 | 7.12 |
| 12 | cis-Sabinene hydrate | 1070 | 14.701 | 0.72 |
| 13 | Linalool a | 1096 | 15.841 | 0.53 |
| 14 | Terpinen-4-ol | 1177 | 18.479 | 0.29 |
| 15 | Thymoquinone | 1252 | 20.816 | 0.19 |
| 16 | Thymol a | 1290 | 22.007 | 0.12 |
| 17 | Carvacrol a | 1299 | 22.382 | 69.89 |
| 18 | a-Cubenene | 1348 | 23.768 | 0.08 |
| 19 | a-Copaene | 1376 | 24.565 | 0.58 |
| 20 | b-Caryophyllene | 1419 | 25.883 | 1.38 |
| 21 | β-Bisabolene | 1505 | 27.302 | 0.14 |
| 22 | δ-Cadinene | 1523 | 29.561 | 0.29 |
| 23 | Thymohydro quinone | 1555 | 30.761 | 1.00 |
| 24 | Caryophyllene oxide | 1583 | 32.548 | 0.20 |
| Total (%) | 99.87 | |||
| Monoterpene hydrocarbons | 24.31 | |||
| Oxygenated monoterpenes | 72.87 | |||
| Sesquiterpene hydrocarbons | 2.69 |
(a) Identification by comparison of retention times and co-injection with authentic compound. (b) R.I. (Retention Indices) from experimental using a SBP-5 column using a homologous series of n-alkanes (C9–C25).
Table 3.
Impact of curative and residual vapour enrichment of chlorine or dittany oil on Botrytis cinerea spore germination and spore production in vitro on PDA medium. Treatments maintained throughout at 11 °C and 95% RH. In each row, mean (±SE) values (n = 9) of plates for the individual vapour enrichment, followed by the same letter, do not differ significantly at p = 0.05.
Table 3.
Impact of curative and residual vapour enrichment of chlorine or dittany oil on Botrytis cinerea spore germination and spore production in vitro on PDA medium. Treatments maintained throughout at 11 °C and 95% RH. In each row, mean (±SE) values (n = 9) of plates for the individual vapour enrichment, followed by the same letter, do not differ significantly at p = 0.05.
| Curative Vapour Enrichment * | |||
|---|---|---|---|
| In Vitro | Control | Chlorine | Dittany Oil |
| Spore production × 105 (mL−1) | 95.29 ± 6.60 a | 111.64 ± 10.89 a | 46.87 ± 14.35 b |
| Spore germination (%) | 68.58 ± 5.03 a | 60.88 ± 6.21 a | 73.11 ± 4.26 a |
| Vapour-Induced Residual Effect ** | |||
| Spore production × 105 (mL−1) | 141.36 ± 16.42 a | 63.26 ± 6.67 b | 25.63 ± 7.93 c |
| Spore germination (%) | 88.51 ± 2.96 a | 47.99 ± 4.63 b | 61.54 ± 7.69 b |
* Curative vapour enrichment: plates initially inoculated with B. cinerea and then subjected to vapours. ** Vapour-induced residual effect: plates pre-exposed to vapours, inoculated with fungi and then subjected to ambient air (control).
Table 4.
Impact of curative and residual vapour enrichment of chlorine or dittany oil on Botrytis cinerea spore germination and spore production on pepper fruit, stored at 11 °C and 95% RH. In each row, mean (±SE) values (n = 9) of fruits for the individual vapour enrichment followed by the same letter do not differ significantly at p = 0.05.
Table 4.
Impact of curative and residual vapour enrichment of chlorine or dittany oil on Botrytis cinerea spore germination and spore production on pepper fruit, stored at 11 °C and 95% RH. In each row, mean (±SE) values (n = 9) of fruits for the individual vapour enrichment followed by the same letter do not differ significantly at p = 0.05.
| Curative Vapour Enrichment * | |||
|---|---|---|---|
| In Vivo | Control | Chlorine | Dittany Oil |
| Spore production × 105 (mL−1) | 34.97 ± 6.25 a | 18.37 ± 3.09 b | 14.11 ± 2.05 b |
| Spore germination (%) | 99.60 ± 0.24 a | 97.20 ± 1.01 a | 91.25 ± 2.39 b |
| Vapour-Induced Residual Effect ** | |||
| Spore production × 105 (mL−1) | 2.85 ± 0.82 a | 1.52 ± 0.41 a | 1.04 ± 0.10 a |
| Spore germination (%) | 98.20 ± 1.56 a | 99.20 ± 0.20 a | 96.76 ± 1.65 a |
* Curative vapour enrichment: fruits inoculated with B. cinerea and then subjected to vapours. ** Vapour-induced residual effect: fruits pre-exposed to vapours, then inoculated with fungi, transferred and stored in ambient air.
Table 5.
Impact of curative vapour enrichment of pure (100%) chlorine or dittany oil on Botrytis cinerea lesion growth and spore germination on pepper fruit, stored at 11 °C and 95% RH. In each row, mean (±SE) values (n = 9) of fruits for the individual vapour enrichment followed by the same letter do not differ significantly at p = 0.05.
Table 5.
Impact of curative vapour enrichment of pure (100%) chlorine or dittany oil on Botrytis cinerea lesion growth and spore germination on pepper fruit, stored at 11 °C and 95% RH. In each row, mean (±SE) values (n = 9) of fruits for the individual vapour enrichment followed by the same letter do not differ significantly at p = 0.05.
| Curative Vapour Enrichment * | |||
|---|---|---|---|
| In Vivo | Control | Chlorine | Dittany Oil |
| Lesion growth (cm2) | 9.48 ± 0.96 a | 3.33 ± 0.68 b | 4.46 ± 0.56 b |
| Spore germination (%) | 96.54 ± 1.06 a | 2.63 ± 0.67 b | 0.25 ± 0.25 b |
* Curative vapour enrichment: fruits inoculated with B. cinerea and then subjected to pure (100%) vapours.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tzortzakis, N. Origanum dictamnus Essential Oil in Vapour or Aqueous Solution Application for Pepper Fruit Preservation against Botrytis cinerea. Agronomy 2024, 14, 257. https://doi.org/10.3390/agronomy14020257
AMA Style
Tzortzakis N. Origanum dictamnus Essential Oil in Vapour or Aqueous Solution Application for Pepper Fruit Preservation against Botrytis cinerea. Agronomy. 2024; 14(2):257. https://doi.org/10.3390/agronomy14020257
Chicago/Turabian StyleTzortzakis, Nikolaos. 2024. "Origanum dictamnus Essential Oil in Vapour or Aqueous Solution Application for Pepper Fruit Preservation against Botrytis cinerea" Agronomy 14, no. 2: 257. https://doi.org/10.3390/agronomy14020257
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.